

Regulation of ssb Gene Expression in Escherichia coli

 ,
,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
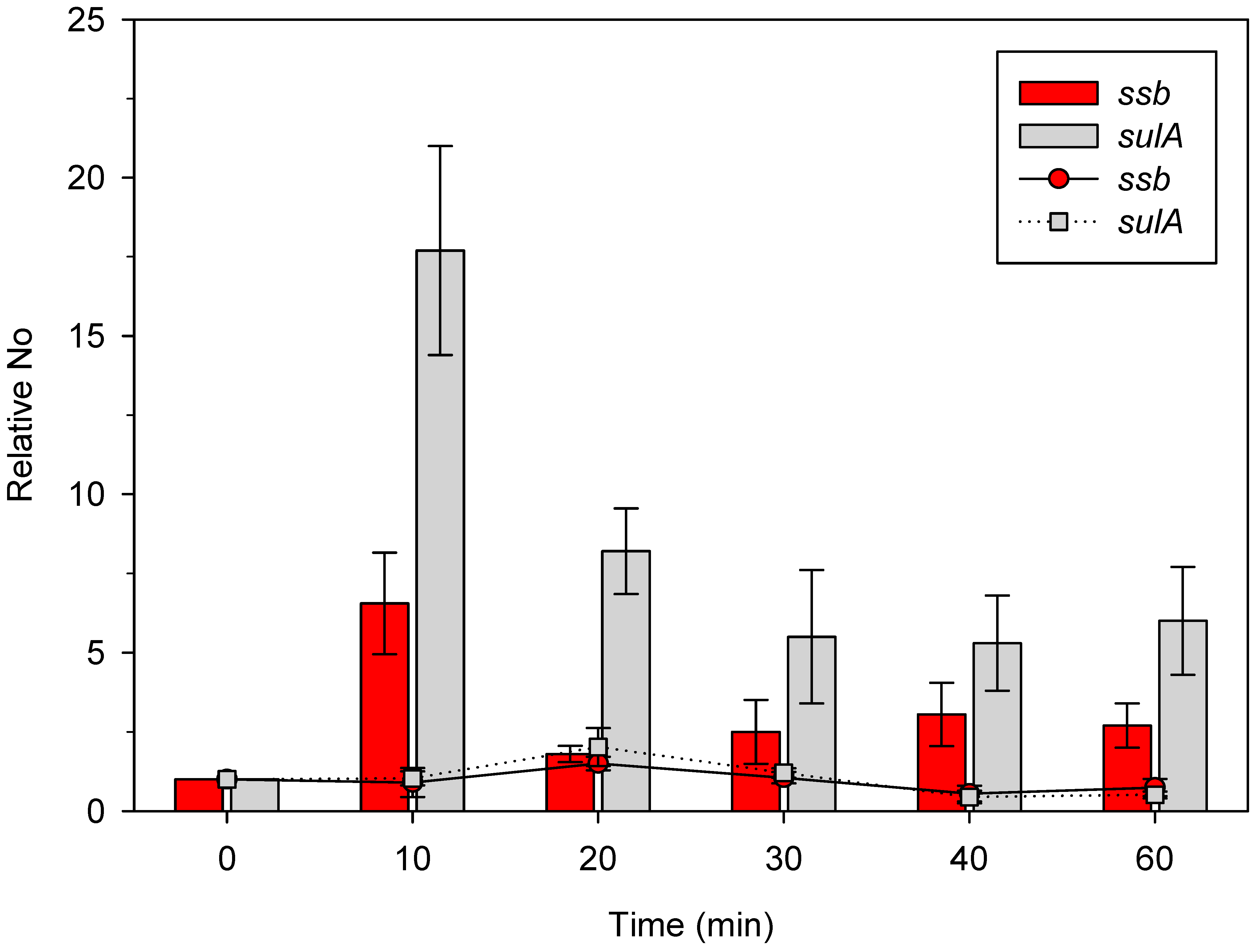
2.1. γ Irradiation Induces ssb and sulA Gene Expression
2.2. UV Induces ssb and sulA Gene Expression
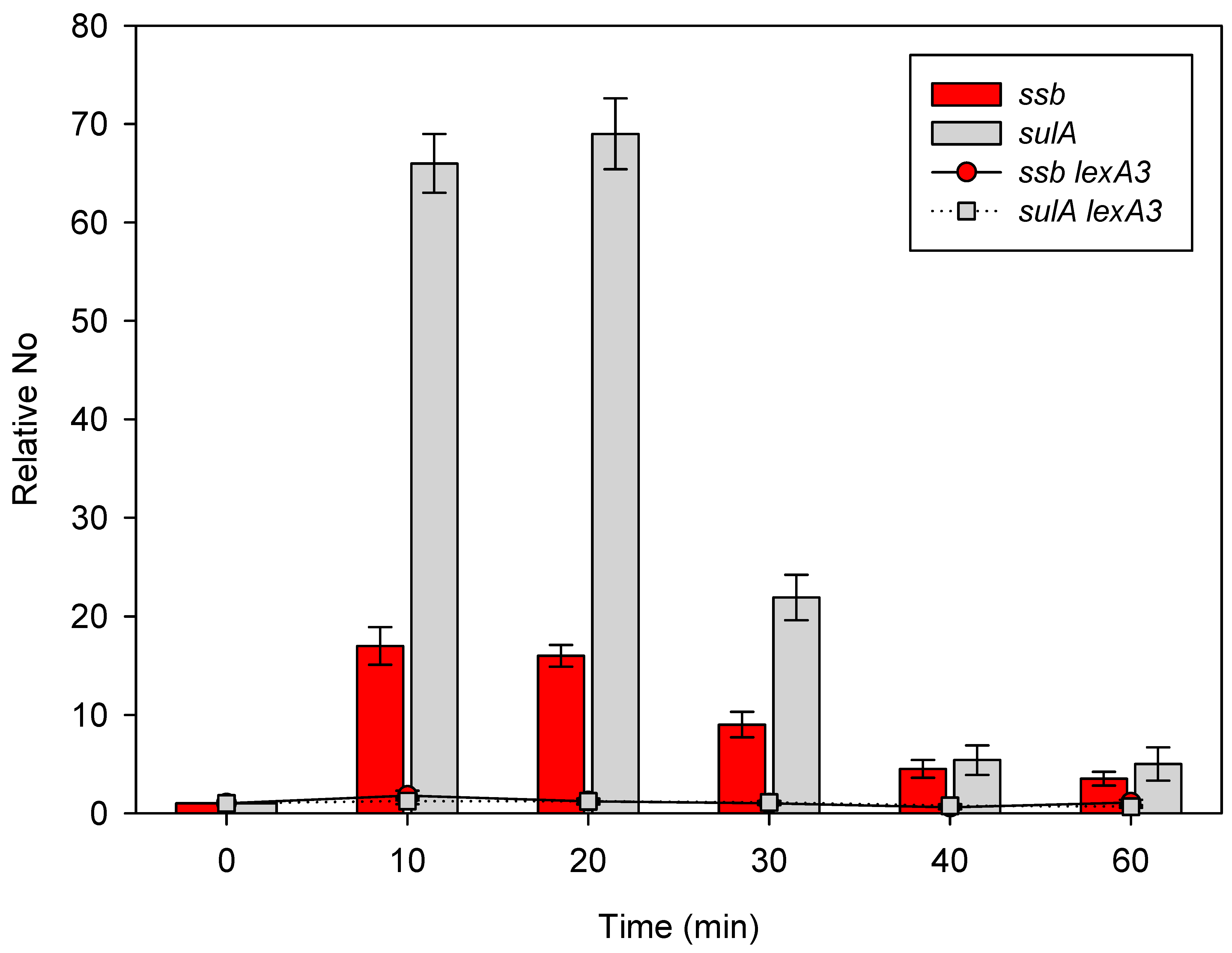
2.3. Expression of the ssb Gene Is Regulated by the LexA Repressor
2.4. The ssb Gene Is Overexpressed in Bacteria with Constitutively Activated SOS Regulon
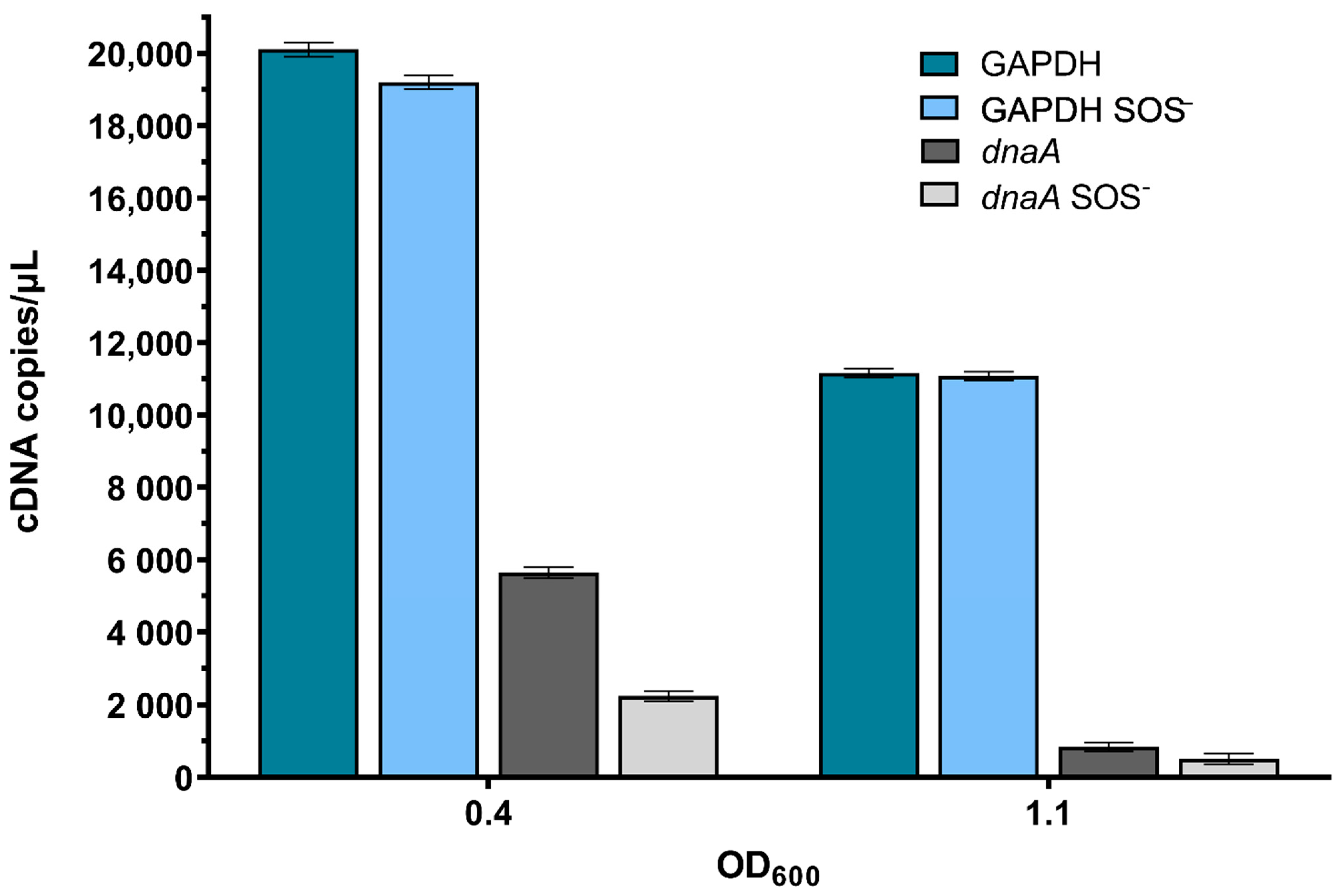
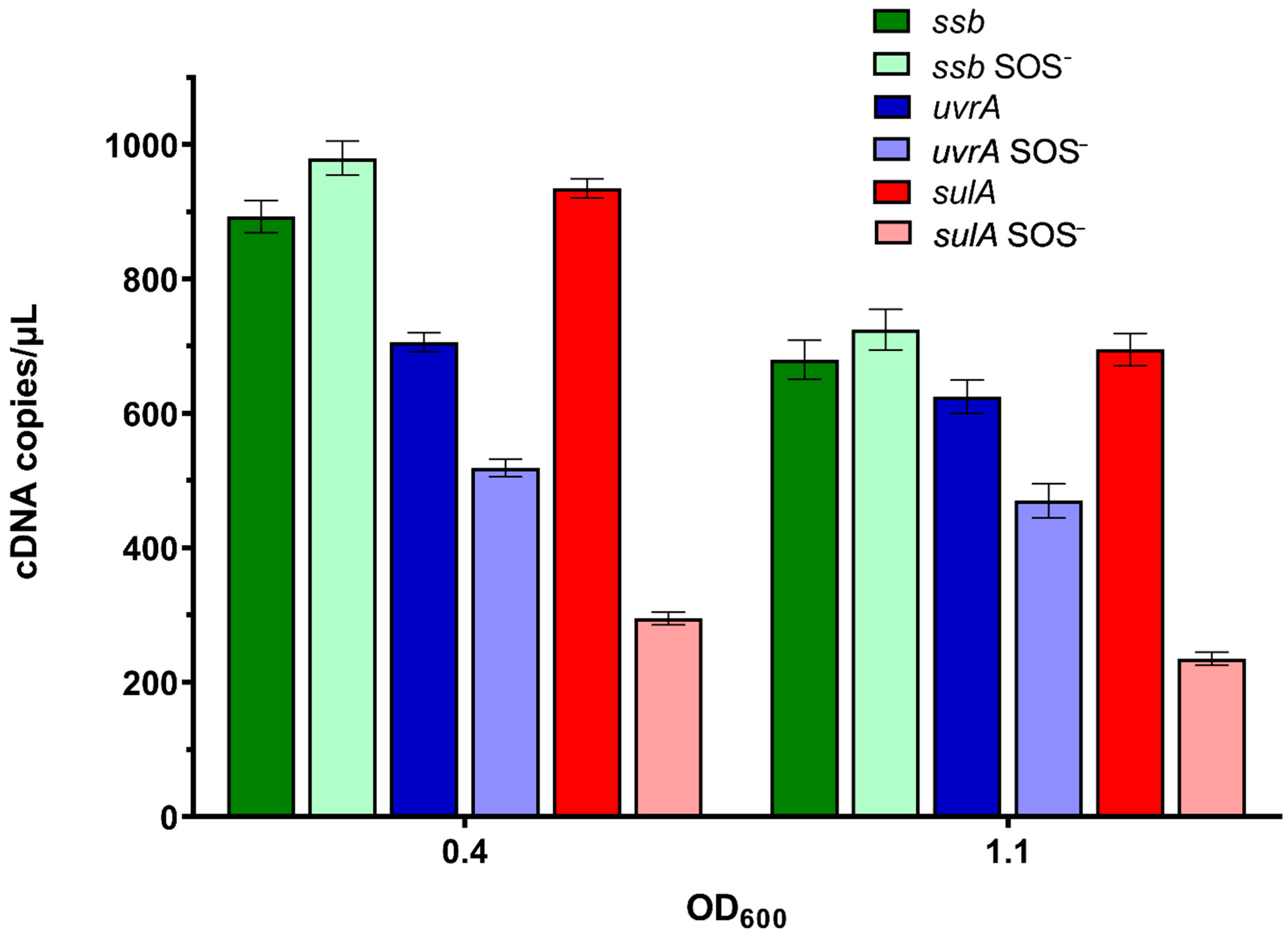
2.5. ssb Gene Expression Is Unlinked to SOS Induction during Undisturbed Bacterial Growth
3. Discussion
4. Materials and Methods
4.1. Strains, Growth Conditions, and Media
4.2. Cell Irradiation by UV and γ-Rays
4.3. RNA Isolation and Reverse Transcription
4.4. Quantitative Real-Time PCR (RT-qPCR) Analysis
4.5. Absolute Quantification of RNA Levels by Microfluidic Digital PCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sigal, N.; Delius, H.; Kornberg, T.; Gefter, M.L.; Alberts, B.A. DNA-unwinding protein isolated from Escherichia coli: Its interaction with DNA and with DNA polymerases. Proc. Natl. Acad. Sci. USA 1972, 69, 3537–3541. [Google Scholar] [CrossRef] [PubMed]
- Molineux, I.J.; Gefter, M.L. Properties of the Escherichia coli DNA-binding (unwinding) protein interaction with nucleolytic enzymes and DNA. J. Mol. Biol. 1975, 98, 811–825. [Google Scholar] [CrossRef]
- Weiner, J.H.; Bertsch, L.L.; Kornberg, A. The deoxyribonucleic acid unwinding protein of Escherichia coli. Properties and functions in replication. J. Biol. Chem. 1975, 250, 1972–1980. [Google Scholar] [CrossRef]
- Meyer, R.R.; Laine, P.S. The single-stranded DNA-binding protein of Escherichia coli. Microbiol. Rev. 1990, 54, 342–380. [Google Scholar] [CrossRef] [PubMed]
- Lohman, T.M.; Ferrari, M.E. Escherichia coli single-stranded DNA-binding protein: Multiple DNA-binding modes and cooperativities. Annu. Rev. Biochem. 1994, 63, 527–570. [Google Scholar] [CrossRef]
- Zou, Y.; Liu, Y.; Wu, X.; Shell, S.M. Functions of human replication protein A (RPA): From DNA replication to DNA damage and stress responses. J. Cell. Physiol. 2006, 208, 267–273. [Google Scholar] [CrossRef]
- Mackay, V.; Linn, S. Selective inhibition of the dnase activity of the recBC enzyme by the DNA binding protein from Escherichia coli. J. Biol. Chem. 1976, 251, 3716–3719. [Google Scholar] [CrossRef]
- Meyer, R.R.; Glassberg, J.; Scott, J.V.; Kornberg, A. A temperature-sensitive single-stranded DNA-binding protein from Escherichia coli. J. Biol. Chem. 1980, 255, 2897–2929. [Google Scholar] [CrossRef]
- Kowalczykowski, S.C.; Dixon, D.A.; Eggleston, A.K.; Lauder, S.D.; Rehrauer, W.M. Biochemistry of homologous recombination in Escherichia coli. Microbiol. Rev. 1994, 58, 401–465. [Google Scholar] [CrossRef]
- Myler, L.R.; Gallardo, I.F.; Zhou, Y.; Gong, F.; Yang, S.H.; Wold, M.S.; Miller, K.M.; Paull, T.T.; Finkelstein, I.J. Single-molecule imaging reveals the mechanism of Exo1 regulation by single-stranded DNA binding proteins. Proc. Natl. Acad. Sci. USA 2016, 113, E1170–E1179. [Google Scholar] [CrossRef]
- Shereda, R.D.; Kozlov, A.G.; Lohman, T.M.; Cox, M.M.; Keck, J.L. SSB as an organizer/mobilizer of genome maintenance complexes. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 289–318. [Google Scholar] [CrossRef]
- von Hippel, P.H.; Delagoutte, E. A general model for nucleic acid helicases and their “coupling” within macromolecular machines. Cell 2001, 104, 177–190. [Google Scholar] [CrossRef]
- Williams, K.R.; Spicer, E.K.; LoPresti, M.B.; Guggenheimer, R.A.; Chase, J.W. Limited proteolysis studies on the Escherichia coli single-stranded DNA binding protein. Evidence for a functionally homologous domain in both the Escherichia coli and T4 DNA binding proteins. J. Biol. Chem. 1983, 258, 3346–3355. [Google Scholar] [CrossRef]
- Raghunathan, S.; Kozlov, A.G.; Lohman, T.M.; Waksman, G. Structure of the DNA binding domain of E. coli SSB bound to ssDNA. Nat. Struct. Biol. 2000, 7, 648–652. [Google Scholar] [CrossRef] [PubMed]
- Costes, A.; Lecointe, F.; McGovern, S.; Quevillon-Cheruel, S.; Polard, P. The C-terminal domain of the bacterial SSB protein acts as a DNA maintenance hub at active chromosome replication forks. PLoS Genet. 2010, 6, e1001238. [Google Scholar] [CrossRef]
- BioCyc. Available online: https://biocyc.org/gene?orgid=ECOLI&id=EG10976 (accessed on 19 June 2022).
- Brandsma, J.A.; Bosch, D.; de Ruÿter, M.; van de Putte, P. Analysis of the regulatory region of the ssb gene of Escherichia coli. Nucleic Acids Res. 1985, 13, 5095–5109. [Google Scholar] [CrossRef]
- Friedberg, E.C.; Walker, G.C.; Siede, W.; Wood, R.D.; Schultz, R.A.; Ellenberger, T. DNA Repair and Mutagenesis, 2nd ed.; ASM Press: Washington, DC, USA, 2006. [Google Scholar]
- Fernández De Henestrosa, A.R.; Ogi, T.; Aoyagi, S.; Chafin, D.; Hayes, J.J.; Ohmori, H.; Woodgate, R. Identification of additional genes belonging to the LexA regulon in Escherichia coli. Mol. Microbiol. 2000, 35, 1560–1572. [Google Scholar] [CrossRef]
- Whittier, R.F.; Chase, J.W. DNA repair in E. coli strains deficient in single-strand DNA binding protein. Mol. Gen. Genet. 1981, 183, 341–347. [Google Scholar] [CrossRef]
- Salles, B.; Paoletti, C.; Villani, G. Lack of single-strand DNA-binding protein amplification under conditions of SOS induction in E. coli. Mol. Gen. Genet. 1983, 189, 175–177. [Google Scholar] [CrossRef]
- Villani, G.; Pierre, A.; Salles, B. Quantification of SSB protein in E. coli and its variation during RECA protein induction. Biochimie 1984, 66, 471–476. [Google Scholar] [CrossRef]
- Perrino, F.W.; Rein, D.C.; Bobst, A.M.; Meyer, R.R. The relative rate of synthesis and levels of single-stranded DNA binding protein during induction of SOS repair in Escherichia coli. Mol. Gen. Genet. 1987, 209, 612–614. [Google Scholar] [CrossRef]
- Moreau, P.L. Effects of overproduction of single-stranded DNA-binding protein on RecA protein-dependent processes in Escherichia coli. J. Mol. Biol. 1987, 194, 621–634. [Google Scholar] [CrossRef]
- Alazard, R.J. Study of the expression of UvrA and SSB proteins in vivo in lambda hybrid phages containing the uvrA and ssbA genes of Escherichia coli. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 1983, 109, 155–168. [Google Scholar] [CrossRef]
- Brandsma, J.A.; Bosch, D.; Backendorf, C.; van de Putte, P. A common regulatory region shared by divergently transcribed genes of the Escherichia coli SOS system. Nature 1983, 305, 243–245. [Google Scholar] [CrossRef] [PubMed]
- Bertrand-Burggraf, E.; Oertel, P.; Schnarr, M.; Daune, M.; Granger-Schnarr, M. Effect of induction of SOS response on expression of pBR322 genes and on plasmid copy number. Plasmid 1989, 22, 163–168. [Google Scholar] [CrossRef]
- Khodursky, A.B.; Peter, B.J.; Schmid, M.B.; DeRisi, J.; Botstein, D.; Brown, P.O.; Cozzarelli, N.R. Analysis of topoisomerase function in bacterial replication fork movement: Use of DNA microarrays. Proc. Natl. Acad. Sci. USA 2000, 97, 9419–9424. [Google Scholar] [CrossRef]
- Khil, P.P.; Camerini-Otero, R.D. Over 1000 genes are involved in the DNA damage response of Escherichia coli. Mol. Microbiol. 2002, 44, 89–105. [Google Scholar] [CrossRef]
- Courcelle, J.; Khodursky, A.; Peter, B.; Brown, P.O.; Hanawalt, P.C. Comparative gene expression profiles following UV exposure in wild-type and SOS-deficient Escherichia coli. Genetics 2001, 158, 41–64. [Google Scholar] [CrossRef]
- Quillardet, P.; Rouffaud, M.A.; Bouige, P. DNA array analysis of gene expression in response to UV irradiation in Escherichia coli. Res. Microbiol. 2003, 154, 559–572. [Google Scholar] [CrossRef]
- Peterson, K.R.; Mount, D.W. Differential repression of SOS genes by unstable LexA41 (Tsl-1) protein causes a “split phenotype” in Escherichia coli K-12. J. Mol. Biol. 1987, 193, 27–40. [Google Scholar] [CrossRef]
- Feliciello, I.; Zahradka, D.; Zahradka, K.; Ivanković, S.; Puc, N.; Đermić, D. RecF, UvrD, RecX and RecN proteins suppress DNA degradation at DNA double-strand breaks in Escherichia coli. Biochimie 2018, 148, 116–126. [Google Scholar] [CrossRef] [PubMed]
- BioCyc. Available online: https://biocyc.org/gene?orgid=ECOLI&id=PD03831 (accessed on 19 June 2022).
- Quiñones, A.; Jüterbock, W.R.; Messer, W. Expression of the dnaA gene of Escherichia coli is inducible by DNA damage. Mol. Gen. Genet. 1991, 227, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Bobst, E.V.; Bobst, A.M.; Perrino, F.W.; Meyer, R.R.; Rein, D.C. Variability in the nucleic acid binding site size and the amount of single-stranded DNA-binding protein in Escherichia coli. FEBS Lett. 1985, 181, 133–137. [Google Scholar] [CrossRef]
- Salles, B.; Defais, M. Signal of induction of recA protein in E. coli. Mutat. Res./DNA Repair Rep. 1984, 131, 53–59. [Google Scholar] [CrossRef]
- Sassanfar, M.; Roberts, J.W. Nature of the SOS-inducing signal in Escherichia coli. The involvement of DNA replication. J. Mol. Biol. 1990, 212, 79–96. [Google Scholar] [CrossRef]
- Chaudhury, A.M.; Smith, G.R. Role of Escherichia coli RecBC enzyme in SOS induction. Mol. Gen. Genet. 1985, 201, 525–528. [Google Scholar] [CrossRef]
- Pennington, J.M.; Rosenberg, S.M. Spontaneous DNA breakage in single living Escherichia coli cells. Nat. Genet. 2007, 39, 797–802, Erratum in Nat. Genet. 2007, 39, 1174. [Google Scholar] [CrossRef]
- Wurihan; Gezi; Brambilla, E.; Wang, S.; Sun, H.; Fan, L.; Shi, Y.; Sclavi, B.; Morigen. DnaA and LexA Proteins Regulate Transcription of the uvrB Gene in Escherichia coli: The Role of DnaA in the Control of the SOS Regulon. Front. Microbiol. 2018, 9, 1212. [Google Scholar] [CrossRef]
- Schmidt, A.; Kochanowski, K.; Vedelaar, S.; Ahrne, E.; Volkmer, B.; Callipo, L.; Heinemann, M. The quantitative and condition-dependent Escherichia coli proteome. Nat. Biotechnol. 2016, 34, 104–110. [Google Scholar] [CrossRef]
- Zhao, T.; Liu, Y.; Wang, Z.; He, R.; Xiang Zhang, J.; Xu, F.; Lei, M.; Deci, M.B.; Nguyen, J.; Bianco, P.R. Super-resolution imaging reveals changes in Escherichia coli SSB localization in response to DNA damage. Genes Cells 2019, 24, 814–826. [Google Scholar] [CrossRef]
- Bachmann, B.J. Pedigrees of some mutant strains of Escherichia coli K-12. Bacteriol. Rev. 1972, 36, 525–557. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.H. A Short Course in Bacterial Genetics; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1992. [Google Scholar]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.; Karlen, Y.; Bakker, O.; van den Hoff, M.J.; Moorman, A.F. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [PubMed]
- Ruijter, J.M.; Pfaffl, M.W.; Zhao, S.; Spiess, A.N.; Boggy, G.; Blom, J.; Rutledge, R.G.; Sisti, D.; Lievens, A.; De Preter, K.; et al. Evaluation of qPCR curve analysis methods for reliable biomarker discovery: Bias, resolution, precision and implications. Methods 2013, 59, 32–46. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feliciello, I.; Đermić, E.; Malović, H.; Ivanković, S.; Zahradka, D.; Ljubić, S.; Procino, A.; Đermić, D. Regulation of ssb Gene Expression in Escherichia coli. Int. J. Mol. Sci. 2022, 23, 10917. https://doi.org/10.3390/ijms231810917
Feliciello I, Đermić E, Malović H, Ivanković S, Zahradka D, Ljubić S, Procino A, Đermić D. Regulation of ssb Gene Expression in Escherichia coli. International Journal of Molecular Sciences. 2022; 23(18):10917. https://doi.org/10.3390/ijms231810917
Chicago/Turabian StyleFeliciello, Isidoro, Edyta Đermić, Helena Malović, Siniša Ivanković, Davor Zahradka, Sven Ljubić, Alfredo Procino, and Damir Đermić. 2022. "Regulation of ssb Gene Expression in Escherichia coli" International Journal of Molecular Sciences 23, no. 18: 10917. https://doi.org/10.3390/ijms231810917
APA StyleFeliciello, I., Đermić, E., Malović, H., Ivanković, S., Zahradka, D., Ljubić, S., Procino, A., & Đermić, D. (2022). Regulation of ssb Gene Expression in Escherichia coli. International Journal of Molecular Sciences, 23(18), 10917. https://doi.org/10.3390/ijms231810917