

Heat Stress Decreases Rice Grain Weight: Evidence and Physiological Mechanisms of Heat Effects Prior to Flowering



Abstract
1. Introduction
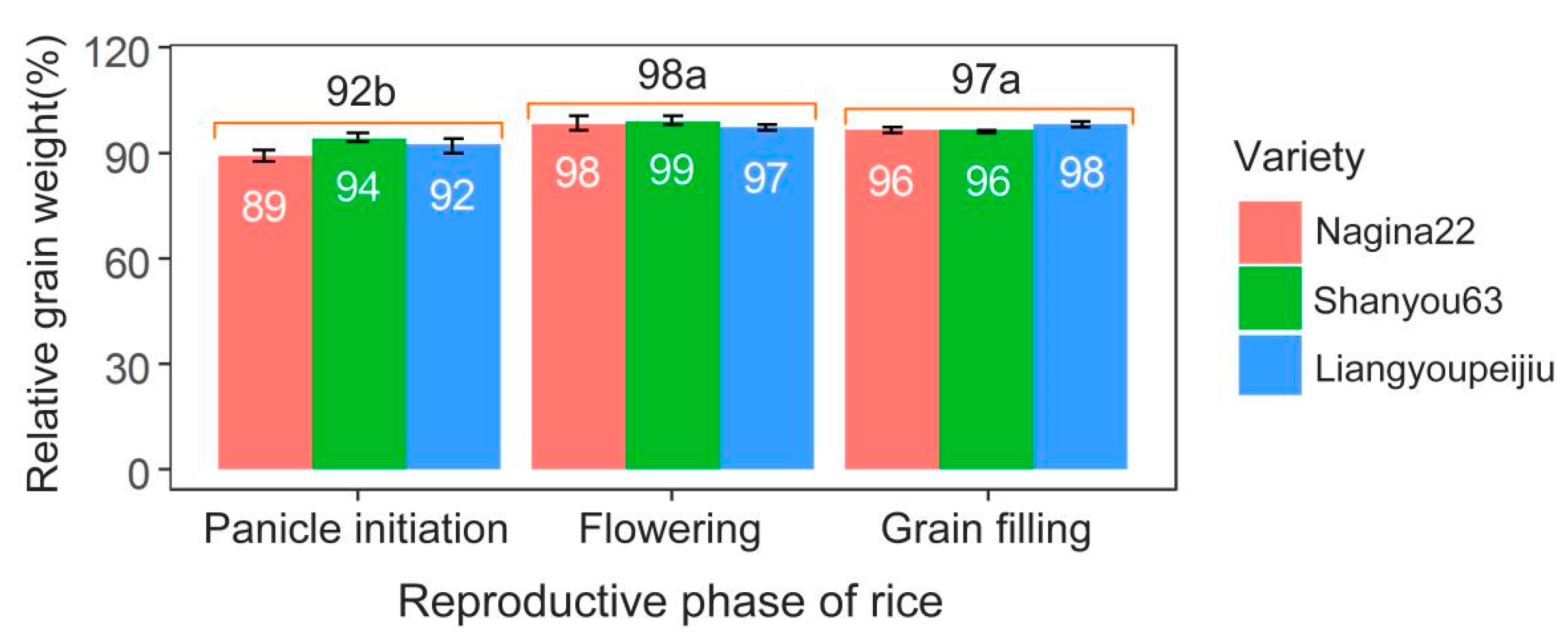
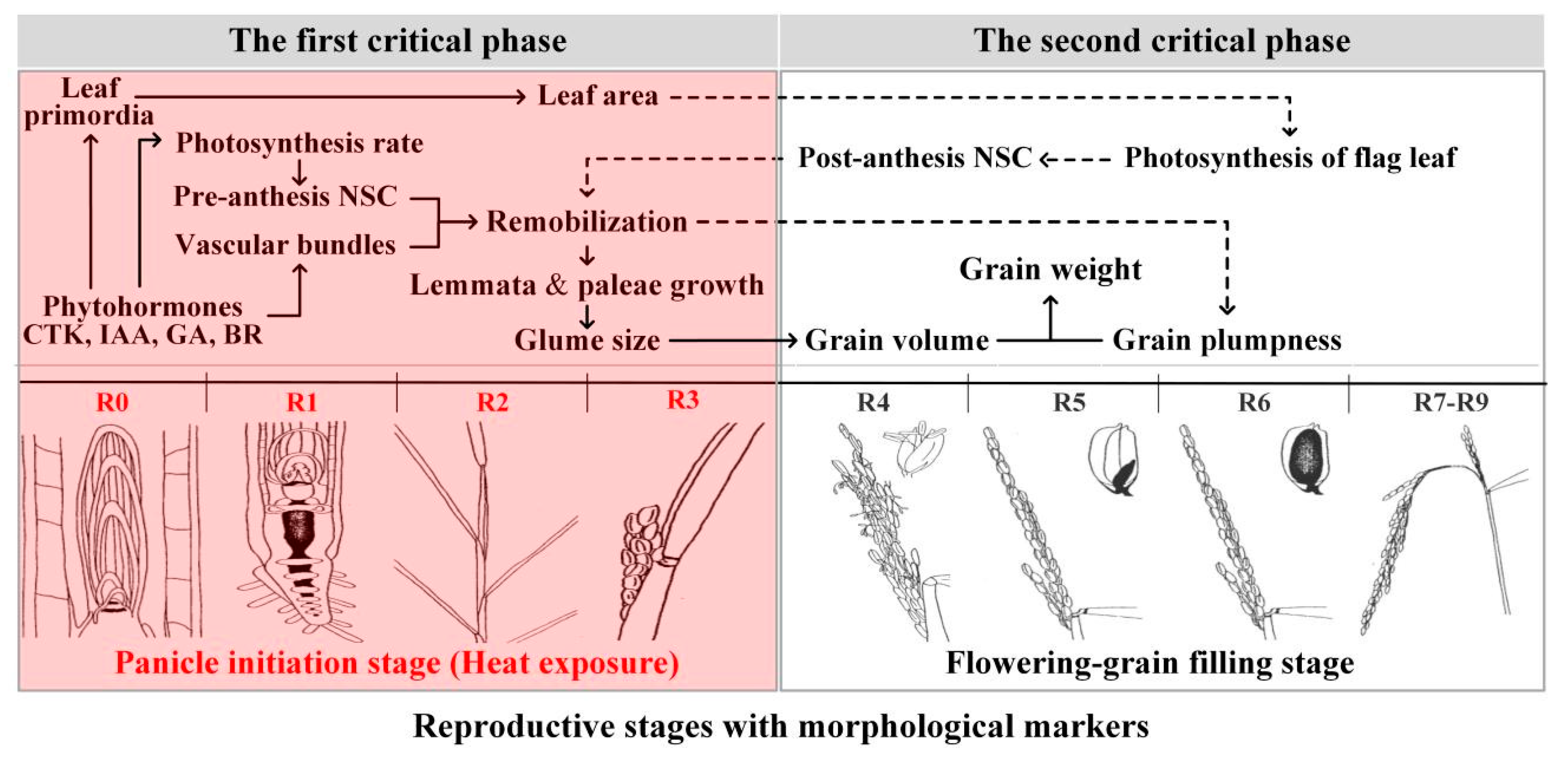
2. Heat Effects on Rice Grain Weight during the Panicle Initiation Stage
3. Heat-Induced Changes in Assimilate Supply Explain the Reduced Rice Grain Weight during the Panicle Initiation Stage
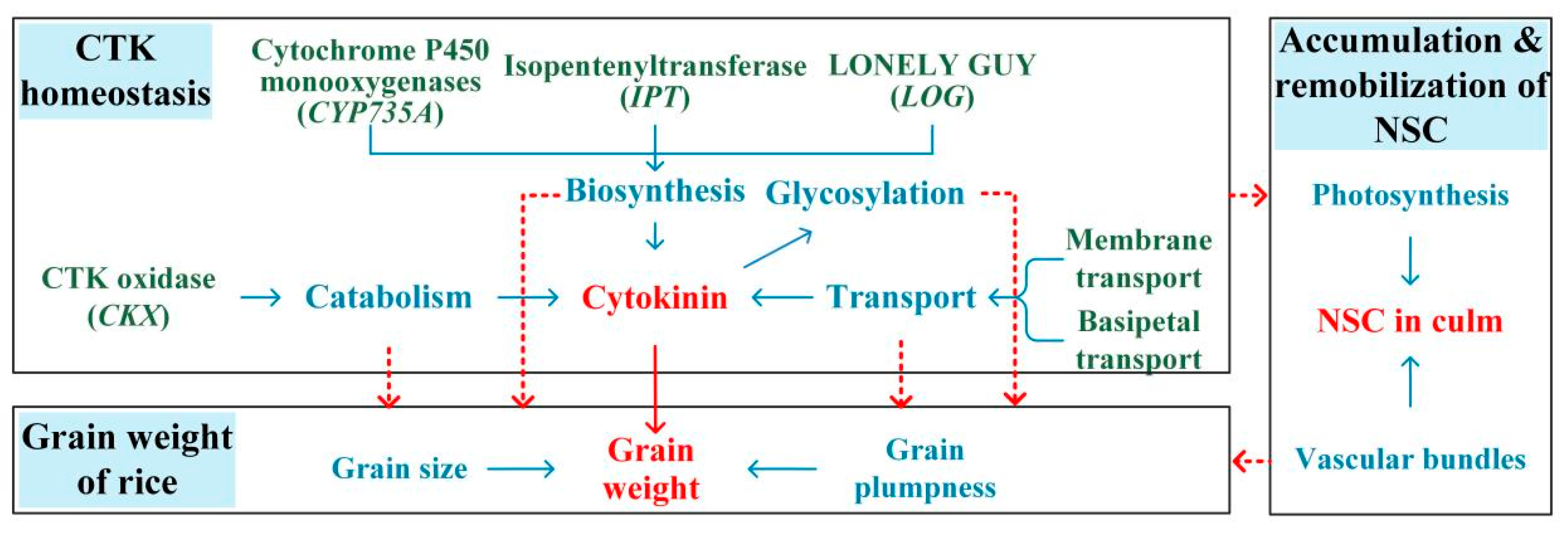
4. Effect of Phytohormones on Grain Weight under Heat during the Panicle Initiation Stage
5. Conclusions and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021; pp. 56–73. [Google Scholar]
- Wang, X.; Li, Y.; Wang, M.; Li, Y.; Gong, X.; Chen, Y.; Chen, Y.; Cao, W. Changes in daily extreme temperature and precipitation events in mainland China from 1960 to 2016 under global warming. Int. J. Climatol. 2021, 41, 1465–1483. [Google Scholar] [CrossRef]
- Dhorde, A.; Wakhare, A. Evidence of long-term climate change at major cities of India during the twentieth century. Int. J. Clim. Chang. Imp. Res. 2009, 1, 15–41. [Google Scholar] [CrossRef]
- Martínez, M.; Serra, C.; BurgueñO, A.; Lana, X. Time trends of daily maximum and minimum temperatures in Catalonia (ne Spain) for the period 1975–2004. Int. J. Climatol. 2010, 30, 267–290. [Google Scholar] [CrossRef]
- Tabari, H.; Talaee, P.H. Recent trends of mean maximum and minimum air temperatures in the western half of Iran. Meteorol. Atmos. Phys. 2011, 111, 121–131. [Google Scholar] [CrossRef]
- Wu, C.; Cui, K.; Li, Q.; Li, L.; Wang, W.; Hu, Q.; Ding, Y.; Li, G.; Fahad, S.; Huang, J.; et al. Estimating the yield stability of heat-tolerant rice genotypes under various heat conditions across reproductive stages: A 5-year case study. Sci. Rep. 2021, 11, 13604. [Google Scholar] [CrossRef]
- Fahad, S.; Ihsan, M.Z.; Khaliq, A.; Daur, I.; Saud, S.; Alzamanan, S.; Nasim, W.; Abdullah, M.; Khan, I.A.; Wu, C.; et al. Consequences of high temperature under changing climate optima for rice pollen characteristics-concepts and perspectives. Arch. Agron. Soil Sci. 2018, 64, 1473–1488. [Google Scholar] [CrossRef]
- Lin, M.; Wang, C.; Zhang, J. Heat injury risk assessment for single-cropping rice in the middle and lower reaches of the Yangtze river under climate change. J. Meteorol. Res. 2016, 30, 426–443. [Google Scholar]
- Wang, Y.; Li, J. The plant architecture of rice (Oryza sativa). Plant Mol. Biol. 2005, 59, 75–84. [Google Scholar] [CrossRef]
- Sánchez, B.; Rasmussen, A.; Porter, J.R. Temperatures and the growth and development of maize and rice: A review. Global Chang. Biol. 2014, 20, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Impa, S.M.; Raju, B.; Hein, N.T.; Sandhu, J.; Prasad, P.V.V.; Walia, H.; Jagadish, S.V.K. High night temperature effects on wheat and rice: Current status and way forward. Plant Cell Environ. 2021, 44, 2049–2065. [Google Scholar] [CrossRef] [PubMed]
- Jagadish, K.S.; Way, D.A.; Sharkey, T.D. Plant heat stress: Concepts directing future research. Plant Cell Environ. 2021, 44, 1992–2005. [Google Scholar] [CrossRef]
- Wu, C.; Cui, K.; Wang, W.; Li, Q.; Fahad, S.; Hu, Q.; Huang, J.; Nie, L.; Peng, S. Heat-induced phytohormone changes are associated with disrupted early reproductive development and reduced yield in rice. Sci. Rep. 2016, 6, 34978. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Chu, C.; Yao, S. The impact of high-temperature stress on rice: Challenges and solutions. Crop J. 2021, 9, 963–976. [Google Scholar] [CrossRef]
- Qi, B.; Wu, C. Potential roles of stigma exsertion on spikelet fertility in rice (Oryza sativa L.) under heat stress. Front. Plant Sci. 2022, 13, 983070. [Google Scholar]
- Ren, Y.; Huang, Z.; Jiang, H.; Wang, Z.; Wu, F.; Xiong, Y.; Yao, J. A heat stress responsive NAC transcription factor heterodimer plays key roles in rice grain filling. J. Exp. Bot. 2021, 72, 2947–2964. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Zhang, Q. Genetic and molecular bases of rice yield. Annu. Rev. Plant Biol. 2010, 61, 421–442. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Tang, S.; Li, G.; Wang, S.; Fahad, S.; Ding, Y. Roles of phytohormone changes in the grain yield of rice plants exposed to heat: A review. PeerJ 2019, 7, e7792. [Google Scholar] [CrossRef] [PubMed]
- Aghamolki, M.T.K.; Yusop, M.K.; Oad, F.C.; Zakikhani, H.; Jaafar, H.Z.; Kharidah, S.; Musa, M.H. Heat stress effects on yield parameters of selected rice cultivars at reproductive growth stages. J. Food Agric. Environ. 2014, 12, 741–746. [Google Scholar]
- Arai-Sanoh, Y.; Ishimaru, T.; Ohsumi, A.; Kondo, M. Effects of soil temperature on growth and root function in rice. Plant Prod. Sci. 2010, 13, 235–242. [Google Scholar] [CrossRef]
- Wang, Y.L.; Zhang, Y.P.; Zeng, Y.H.; Wu, H.; Xiang, J.; Chen, H.Z.; Zhang, Y.K.; Zhu, D.F. Effect of high temperature stress on rice spikelet differentiation and degeneration during panicle initiation stage. Chin. J. Agrometeorol. 2015, 36, 724–731. [Google Scholar]
- Luo, Z.Q.; Shi, C.L.; Jiang, M.; Liu, Y.; Xuan, S.L.; Jin, Z.Q. Effect of high temperature on rice dry matter partition and yield component during booting stage. Chin. J. Agrometeorol. 2016, 37, 326–334. [Google Scholar]
- Zhang, C.X.; Feng, B.H.; Chen, T.T.; Zhang, X.F.; Tao, L.X.; Fu, G.F. Sugars, antioxidant enzymes and IAA mediate salicylic acid to prevent rice spikelet degeneration caused by heat stress. Plant Growth Regul. 2017, 83, 313–323. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, J.; Chen, L.; Huang, X.; Cheng, Z.; Han, B.; Zhang, Q.; Wu, C. Rice functional genomics research: Past decade and future. Mol. Plant 2018, 11, 359–380. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Henry, A.; Sreenivasulu, N. Rice yield formation under high day and night temperatures—A prerequisite to ensure future food security. Plant Cell Environ. 2020, 43, 1595–1608. [Google Scholar] [CrossRef]
- Wu, C.; Song, Y.; Qi, B.; Fahad, S. Effects of asymmetric heat on grain quality during the panicle initiation stage in contrasting rice genotypes. J. Plant Growth Regul. 2022, 1–7. [Google Scholar] [CrossRef]
- Pan, J.; Cui, K.; Wei, D.; Huang, J.; Xiang, J.; Nie, L. Relationships of non-structural carbohydrates accumulation and translocation with yield formation in rice recombinant inbred lines under two nitrogen levels. Physiol. Plantarum 2010, 141, 321–331. [Google Scholar] [CrossRef]
- Hermann, R.-D.; Gabriel, G.-V. Response of rice plants to heat stress during initiation of panicle primordia or grain-filling phases. J. Stress Physiol. Biochem. 2013, 9, 318–325. [Google Scholar]
- Laza, M.R.C.; Sakai, H.; Cheng, W.; Tokida, T.; Peng, S.; Hasegawa, T. Differential response of rice plants to high night temperatures imposed at varying developmental phases. Agric. Forest Meteorol. 2015, 209–210, 69–77. [Google Scholar] [CrossRef]
- Zhen, F.; Zhou, J.; Mahmood, A.; Wang, W.; Chang, X.; Liu, B.; Liu, L.; Cao, W.; Zhu, Y.; Tang, L. Quantifying the effects of short-term heat stress at booting stage on nonstructural carbohydrates remobilization in rice. Crop J. 2020, 8, 194–212. [Google Scholar] [CrossRef]
- Takeoka, Y.; Hiroi, K.; Kitano, H.; Wada, T. Pistil hyperplasia in rice spikelets as affected by heat stress. Sexual Plant Reprod. 1991, 4, 39–43. [Google Scholar] [CrossRef]
- Zhang, G.L.; Chen, L.Y.; Zhang, S.T.; Zheng, H.; Liu, G.H. Effects of high temperature stress on microscopic and ultrastructural characteristics of mesophyll cells in flag leaves of rice. Rice Sci. 2009, 16, 65–71. [Google Scholar] [CrossRef]
- Hu, Q.; Wencheng, W.; Lu, Q.; Huang, J.; Peng, S.; Cui, K. Abnormal anther development leads to lower spikelet fertility in rice (Oryza sativa L.) under high temperature during the panicle initiation stage. BMC Plant Biol. 2021, 21, 428. [Google Scholar] [CrossRef] [PubMed]
- Counce, P.A.; Bryant, R.J.; Bergman, C.J.; Bautista, R.C.; Wang, Y.J.; Siebenmorgen, T.J.; Kak, M.; Jfc, M. Rice milling quality, grain dimensions, and starch branching as affected by high night temperatures. Cereal Chem 2005, 82, 645–648. [Google Scholar] [CrossRef]
- Morita, S.; Yonemaru, J.I.; Takanashi, J.I. Grain growth and endosperm cell size under high night temperatures in rice (Oryza sativa L.). Ann. Bot. 2005, 95, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Li, L.Y. Effects of High Temperature Treatments at Different Growth Stages on the Growth and Yield Formation of Rice. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2015. [Google Scholar]
- Kumar, A.; Nayak, A.K.; Pani, D.R.; Das, B.S. Physiological and morphological responses of four different rice cultivars to soil water potential based deficit irrigation management strategies. Field Crops Res. 2017, 205, 78–94. [Google Scholar] [CrossRef]
- Zhang, B.; Li, F.M.; Huang, G.; Cheng, Z.Y.; Zhang, Y. Yield performance of spring wheat improved by regulated deficit irrigation in an arid area. Agric. Water Manag. 2006, 79, 28–42. [Google Scholar] [CrossRef]
- Wu, C.; Cui, K.; Wang, W.; Li, Q.; Fahad, S.; Hu, Q.; Huang, J.; Nie, L.; Mohapatra, P.K.; Peng, S. Heat-induced cytokinin transportation and degradation are associated with reduced panicle cytokinin expression and fewer spikelets per panicle in rice. Front. Plant Sci. 2017, 8, 371. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.C. Mechanism and regulation in the filling of inferior spikelets of rice. Acta Agron. Sin. 2010, 36, 2011–2019. [Google Scholar]
- Yang, L.X.; Wang, Y.L.; Huang, J.Y.; Zhou, X.D.; Xu, J.K.; Gu, H.; Yang, J.H.; Dong, G.C. Dynamic change and influencing factors of hull size in Yangdao 6, a rice variety. Acta Agron. Sin. 2003, 29, 853–859. [Google Scholar]
- Castillo, F.M.; Vásquez, S.C.; Calderini, D.F. Does the pre-flowering period determine the potential grain weight of sunflower? Field Crops Res. 2017, 212, 23–33. [Google Scholar] [CrossRef]
- Ishimaru, K.; Hirotsu, N.; Madoka, Y.; Murakami, N.; Hara, N.; Onodera, H.; Kashiwagi, T.; Ujiie, K.; Shimizu, B.; Onishi, A. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield. Nat. Genet. 2013, 45, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhao, B.; Yuan, D.; Duan, M.; Li, C. Rice zinc finger protein DST enhances grain production through controlling Gn1a/OsCKX2 expression. Proc. Natl. Acad. Sci. USA 2013, 110, 3167–3172. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Shen, Y.; Zheng, M.; Yang, C.; Chen, Y.; Feng, Z.; Liu, X.; Liu, S.; Chen, Z.; Lei, C. Gene SGL, encoding a kinesin-like protein with transactivation activity, is involved in grain length and plant height in rice. Plant Cell Rep. 2014, 33, 235–244. [Google Scholar] [CrossRef]
- Itoh, J.I.; Hibara, K.I.; Sato, Y.; Nagato, Y. Developmental role and auxin responsiveness of class III homeodomain leucine zipper gene family members in rice. Plant Physiol. 2008, 147, 1960–1975. [Google Scholar] [CrossRef] [PubMed]
- Mangieri, M.A.; Hall, A.J.; Striker, G.G.; Chimenti, C.A. Cytokinins: A key player in determining differences in patterns of canopy senescence in Stay-Green and Fast Dry-Down sunflower (Helianthus annuus L.) hybrids. Eur. J. Agron. 2017, 86, 60–70. [Google Scholar] [CrossRef]
- Dawood, M.F.A.; Abu-Elsaoud, A.M.; Sofy, M.R.; Mohamed, H.I.; Soliman, M.H. Appraisal of kinetin spraying strategy to alleviate the harmful effects of UVC stress on tomato plants. Environ. Sci. Pollut. R. 2022, 29, 52378–52398. [Google Scholar] [CrossRef]
- Groover, A.T.; Pattishall, A.; Jones, A.M. IAA8 expression during vascular cell differentiation. Plant Mol. Biol. 2003, 51, 427–435. [Google Scholar] [CrossRef]
- Mähönen, A.P.; Bishopp, A.; Higuchi, M.; Nieminen, K.M.; Kinoshita, K.; Törmäkangas, K.; Ikeda, Y.; Oka, A.; Kakimoto, T.; Helariutta, Y. Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science 2006, 311, 94–98. [Google Scholar] [CrossRef]
- Joshi, R.; Sahoo, K.K.; Tripathi, A.K.; Kumar, R.; Gupta, B.K.; Pareek, A.; Singla-Pareek, S.L. Knockdown of an inflorescence meristem-specific cytokinin oxidase—OsCKX2 in rice reduces yield penalty under salinity stress condition. Plant Cell Environ. 2017, 41, 936–946. [Google Scholar] [CrossRef]
- Xiao, Y.; Liu, D.; Zhang, G.; Gao, S.; Liu, L.; Xu, F.; Che, R.; Wang, Y.; Tong, H.; Chu, C. Big Grain3, encoding a purine permease, regulates grain size via modulating cytokinin transport in rice. J. Integr. Plant Biol. 2019, 61, 581–597. [Google Scholar] [CrossRef]
- Nayar, S.; Sharma, R.; Tyagi, A.K.; Kapoor, S. Functional delineation of rice MADS29 reveals its role in embryo and endosperm development by affecting hormone homeostasis. J. Exp. Bot. 2013, 64, 4239–4253. [Google Scholar] [CrossRef] [PubMed]
- Che, R.; Tong, H.; Shi, B.; Liu, Y.; Fang, S.; Liu, D.; Xiao, Y.; Hu, B.; Liu, L.; Wang, H. Control of grain size and rice yield by GL2-mediated brassinosteroid responses. Nat. Plants 2015, 2, 15195. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Wu, C.; Wang, C.; Roh, J.; Zhang, L.; Chen, J.; Zhang, S.; Zhang, H.; Yang, C.; Hu, J. SLG controls grain size and leaf angle by modulating brassinosteroid homeostasis in rice. J. Exp. Bot. 2016, 67, 4241–4253. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, J.; Zheng, X.; Wu, F.; Lin, Q.; Heng, Y.; Tian, P.; Cheng, Z.; Yu, X.; Zhou, K. GW5 acts in the brassinosteroid signalling pathway to regulate grain width and weight in rice. Nat. Plants 2017, 3, 17043. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhang, M.; Wang, Y.; Liang, J. RGB1 regulates rice panicle architecture and grain filling through monitoring cytokinin level in inflorescence meristem and grain abscisic acid level during filling stage. Rice Sci. 2021, 28, 317–321. [Google Scholar]
- Guo, T.; Lu, Z.Q.; Shan, J.X.; Ye, W.W.; Lin, H.X. ERECTA1 acts upstream of the OsMKKK10-OsMKK4-OsMPK6 cascade to control spikelet number by regulating cytokinin metabolism in rice. Plant Cell 2020, 32, 2763–2779. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhang, J.; Yu, G.; Lu, X.; Tang, W. Endoplasmic reticulum-localized PURINE PERMEASE1 regulates plant height and grain weight by modulating cytokinin distribution in rice. Front. Plant Sci. 2020, 11, 618560. [Google Scholar] [CrossRef]
- Yin, W.; Xiao, Y.; Niu, M.; Meng, W.; Tong, H. ARGONAUTE2 enhances grain length and salt tolerance by activating BIG GRAIN3 to modulate cytokinin distribution in rice. Plant Cell 2020, 32, 2292–2306. [Google Scholar] [CrossRef]
- Tsago, Y.; Chen, Z.; Cao, H.; Sunusi, M.; Khan, A.U.; Shi, C.; Jin, X. Rice gene, OsCKX2-2, regulates inflorescence and grain size by increasing endogenous cytokinin content. Plant Growth Regul. 2020, 92, 283–294. [Google Scholar] [CrossRef]
- Liu, K.; Li, M.; Zhang, B.; Cui, Y.; Wang, M. Poaceae orthologs of rice OsSGL, DUF1645 domain-containing genes, positively regulate drought tolerance, grain length and weight in rice. Res. Sq. 2020, 29, 257–267. [Google Scholar] [CrossRef]
- Wang, M.; Lu, X.; Xu, G.; Yin, X.; Cui, Y.; Huang, L.; Rocha, P.S.C.F.; Xia, X. OsSGL, a novel pleiotropic stress-related gene enhances grain length and yield in rice. Sci. Rep. 2016, 6, 38157. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Y.; Mi, X.F.; Shan, J.X.; Lin, H.X. The QTL GNP1 encodes GA20ox1, which increases grain number and yield by increasing cytokinin activity in rice panicle meristems. PLoS Genet. 2016, 12, e1006386. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Xiong, L. Characterization of a purine permease family gene OsPUP7 involved in growth and development control in rice. J. Integr. Plant Biol. 2013, 55, 1119–1135. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.L.; Dong, N.Q.; Guo, T.; Ye, W.W.; Shan, J.X.; Lin, H.X. A quantitative trait locus GW6 controls rice grain size and yield through the gibberellin pathway. Plant J. 2020, 103, 1174–1188. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Cheng, Z.; Liu, L.; Wang, M.; You, X.; Wang, J.; Zhang, F.; Zhou, C.; Zhang, Z.; Zhang, H.; et al. Small Grain and Dwarf 2, encoding an HD-Zip II family transcription factor, regulates plant development by modulating gibberellin biosynthesis in rice. Plant Sci. 2019, 288, 110208. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shi, S.; Tao, Q.; Tao, Y.; Miao, J.; Peng, X.; Li, C.; Yang, Z.; Zhou, Y.; Liang, G. OsGASR9 positively regulates grain size and yield in rice (Oryza sativa). Plant Sci. 2019, 286, 17–27. [Google Scholar] [CrossRef]
- Tamiru, M.; Undan, J.R.; Takagi, H.; Abe, A.; Yoshida, K.; Undan, J.Q.; Natsume, S.; Uemura, A.; Saitoh, H.; Matsumura, H.; et al. A cytochrome P450, OsDSS1, is involved in growth and drought stress responses in rice (Oryza sativa L.). Plant Mol. Biol. 2015, 88, 85–99. [Google Scholar] [CrossRef]
- Ho, S.L.; Huang, L.F.; Lu, C.A.; He, S.L.; Wang, C.C.; Yu, S.P.; Chen, J.; Yu, S.M. Sugar starvation- and GA-inducible calcium-dependent protein kinase 1 feedback regulates GA biosynthesis and activates a 14-3-3 protein to confer drought tolerance in rice seedlings. Plant Mol. Biol. 2013, 81, 347–361. [Google Scholar] [CrossRef]
- Qiao, J.; Jiang, H.; Lin, Y.; Shang, L.; Qian, Q. A novel miR167a-OsARF6-OsAUX3 module regulates grain length and weight in rice. Mol. Plant 2021, 14, 1683–1698. [Google Scholar] [CrossRef]
- Dong, N.Q.; Sun, Y.; Guo, T.; Shi, C.L.; Lin, H.X. UDP-glucosyltransferase regulates grain size and abiotic stress tolerance associated with metabolic flux redirection in rice. Nat. Commun. 2020, 11, 2629. [Google Scholar] [CrossRef]
- Guo, T.; Chen, K.; Dong, N.-Q.; Ye, W.-W.; Shan, J.-X.; Lin, H.-X. TILLERING AND SMALL GRAIN 1 dominates the tryptophan aminotransferase family required for local auxin biosynthesis in rice. J. Integr. Plant Biol. 2020, 62, 581–600. [Google Scholar] [CrossRef]
- Hu, Z.; Lu, S.J.; Wang, M.J.; He, H.; Le, S.; Wang, H.; Liu, X.H.; Ling, J.; Sun, J.L.; Xin, X. A novel QTL qTGW3 encodes the GSK3/SHAGGY-like kinase OsGSK5/OsSK41 that interacts with OsARF4 to negatively regulate grain size and weight in rice. Mol. Plant 2018, 11, 736–749. [Google Scholar] [CrossRef]
- Liu, X.; Yang, C.Y.; Miao, R.; Zhou, C.L.; Cao, P.H.; Lan, J.; Zhu, X.J.; Mou, C.L.; Huang, Y.S.; Liu, S.J.; et al. DS1/OsEMF1 interacts with OsARF11 to control rice architecture by regulation of brassinosteroid signaling. Rice 2018, 11, 46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, J.; Tang, Z.; Sun, X.; Zhang, H.; Yu, J.; Yao, G.; Li, G.; Guo, H.; Li, J.; et al. Gnp4/LAX2, a RAWUL protein, interferes with the OsIAA3–OsARF25 interaction to regulate grain length via the auxin signaling pathway in rice. J. Exp. Bot. 2018, 69, 4723–4737. [Google Scholar] [CrossRef]
- Aya, K.; Hobo, T.; Sato-Izawa, K.; Ueguchi-Tanaka, M.; Kitano, H.; Matsuoka, M. A novel AP2-type transcription factor, SMALL ORGAN SIZE1, controls organ size downstream of an auxin signaling pathway. Plant Cell Physiol. 2014, 55, 897–912. [Google Scholar] [CrossRef] [PubMed]
- Hirano, K.; Yoshida, H.; Aya, K.; Kawamura, M.; Hayashi, M.; Hobo, T.; Sato-Izawa, K.; Kitano, H.; Ueguchi-Tanaka, M.; Matsuoka, M. SMALL ORGAN SIZE 1 and SMALL ORGAN SIZE 2/DWARF and LOW-TILLERING form a complex to integrate auxin and brassinosteroid signaling in rice. Mol. Plant 2017, 10, 590–604. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Tong, H.; Xiao, Y.; Che, R.; Xu, F.; Hu, B.; Liang, C.; Chu, J.; Li, J.; Chu, C. Activation of Big Grain1 significantly improves grain size by regulating auxin transport in rice. Proc. Natl. Acad. Sci. USA 2015, 112, 11102. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Fang, J.; Ou, S.; Gao, S.; Zhang, F.; Du, L.; Xiao, Y.; Wang, H.; Sun, X.; Chu, J.; et al. Variations in CYP78A13 coding region influence grain size and yield in rice. Plant Cell Environ. 2015, 38, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-H.; Yoo, S.-C.; Zhang, H.; Pandeya, D.; Koh, H.-J.; Hwang, J.-Y.; Kim, G.-T.; Paek, N.-C. The rice narrow leaf2 and narrow leaf3 loci encode WUSCHEL-related homeobox 3A (OsWOX3A) and function in leaf, spikelet, tiller and lateral root development. New Phytol. 2013, 198, 1071–1084. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fan, X.; Song, W.; Zhang, Y.; Xu, G. Over-expression of OsPIN2 leads to increased tiller numbers, angle and shorter plant height through suppression of OsLAZY1. Plant Biotechnol. J. 2012, 10, 139–149. [Google Scholar] [CrossRef]
- Li, Y.; Huang, R.; Li, J.; Huang, X.; Zeng, X.; Zhao, D. DWARF and SMALL SEED1, a novel allele of OsDWARF, controls rice plant architecture, seed size, and chlorophyll biosynthesis. Phyton 2021, 90, 111–127. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, R.; Xing, Y.; Xu, Y.; Xiong, D.; Wang, Y.; Yao, S. Separable regulation of POW1 in grain size and leaf angle development in rice. Plant Biotechnol. J. 2021, 19, 2517–2531. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Xu, H.; Li, B.; Shang, Y.; Wei, M.; Zhang, S.; Zhao, C.; Qin, R.; Cui, F.; Wu, Y. The brassinosteroid biosynthesis gene, ZmD11, increases seed size and quality in rice and maize. Plant Physiol. Biochem. 2021, 160, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Morinaka, Y.; Sakamoto, T.; Inukai, Y.; Agetsuma, M.; Kitano, H.; Ashikari, M.; Matsuoka, M. Morphological alteration caused by brassinosteroid insensitivity increases the biomass and grain production of rice. Plant Physiol. 2006, 141, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Zhu, W.; Tong, Y. Knock-down the expression of brassinosteroid receptor TaBRI1 reduces photosynthesis, tolerance to high light and high temperature stresses and grain yield in wheat. Plants 2020, 9, 840. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Yan, J.; Su, J.; Liu, H.; Hu, C.; Li, G.; Wang, F.; Lin, Y. Novel OsGRAS19 mutant, D26, positively regulates grain shape in rice (Oryza sativa). Funct. Plant Biol. 2019, 46, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Qin, P.; Peng, Y.; Ma, B.; Hu, J.; Fan, S.; Hu, B.; Zhang, G.; Yuan, H.; Yan, W. A single nucleotide substitution at 5’-UTR of GSN1 represses its translation and leads to an increase of grain length in rice. J. Genet. Genom. 2019, 46, 105–108. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiong, Y.; Liu, R.; Xue, H.W.; Yang, Z. The Rho-family GTPase OsRac1 controls rice grain size and yield by regulating cell division. Proc. Natl. Acad. Sci. USA 2019, 116, 16121–16126. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; Zeng, D.; Yang, C.; Akhter, D.; Alamin, M.; Jin, X.; Shi, C. LTBSG1, a new allele of BRD2, regulates panicle and grain development in rice by brassinosteroid biosynthetic pathway. Genes 2018, 9, 292. [Google Scholar] [CrossRef] [PubMed]
- Segami, S.; Takehara, K.; Yamamoto, T.; Kido, S.; Kondo, S.; Iwasaki, Y.; Miura, K. Overexpression of SRS5 improves grain size of brassinosteroid-related dwarf mutants in rice (Oryza sativa L.). Breed. Sci. 2017, 67, 393–397. [Google Scholar] [CrossRef]
- Wang, N.; Xing, Y.; Lou, Q.; Feng, P.; Liu, S.; Zhu, M.; Yin, W.; Fang, S.; Lin, Y.; Zhang, T.; et al. Dwarf and short grain 1, encoding a putative U-box protein regulates cell division and elongation in rice. J. Plant Physiol. 2017, 209, 84–94. [Google Scholar] [CrossRef]
- Xiao, Y.; Liu, D.; Zhang, G.; Tong, H.; Chu, C. Brassinosteroids regulate OFP1, a DLT interacting protein, to modulate plant architecture and grain morphology in rice. Front. Plant Sci. 2017, 8, 1698. [Google Scholar] [CrossRef]
- Yu, H.; Banpu, R.; Wang, Z.; Ren, D.; Zhang, Y.; Leng, Y.; Zeng, D.; Hu, J.; Zhang, G.; Zhu, L. Fine mapping of a novel defective glume 1 (dg1) mutant, which affects vegetative and spikelet development in rice. Front. Plant Sci. 2017, 8, 486. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Wu, C.; Fu, Y.; Hu, G.; He, Z.; Liu, W. Novel rice mutants overexpressing the brassinosteroid catabolic gene CYP734A4. Plant Mol. Biol. 2017, 93, 197–208. [Google Scholar] [CrossRef]
- Fang, N.; Xu, R.; Huang, L.; Zhang, B.; Duan, P.; Li, N.; Luo, Y.; Li, Y. SMALL GRAIN 11 controls grain size, grain number and grain yield in rice. Rice 2016, 9, 64. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Feng, Z.M.; Zhou, C.L.; Ren, Y.K.; Mou, C.L.; Wu, T.; Yang, C.Y.; Liu, S.J.; Jiang, L.; Wan, J.M. Brassinosteroid (BR) biosynthetic gene lhdd10 controls late heading and plant height in rice (Oryza sativa L.). Plant Cell Rep. 2016, 35, 357–368. [Google Scholar] [CrossRef]
- Wu, Y.; Fu, Y.; Zhao, S.; Gu, P.; Zhu, Z.; Sun, C.; Tan, L. CLUSTERED PRIMARY BRANCH 1, a new allele of DWARF11, controls panicle architecture and seed size in rice. Plant Biotechnol. J. 2016, 14, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hua, L.; Dong, S.; Chen, H.; Zhu, X.; Jiang, J.E.; Zhang, F.; Li, Y.; Fang, X.; Chen, F. OsMAPK6, a mitogen-activated protein kinase, influences rice grain size and biomass production. Plant J. 2015, 84, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Rao, Y.; Xu, J.; Hu, S.; Fang, Y.; Yu, H.; Pan, J.; Liu, R.; Ren, D.; Wang, X.; et al. Characterization and cloning of SMALL GRAIN 4, a novel DWARF11 allele that affects brassinosteroid biosynthesis in rice. Sci. Bull. 2015, 60, 905–915. [Google Scholar] [CrossRef]
- Xu, C.J.; Liu, Y.; Li, Y.B.; Xu, X.D.; Xu, C.G.; Li, X.H.; Xiao, J.H.; Zhang, Q.F. Differential expression of GS5 regulates grain size in rice. J. Exp. Bot. 2015, 66, 2611–2623. [Google Scholar] [CrossRef] [PubMed]
- Duan, P.; Rao, Y.; Zeng, D.; Yang, Y.; Xu, R.; Zhang, B.; Dong, G.; Qian, Q.; Li, Y. SMALL GRAIN 1, which encodes a mitogen-activated protein kinase kinase 4, influences grain size in rice. Plant J. 2014, 77, 547–557. [Google Scholar] [CrossRef]
- Kitagawa, K.; Kurinami, S.; Oki, K.; Abe, Y.; Ando, T.; Kono, I.; Yano, M.; Kitano, H.; Iwasaki, Y. A novel kinesin 13 protein regulating rice seed length. Plant Cell Physiol. 2010, 51, 1315–1329. [Google Scholar] [CrossRef]
- Xu, F.X.; Xiong, H.; Zhang, L.; Zhu, Y.; Liu, M.; Jiang, P.; Guo, X.; Zhou, X. Research progresses and prospects of sink-source structures and optimal regulation of high-yield varieties of hybrid rice in China. Chin. J. Eco-Agric. 2016, 24, 1285–1299. [Google Scholar]
- Zhen, F.; Liu, Y.; Ali, I.; Liu, B.; Zhu, Y. Short-term heat stress at booting stage inhibited nitrogen remobilization to grain in rice. J. Agric. Food Res. 2020, 2, 100066. [Google Scholar] [CrossRef]
- Liu, Q.; Ma, H.; Sun, Z.; Lin, X.; Zhou, X. Translocation efficiencies and allocation of nitrogen, phosphorous and potassium in rice as affected by silicon fertilizer under high daytime temperature. J. Agron. Crop Sci. 2019, 205, 188–201. [Google Scholar] [CrossRef]
- Yang, J.; Chen, X.R.; Zhu, C.L.; Peng, X.S.; He, X.P.; Fu, J.R.; Ouyang, L.J.; Bian, J.M.; Hu, L.F.; He, H.H. Effects of nitrogen level and high temperature at late booting stage on yield and physiological characteristics of two early rice cultivars. Chin. J. Rice Sci. 2014, 28, 523–533. [Google Scholar]
- Liu, Q.; Sun, Z.; Xin, C.; Ma, J.Q. Effects of silicon on dry matter remobilization, distribution and grain yield of rice under high air temperature. J. Nucl. Agric. Sci. 2016, 30, 1833–1839. [Google Scholar]
- Gunawardena, T.; Fukai, S. The interaction of nitrogen application and temperature during reproductive stage on spikelet sterility in field-grown rice. Aust. J. Agric. Res. 2005, 56, 625–636. [Google Scholar] [CrossRef]
- Ding, C.Q.; You, J.; Chen, L.; Wang, S.H.; Ding, Y.F. Nitrogen fertilizer increases spikelet number per panicle by enhancing cytokinin synthesis in rice. Plant Cell Rep. 2014, 33, 363–371. [Google Scholar] [CrossRef]
- Markovich, O.; Steiner, E.; Kouřil, Š.; Tarkowski, P.; Aharoni, A.; Elbaum, R. Silicon promotes cytokinin biosynthesis and delays senescence in Arabidopsis and Sorghum. Plant Cell Environ. 2017, 40, 1189–1196. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genes | Regulated Trait of Grain Size | Regulated Phytohormones | Verified Stresses | References |
|---|---|---|---|---|
| RGB1 | Grain length | CTK | — | [57] |
| OsER1 | Grain length | CTK | — | [58] |
| OsPUP1 | Grain length, grain width | CTK | — | [59] |
| AGO2 | Grain length | CTK | — | [60] |
| OsCKX2 | Grain length, grain width | CTK | Salinity, drought | [51,61] |
| OsSGL | Grain length | CTK | — | [62,63] |
| OsPUP4 | Grain width, grain thickness, grain length | CTK | — | [52] |
| GNP1/GA20ox1 | Grain length | CTK, GA | — | [64] |
| OsMADS29 | Grain length | CTK | — | [53] |
| OsPUP7 | Grain length, grain width | CTK | — | [65] |
| DST | Grain length, grain width | CTK | Salinity, drought | [44] |
| GW6 | Grain length, grain width | GA | — | [66] |
| SGD2 | Grain length, grain width | GA | — | [67] |
| OsGASR9 | Grain length, grain width, grain thickness | GA | — | [68] |
| DSS1/CYP96B | Grain length, grain width | GA, ABA | Drought | [69] |
| SGL | Grain length, grain width | GA | — | [45] |
| OsCDPK1 | Grain length, grain width | GA | Drought | [70] |
| OsARF6/OsAUX3 | Grain length | IAA | — | [71] |
| GSA1 | Grain length, grain width | IAA | Heat, salinity, drought | [72] |
| TSG1/FIB | Grain length, grain width | IAA | — | [73] |
| OsSK41/OsGSK5 | Grain length, grain width | IAA | — | [74] |
| DS1/OsEMF1 | Grain length, grain width | IAA, BR | — | [75] |
| Gnp4/LAX2 | Grain length | IAA | — | [76] |
| SMOS1/DLT | Grain length | IAA, BR | — | [77,78] |
| BG1 | Grain length, grain width | IAA | — | [79] |
| CYP78A13 | Grain length, grain width, grain thickness | IAA | — | [80] |
| TGW6 | Grain length | IAA | — | [43] |
| NAL2/3 | Grain width | IAA | — | [81] |
| OsPIN2 | Grain length, grain width | IAA | — | [82] |
| DSS1/OsDWARF | Grain length, grain width | BR | — | [83] |
| POW1 | Grain width, grain thickness, grain length | BR | — | [84] |
| ZmD11 | Grain length | BR | — | [85] |
| OsBRI1 | Grain length, grain width | BR | Heat, high light | [86,87] |
| OsGRAS19/D26 | Grain length, grain width | BR | — | [88] |
| GSN1 | Grain length | BR | — | [89] |
| OsRac1 | Grain length, grain width | BR | — | [90] |
| LTBSG1/BRD2 | Grain length, grain width | BR | — | [91] |
| SRS5 | Grain length | BR | — | [92] |
| GW5 | Grain width | BR | — | [56] |
| DSG1 | Grain length, grain width | BR | — | [93] |
| OFP1 | Grain width | BR | — | [94] |
| DG1 | Grain length, grain width | BR | — | [95] |
| CYP734A4 | Grain length | BR | — | [96] |
| SMG11/D2 | Grain length, grain width | BR | — | [97] |
| SLG | Grain length, grain width | BR | — | [55] |
| LHDD10/BRD2 | Grain length, grain width | BR | — | [98] |
| CPB1/D11 | Grain length, grain width | BR | — | [99] |
| GL2 | Grain length, grain width | BR | — | [54] |
| OsMAPK6 | Grain length | BR | — | [100] |
| sg4/D11 | Grain length | BR | — | [101] |
| GS5 | Grain width | BR | — | [102] |
| OsMKK4/SMG1 | Grain length, grain width | BR | — | [103] |
| SRS3 | Grain length, grain width | BR | — | [104] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.; Cui, K.; Fahad, S. Heat Stress Decreases Rice Grain Weight: Evidence and Physiological Mechanisms of Heat Effects Prior to Flowering. Int. J. Mol. Sci. 2022, 23, 10922. https://doi.org/10.3390/ijms231810922
Wu C, Cui K, Fahad S. Heat Stress Decreases Rice Grain Weight: Evidence and Physiological Mechanisms of Heat Effects Prior to Flowering. International Journal of Molecular Sciences. 2022; 23(18):10922. https://doi.org/10.3390/ijms231810922
Chicago/Turabian StyleWu, Chao, Kehui Cui, and Shah Fahad. 2022. "Heat Stress Decreases Rice Grain Weight: Evidence and Physiological Mechanisms of Heat Effects Prior to Flowering" International Journal of Molecular Sciences 23, no. 18: 10922. https://doi.org/10.3390/ijms231810922
APA StyleWu, C., Cui, K., & Fahad, S. (2022). Heat Stress Decreases Rice Grain Weight: Evidence and Physiological Mechanisms of Heat Effects Prior to Flowering. International Journal of Molecular Sciences, 23(18), 10922. https://doi.org/10.3390/ijms231810922