
Responses of Ileal and Fecal Microbiota to Withdrawal of Pancreatic Enzyme Replacement Therapy in a Porcine Model of Exocrine Pancreatic Insufficiency

 , , ,
, , ,
Abstract
1. Introduction
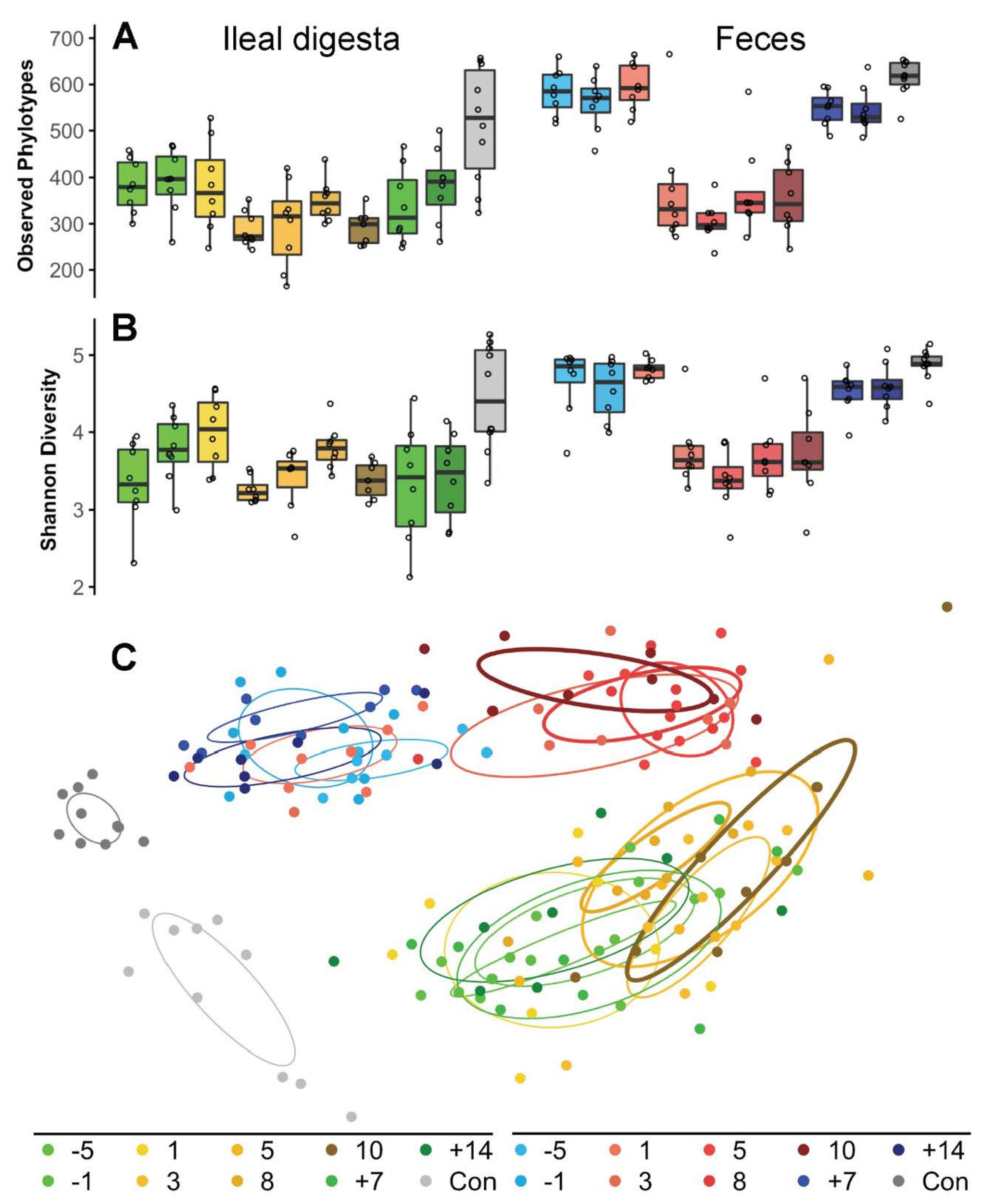
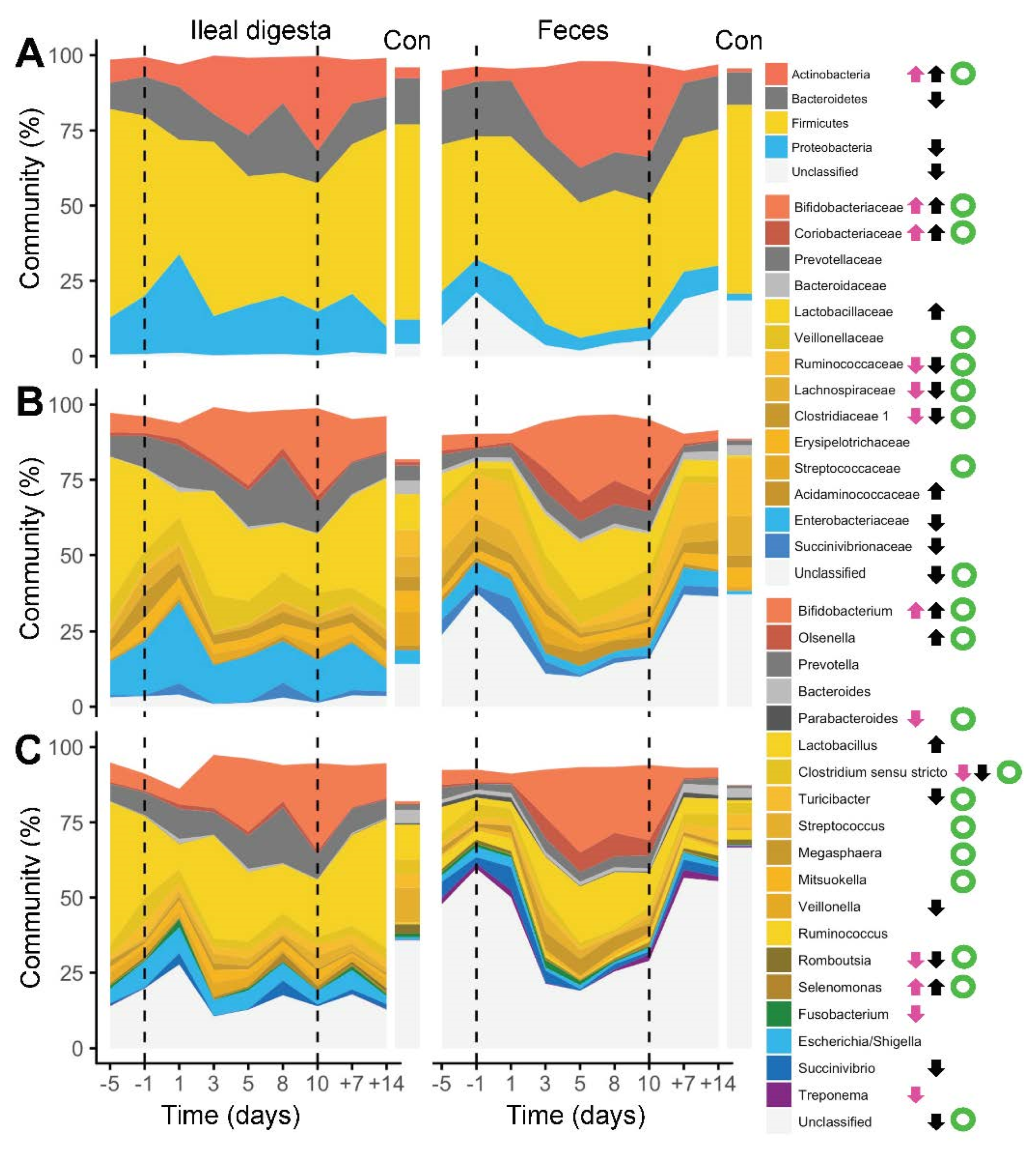
2. Results
3. Discussion
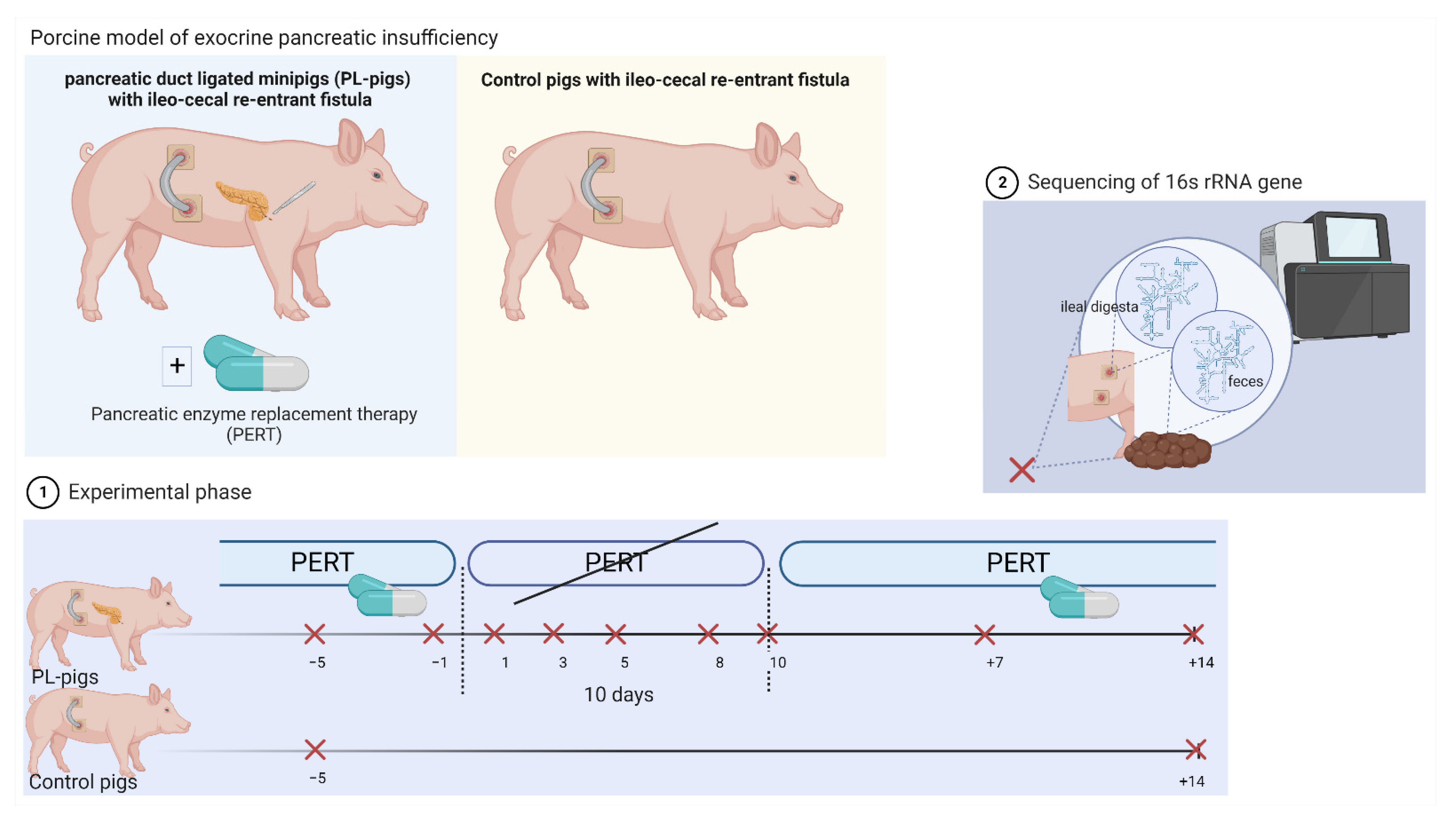
4. Materials and Methods
4.1. Animal Model
4.2. Microbiota Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Struyvenberg, M.R.; Martin, C.R.; Freedman, S.D. Practical guide to exocrine pancreatic insufficiency–breaking the myths. BMC Med. 2017, 15, 29. [Google Scholar] [CrossRef] [PubMed]
- Frost, F.; Weiss, F.U.; Sendler, M.; Kacprowski, T.; Rühlemann, M.; Bang, C.; Franke, A.; Völker, U.; Völzke, H.; Lamprecht, G. The Gut Microbiome in Patients With Chronic Pancreatitis Is Characterized by Significant Dysbiosis and Overgrowth by Opportunistic Pathogens. Clin. Transl. Gastroenterol. 2020, 11, e00232. [Google Scholar] [CrossRef] [PubMed]
- Pietzner, M.; Budde, K.; Rühlemann, M.; Völzke, H.; Homuth, G.; Weiss, F.U.; Lerch, M.M.; Frost, F. Exocrine Pancreatic Function Modulates Plasma Metabolites Through Changes in Gut Microbiota Composition. J. Clin. Endocrinol. Metab. 2021, 106, e2290–e2298. [Google Scholar] [CrossRef] [PubMed]
- Frost, F.; Kacprowski, T.; Rühlemann, M.; Bülow, R.; Kühn, J.-P.; Franke, A.; Heinsen, F.-A.; Pietzner, M.; Nauck, M.; Völker, U. Impaired exocrine pancreatic function associates with changes in intestinal microbiota composition and diversity. Gastroenterology 2019, 156, 1010–1015. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.-H.; Meng, Y.-T.; Xu, J.-J.; Fang, X.; Zhao, J.-L.; Zhou, W.; Zhao, J.; Han, J.-C.; Zhang, L.; Wang, K.-X. Altered diversity and composition of gut microbiota in Chinese patients with chronic pancreatitis. Pancreatology 2020, 20, 16–24. [Google Scholar] [CrossRef]
- Pan, L.-l.; Li, B.-b.; Pan, X.-h.; Sun, J. Gut microbiota in pancreatic diseases: Possible new therapeutic strategies. Acta Pharmacol. Sin. 2021, 42, 1027–1039. [Google Scholar] [CrossRef]
- Adolph, T.E.; Mayr, L.; Grabherr, F.; Schwärzler, J.; Tilg, H. Pancreas-microbiota cross talk in health and disease. Annu. Rev. Nutr. 2019, 39, 249–266. [Google Scholar] [CrossRef]
- El Kurdi, B.; Babar, S.; El Iskandarani, M.; Bataineh, A.; Lerch, M.M.; Young, M.; Singh, V.P. Factors that affect prevalence of small intestinal bacterial overgrowth in chronic pancreatitis: A systematic review, meta-analysis, and meta-regression. Clin. Transl. Gastroenterol. 2019, 10, e00072. [Google Scholar] [CrossRef]
- Bures, J.; Cyrany, J.; Kohoutova, D.; Förstl, M.; Rejchrt, S.; Kvetina, J.; Vorisek, V.; Kopacova, M. Small intestinal bacterial overgrowth syndrome. World J. Gastroenterol. 2010, 16, 2978. [Google Scholar] [CrossRef]
- Demeyer, S.; De Boeck, K.; Witters, P.; Cosaert, K. Beyond pancreatic insufficiency and liver disease in cystic fibrosis. Eur. J. Pediatr. 2016, 175, 881–894. [Google Scholar] [CrossRef]
- De la Iglesia-García, D.; Huang, W.; Szatmary, P.; Baston-Rey, I.; Gonzalez-Lopez, J.; Prada-Ramallal, G.; Mukherjee, R.; Nunes, Q.M.; Domínguez-Muñoz, J.E.; Sutton, R. Efficacy of pancreatic enzyme replacement therapy in chronic pancreatitis: Systematic review and meta-analysis. Gut 2017, 66, 1354–1355. [Google Scholar] [CrossRef] [PubMed]
- Diéguez-Castillo, C.; Jiménez-Luna, C.; Prados, J.; Martín-Ruiz, J.L.; Caba, O. State of the art in exocrine pancreatic insufficiency. Medicina 2020, 56, 523. [Google Scholar] [CrossRef] [PubMed]
- Ritz, S.; Hahn, D.; Wami, H.T.; Tegelkamp, K.; Dobrindt, U.; Schnekenburger, J. Gut microbiome as a response marker for pancreatic enzyme replacement therapy in a porcine model of exocrine pancreas insufficiency. Microb. Cell Fact 2020, 19, 523. [Google Scholar] [CrossRef] [PubMed]
- Luu, A.M.; Brock, A.; Ritz, S.; Junghänel, S.; Aldag, I.; Edskes, S.; Hartmann, M.; Hessler, M.; Praktiknjo, M.; Arnemann, P. Long term follow-up of a simplified and less burdened pancreatic duct ligation model of exocrine pancreatic insufficiency in Goettingen Minipigs. BMC Gastroenterol. 2020, 20, 221. [Google Scholar] [CrossRef]
- Gregory, P.C.; Hoffmann, K.; Kamphues, J.; Mößeler, A. The pancreatic duct ligated (mini) pig as a model for pancreatic exocrine insufficiency in man. Pancreas 2016, 45, 1213–1226. [Google Scholar] [CrossRef]
- Mößeler, A.; Kamphues, J. Black-Box Gastrointestinal Tract—Needs and Prospects of Gaining Insights of Fate of Fat, Protein, and Starch in Case of Exocrine Pancreatic Insufficiency by Using Fistulated Pigs. Nutrients 2017, 9, 150. [Google Scholar] [CrossRef]
- Kammlott, E.; Karthoff, J.; Stemme, K.; Gregory, P.; Kamphues, J. Experiments to optimize enzyme substitution therapy in pancreatic duct-ligated pigs. J. Anim. Physiol. Anim. Nutr. 2005, 89, 105–108. [Google Scholar] [CrossRef]
- Gesualdo, M.; Rizzi, F.; Bonetto, S.; Rizza, S.; Cravero, F.; Saracco, G.M.; De Angelis, C.G. Pancreatic Diseases and Microbiota: A Literature Review and Future Perspectives. J. Clin. Med. 2020, 9, 3535. [Google Scholar] [CrossRef]
- Kruszewska, D.; Ljungh, Å.; Hynes, S.O.; Pierzynowski, S.G. Effect of the antibacterial activity of pig pancreatic juice on human multiresistant bacteria. Pancreas 2004, 28, 191–199. [Google Scholar] [CrossRef]
- Mandischer, C. Untersuchungen am Pankreasgangligierten Schwein zur Verdaulichkeit (Praecaecal/In Toto) Eines Stärkereichen Mischfutters unter dem Einfluss zwei Verschiedener Oral Verabreichter Enzympräparationen. Ph.D. Thesis, University of Veterinary Medicine Hanover, Foundation, Hanover, Germany, 2002. [Google Scholar]
- Fuente-Dege, A.M. Untersuchungen an Pankreasgangligierten Schweinen zu Beziehungen Zwischen der Verdaulichkeit von Nährstoffen im Praecaecalen Bereich und der Verdaulichkeit über den Gesamten Verdauungstrakt. Ph.D. Thesis, University of Veterinary Medicine Hannover, Foundation, Hannover, Germany, 2003. [Google Scholar]
- Mößeler, A.K.; Gregory, P.C.; Kamphues, J. Impact of exocrine pancreas on digestion in the pig. Übers. Tierernährg. 2006, 34, 57. [Google Scholar]
- Uhr, G.T.; Dohnalová, L.; Thaiss, C.A. The dimension of time in host-microbiome interactions. mSystems 2019, 4, e00216–e00218. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Materna, A.C.; Friedman, J.; Campos-Baptista, M.I.; Blackburn, M.C.; Perrotta, A.; Erdman, S.E.; Alm, E.J. Host lifestyle affects human microbiota on daily timescales. Genome Biol. 2014, 15, e00216-18. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.J.; Vangay, P.; Al-Ghalith, G.A.; Hillmann, B.M.; Ward, T.L.; Shields-Cutler, R.R.; Kim, A.D.; Shmagel, A.K.; Syed, A.N.; Students, P.M.C. Daily sampling reveals personalized diet-microbiome associations in humans. Cell Host Microbe 2019, 25, 789–802.e5. [Google Scholar] [CrossRef] [PubMed]
- Trosvik, P.; De Muinck, E.J.; Stenseth, N.C. Biotic interactions and temporal dynamics of the human gastrointestinal microbiota. ISME J. 2015, 9, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Vital, M.; Howe, A.; Bergeron, N.; Krauss, R.M.; Jansson, J.K.; Tiedje, J.M. Metagenomic insights into the degradation of resistant starch by human gut microbiota. Appl. Env. Microbiol. 2018, 84, e01562-18. [Google Scholar] [CrossRef]
- Dethlefsen, L.; Relman, D.A. Incomplete recovery and individualized responses of the human distal gut microbiota to repeated antibiotic perturbation. Proc. Natl. Acad. Sci. USA 2011, 108, 4554–4561. [Google Scholar] [CrossRef]
- Shade, A.; Peter, H.; Allison, S.D.; Baho, D.; Berga, M.; Bürgmann, H.; Huber, D.H.; Langenheder, S.; Lennon, J.T.; Martiny, J.B. Fundamentals of microbial community resistance and resilience. Front. Microbiol. 2012, 3, 417. [Google Scholar] [CrossRef]
- Vernocchi, P.; Del Chierico, F.; Russo, A.; Majo, F.; Rossitto, M.; Valerio, M.; Casadei, L.; La Storia, A.; De Filippis, F.; Rizzo, C. Gut microbiota signatures in cystic fibrosis: Loss of host CFTR function drives the microbiota enterophenotype. PLoS ONE 2018, 13, e0208171. [Google Scholar] [CrossRef]
- Burke, D.; Fouhy, F.; Harrison, M.; Rea, M.C.; Cotter, P.D.; O’Sullivan, O.; Stanton, C.; Hill, C.; Shanahan, F.; Plant, B.J. The altered gut microbiota in adults with cystic fibrosis. BMC Microbiol. 2017, 17, 58. [Google Scholar] [CrossRef]
- Rose, E.C.; Blikslager, A.T.; Ziegler, A.L. Porcine Models of the Intestinal Microbiota: The Translational Key to Understanding How Gut Commensals Contribute to Gastrointestinal Disease. Front. Vet. Sci. 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Gavini, F.; Delcenserie, V.; Kopeinig, K.; Pollinger, S.; Beerens, H.; Bonaparte, C.; Upmann, M. Bifidobacterium species isolated from animal feces and from beef and pork meat. J. Food Prot. 2006, 69, 871–877. [Google Scholar] [CrossRef] [PubMed]
- O’callaghan, A.; Van Sinderen, D. Bifidobacteria and their role as members of the human gut microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef]
- Wang, Z.; Bai, Y.; Pi, Y.; Gerrits, W.J.; de Vries, S.; Shang, L.; Tao, S.; Zhang, S.; Han, D.; Zhu, Z. Xylan alleviates dietary fiber deprivation-induced dysbiosis by selectively promoting Bifidobacterium pseudocatenulatum in pigs. Microbiome 2021, 9, 227. [Google Scholar] [CrossRef] [PubMed]
- Isaiah, A.; Parambeth, J.C.; Steiner, J.M.; Lidbury, J.A.; Suchodolski, J.S. The fecal microbiome of dogs with exocrine pancreatic insufficiency. Anaerobe 2017, 45, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Scotta, M.; Marzani, M.; Maggiore, G.; De Giacomo, C.; D’Eril, G.M.; Moratti, R. Fecal chymotrypsin: A new diagnostic test for exocrine pancreatic insufficiency in children with cystic fibrosis. Clin. Biochem. 1985, 18, 233–234. [Google Scholar] [CrossRef]
- Lankisch, P.; Schmidt, I.; König, H.; Lehnick, D.; Knollmann, R.; Löhr, M.; Liebe, S. Faecal elastase 1: Not helpful in diagnosing chronic pancreatitis associated with mild to moderate exocrine pancreatic insufficiency. Gut 1998, 42, 551–554. [Google Scholar] [CrossRef]
- Million, M.; Tidjani Alou, M.; Khelaifia, S.; Bachar, D.; Lagier, J.-C.; Dione, N.; Brah, S.; Hugon, P.; Lombard, V.; Armougom, F. Increased gut redox and depletion of anaerobic and methanogenic prokaryotes in severe acute malnutrition. Sci. Rep. 2016, 6, 26051. [Google Scholar] [CrossRef]
- Wang, W.; Chen, L.; Zhou, R.; Wang, X.; Song, L.; Huang, S.; Wang, G.; Xia, B. Increased proportions of Bifidobacterium and the Lactobacillus group and loss of butyrate-producing bacteria in inflammatory bowel disease. J. Clin. Microbiol. 2014, 52, 398–406. [Google Scholar] [CrossRef]
- Joly, F.; Mayeur, C.; Bruneau, A.; Noordine, M.-L.; Meylheuc, T.; Langella, P.; Messing, B.; Duée, P.-H.; Cherbuy, C.; Thomas, M. Drastic changes in fecal and mucosa-associated microbiota in adult patients with short bowel syndrome. Biochimie 2010, 92, 753–761. [Google Scholar] [CrossRef]
- Bunte, S.; Grone, R.; Keller, B.; Keller, C.; Galvez, E.; Strowig, T.; Kamphues, J.; Hankel, J. Intestinal microbiota of fattening pigs offered non-fermented and fermented liquid feed with and without the supplementation of non-fermented coarse cereals. Microorganisms 2020, 8, 638. [Google Scholar] [CrossRef] [PubMed]
- Mößeler, A.; Vagt, S.; Beyerbach, M.; Kamphues, J. Starch origin and thermal processing affect starch digestion in a minipig model of pancreatic exocrine insufficiency. Gastroenterol. Res. Pract. 2015, 2015, 872872. [Google Scholar] [CrossRef] [PubMed]
- Vagt, S. In Vivo and in Vitro Investigations on Prececal Starch Digestion (Related to Source of Starch and Thermal Treatment) in Pancreatic Duct Ligated Minipigs—Used as a Model for Human Exocrine Pancreatic Insufficiency. Ph.D. Thesis, University of Veterinary Medicine, Foundation Hannover, Hanover, Germany, 2014. [Google Scholar]
- Bohm, M.; Siwiec, R.M.; Wo, J.M. Diagnosis and management of small intestinal bacterial overgrowth. Nutr. Clin. Pract. 2013, 28, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Lugli, G.A.; Duranti, S.; Turroni, F.; Bottacini, F.; Mangifesta, M.; Sanchez, B.; Viappiani, A.; Mancabelli, L.; Taminiau, B. Genomic encyclopedia of type strains of the genus Bifidobacterium. Appl. Environ. Microbiol. 2014, 80, 6290–6302. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Lugli, G.A.; Duranti, S.; Turroni, F.; Mancabelli, L.; Ferrario, C.; Mangifesta, M.; Hevia, A.; Viappiani, A.; Scholz, M. Bifidobacteria exhibit social behavior through carbohydrate resource sharing in the gut. Sci. Rep. 2015, 5, 15782. [Google Scholar] [CrossRef]
- Bouhnik, Y.; Raskine, L.; Simoneau, G.; Vicaut, E.; Neut, C.; Flourié, B.; Brouns, F.; Bornet, F.R. The capacity of nondigestible carbohydrates to stimulate fecal bifidobacteria in healthy humans: A double-blind, randomized, placebo-controlled, parallel-group, dose-response relation study. Am. J. Clin. Nutr. 2004, 80, 1658–1664. [Google Scholar] [CrossRef]
- Metzler-Zebeli, B.; Canibe, N.; Montagne, L.; Freire, J.; Bosi, P.; Prates, J.A.; Tanghe, S.; Trevisi, P. Resistant starch reduces large intestinal pH and promotes fecal lactobacilli and bifidobacteria in pigs. Animal 2019, 13, 64–73. [Google Scholar] [CrossRef]
- Butteiger, D.N.; Hibberd, A.A.; McGraw, N.J.; Napawan, N.; Hall-Porter, J.M.; Krul, E.S. Soy protein compared with milk protein in a western diet increases gut microbial diversity and reduces serum lipids in golden syrian hamsters. J. Nutr. 2015, 146, 697–705. [Google Scholar] [CrossRef]
- Dominika, Ś.; Arjan, N.; Karyn, R.P.; Henryk, K. The study on the impact of glycated pea proteins on human intestinal bacteria. Int. J. Food Microbiol. 2011, 145, 267–272. [Google Scholar] [CrossRef]
- Picard, C.; Fioramonti, J.; Francois, A.; Robinson, T.; Neant, F.; Matuchansky, C. bifidobacteria as probiotic agents–physiological effects and clinical benefits. Aliment. Pharmacol. Ther. 2005, 22, 495–512. [Google Scholar] [CrossRef]
- Tabeling, R.; Gregory, P.; Kamphues, J. Studies on nutrient digestibilities (pre-caecal and total) in pancreatic duct-ligated pigs and the effects of enzyme substitution. J. Anim. Physiol. Anim. Nutr. 1999, 82, 251–263. [Google Scholar] [CrossRef]
- Easter, R.A.; Tanksley Jr, T.D. A technique for re-entrant ileocecal cannulation of swine. J. Anim. Sci. 1973, 36, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Hazem, A.; Drochner, W. Extracorporeal ileocecal bypass avoiding the ileocecal value in swine. Z. Versuchstierkd. 1976, 18, 303–306. [Google Scholar]
- Rath, S.; Heidrich, B.; Pieper, D.H.; Vital, M. Uncovering the trimethylamine-producing bacteria of the human gut microbiota. Microbiome 2017, 5, 54. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Schütte, K.; Koch, N.; Vilchez-Vargas, R.; Wos-Oxley, M.L.; Oxley, A.P.; Vital, M.; Malfertheiner, P.; Pieper, D.H. The active bacterial assemblages of the upper GI tract in individuals with and without Helicobacter infection. Gut 2018, 67, 216–225. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Env. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Evol. Biol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| PL-Pigs (n = 8) | Control Pigs (n = 5) | |||
|---|---|---|---|---|
| Range | Mean ± SD | Range | Mean ± SD | |
| Age (years) | 1–5 | 3.38 ± 1.30 | 2–5 | 3.60 ± 1.34 |
| Body weight (kg) | 21.6–29.3 | 26.4 ± 2.89 | 24.8–32.6 | 27.9 ± 4.06 |
| Time after surgery (years) | 1–4 | 2.63 ± 1.06 | 1–4 | 2.20 ± 1.30 |
| Chymotrypsin activity in feces (U/g) | 0.106–0.424 | 0.225 ± 0.155 | 2.4–26.9 | 10.5 ± 9.70 |
| ID | Annotation | M_Ildig (%) | M_fec (%) | |||
|---|---|---|---|---|---|---|
| 2 | B. thermophilum * | 7.36 | 7.39 | ↑ | ↑ | ◯ |
| 9 | B. pseudolongum * | 1.95 | 1.13 | ↑ | ↑ | ◯ |
| 10 | B. thermophilum * | 1.63 | 1.65 | ↑ | ↑ | ◯ |
| 18 | B. boum * | 1.04 | 0.52 | ↑ | ↑ | ◯ |
| 28 | Clostridium sensu stricto | 0.62 | 0.43 | ↓ | ↓ | ◯ |
| 49 | Holdemanella | 0.43 | 0.27 | ↓ | ↓ | ◯ |
| 99 | Alloprevotella | 0.32 | 0.01 | ↓ | ↓ | ◯ |
| 40 | Clostridium sensu stricto | 0.32 | 0.40 | ↓ | ↓ | ◯ |
| 67 | Selenomonas | 0.30 | 0.20 | ↑ | ↑ | ◯ |
| 70 | Mitsuokella | 0.28 | 0.22 | ↑ | ↑ | ◯ |
| 87 | Clostridium sensu stricto | 0.21 | 0.12 | ↓ | ↓ | ◯ |
| 61 | Prevotellaceae | 0.20 | 0.37 | ↑ | ↑ | ◯ |
| 95 | Romboutsia | 0.16 | 0.18 | ↓ | ↓ | ◯ |
| 132 | Prevotellaceae | 0.09 | 0.13 | ↑ | ↑ | ◯ |
| 108 | Bacteroidales | 0.05 | 0.22 | ↓ | ↓ | ◯ |
| 110 | Solobacterium | 0.05 | 0.25 | ↑ | ↑ | ◯ |
| 109 | Bacteroidales | 0.04 | 0.24 | ↓ | ↓ | ◯ |
| 123 | Parabacteroides | 0.03 | 0.22 | ↓ | ↓ | |
| 25 | Prevotella | 1.20 | 0.10 | ↑ | ||
| 37 | Lactobacillus | 0.78 | 0.04 | ↓ | ||
| 42 | Prevotella | 0.72 | 0.04 | ↑ | ||
| 1 | Lactobacillus | 16.41 | 7.25 | ↑ | ◯ | |
| 5 | Succinivibrio | 1.37 | 2.90 | ↓ | ||
| 4 | Turicibacter | 2.99 | 2.31 | ↓ | ◯ | |
| 14 | Bacteria | 0.14 | 2.22 | ↓ | ||
| 17 | Bacteria | 0.01 | 1.82 | ↓ | ||
| 16 | Olsenella | 0.46 | 1.55 | ↑ | ◯ | |
| 20 | Bacteroidetes | 0.06 | 1.50 | ↓ | ||
| 3 | Escherichia/Shigella | 4.22 | 1.46 | ↓ | ||
| 26 | Lachnospiraceae | 0.00 | 1.35 | ↓ | ||
| 19 | Acidaminococcaceae | 0.33 | 1.06 | ↑ | ◯ | |
| 8 | Enterobacteriaceae | 2.63 | 0.81 | ↓ | ||
| 51 | Bacteria | 0.00 | 0.72 | ↓ | ||
| 48 | Bacteria | 0.07 | 0.70 | ↑ | ◯ | |
| 39 | Clostridiales | 0.18 | 0.63 | ↑ | ◯ | |
| 44 | Bacteria | 0.05 | 0.55 | ↑ | ||
| 31 | Lactobacillus | 0.67 | 0.31 | ↑ | ||
| 34 | Lactobacillus | 0.60 | 0.30 | ↑ | ||
| 21 | Lactobacillus | 1.13 | 0.24 | ↑ | ||
| 36 | Lactobacillus | 0.68 | 0.18 | ↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hankel, J.; Mößeler, A.; Hartung, C.B.; Rath, S.; Schulten, L.; Visscher, C.; Kamphues, J.; Vital, M. Responses of Ileal and Fecal Microbiota to Withdrawal of Pancreatic Enzyme Replacement Therapy in a Porcine Model of Exocrine Pancreatic Insufficiency. Int. J. Mol. Sci. 2022, 23, 11700. https://doi.org/10.3390/ijms231911700
Hankel J, Mößeler A, Hartung CB, Rath S, Schulten L, Visscher C, Kamphues J, Vital M. Responses of Ileal and Fecal Microbiota to Withdrawal of Pancreatic Enzyme Replacement Therapy in a Porcine Model of Exocrine Pancreatic Insufficiency. International Journal of Molecular Sciences. 2022; 23(19):11700. https://doi.org/10.3390/ijms231911700
Chicago/Turabian StyleHankel, Julia, Anne Mößeler, Clara Berenike Hartung, Silke Rath, Lisa Schulten, Christian Visscher, Josef Kamphues, and Marius Vital. 2022. "Responses of Ileal and Fecal Microbiota to Withdrawal of Pancreatic Enzyme Replacement Therapy in a Porcine Model of Exocrine Pancreatic Insufficiency" International Journal of Molecular Sciences 23, no. 19: 11700. https://doi.org/10.3390/ijms231911700
APA StyleHankel, J., Mößeler, A., Hartung, C. B., Rath, S., Schulten, L., Visscher, C., Kamphues, J., & Vital, M. (2022). Responses of Ileal and Fecal Microbiota to Withdrawal of Pancreatic Enzyme Replacement Therapy in a Porcine Model of Exocrine Pancreatic Insufficiency. International Journal of Molecular Sciences, 23(19), 11700. https://doi.org/10.3390/ijms231911700