

CD3+/CD56+ NKT-like Cells Show Imbalanced Control Immediately after Exercise in Delayed-Onset Muscle Soreness

 , , , and
, , , and
Abstract
1. Introduction
2. Results
2.1. Study Population
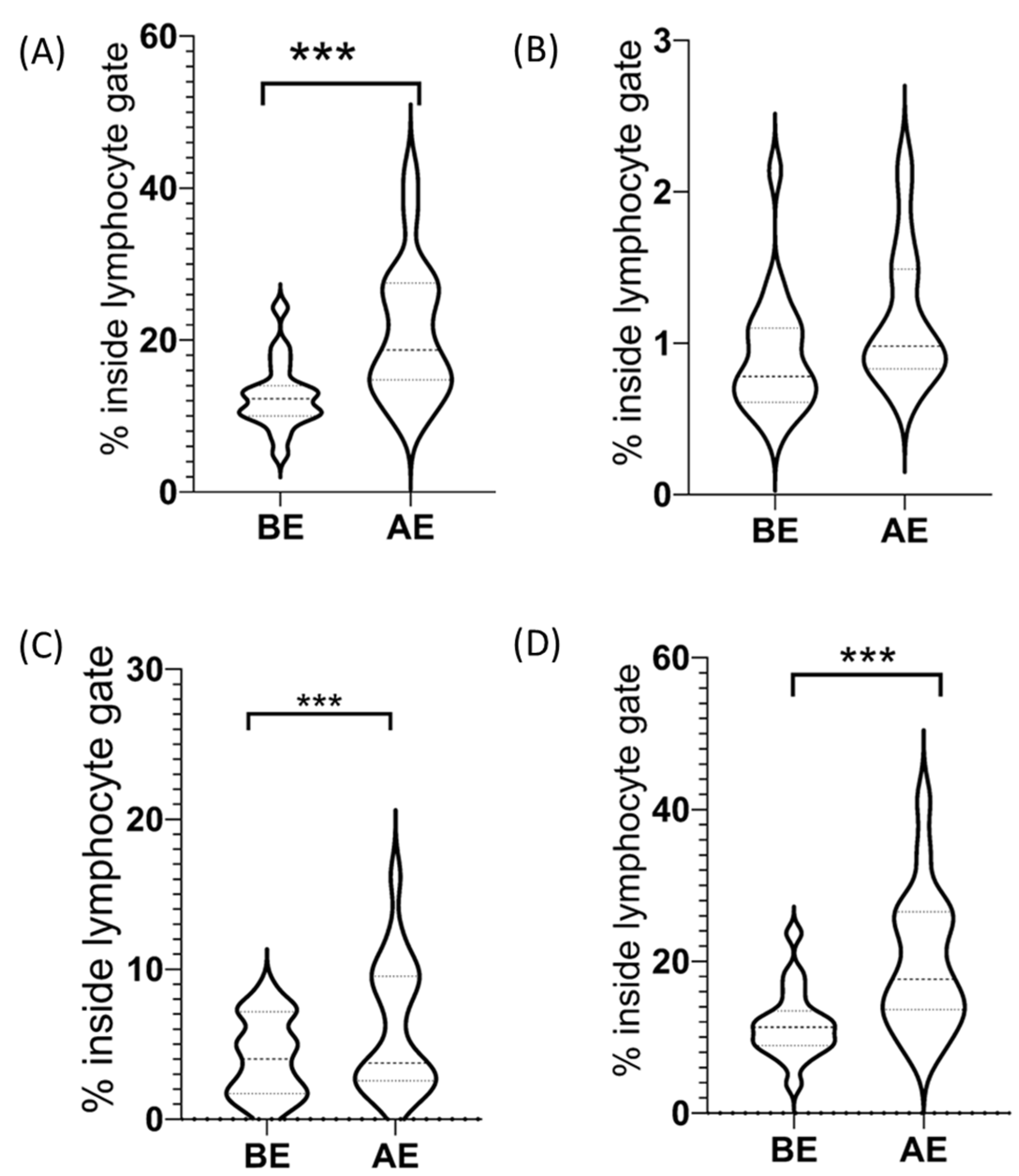
2.2. CD3−/CD56+ NK Cells among Lymphocytes
2.2.1. CD3−/CD56dim+ NK Cells among Lymphocytes
2.2.2. CD3−/CD56bright+ NK Cells among Lymphocytes
2.3. CD3+ T Cells among Lymphocytes
CD3+/CD56+ NKT-like Cells among Lymphocytes
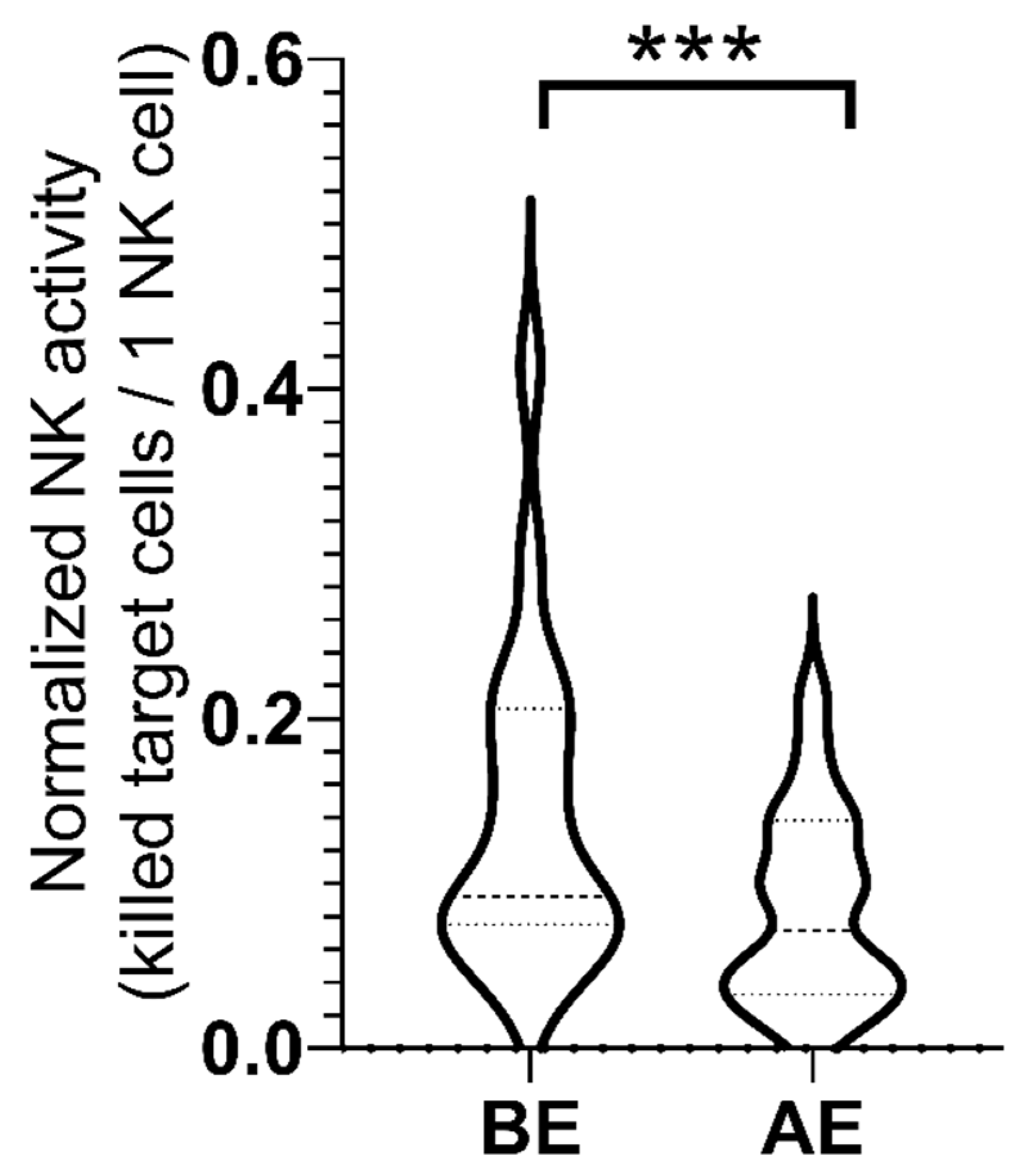
2.4. NK Cell Activity
3. Discussion
Limitations
4. Materials and Methods
4.1. Study Design
4.2. Participants
4.3. Procedures
4.3.1. Sports Cardiology Screening with Wall-Sit Test
4.3.2. Natural Killer Cell Activity
4.3.3. PBMC Seperation
4.3.4. Immunophenotyping of Circulating NK Cells
4.3.5. Flow Cytometry Measurements
4.4. Statistical Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clarkson, P.M.; Nosaka, K.; Braun, B. Muscle function after exercise-induced muscle damage and rapid adaptation. Med. Sci. Sports Exerc. 1992, 24, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Torres, R.; Vasques, J.; Duarte, J.A.; Cabri, J.M. Knee proprioception after exercise-induced muscle damage. Int. J. Sports Med. 2010, 31, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Deschenes, M.R.; Brewer, R.E.; Bush, J.A.; McCoy, R.W.; Volek, J.S.; Kraemer, W.J. Neuromuscular disturbance outlasts other symptoms of exercise-induced muscle damage. J. Neurol. Sci. 2000, 174, 92–99. [Google Scholar] [CrossRef]
- Gleeson, M.; Blannin, A.K.; Walsh, N.P.; Field, C.N.; Pritchard, J.C. Effect of exercise-induced muscle damage on the blood lactate response to incremental exercise in humans. Eur. J. Appl Physiol. Occup. Physiol. 1998, 77, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Mizumura, K.; Taguchi, T. Delayed onset muscle soreness: Involvement of neurotrophic factors. J. Physiol. Sci. 2016, 66, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.; Hume, P.; Maxwell, L. Delayed onset muscle soreness: Treatment strategies and performance factors. Sports Med. 2003, 33, 145–164. [Google Scholar] [CrossRef] [PubMed]
- Romero-Moraleda, B.; La Touche, R.; Lerma-Lara, S.; Ferrer-Pena, R.; Paredes, V.; Peinado, A.B.; Munoz-Garcia, D. Neurodynamic mobilization and foam rolling improved delayed-onset muscle soreness in a healthy adult population: A randomized controlled clinical trial. PeerJ 2017, 5, e3908. [Google Scholar] [CrossRef]
- Luján, R.; Carrera, R.A.; Moreira, O.; Candia Sosa, K.; De Paz Fernandez, J.A. Massage in The Prevention and Treatment of Delayed Onset Muscle Soreness: An Updated Systematic Review. Rev. Ib. CC. Act. Fís. Dep. 2017, 6, 38–56. [Google Scholar]
- Sonkodi, B.; Berkes, I.; Koltai, E. Have We Looked in the Wrong Direction for More Than 100 Years? Delayed Onset Muscle Soreness Is, in Fact, Neural Microdamage Rather Than Muscle Damage. Antioxidants 2020, 9, 212. [Google Scholar] [CrossRef]
- Close, G.L.; Ashton, T.; Cable, T.; Doran, D.; MacLaren, D.P. Eccentric exercise, isokinetic muscle torque and delayed onset muscle soreness: The role of reactive oxygen species. Eur. J. Appl. Physiol. 2004, 91, 615–621. [Google Scholar] [CrossRef]
- Lippi, L.; de Sire, A.; Mezian, K.; Curci, C.; Perrero, L.; Turco, A.; Andaloro, S.; Ammendolia, A.; Fusco, N.; Invernizzi, M. Impact of exercise training on muscle mitochondria modifications in older adults: A systematic review of randomized controlled trials. Aging Clin. Exp. Res. 2022, 34, 1495–1510. [Google Scholar] [CrossRef] [PubMed]
- Sonkodi, B. Delayed Onset Muscle Soreness and Critical Neural Microdamage-Derived Neuroinflammation. Biomolecules 2022, 12, 1207. [Google Scholar] [CrossRef]
- Pappas, A.; Tsiokanos, A.; Fatouros, I.G.; Poulios, A.; Kouretas, D.; Goutzourelas, N.; Giakas, G.; Jamurtas, A.Z. The Effects of Spirulina Supplementation on Redox Status and Performance Following a Muscle Damaging Protocol. Int. J. Mol. Sci. 2021, 22, 3559. [Google Scholar] [CrossRef] [PubMed]
- Ziemkiewicz, N.; Hilliard, G.; Pullen, N.A.; Garg, K. The Role of Innate and Adaptive Immune Cells in Skeletal Muscle Regeneration. Int. J. Mol. Sci. 2021, 22, 3265. [Google Scholar] [CrossRef]
- Camus, G.; Deby-Dupont, G.; Duchateau, J.; Deby, C.; Pincemail, J.; Lamy, M. Are similar inflammatory factors involved in strenuous exercise and sepsis? Intensive Care Med. 1994, 20, 602–610. [Google Scholar] [CrossRef]
- Borghi, S.M.; Bussulo, S.K.D.; Pinho-Ribeiro, F.A.; Fattori, V.; Carvalho, T.T.; Rasquel-Oliveira, F.S.; Zaninelli, T.H.; Ferraz, C.R.; Casella, A.M.B.; Cunha, F.Q.; et al. Intense Acute Swimming Induces Delayed-Onset Muscle Soreness Dependent on Spinal Cord Neuroinflammation. Front. Pharmacol. 2021, 12, 734091. [Google Scholar] [CrossRef]
- Bennett, G.J.; Liu, G.K.; Xiao, W.H.; Jin, H.W.; Siau, C. Terminal arbor degeneration--a novel lesion produced by the antineoplastic agent paclitaxel. Eur. J. Neurosci. 2011, 33, 1667–1676. [Google Scholar] [CrossRef]
- Sonkodi, B.; Kopa, Z.; Nyirady, P. Post Orgasmic Illness Syndrome (POIS) and Delayed Onset Muscle Soreness (DOMS): Do They Have Anything in Common? Cells 2021, 10, 1867. [Google Scholar] [CrossRef]
- De Amicis, K.; Costa, P.R.; Figo, D.D.; De Lima, C.M.F.; Castro, F.F.M.; Kalil, J.; Santos, K.S.; Galvao, C.E.S. Immunophenotypical Characterization of a Brazilian POIS (Post-Orgasmic Illness Syndrome) Patient: Adding More Pieces to Puzzle. J. Sex Marital Ther. 2020, 46, 227–233. [Google Scholar] [CrossRef]
- Sonkodi, B. Delayed Onset Muscle Soreness (DOMS): The Repeated Bout Effect and Chemotherapy-Induced Axonopathy May Help Explain the Dying-Back Mechanism in Amyotrophic Lateral Sclerosis and Other Neurodegenerative Diseases. Brain Sci. 2021, 11, 108. [Google Scholar] [CrossRef]
- Sonkodi, B.; Hortobágyi, T. Amyotrophic lateral sclerosis and delayed onset muscle soreness in light of the impaired blink and stretch reflexes – watch out for Piezo2. Open Med. 2022, 17, 397–402. [Google Scholar] [CrossRef]
- Ingre, C.; Roos, P.M.; Piehl, F.; Kamel, F.; Fang, F. Risk factors for amyotrophic lateral sclerosis. Clin. Epidemiol. 2015, 7, 181–193. [Google Scholar] [CrossRef]
- Rolfes, L.; Schulte-Mecklenbeck, A.; Schreiber, S.; Vielhaber, S.; Herty, M.; Marten, A.; Pfeuffer, S.; Ruck, T.; Wiendl, H.; Gross, C.C.; et al. Amyotrophic lateral sclerosis patients show increased peripheral and intrathecal T-cell activation. Brain Commun. 2021, 3, fcab157. [Google Scholar] [CrossRef]
- Rentzos, M.; Evangelopoulos, E.; Sereti, E.; Zouvelou, V.; Marmara, S.; Alexakis, T.; Evdokimidis, I. Alterations of T cell subsets in ALS: A systemic immune activation? Acta Neurol. Scand. 2012, 125, 260–264. [Google Scholar] [CrossRef]
- Maruyama, H.; Morino, H.; Ito, H.; Izumi, Y.; Kato, H.; Watanabe, Y.; Kinoshita, Y.; Kamada, M.; Nodera, H.; Suzuki, H.; et al. Mutations of optineurin in amyotrophic lateral sclerosis. Nature 2010, 465, 223–226. [Google Scholar] [CrossRef]
- Millard, A.L.; Valli, P.V.; Stussi, G.; Mueller, N.J.; Yung, G.P.; Seebach, J.D. Brief Exercise Increases Peripheral Blood NK Cell Counts without Immediate Functional Changes, but Impairs their Responses to ex vivo Stimulation. Front. Immunol. 2013, 4, 125. [Google Scholar] [CrossRef]
- Pizza, F.X.; Mitchell, J.B.; Davis, B.H.; Starling, R.D.; Holtz, R.W.; Bigelow, N. Exercise-induced muscle damage: Effect on circulating leukocyte and lymphocyte subsets. Med. Sci. Sports Exerc. 1995, 27, 363–370. [Google Scholar] [CrossRef]
- Malm, C.; Lenkei, R.; Sjodin, B. Effects of eccentric exercise on the immune system in men. J. Appl. Physiol. 1999, 86, 461–468. [Google Scholar] [CrossRef]
- Jiang, Y.; Cui, X.; Cui, C.; Zhang, J.; Zhou, F.; Zhang, Z.; Fu, Y.; Xu, J.; Chu, Z.; Liu, J.; et al. The function of CD3+CD56+ NKT-like cells in HIV-infected individuals. BioMed Res. Int. 2014, 2014, 863625. [Google Scholar] [CrossRef]
- Doherty, D.G.; O’Farrelly, C. Innate and adaptive lymphoid cells in the human liver. Immunol. Rev. 2000, 174, 5–20. [Google Scholar] [CrossRef]
- Heck, T.G.; Scomazzon, S.P.; Nunes, P.R.; Scholer, C.M.; da Silva, G.S.; Bittencourt, A.; Faccioni-Heuser, M.C.; Krause, M.; Bazotte, R.B.; Curi, R.; et al. Acute exercise boosts cell proliferation and the heat shock response in lymphocytes: Correlation with cytokine production and extracellular-to-intracellular HSP70 ratio. Cell Stress Chaperones 2017, 22, 271–291. [Google Scholar] [CrossRef]
- Kaszubowska, L.; Foerster, J.; Kmiec, Z. NKT-like (CD3 + CD56+) cells differ from T cells in expression level of cellular protective proteins and sensitivity to stimulation in the process of ageing. Immun. Ageing 2022, 19, 18. [Google Scholar] [CrossRef] [PubMed]
- Martinez de Toda, I.; De la Fuente, M. The role of Hsp70 in oxi-inflamm-aging and its use as a potential biomarker of lifespan. Biogerontology 2015, 16, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, R.S.; Veras, F.P.; Ferreira, D.W.; Sant’Anna, M.B.; Lollo, P.C.B.; Cunha, T.M.; Galdino, G. Involvement of the Hsp70/TLR4/IL-6 and TNF-alpha pathways in delayed-onset muscle soreness. J. Neurochem. 2020, 155, 29–44. [Google Scholar] [CrossRef] [PubMed]
- De Lemos Muller, C.H.; de Matos, J.R.; Grigolo, G.B.; Schroeder, H.T.; Rodrigues-Krause, J.; Krause, M. Exercise Training for the Elderly: Inflammaging and the Central Role for HSP70. J. Sci. Sport Exerc. 2019, 1, 97–115. [Google Scholar] [CrossRef]
- Fang, H.; Wu, Y.; Huang, X.; Wang, W.; Ang, B.; Cao, X.; Wan, T. Toll-like receptor 4 (TLR4) is essential for Hsp70-like protein 1 (HSP70L1) to activate dendritic cells and induce Th1 response. J. Biol. Chem. 2011, 286, 30393–30400. [Google Scholar] [CrossRef] [PubMed]
- Djavaheri-Mergny, M.; Amelotti, M.; Mathieu, J.; Besancon, F.; Bauvy, C.; Souquere, S.; Pierron, G.; Codogno, P. NF-kappaB activation represses tumor necrosis factor-alpha-induced autophagy. J. Biol. Chem. 2006, 281, 30373–30382. [Google Scholar] [CrossRef] [PubMed]
- Rubinsztein, D.C.; Marino, G.; Kroemer, G. Autophagy and aging. Cell 2011, 146, 682–695. [Google Scholar] [CrossRef]
- Nimmervoll, B.; Chtcheglova, L.A.; Juhasz, K.; Cremades, N.; Aprile, F.A.; Sonnleitner, A.; Hinterdorfer, P.; Vigh, L.; Preiner, J.; Balogi, Z. Cell surface localised Hsp70 is a cancer specific regulator of clathrin-independent endocytosis. FEBS Lett. 2015, 589, 2747–2753. [Google Scholar] [CrossRef]
- Hantschel, M.; Pfister, K.; Jordan, A.; Scholz, R.; Andreesen, R.; Schmitz, G.; Schmetzer, H.; Hiddemann, W.; Multhoff, G. Hsp70 plasma membrane expression on primary tumor biopsy material and bone marrow of leukemic patients. Cell Stress Chaperones 2000, 5, 438–442. [Google Scholar] [CrossRef]
- Multhoff, G.; Botzler, C.; Jennen, L.; Schmidt, J.; Ellwart, J.; Issels, R. Heat shock protein 72 on tumor cells: A recognition structure for natural killer cells. J. Immunol. 1997, 158, 4341–4350. [Google Scholar]
- Vega, V.L.; Rodriguez-Silva, M.; Frey, T.; Gehrmann, M.; Diaz, J.C.; Steinem, C.; Multhoff, G.; Arispe, N.; De Maio, A. Hsp70 translocates into the plasma membrane after stress and is released into the extracellular environment in a membrane-associated form that activates macrophages. J. Immunol. 2008, 180, 4299–4307. [Google Scholar] [CrossRef]
- Buyan, A.; Cox, C.D.; Barnoud, J.; Li, J.; Chan, H.S.M.; Martinac, B.; Marrink, S.J.; Corry, B. Piezo1 Forms Specific, Functionally Important Interactions with Phosphoinositides and Cholesterol. Biophys. J. 2020, 119, 1683–1697. [Google Scholar] [CrossRef]
- Chong, J.; De Vecchis, D.; Hyman, A.J.; Povstyan, O.V.; Ludlow, M.J.; Shi, J.; Beech, D.J.; Kalli, A.C. Modeling of full-length Piezo1 suggests importance of the proximal N-terminus for dome structure. Biophys. J. 2021, 120, 1343–1356. [Google Scholar] [CrossRef]
- Sonkodi, B.; Resch, M.D.; Hortobágyi, T. Is the Sex Difference a Clue to the Pathomechanism of Dry Eye Disease? Watch out for the NGF-TrkA-Piezo2 Signaling Axis and the Piezo2 Channelopathy. J. Mol. Neurosci. 2022, 72, 1598–1608. [Google Scholar] [CrossRef]
- Mosesson, Y.; Mills, G.B.; Yarden, Y. Derailed endocytosis: An emerging feature of cancer. Nat. Rev. Cancer 2008, 8, 835–850. [Google Scholar] [CrossRef]
- Vincent, J.A.; Wieczerzak, K.B.; Gabriel, H.M.; Nardelli, P.; Rich, M.M.; Cope, T.C. A novel path to chronic proprioceptive disability with oxaliplatin: Distortion of sensory encoding. Neurobiol. Dis. 2016, 95, 54–65. [Google Scholar] [CrossRef][Green Version]
- Vincent, J.A.; Nardelli, P.; Gabriel, H.M.; Deardorff, A.S.; Cope, T.C. Complex impairment of IA muscle proprioceptors following traumatic or neurotoxic injury. J. Anat. 2015, 227, 221–230. [Google Scholar] [CrossRef]
- Bullinger, K.L.; Nardelli, P.; Pinter, M.J.; Alvarez, F.J.; Cope, T.C. Permanent central synaptic disconnection of proprioceptors after nerve injury and regeneration. II. Loss of functional connectivity with motoneurons. J. Neurophysiol. 2011, 106, 2471–2485. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, F.J.; Titus-Mitchell, H.E.; Bullinger, K.L.; Kraszpulski, M.; Nardelli, P.; Cope, T.C. Permanent central synaptic disconnection of proprioceptors after nerve injury and regeneration. I. Loss of VGLUT1/IA synapses on motoneurons. J. Neurophysiol. 2011, 106, 2450–2470. [Google Scholar] [CrossRef]
- Chang, W.; Gu, J.G. Role of microtubules in Piezo2 mechanotransduction of mouse Merkel cells. J. Neurophysiol. 2020, 124, 1824–1831. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.H.; Lukacs, V.; de Nooij, J.C.; Zaytseva, D.; Criddle, C.R.; Francisco, A.; Jessell, T.M.; Wilkinson, K.A.; Patapoutian, A. Piezo2 is the principal mechanotransduction channel for proprioception. Nat. Neurosci. 2015, 18, 1756–1762. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.J.; Rinaldi, S.; Costigan, M.; Oh, S.B. Cytotoxic Immunity in Peripheral Nerve Injury and Pain. Front. Neurosci. 2020, 14, 142. [Google Scholar] [CrossRef] [PubMed]
- Sonkodi, B.; Hegedus, A.; Kopper, B.; Berkes, I. Significantly Delayed Medium-Latency Response of the Stretch Reflex in Delayed-Onset Muscle Soreness of the Quadriceps Femoris Muscles Is Indicative of Sensory Neuronal Microdamage. J. Funct. Morphol. Kinesiol. 2022, 7, 43. [Google Scholar] [CrossRef]
- Liu, M.; Li, Y.; Zhong, J.; Xia, L.; Dou, N. The effect of IL-6/Piezo2 on the trigeminal neuropathic pain. Aging 2021, 13, 13615–13625. [Google Scholar] [CrossRef]
- Sonkodi, B.; Bardoni, R.; Poór, G. Osteoporosis in Light of a New Mechanism Theory of Delayed Onset Muscle Soreness and Non-Contact Anterior Cruciate Ligament Injury. Int. J. Mol. Sci. 2022, 23, 9046. [Google Scholar] [CrossRef]
- Wang, X.; Chen, X.; Rodenkirch, L.; Simonson, W.; Wernimont, S.; Ndonye, R.M.; Veerapen, N.; Gibson, D.; Howell, A.R.; Besra, G.S.; et al. Natural killer T-cell autoreactivity leads to a specialized activation state. Blood 2008, 112, 4128–4138. [Google Scholar] [CrossRef][Green Version]
- Hody, S.; Croisier, J.L.; Bury, T.; Rogister, B.; Leprince, P. Eccentric Muscle Contractions: Risks and Benefits. Front. Physiol. 2019, 10, 536. [Google Scholar] [CrossRef]
- Morgan, D.L.; Allen, D.G. Early events in stretch-induced muscle damage. J. Appl. Physiol. 1999, 87, 2007–2015. [Google Scholar] [CrossRef]
- Pollak, K.A.; Swenson, J.D.; Vanhaitsma, T.A.; Hughen, R.W.; Jo, D.; White, A.T.; Light, K.C.; Schweinhardt, P.; Amann, M.; Light, A.R. Exogenously applied muscle metabolites synergistically evoke sensations of muscle fatigue and pain in human subjects. Exp. Physiol. 2014, 99, 368–380. [Google Scholar] [CrossRef]
- Hu, J.; Chiang, L.Y.; Koch, M.; Lewin, G.R. Evidence for a protein tether involved in somatic touch. EMBO J. 2010, 29, 855–867. [Google Scholar] [CrossRef]
- Gibson, W.; Arendt-Nielsen, L.; Taguchi, T.; Mizumura, K.; Graven-Nielsen, T. Increased pain from muscle fascia following eccentric exercise: Animal and human findings. Exp. Brain Res. 2009, 194, 299–308. [Google Scholar] [CrossRef]
- Nencini, S.; Morgan, M.; Thai, J.; Jobling, A.I.; Mazzone, S.B.; Ivanusic, J.J. Piezo2 Knockdown Inhibits Noxious Mechanical Stimulation and NGF-Induced Sensitization in A-Delta Bone Afferent Neurons. Front. Physiol. 2021, 12, 644929. [Google Scholar] [CrossRef]
- Bunting, M.D.; Vyas, M.; Requesens, M.; Langenbucher, A.; Schiferle, E.B.; Manguso, R.T.; Lawrence, M.S.; Demehri, S. Extracellular matrix proteins regulate NK cell function in peripheral tissues. Sci. Adv. 2022, 8, eabk3327. [Google Scholar] [CrossRef]
- Ralainirina, N.; Brons, N.H.; Ammerlaan, W.; Hoffmann, C.; Hentges, F.; Zimmer, J. Mouse natural killer (NK) cells express the nerve growth factor receptor TrkA, which is dynamically regulated. PLoS ONE 2010, 5, e15053. [Google Scholar] [CrossRef]
- Geng, J.; Shi, Y.; Zhang, J.; Yang, B.; Wang, P.; Yuan, W.; Zhao, H.; Li, J.; Qin, F.; Hong, L.; et al. TLR4 signalling via Piezo1 engages and enhances the macrophage mediated host response during bacterial infection. Nat. Commun. 2021, 12, 3519. [Google Scholar] [CrossRef] [PubMed]
- Qorri, B.; Kalaydina, R.V.; Velickovic, A.; Kaplya, Y.; Decarlo, A.; Szewczuk, M.R. Agonist-Biased Signaling via Matrix Metalloproteinase-9 Promotes Extracellular Matrix Remodeling. Cells 2018, 7, 117. [Google Scholar] [CrossRef]
- Sonkodi, B. Should We Void Lactate in the Pathophysiology of Delayed Onset Muscle Soreness? Not So Fast! Let’s See a Neurocentric View! Metabolites 2022, 12, 857. [Google Scholar] [CrossRef]
- Kumar, A.; Pyaram, K.; Yarosz, E.L.; Hong, H.; Lyssiotis, C.A.; Giri, S.; Chang, C.H. Enhanced oxidative phosphorylation in NKT cells is essential for their survival and function. Proc. Natl. Acad. Sci. USA 2019, 116, 7439–7448. [Google Scholar] [CrossRef]
- Magistretti, P.J.; Pellerin, L. Astrocytes Couple Synaptic Activity to Glucose Utilization in the Brain. News Physiol. Sci. 1999, 14, 177–182. [Google Scholar] [CrossRef]
- Belanger, M.; Allaman, I.; Magistretti, P.J. Brain energy metabolism: Focus on astrocyte-neuron metabolic cooperation. Cell Metab. 2011, 14, 724–738. [Google Scholar] [CrossRef] [PubMed]
- Mason, S. Lactate Shuttles in Neuroenergetics-Homeostasis, Allostasis and Beyond. Front. Neurosci. 2017, 11, 43. [Google Scholar] [CrossRef] [PubMed]
- Csulak, E.; Petrov, A.; Kovats, T.; Tokodi, M.; Lakatos, B.; Kovacs, A.; Staub, L.; Suhai, F.I.; Szabo, E.L.; Dohy, Z.; et al. The Impact of COVID-19 on the Preparation for the Tokyo Olympics: A Comprehensive Performance Assessment of Top Swimmers. Int. J. Environ. Res. Public Health 2021, 18, 9770. [Google Scholar] [CrossRef]
- Zarcone, D.; Tilden, A.B.; Cloud, G.; Friedman, H.M.; Landay, A.; Grossi, C.E. Flow cytometry evaluation of cell-mediated cytotoxicity. J. Immunol. Methods 1986, 94, 247–255. [Google Scholar] [CrossRef]
- Zamai, L.; Mariani, A.R.; Zauli, G.; Rodella, L.; Rezzani, R.; Manzoli, F.A.; Vitale, M. Kinetics of in vitro natural killer activity against K562 cells as detected by flow cytometry. Cytometry 1998, 32, 280–285. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Analyte | Before Exercise (BE) | After Exercise (AE) | Significance |
|---|---|---|---|
| CD3−/CD56+ NK cells (% inside lymphocyte gate ± SE) | 12.60 ± 1.01 | 20.96 ± 2.04 | p = 0.0001 |
| CD3+/CD56+ NKT-like cells (% inside lymphocyte gate ± SE) | 4.13 ± 0.59 | 6.14 ± 0.98 | p = 0.0004 |
| CD3−/CD56dim+ NK cell subset (% inside lymphocyte gate ± SE) | 11.69 ± 1.04 | 19.79 ± 2.05 | p = 0.0002 |
| CD3−/CD56bright+ NK cell subset (% inside lymphocyte gate ± SE) | 0.91 ± 0.09 | 1.16 ± 0.10 | n.s. (p = 0.1) |
| Normalized NK cell activity (killed target cells/1 NK cell ± SE) | 0.14 ± 0.02 | 0.08 ± 0.01 | p = 0.0009 |
| CD3+ T cells (% inside lymphocyte gate ± SE) | 65.54 ± 1.53 | 58.03 ± 2.02 | p < 0.0001 |
| Age (years) | 16.5 ± 2.8 |
| Female (n,%) | 10 (53%) |
| Training (hours/week) | 21 ± 2.5 |
| Training History (years) | 11.7 ± 3.2 |
| Last Training before Exam (h) | 22.7 ± 14.1 |
| Last Training Length (min) | 105 ± 39.5 |
| Wall-Sit Time (s) | 127.1 ± 46.2 |
| Muscle Fever Pre-Exercise (VAS 1-10) | 1.42 ± 2.0 |
| Muscle Fever Post-Exercise 1th day (VAS 1-10) | 3.2 ± 2.6 |
| Muscle Fever Post-Exercise 2nd day (VAS 1-10) | 2.9 ±2.5 |
| CK before Exercise (U/L) | 202 ± 125 |
| CK after Exercise (U/L) | 219 ± 123 |
| CKMB before Exercise (U/L) | 18 ± 3.9 |
| CKMB after Exercise (U/L) | 19 ± 3.3 |
| Troponin T before Exercise (ng/mL) | 9.4 ± 8.0 |
| Troponin T after Exercise (ng/mL) | 9.5 ± 7.4 |
| Load Time (min) | 13.4 ± 1.1 |
| Peak Lactate (mmol/L) | 10.4 ± 2.6 |
| Restitution Lactate (mmol/L) | 9.6 ± 2.7 |
| VO2 Max Male (mL/kg/min) | 58.2 ± 1.8 |
| VO2 Max Female (mL/kg/min) | 53.4 ± 3.1 |
| VO2 Max (mL/kg/min) | 55.1 ± 3.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sonkodi, B.; Pállinger, É.; Radovits, T.; Csulak, E.; Shenker-Horváth, K.; Kopper, B.; Buzás, E.I.; Sydó, N.; Merkely, B. CD3+/CD56+ NKT-like Cells Show Imbalanced Control Immediately after Exercise in Delayed-Onset Muscle Soreness. Int. J. Mol. Sci. 2022, 23, 11117. https://doi.org/10.3390/ijms231911117
Sonkodi B, Pállinger É, Radovits T, Csulak E, Shenker-Horváth K, Kopper B, Buzás EI, Sydó N, Merkely B. CD3+/CD56+ NKT-like Cells Show Imbalanced Control Immediately after Exercise in Delayed-Onset Muscle Soreness. International Journal of Molecular Sciences. 2022; 23(19):11117. https://doi.org/10.3390/ijms231911117
Chicago/Turabian StyleSonkodi, Balázs, Éva Pállinger, Tamás Radovits, Emese Csulak, Kinga Shenker-Horváth, Bence Kopper, Edit I. Buzás, Nóra Sydó, and Béla Merkely. 2022. "CD3+/CD56+ NKT-like Cells Show Imbalanced Control Immediately after Exercise in Delayed-Onset Muscle Soreness" International Journal of Molecular Sciences 23, no. 19: 11117. https://doi.org/10.3390/ijms231911117
APA StyleSonkodi, B., Pállinger, É., Radovits, T., Csulak, E., Shenker-Horváth, K., Kopper, B., Buzás, E. I., Sydó, N., & Merkely, B. (2022). CD3+/CD56+ NKT-like Cells Show Imbalanced Control Immediately after Exercise in Delayed-Onset Muscle Soreness. International Journal of Molecular Sciences, 23(19), 11117. https://doi.org/10.3390/ijms231911117