
Embryogenic Stem Cell Identity after Protoplast Isolation from Daucus carota and Recovery of Regeneration Ability through Protoplast Culture

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
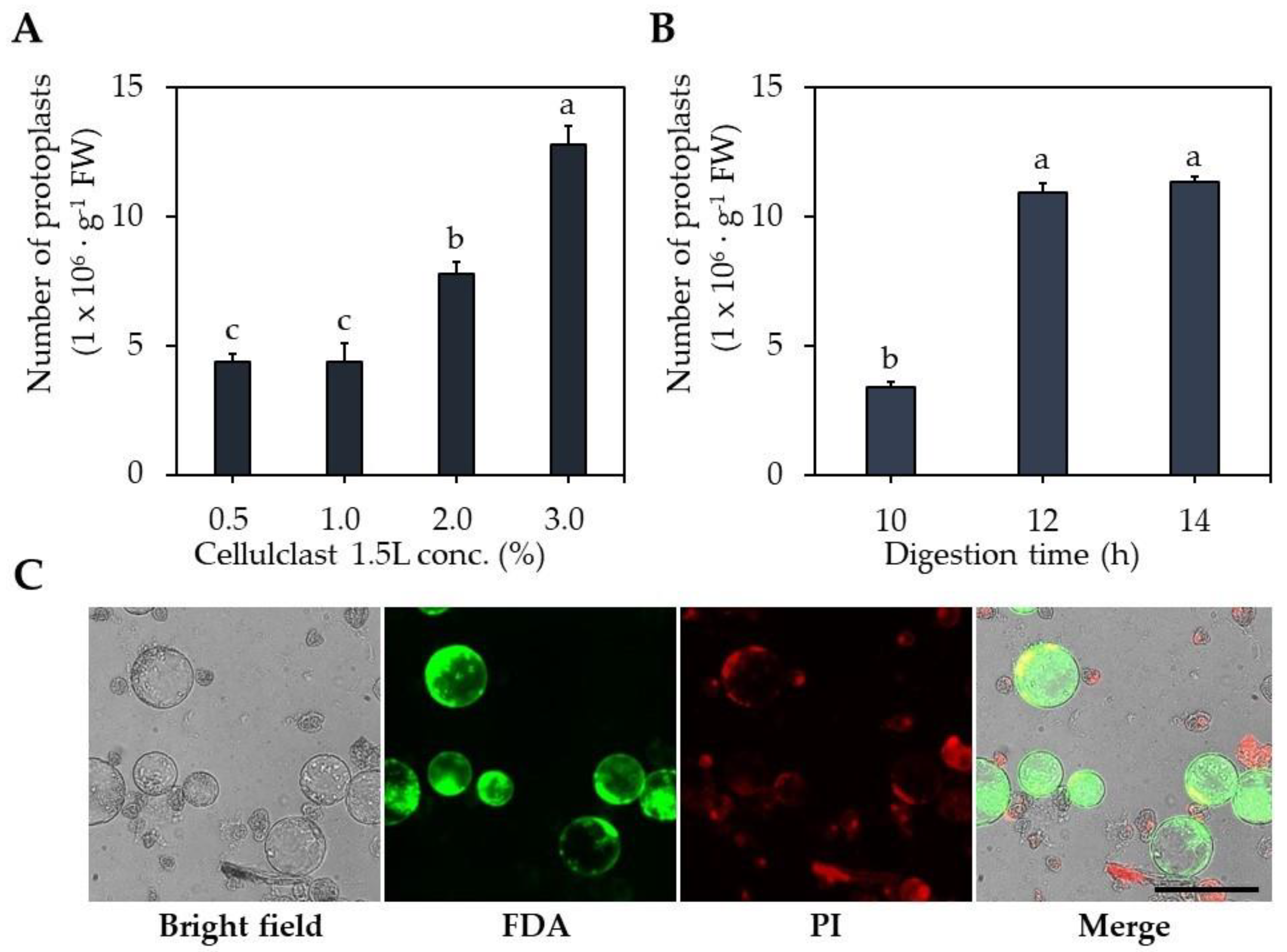
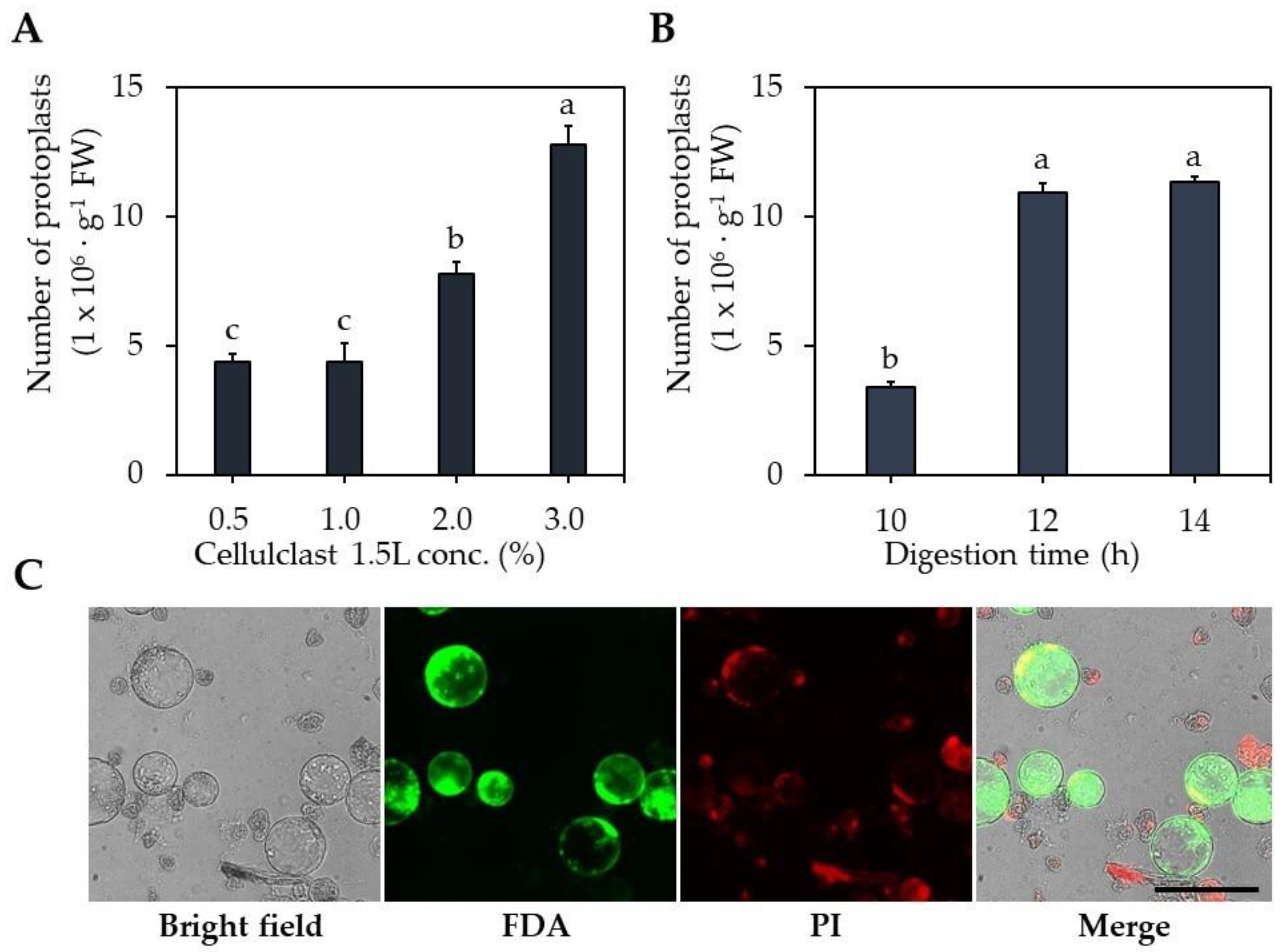
2.1. Optimization of Protoplast Isolation
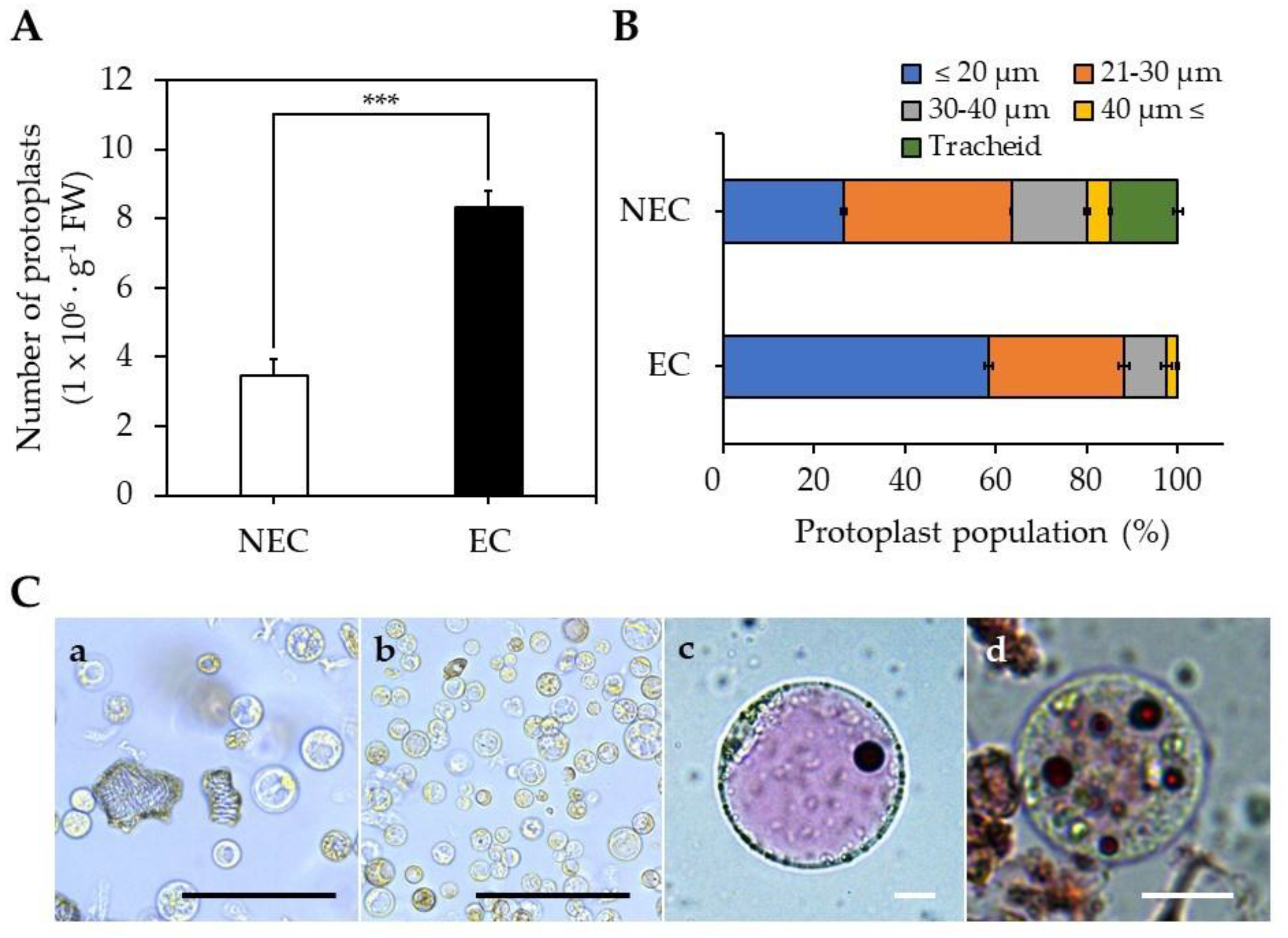
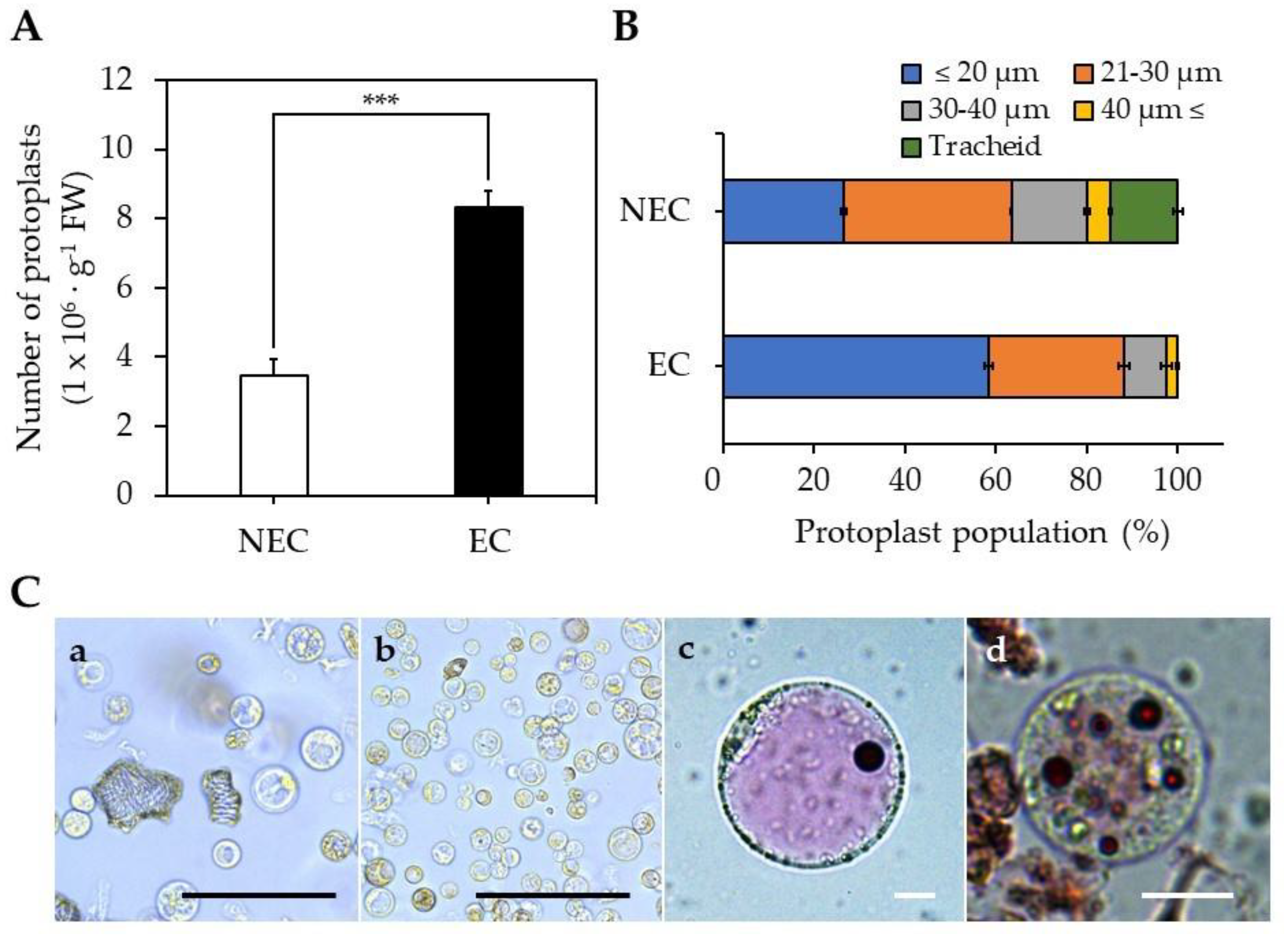
2.2. Isolation of Protoplasts from EC and NEC
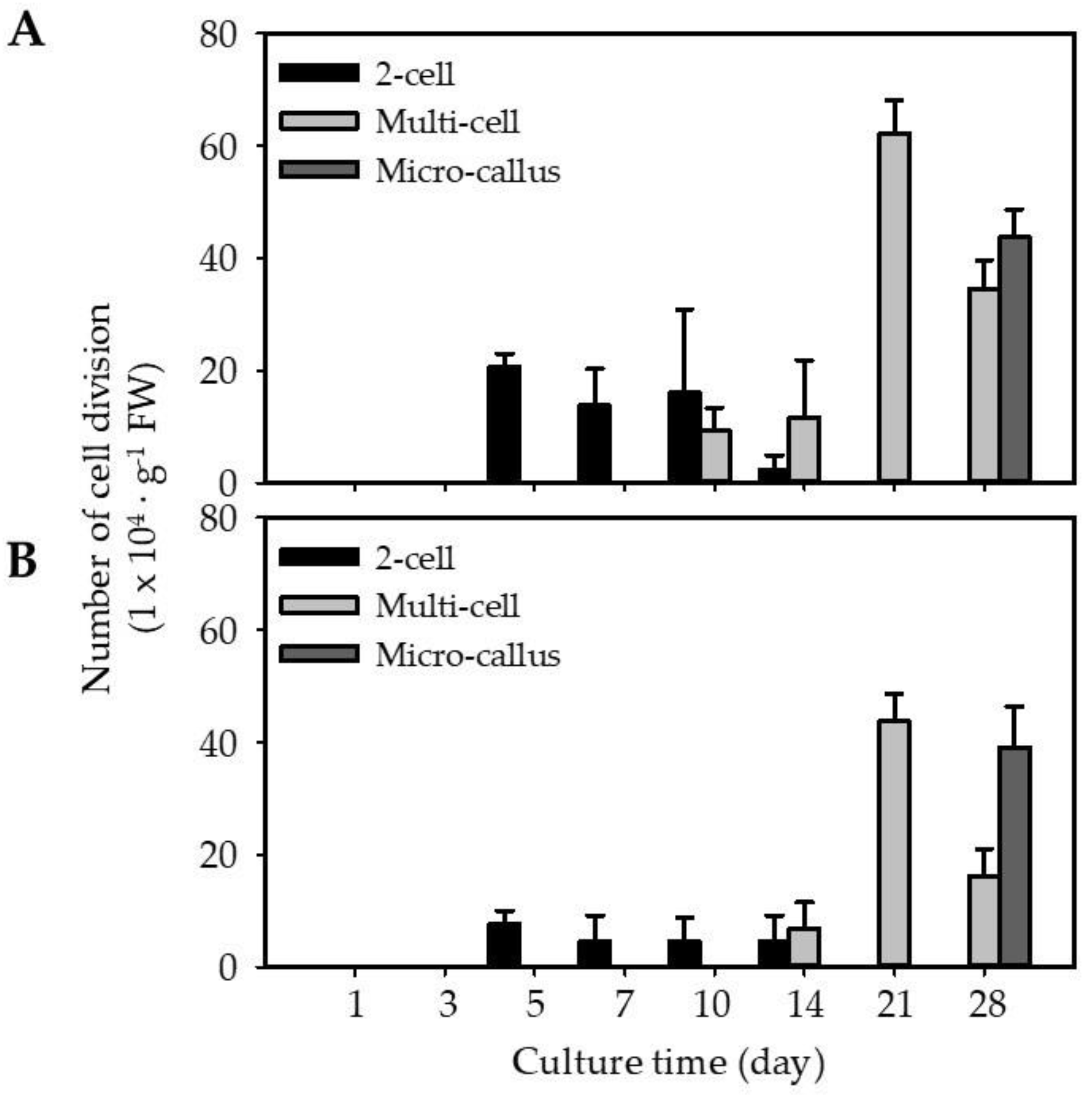
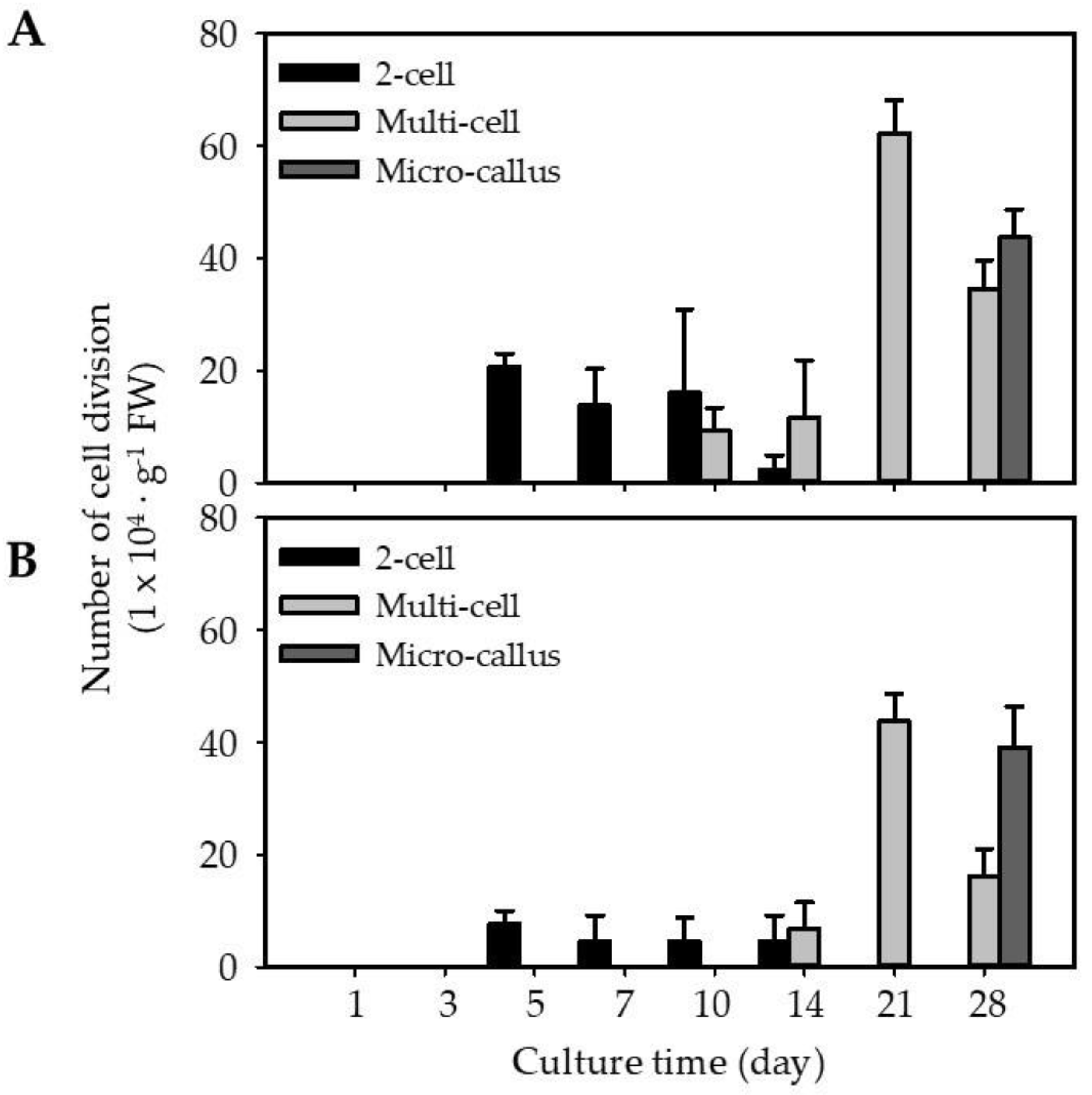
2.3. Regeneration of EC and NEC-Derived Protoplasts
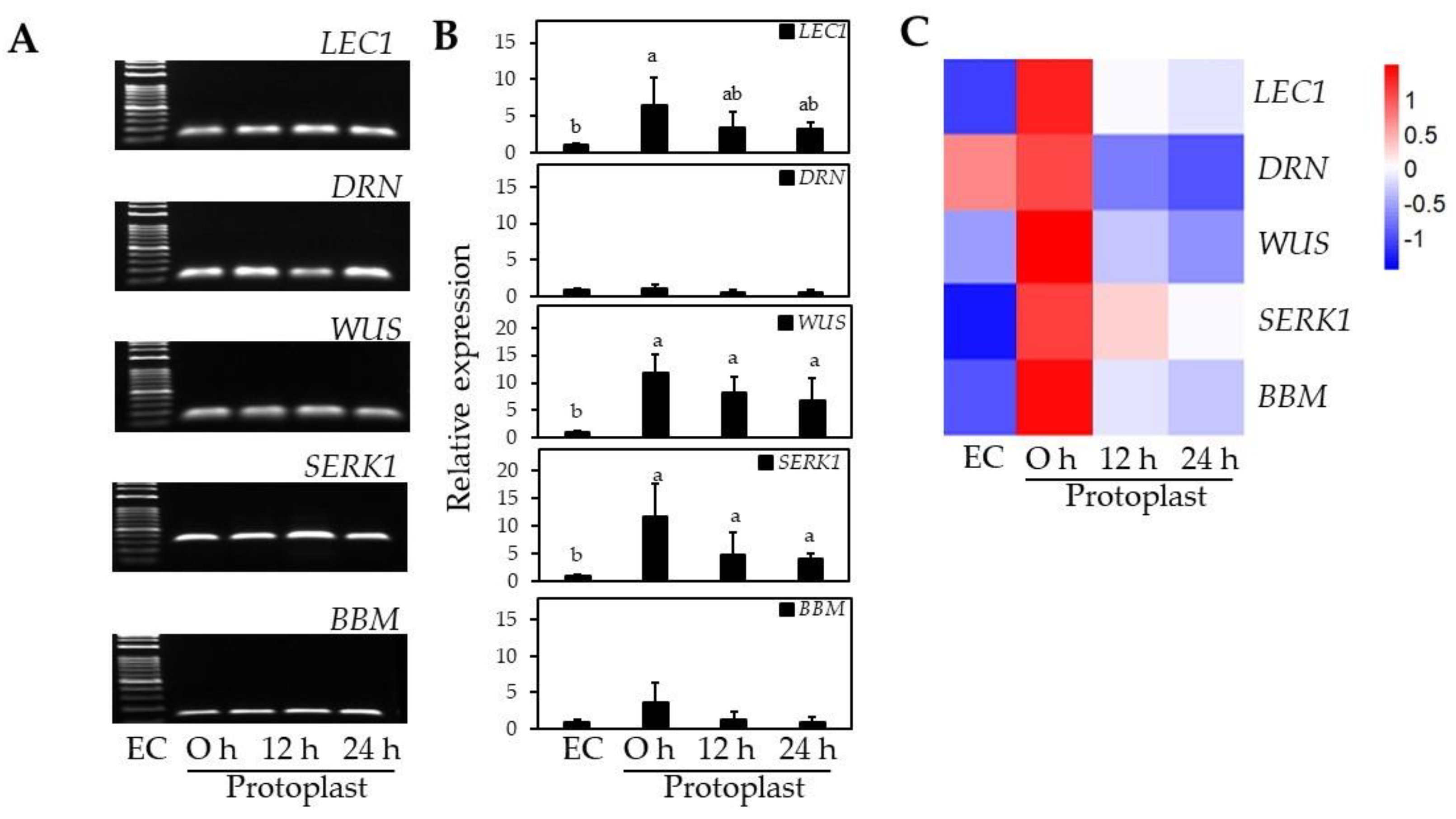
2.4. Regeneration Capacity of EC-pt-TAL and EC-Specific Gene Expression
3. Materials and Methods
3.1. Plant Materials
3.2. Optimization of Protoplast Isolation
3.3. EC- and NEC-Derived Protoplast Isolation and Culture
3.3.1. Protoplast Isolation
3.3.2. Protoplast Culture Using Thin Alginate Layer (TAL) System
3.3.3. Plant Regeneration from EC-TAL, EC-pt-TAL, and NEC-pt-TAL
3.3.4. Microscopy and Staining
3.3.5. EC-Specific Gene Expression
3.3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, M.B.; Bhalla, P.L. Plant stem cells carve their own niche. Trends Plant Sci. 2006, 11, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Verdeil, J.; Alemanno, L.; Niemenak, N.; Tranbarger, T. Pluripotent versus totipotent plant stem cells: Dependence versus autonomy? Trends Plant Sci. 2007, 12, 245–252. [Google Scholar] [CrossRef]
- Passamani, L.Z.; Reis, R.S.; Vale, E.M.; Sousa, K.R.; Aragão, V.P.M.; Santa-Catarina, C.; Silveira, V. Long-term culture with 2,4-dichlorophenoxyacetic acid affects embryogenic competence in sugarcane callus via changes in starch, polyamine and protein profiles. Plant Cell Tissue Organ Cult. 2020, 140, 415–429. [Google Scholar] [CrossRef]
- Weckx, S.; Inzé, D.; Maene, L. Tissue culture of oil palm: Finding the balance between mass propagation and somaclonal variation. Front. Plant Sci. 2019, 10, 722. [Google Scholar] [CrossRef]
- Tacán, M.; Tapia, C.; Pérez, C.; Zambrano, E.; Mendoza, A.; Monteros-Altamirano, Á.; Sørensen, M. Cryopreservation of Arachis Hypogaea, L. varieties, from the INIAP-ecuador germplasm bank. Preprints 2021, 2021120393. [Google Scholar] [CrossRef]
- Godel-Jędrychowska, K.; Maćkowska, K.; Kurczyńska, E.; Grzebelus, E. Composition of the reconstituted cell wall in protoplast-derived cells of daucus is affected by phytosulfokine (PSK). Int. J. Mol. Sci. 2019, 20, 5490. [Google Scholar] [CrossRef]
- Adedeji, O.S.; Naing, A.H.; Kil Kim, C. Protoplast isolation and shoot regeneration from protoplast-derived calli of Chrysanthemum cv. White ND. Plant Cell Tissue Organ Cult. 2020, 141, 571–581. [Google Scholar] [CrossRef]
- Rahmani, M.-S.; Pijut, P.M.; Shabanian, N. Protoplast isolation and genetically true-to-type plant regeneration from leaf- and callus-derived protoplasts of Albizia julibrissin. Plant Cell Tissue Organ Cult. 2016, 127, 475–488. [Google Scholar] [CrossRef]
- Kim, S.W.; Oh, S.C.; In, D.S.; Liu, J.R. Plant regeneration of rose (Rosa hybridia) from embryogenic cell-derived protoplasts. Plant Cell Tissue Organ Cult. 2003, 73, 15–19. [Google Scholar] [CrossRef]
- Bertini, E.; Tornielli, G.B.; Pezzotti, M.; Zenoni, S. Regeneration of plants from embryogenic callus-derived protoplasts of Garganega and Sangiovese grapevine (Vitis vinifera L.) cultivars. Plant Cell Tissue Organ Cult. 2019, 138, 239–246. [Google Scholar] [CrossRef]
- Karamian, R.; Ebrahimzadeh, H. Plantlet regeneration from protoplast-derived embryogenic calli of Crocus cancellatus. Plant Cell Tissue Organ Cult. 2001, 65, 115–121. [Google Scholar] [CrossRef]
- Tang, K.; Sun, X.; An, D.; Power, J.B.; Cocking, E.C.; Davey, M.R. A simple and rapid procedure to establish embryogenic cell suspensions as a source of protoplasts for efficient plant regeneration from two Chinese commercial rice cultivars. Plant Cell Tissue Organ Cult. 2001, 66, 149–153. [Google Scholar] [CrossRef]
- Du, L.; Bao, M. Plant regeneration from protoplasts isolated from embryogenic suspension cultured cells of Cinnamomum camphora L. Plant Cell Reports. 2005, 24, 462–467. [Google Scholar] [CrossRef]
- Guo, H.; Guo, H.; Zhang, L.; Tang, Z.; Yu, X.; Wu, J.; Zeng, F. Metabolome and Transcriptome Association Analysis Reveals Dynamic Regulation of Purine Metabolism and Flavonoid Synthesis in Transdifferentiation during Somatic Embryogenesis in Cotton. Int. J. Mol. Sci. 2019, 20, 2070. [Google Scholar] [CrossRef]
- Kumar, S.; Ruggles, A.; Logan, S.; Mazarakis, A.; Tyson, T.; Bates, M.; Grosse, C.; Reed, D.; Li, Z.; Grimwood, J.; et al. Comparative Transcriptomics of Non-Embryogenic and Embryogenic Callus in Semi-Recalcitrant and Non-Recalcitrant Upland Cotton Lines. Plants 2021, 10, 1775. [Google Scholar] [CrossRef]
- Liu, B.; Shan, X.; Wu, Y.; Su, S.; Li, S.; Liu, H.; Han, J.; Yuan, Y. iTRAQ-Based Quantitative Proteomic Analysis of Embryogenic and Non-embryogenic Calli Derived from a Maize (Zea mays L.) Inbred Line Y423. Int. J. Mol. Sci. 2018, 19, 4004. [Google Scholar] [CrossRef]
- Shim, S.; Kim, H.K.; Bae, S.H.; Lee, H.; Lee, H.J.; Jung, Y.J.; Seo, P.J. Transcriptome comparison between pluripotent and non-pluripotent calli derived from mature rice seeds. Sci. Rep. 2020, 10, 21257. [Google Scholar] [CrossRef]
- Wen, L.; Li, W.; Parris, S.; West, M.; Lawson, J.; Smathers, M.; Li, Z.; Jones, D.; Jin, S.; Saski, C.A. Transcriptomic profiles of non-embryogenic and embryogenic callus cells in a highly regenerative upland cotton line (Gossypium hirsutum L.). BMC Dev. Biol. 2020, 20, 25. [Google Scholar] [CrossRef]
- Jha, P.; Ochatt, S.J.; Kumar, V. WUSCHEL: A master regulator in plant growth signaling. Plant Cell Rep. 2020, 39, 431–444. [Google Scholar] [CrossRef]
- Ma, W.; Yi, F.; Xiao, Y.; Yang, G.; Chen, F.; Wang, J. Isolation of leaf mesophyll protoplasts optimized by orthogonal design for transient gene expression in Catalpa bungei. Sci. Hortic. 2020, 274, 109684. [Google Scholar] [CrossRef]
- Bai, L.; Cheng, Y.; She, J.; He, Z.; Liu, H.; Zhang, G.; Cao, R.; Chen, Y. Development of an efficient protoplast isolation and transfection system for castor bean (Ricinus communis L.). Plant Cell Tissue Organ Cult. 2020, 143, 457–464. [Google Scholar] [CrossRef]
- Cosgrove, D.C.; Jarvis, M.C. Comparative structure and biomechanics of plant primary and secondary cell walls. Front. Plant Sci. 2012, 3, 204. [Google Scholar] [CrossRef] [PubMed]
- Krumm, C.; Pfaendtner, J.; Dauenhauer, P.J. Millisecond pulsed films unify the mechanisms of cellulose fragmentation. Chem. Mater. 2016, 28, 3108–3114. [Google Scholar] [CrossRef]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef]
- Mota-NarvÃ, L.A.; del Rosario Ayora-Talavera, T.; Santana-Buzzy, N.; López-Puc, G. Isolation and regeneration of protoplasts from leaf explants of Rhyncholaelia digbyana. Afr. J. Biotechnol. 2018, 17, 1082–1089. [Google Scholar]
- Ren, R.; Gao, J.; Lu, C.; Wei, Y.; Jin, J.; Wong, S.-M.; Zhu, G.; Yang, F. Highly efficient protoplast isolation and transient expression system for functional characterization of flowering related genes in Cymbidium orchids. Int. J. Mol. Sci. 2020, 21, 2264. [Google Scholar] [CrossRef]
- Moghaddam, P.R.; Wilman, D. Cell wall thickness and cell dimensions in plant parts of eight forage species. J. Agric. Sci. 1998, 131, 59–67. [Google Scholar] [CrossRef]
- Simpson, K.; Fuentes, P.; Quiroz-Iturra, L.F.; Flores-Ortiz, C.; Contreras, R.; Handford, M.; Stange, C. Unraveling the induction of phytoene synthase 2 expression by salt stress and abscisic acid in Daucus carota. J. Exp. Bot. 2018, 69, 4113–4126. [Google Scholar] [CrossRef]
- Nguyen, T.K.; Ha, S.T.T.; Lim, J. Optimization of protoplast isolation methods from leaves in Chrysanthemum morifolium ‘Jinba’. Ann. Biol. Sci. 2019, 7, 16–19. [Google Scholar]
- Mallick, P.; Ghosh, S.; Chattaraj, S.; Sikdar, S.R. Isolation of mesophyll protoplast from Indian mulberry (Morus alba L) cv. S 1635. J. Environ. Sociobiol. 2016, 13, 217–222. [Google Scholar]
- Dovzhenko, A.; Koop, H.-U. Sugarbeet (Beta vulgaris L.): Shoot regeneration from callus and callus protoplasts. Planta 2003, 217, 374–381. [Google Scholar] [CrossRef]
- Klimek-Chodacka, M.; Kadluczka, D.; Lukasiewicz, A.; Malec-Pala, A.; Baranski, R.; Grzebelus, E. Effective callus induction and plant regeneration in callus and protoplast cultures of Nigella damascena L. Plant Cell Tissue Organ Cult. 2020, 143, 693–707. [Google Scholar] [CrossRef]
- Kanwar, K.; Bhardwaj, A.; Deepika, R. Efficient regeneration of plantlets from callus and mesophyll derived protoplasts of Robinia pseudoacacia L. Plant Cell Tissue Organ Cult. 2009, 96, 95–103. [Google Scholar] [CrossRef]
- Bartos, P.M.C.; Gomes, H.T.; Do Amaral, L.I.V.; Teixeira, J.B.; Scherwinski-Pereira, J.E. Biochemical events during somatic embryogenesis in Coffea arabica L. 3 Biotech 2018, 8, 209. [Google Scholar] [CrossRef]
- Pinto, R.T.; Freitas, N.C.; Máximo, W.P.F.; Cardoso, T.B.; Prudente, D.D.O.; Paiva, L.V. Genome-wide analysis, transcription factor network approach and gene expression profile of GH3 genes over early somatic embryogenesis in Coffea spp. BMC Genom. 2019, 20, 812. [Google Scholar] [CrossRef]
- Dinneny, J.R.; Benfey, P.N. Plant Stem Cell Niches: Standing the Test of Time. Cell 2008, 132, 553–557. [Google Scholar] [CrossRef]
- Fehér, A. Callus, Dedifferentiation, Totipotency, somatic embryogenesis: What these terms mean in the era of molecular plant biology? Front. Plant Sci. 2019, 10, 536. [Google Scholar] [CrossRef]
- Aggarwal, S.; Sardana, C.; Ozturk, M.; Sarwat, M. Plant stem cells and their applications: Special emphasis on their marketed products. 3 Biotech 2020, 10, 291. [Google Scholar] [CrossRef]
- Hatfield, S.D.; Shcherbata, H.R.; Fischer, K.A.; Nakahara, K.; Carthew, R.W.; Ruohola-Baker, H. Stem cell division is regulated by the microRNA pathway. Nature 2005, 435, 974–978. [Google Scholar] [CrossRef]
- Xu, M.; Du, Q.; Tian, C.; Wang, Y.; Jiao, Y. Stochastic gene expression drives mesophyll protoplast regeneration. Sci. Adv. 2021, 7, eabg8466. [Google Scholar] [CrossRef]
- Pérez-Núñez, M.T.; Souza, R.; Sáenz, L.; Chan, J.L.; Zúñiga-Aguilar, J.J.; Oropeza, C. Detection of a SERK-like gene in coconut and analysis of its expression during the formation of embryogenic callus and somatic embryos. Plant Cell Rep. 2009, 28, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Takahashi, M.; Fujiwara, S.; Mitsuda, N.; Ohme-Takagi, M. Improving the efficiency of adventitious shoot induction and somatic embryogenesis via modification of WUSCHEL and LEAFY COTYLEDON 1. Plants 2020, 9, 1434. [Google Scholar] [CrossRef] [PubMed]
- Jha, P.; Kumar, V. BABY BOOM (BBM): A candidate transcription factor gene in plant biotechnology. Biotechnol. Lett. 2018, 40, 1467–1475. [Google Scholar] [CrossRef]
- Méndez-Hernández, H.A.; Ledezma-Rodríguez, M.; Avilez-Montalvo, R.N.; Juárez-Gómez, Y.L.; Skeete, A.; Avilez-Montalvo, J.; De-la-Peña, C.; Loyola-Vargas, V.M. Signaling overview of plant somatic embryogenesis. Front. Plant Sci. 2019, 10, 77. [Google Scholar] [CrossRef]
- Kusnandar, A.S.; Itoh, J.-I.; Sato, Y.; Honda, E.; Hibara, K.-I.; Kyozuka, J.; Naramoto, S. NARROW AND DWARF LEAF 1, the orthologue of Arabidopsis ENHANCER OF SHOOT REGENERATION1/DORNRÖSCHEN, mediates leaf development and maintenance of the shoot apical meristem in Oryza sativa L. Plant Cell Physiol. 2021, 63, 265–278. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Jeong, Y.Y.; Lee, H.-Y.; Kim, S.W.; Noh, Y.-S.; Seo, P.J. Optimization of protoplast regeneration in the model plant Arabidopsis thaliana. Plant Methods 2021, 17, 21. [Google Scholar] [CrossRef]
- Lee, E.-K.; Jin, Y.-W.; Park, J.H.; Yoo, Y.M.; Hong, S.M.; Amir, R.; Yan, Z.; Kwon, E.; Elfick, A.; Tomlinson, S.R.; et al. Cultured cambial meristematic cells as a source of plant natural products. Nat. Biotechnol. 2010, 28, 1213–1217. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, J.-E.; Lee, H.-S.; Lee, H.; Cho, H.; Park, S.-Y. Embryogenic Stem Cell Identity after Protoplast Isolation from Daucus carota and Recovery of Regeneration Ability through Protoplast Culture. Int. J. Mol. Sci. 2022, 23, 11556. https://doi.org/10.3390/ijms231911556
Han J-E, Lee H-S, Lee H, Cho H, Park S-Y. Embryogenic Stem Cell Identity after Protoplast Isolation from Daucus carota and Recovery of Regeneration Ability through Protoplast Culture. International Journal of Molecular Sciences. 2022; 23(19):11556. https://doi.org/10.3390/ijms231911556
Chicago/Turabian StyleHan, Jong-Eun, Han-Sol Lee, Hyoshin Lee, Hyunwoo Cho, and So-Young Park. 2022. "Embryogenic Stem Cell Identity after Protoplast Isolation from Daucus carota and Recovery of Regeneration Ability through Protoplast Culture" International Journal of Molecular Sciences 23, no. 19: 11556. https://doi.org/10.3390/ijms231911556
APA StyleHan, J.-E., Lee, H.-S., Lee, H., Cho, H., & Park, S.-Y. (2022). Embryogenic Stem Cell Identity after Protoplast Isolation from Daucus carota and Recovery of Regeneration Ability through Protoplast Culture. International Journal of Molecular Sciences, 23(19), 11556. https://doi.org/10.3390/ijms231911556