
Apoptotic Body-Rich Media from Tenocytes Enhance Proliferation and Migration of Tenocytes and Bone Marrow Stromal Cells

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
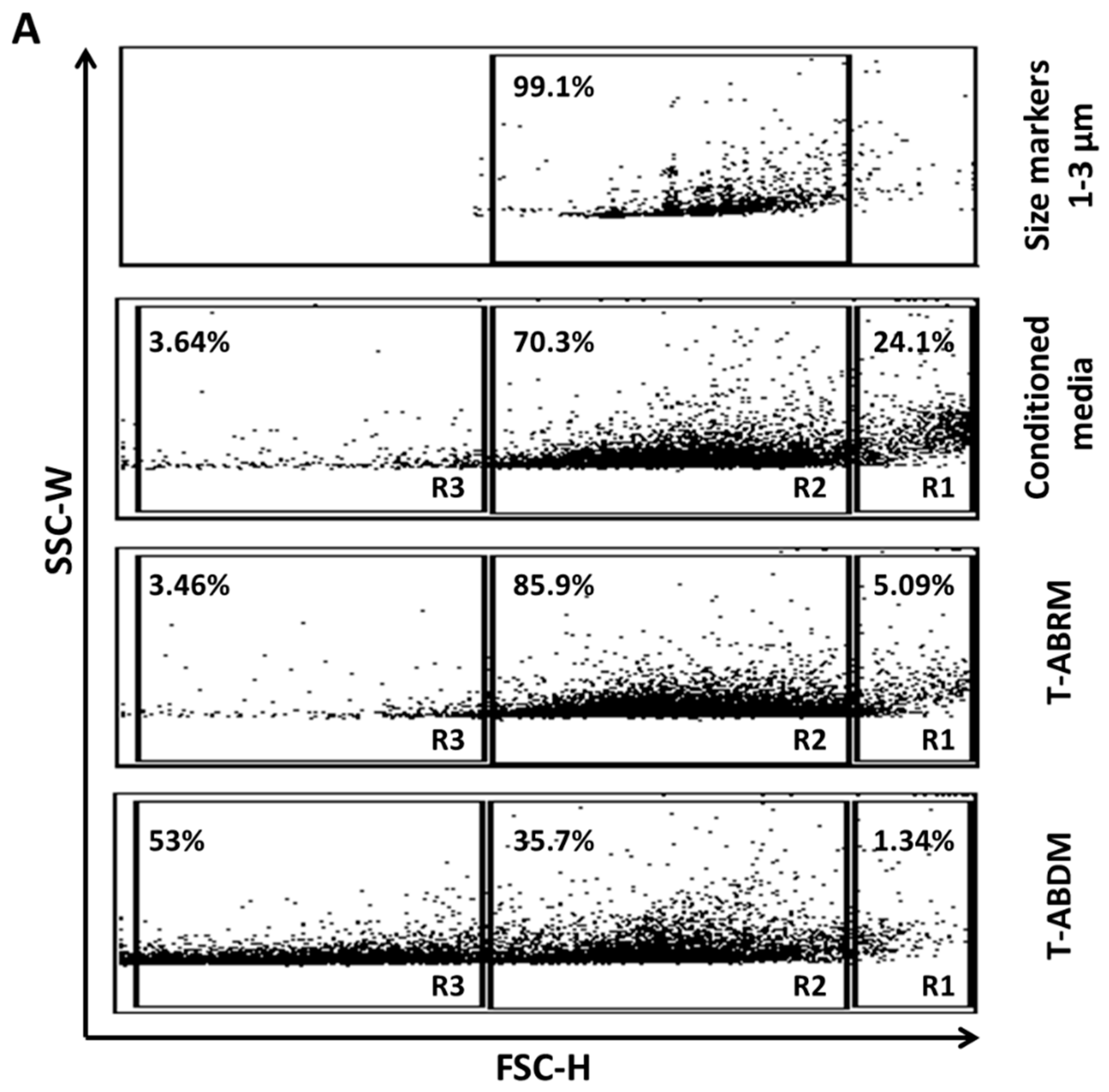
2.1. Characterization of a Cell Culture Conditioning Medium
2.2. T-ABRM and T-ABDM-Mediated Cell Proliferation
2.3. Cell Migration under T-ABRM and T-ABDM Stimulation
2.4. Gene Expression Assay
3. Discussion
4. Materials and Methods
4.1. Isolation of Apoptotic Bodies Derived from Tendon Cells
4.2. Characterization of T-ABRM by Flow Cytometry and Immunohistochemistry
4.3. Measurement of Cell Proliferation and Viability
4.4. Measurement of Cell Migration
4.5. RT and Quantitative Real-Time PCR
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- Calve, S.; Dennis, R.G.; Kosnik, P.E., II; Baar, K.; Grosh, K.; Arruda, E.M. Engineering of functional tendon. Tissue Eng. 2004, 10, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Davis, W.T.; Fletcher, S.A.; Guillamondegui, O.D. Musculoskeletal occupational injury among surgeons: Effects for patients, providers, and institutions. J. Surg. Res. 2014, 189, 207–212.e206. [Google Scholar] [CrossRef] [PubMed]
- Elliot, D. Primary flexor tendon repair–operative repair, pulley management and rehabilitation. J. Hand Surg. 2002, 27, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Gross, G.; Hoffmann, A. Therapeutic strategies for tendon healing based on novel biomaterials, factors and cells. Pathobiol. J. Immunopathol. Mol. Cell. Biol. 2013, 80, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Budovec, J.J.; Sudakoff, G.S.; Dzwierzynski, W.W.; Matloub, H.S.; Sanger, J.R. Sonographic differentiation of digital tendon rupture from adhesive scarring after primary surgical repair. J. Hand Surg. Am. 2006, 31, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Elliot, D.; Giesen, T. Primary flexor tendon surgery: The search for a perfect result. Hand Clin. 2013, 29, 191–206. [Google Scholar] [CrossRef]
- Dahlgren, L.A.; van der Meulen, M.C.; Bertram, J.E.; Starrak, G.S.; Nixon, A.J. Insulin-like growth factor-i improves cellular and molecular aspects of healing in a collagenase-induced model of flexor tendinitis. J. Orthop. Res. 2002, 20, 910–919. [Google Scholar] [CrossRef]
- Godbout, C.; Bilodeau, R.; Van Rooijen, N.; Bouchard, P.; Frenette, J. Transient neutropenia increases macrophage accumulation and cell proliferation but does not improve repair following intratendinous rupture of achilles tendon. J. Orthop. Res. 2010, 28, 1084–1091. [Google Scholar] [CrossRef]
- Dyment, N.A.; Liu, C.F.; Kazemi, N.; Aschbacher-Smith, L.E.; Kenter, K.; Breidenbach, A.P.; Shearn, J.T.; Wylie, C.; Rowe, D.W.; Butler, D.L. The paratenon contributes to scleraxis-expressing cells during patellar tendon healing. PLoS ONE 2013, 8, e59944. [Google Scholar] [CrossRef]
- Boyer, M.I.; Goldfarb, C.A.; Gelberman, R.H. Recent progress in flexor tendon healing. The modulation of tendon healing with rehabilitation variables. J. Hand Ther. 2005, 18, 80–85. [Google Scholar] [CrossRef]
- Gelberman, R.H. Flexor tendon physiology: Tendon nutrition and cellular activity in injury and repair. Instr. Course Lect. 1985, 34, 351–360. [Google Scholar] [PubMed]
- Lutsky, K.F.; Giang, E.L.; Matzon, J.L. Flexor tendon injury, repair and rehabilitation. Orthop. Clin. N. Am. 2015, 46, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Simpson, D.; Meurer, L.; Braza, D. Meeting the scholarly project requirement-application of scholarship criteria beyond research. J. Grad. Med. Educ. 2012, 4, 111–112. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.C.; Cullen, S.P.; Martin, S.J. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell Biol. 2008, 9, 231–241. [Google Scholar] [CrossRef]
- Sharma, P.; Maffulli, N. Biology of tendon injury: Healing, modeling and remodeling. J. Musculoskelet. Neuronal Interact. 2006, 6, 181–190. [Google Scholar]
- Muller, S.A.; Todorov, A.; Heisterbach, P.E.; Martin, I.; Majewski, M. Tendon healing: An overview of physiology, biology, and pathology of tendon healing and systematic review of state of the art in tendon bioengineering. Knee Surg. Sports Traumatol. Arthrosc. Off. J. ESSKA 2013, 23, 2097–2105. [Google Scholar] [CrossRef]
- Asakawa, D.S.; Pappas, G.P.; Blemker, S.S.; Drace, J.E.; Delp, S.L. Cine phase-contrast magnetic resonance imaging as a tool for quantification of skeletal muscle motion. Semin. Musculoskelet. Radiol. 2003, 7, 287–295. [Google Scholar]
- Hatfield, T.R.; Klaus, D.M.; Simske, S.J. An ultrasonic methodology for muscle cross section measurement to support space flight. Microgravity Sci. Technol. 2004, 15, 3–11. [Google Scholar] [CrossRef]
- Wang, J.H.; Wang, J.H.C. Mechanobiology of tendon. J. Biomech. 2006, 39, 1563–1582. [Google Scholar] [CrossRef]
- Molloy, T.; Wang, Y.; Murrell, G. The roles of growth factors in tendon and ligament healing. Sports Med. 2003, 33, 381–394. [Google Scholar] [CrossRef]
- Rothrauff, B.B.; Guang, Y.; Tuan, R.S. Chapter 2—Tendon resident cells—Functions and features in section i—Developmental biology and physiology of tendons. In Tendon Regeneration; Gomes, M.E., Reis, R.L., Rodrigues, M.T., Eds.; Academic Press: Boston, MA, USA, 2015. [Google Scholar]
- Jager, R.; Fearnhead, H.O. “Dead cells talking”: The silent form of cell death is not so quiet. Biochem. Res. Int. 2012, 2012, 453838. [Google Scholar] [CrossRef] [PubMed]
- Maiseyeu, A.; Bagalkot, V. In vitro uptake of apoptotic body mimicking phosphatidylserine-quantum dot micelles by monocytic cell line. Nanoscale Res. Lett. 2014, 9, 176. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.S.; Kobayashi, C.; Agata, K.; Ikeo, K.; Gojobori, T. Detection of apoptosis during planarian regeneration by the expression of apoptosis-related genes and tunel assay. Gene 2004, 333, 15–25. [Google Scholar] [CrossRef]
- Fan, Y.; Bergmann, A. Distinct mechanisms of apoptosis-induced compensatory proliferation in proliferating and differentiating tissues in the drosophila eye. Dev. Cell 2008, 14, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Huang, Q.; Chen, J.; Peng, Y.; Roop, D.R.; Bedford, J.S.; Li, C.Y. Apoptotic cells activate the “phoenix rising” pathway to promote wound healing and tissue regeneration. Sci. Signal. 2010, 3, ra13. [Google Scholar] [CrossRef] [PubMed]
- Jo, C.H.; Lee, Y.G.; Shin, W.H.; Kim, H.; Chai, J.W.; Jeong, E.C.; Kim, J.E.; Shim, H.; Shin, J.S.; Shin, I.S.; et al. Intra-articular injection of mesenchymal stem cells for the treatment of osteoarthritis of the knee: A proof-of-concept clinical trial. Stem Cells 2014, 32, 1254–1266. [Google Scholar] [CrossRef]
- Chang, C.J.; Yen, M.L.; Chen, Y.C.; Chien, C.C.; Huang, H.I.; Bai, C.H.; Yen, B.L. Placenta-derived multipotent cells exhibit immunosuppressive properties that are enhanced in the presence of interferon-gamma. Stem Cells 2006, 24, 2466–2477. [Google Scholar] [CrossRef]
- Selek, O.; Buluc, L.; Muezzinoglu, B.; Ergun, R.E.; Ayhan, S.; Karaoz, E. Mesenchymal stem cell application improves tendon healing via anti-apoptotic effect (animal study). Acta Orthop. Traumatol. Turc. 2014, 48, 187–195. [Google Scholar] [CrossRef]
- Hristov, M.; Erl, W.; Linder, S.; Weber, P.C. Apoptotic bodies from endothelial cells enhance the number and initiate the differentiation of human endothelial progenitor cells in vitro. Blood 2004, 104, 2761–2766. [Google Scholar] [CrossRef]
- Barron, L.; Wynn, T.A. Fibrosis is regulated by th2 and th17 responses and by dynamic interactions between fibroblasts and macrophages. American journal of physiology. Gastrointest. Liver Physiol. 2011, 300, G723–G728. [Google Scholar] [CrossRef]
- Song, J. Emt or apoptosis: A decision for tgf-beta. Cell Res. 2007, 17, 289–290. [Google Scholar] [CrossRef] [PubMed]
- Thoreson, A.R.; Hiwatari, R.; An, K.N.; Amadio, P.C.; Zhao, C. The effect of 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide suture coating on tendon repair strength and cell viability in a canine model. J. Hand. Surg. Am. 2015, 40, 1986–1991. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.E.; Azevedo, D.O.; Campos, L.A.; Zanuncio, J.C.; Serrao, J.E. Melipona quadrifasciata (hymenoptera: Apidae) fat body persists through metamorphosis with a few apoptotic cells and an increased autophagy. Protoplasma 2015, 252, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Chieh, H.F.; Bakri, K.; Ikeda, J.; Sun, Y.L.; Moran, S.L.; An, K.N.; Amadio, P.C. The effects of bone marrow stromal cell transplants on tendon healing in vitro. Med. Eng. Phys. 2009, 31, 1271–1275. [Google Scholar] [CrossRef][Green Version]
- Hayashi, M.; Zhao, C.; An, K.N.; Amadio, P.C. The effects of growth and differentiation factor 5 on bone marrow stromal cell transplants in an in vitro tendon healing model. J. Hand Surg. Eur. Vol. 2011, 36, 271–279. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, C.; Gingery, A.; Amadio, P.C.; An, K.-N.; Moran, S.L.; Zhao, C. Apoptotic Body-Rich Media from Tenocytes Enhance Proliferation and Migration of Tenocytes and Bone Marrow Stromal Cells. Int. J. Mol. Sci. 2022, 23, 11475. https://doi.org/10.3390/ijms231911475
Dong C, Gingery A, Amadio PC, An K-N, Moran SL, Zhao C. Apoptotic Body-Rich Media from Tenocytes Enhance Proliferation and Migration of Tenocytes and Bone Marrow Stromal Cells. International Journal of Molecular Sciences. 2022; 23(19):11475. https://doi.org/10.3390/ijms231911475
Chicago/Turabian StyleDong, Chenhui, Anne Gingery, Peter C. Amadio, Kai-Nan An, Steven L. Moran, and Chunfeng Zhao. 2022. "Apoptotic Body-Rich Media from Tenocytes Enhance Proliferation and Migration of Tenocytes and Bone Marrow Stromal Cells" International Journal of Molecular Sciences 23, no. 19: 11475. https://doi.org/10.3390/ijms231911475
APA StyleDong, C., Gingery, A., Amadio, P. C., An, K.-N., Moran, S. L., & Zhao, C. (2022). Apoptotic Body-Rich Media from Tenocytes Enhance Proliferation and Migration of Tenocytes and Bone Marrow Stromal Cells. International Journal of Molecular Sciences, 23(19), 11475. https://doi.org/10.3390/ijms231911475