

Participation of the Serotonergic System and Brain-Derived Neurotrophic Factor in the Antidepressant-like Effect of Flavonoids

 , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Serotonergic System and BDNF Involved in Depression Etiology and in Response to Antidepressants
3. Serotonergic System
4. Brain-Derived Neurotrophic Factor
5. Searching for Alternatives to Antidepressant Drugs
6. General Information of Flavonoids
6.1. Flavonols
6.2. Flavones
6.3. Flavanones
6.4. Flavanols
6.5. Isoflavones
6.6. Chalcones
6.7. Anthocyanidins
7. Pharmacokinetics of Flavonoids and Their Entry into the CNS
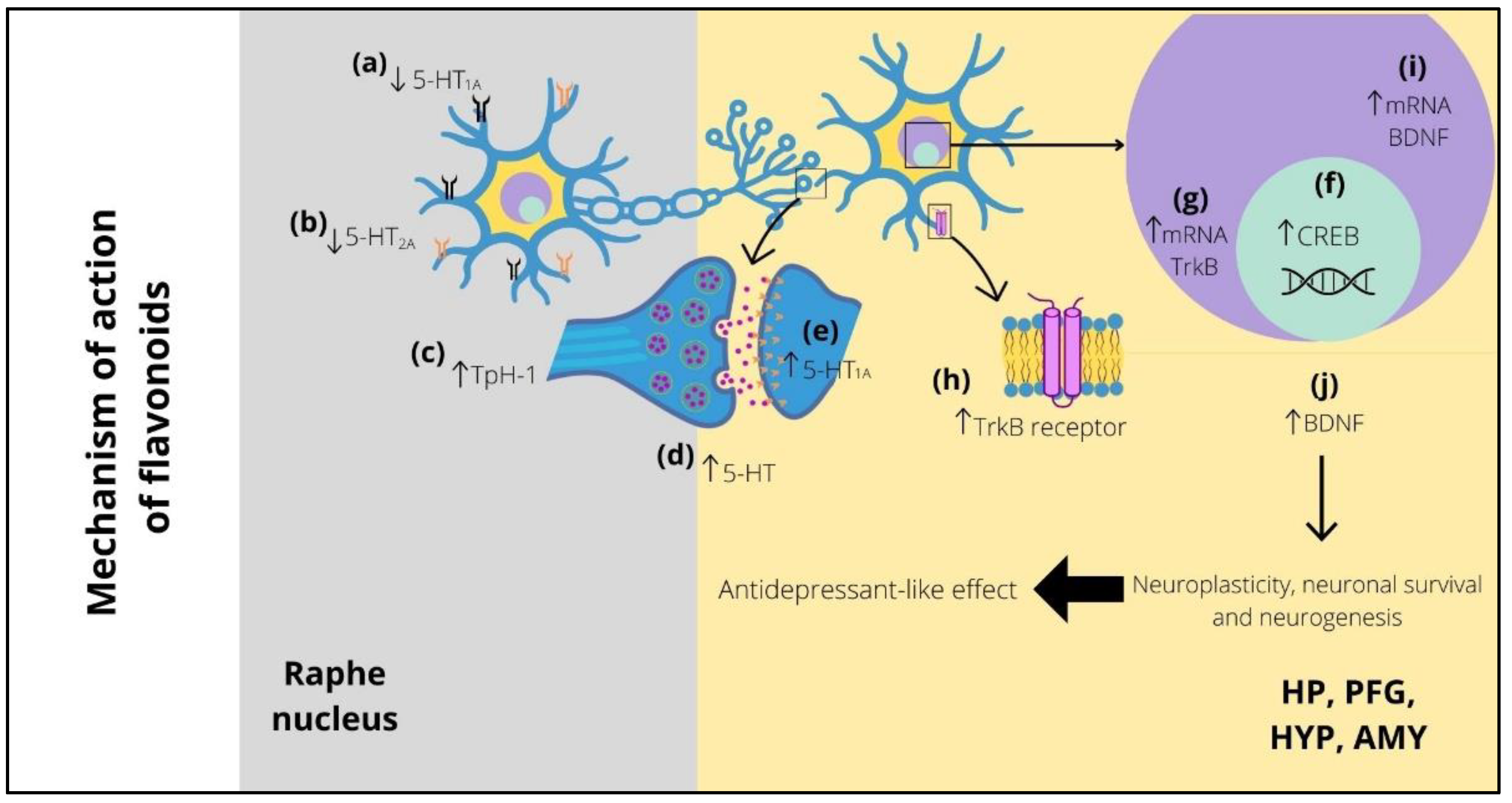
8. Participation of Serotonergic System in the Antidepressant-like Effect of Flavonoids

{kind=link}
| Flavonoid | Experimental Subjects | Treatment | Behavioral Effect | Effect on Serotonergic System | Reference |
|---|---|---|---|---|---|
| Astilbin (taxifolin-3-O-rhamnoside) | Adult male C57BL/6J mice | 10, 20, and 40 mg/kg (i.p.) for 21 days | ↓ TTI in FST and TST ↑ Sucrose intake | ↑ 5-HT in frontal cortex | [168] |
| Hesperidin (3,5,7-trihydroxyflavanone-7-rhamnoglucoside) | Male adult Swiss mice | 0.1, 0.3, and 1 mg/kg (i.p.) S.D. 30 min before behavioral test | ↓ TTI in FST and TST | Pretreatment with pCPA (100 mg/kg, i.p.) prevents antidepressant-like effect | [169] |
| Adult male Wistar rats with hyperglycemia induced by streptozotocin | 25, 50, and 100 mg/kg (p.o.) for 21 days | ↓ TTI in FST | ↑ Brain levels of 5-HT | [170] | |
| Male Swiss Albino mice | 1 mg/kg (i.p.) for 14 days | ↓ TTI in FST and TST | ↑ 5-HT in HP and cerebral cortex | [171] | |
| Old male Sprague-Dawley rats | 20, 50, and 100 mg/kg (i.p.) for 14 days | ↑ Sucrose intake ↓ TTI in FST | ↑ 5-HT in HP, PFC, and amygdala | [172] | |
| Rutin (quercetin-3-O-rhamnosylglucoside) | Male Swiss mice | 0.01, 0.1, 0.3, 1, 3, and 10 mg/kg (p.o.) 60 min before the behavioral test | ↓ TTI in FST | Pretreatment with pCPA (100 mg/kg, i.p.) prevents antidepressant-like effect | [173] |
| Five weeks old male Sprague Dawley rats | 225 mg/kg (p.o.) for 28 days | ↓ TTI ↑ Swimming time in FST | ↑ 5-HT in frontal cortex, HP, striatum, and amygdala | [174] | |
| Icariin (7-(β-D-Glucopyranosyloxy)-5-hydroxy-4′-methoxy-8-(3-methylbut-2-en-1-yl)-3-(α-L-rhamnopyranosyloxy) flavone) | Adult male Wistar rats | 30 and 60 mg/kg (p.o.) for 5 weeks | ↑ Sucrose intake | ↑ 5-HT1A mRNA levels in HP and frontal cortex | [175] |
| Orientin (luteolin-8-C-glucoside) | Adult male Kunming mice | 20 and 40 mg/kg (p.o.) 3 weeks | ↑ Sucrose intake | ↑ 5-HT in HP and PFC | [40] |
| Hyperoside (quercetin 3-galactoside) | Male Albino Swiss mice | 3.75 mg/kg (i.p.) 60 min before the behavioral test | ↓ TTI in FST and TST | Pretreatment with pCPA (100 mg/kg, i.p.) prevented antidepressant-like effect of hyperoside | [176] |
| Quercetin | Male Swiss Albino mice | 25 mg/kg (p.o.) for 4 weeks | ↓ TTI in FST and TST | ↑ Brain levels of 5-HT | [100] |
| Fisetin | Male ICR mice | 10 and 20 mg/kg (p.o.) 60 min before behavioral test | ↓ TTI in FST and TST | ↑ 5-HT in frontal cortex and HP | [166] |
| Vixetin (apigenin-8-C-glucopyranoside) | Adult male BALB/c mice | 10, 20, and 30 mg/kg (p.o.) 60 min before behavioral test | ↓ TTI in FST and TST | Pretreatment with NAN 190, a 5-HT1A antagonist (0.5 mg/kg, i.p.) prevented antidepressant-like effect of vixetin | [177] |
| Apigenin | Male ICR mice | 7, 10, 14, and 20 mg/kg (p.o.) for 2 weeks | ↓ TTI in FST ↑ Sucrose intake | ↑ 5-HT in PFC, HP, hypothalamus and nucleus accumbens of rats exposed to CMS | [178] |
| Albino mice (either sex) | 25 and 50 mg/kg (p.o.) 24, 5, and 1 h before the behavioral test | ↓ TTI in TST and FST | Pretreatment with pCPA (100 mg/kg, i.p.) prevented antidepressant-like effect of apigenin | [179] | |
| Naringenin (4′,5,7-trihydroxyflavanone-7-rhamnoglucoside) | Male ICR mice | 10, 20, and 50 mg/kg (p.o.) 60 min before the behavioral test | ↓ TTI in TST | Pretreatment with pCPA (100 mg/kg, i.p.) prevented antidepressant-like effect of naringenin | [180] |
| Three months old BALB/c male mice | 25, 50, and 100 mg/kg (p.o.) for 14 days | ↑ Sucrose intake ↓ TTI in FST | ↑ 5-HT in cortex and HP | [181] | |
| Silibinin | 6–8 weeks old Kunming mice | 100, 200, and 400 mg/kg (p.o.) for 3 weeks | ↓ TTI in TST and FST | ↑ 5-HT in PFC and HP | [182] |
| Eight weeks old male Sprague Dawley rats | 25, 50, and 100 mg/kg (i.p.) for 14 days | ↓ TTI in the FST ↑ Sucrose intake | ↑ 5-HT in HP and amygdala, and enhanced expression of TpH-1 mRNA in HP | [183] | |
| Chrysin | Male C57B/6J mice | 5 and 20 mg/kg (p.o.) for 14 days | ↓ TTI in FST | ↑ 5-HT in HP | [130] |
| Adult female C57BL/6 mice | 20 mg/kg (p.o.) for 28 days | ↓ TTI in FST and TST | ↑ 5-HT in PFC and HP | [184] | |
| Male Wistar rats | 5 mg/kg (p.o.) for 28 days | ↓ TTI in FST | ↓ 5-HT1A and 5-HT2A mRNA in raphe nucleus ↑ 5-HT1A mRNA in HP | [185] | |
| Nobiletin | Male ICR mice | 25, 50, and 100 mg/kg (p.o.) 60 min before the behavioral test | ↓ TTI in FST and TST | Pretreatment with WAY 100,635 (7.1 mg/kg, s.c., a serotonin 5-HT1A receptor antagonist) and cyproheptadine (3 mg/kg, i.p., a serotonin 5-HT2 receptor antagonist) prevented antidepressant-like effect of Nobiletin | [186] |
| Liquiritin (7-Hydroxyflavanone 4′-O-glucoside) and Isoliquiritin (2′,4,4′-Trihydroxychalcone 4-glucoside) | Mice | 10, 20, and 40 mg/kg (p.o.) 30 min before the behavioral test | ↓ TTI in FST and TST | ↑ 5-HT in HP, hypothalamus and cortex | [187] |
9. BDNF Implicated in the Antidepressant-like Effect of Flavonoids
| Flavonoid | Experimental Subjects and Condition | Dose and Treatment Duration | Behavioral Effects Related with Antidepressant Activity | Effect on BDNF | Reference |
|---|---|---|---|---|---|
| Baicalein | Male Sprague–Dawley rats exposed to restrain stress | 10, 20, and 40 mg/kg (i.p.) for 14 days | ↓ TTI in FST ↑ Sucrose intake | ↑ BDNF levels in HP | [192] |
| Male C57BL/6J mice with depressive-like behavior induced by rotetone | 300 mg/kg (p.o.) for 4 weeks | ↓ TTI in TST ↑ Sucrose intake | ↑ BDNF levels in HP ↑ TrkB phosphorylation | [193] | |
| Icariin | Male C57BL/6J mice exposed to social defeat | 5 and 10 mg/kg (p.o.) for 28 days | ↑ Interaction time | ↑ BDNF mRNA in HP | [194] |
| Hesperidin | Male adult Swiss mice | 0.3 and 1 mg/kg (i.p.) for 21 days | ↓ TTI in FST | ↑ BDNF levels in HP | [167] |
| Astibilin | Male C57BL/6J mice exposed to CUMS | 10, 20, and 40 mg/kg (i.p.) for 21 days | ↓ TTI in FST and TST ↑ Sucrose intake | ↑ BDNF levels in frontal cortex | [168] |
| Naringenin | Male ICR mice exposed to CUMS | 10 and 20 mg/kg (p.o.) for 21 days | ↑ Sucrose intake | ↑ BDNF and mRNA BDNF in HP | [195] |
| 7,8-dihydroxiflavone | Male C57BL/6 mice submitted to CMS | 10 and 20 mg/kg (i.p.) for 28 days | ↑ Sucrose intake | ↑ BDNF in HP and PFC Agonist to the TrkB receptor | [196] |
| Chrysin | Female C57BL/6J mice exposed to CUMS | 5 and 20 mg/kg (p.o.) for 28 days | ↑ Sucrose intake ↓ TTI in FST | ↑ BDNF and NGF in HP and PFC | [130] |
| Orientin | Male Kunming mice | 20 and 40 mg/kg (p.o.) for 21 days | ↑ Sucrose intake ↓ TTI in FST and TST | ↑ BDNF in HP and PFC | [40] |
| 3,5,6,7,8,30,40-Heptame thoxyflavone | C57BL/6 mice | 50 mg/kg (s.c.) for 25 days | ↓ TTI in FST and TST | ↑ BDNF in HP | [197] |
| Apigenin | Male ICR mice | 20 and 40 mg/kg (p.o.) for 21 days | ↑ Sucrose intake ↓ TTI in FST | ↑ BDNF in HP | [198] |
| Fisetin | Male ICR mice | 5 mg/kg (p.o.) for 21 days | ↓ TTI in FST and TST | ↑ Activation of TrkB receptor in HP | [199] |
| Silibinin | Male Sprague-Dawley rats with depression-like behavior induced by Aβ1-42 oligomers | 50 and 100 mg/kg (p.o.) for 15 days | ↓ TTI in FST and TST | ↑ BDNF and TrkB receptor expression in HP | [200] |
| Quercetin | Female C57BL/6J ERα-KO mice | 10 mg/kg (p.o.) for 10 weeks | ↓ TTI in FST and TST | ↑ BDNF and TrkB receptor expression | [201] |
10. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Boden, M.; Zimmerman, L.; Azevedo, K.J.; Ruzek, J.I.; Gala, S.; Magid, H.S.A.; McLean, C.P. Addressing the mental health impact of COVID-19 through population health. Clin. Psychol. Rev. 2021, 85, 102006. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Depression. Available online: https://www.who.int/news-room/fact-sheets/detail/depression (accessed on 18 May 2022).
- Senra, H.; McPherson, S. Depression in disabling medical conditions–current perspectives. Int. Rev. Psychiatry 2021, 33, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Chiu, M.; Vigod, S.; Rahman, F.; Wilton, A.S.; Lebenbaum, M.; Kurdyak, P. Mortality risk associated with psychological distress and major depression: A population-based cohort study. J. Affect. Disord. 2018, 234, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Bahramsoltani, R.; Farzaei, M.H.; Farahani, M.S.; Rahimi, R. Phytochemical constituents as future antidepressants: A comprehensive review. Rev. Neurosci. 2015, 26, 699–719. [Google Scholar] [CrossRef]
- Ali, A.M.; Hendawy, A.O. So, antidepressant drugs have serious adverse effects, but what are the alternatives. Nov. Appro. Drug Des. Dev. 2018, 4, 555636. [Google Scholar] [CrossRef]
- Artigas, F.; Bortolozzi, A.; Celada, P. Can we increase speed and efficacy of antidepressant treatments? Part I: General aspects and monoamine-based strategies. Eur. Neuropsychopharmacol. 2018, 28, 445–456. [Google Scholar] [CrossRef]
- Quentin, E.; Belmer, A.; Maroteaux, L. Somato-dendritic regulation of raphe serotonin neurons; a key to antidepressant action. Front. Neurosci. 2018, 12, 982. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.J.; Boulle, F.; Steinbusch, H.W.; van den Hove, D.L.; Kenis, G.; Lanfumey, L. Neurotrophic factors and neuroplasticity pathways in the pathophysiology and treatment of depression. Psychopharmacology 2018, 235, 2195–2220. [Google Scholar] [CrossRef]
- Mondal, A.C.; Fatima, M. Direct and indirect evidence of BDNF and NGF as key modulators in depression: Role of antidepressants treatment. Int. J. Neurosci. 2018, 129, 283–296. [Google Scholar] [CrossRef]
- Olchanski, N.; Myers, M.M.; Halseth, M.; Cyr, P.L.; Bockstedt, L.; Goss, T.F.; Howland, R.H. The economic burden of treatment-resistant depression. Clin. Ther. 2013, 35, 512–522. [Google Scholar] [CrossRef]
- Malhi, G.S.; Morris, G.; Bell, E.; Hamilton, A. A new paradigm for achieving a rapid antidepressant response. Drugs 2020, 80, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Wichniak, A.; Wierzbicka, A.; Walęcka, M.; Jernajczyk, W. Effects of antidepressants on sleep. Curr. Psychiatry Rep. 2017, 19, 63. [Google Scholar] [CrossRef] [PubMed]
- Rothmore, J. Antidepressant-induced sexual dysfunction. Med. J. Aust. 2020, 212, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Bae, H. Therapeutic effects of phytochemicals and medicinal herbs on depression. Biomed. Res. Int. 2017, 2017, 6596241. [Google Scholar] [CrossRef] [PubMed]
- German-Ponciano, L.J.; Rosas-Sánchez, G.U.; Rivadeneyra-Domínguez, E.; Rodríguez-Landa, J.F. Advances in the preclinical study of some flavonoids as potential antidepressant agents. Scientifica 2018, 2963565, 2963565. [Google Scholar] [CrossRef]
- Pannu, A.; Sharma, P.C.; Thakur, V.K.; Goyal, R.K. Emerging role of flavonoids as the treatment of depression. Biomolecules 2021, 11, 1825. [Google Scholar] [CrossRef]
- World Health Organization. Investing in Mental Health: Evidence for Action. Available online: https://www.who.int/publications/i/item/9789241564618 (accessed on 18 May 2022).
- Dean, J.; Keshavan, M. The neurobiology of depression: An integrated view. Asian J. Psychiatry 2017, 27, 101–111. [Google Scholar] [CrossRef]
- Ferrari, F.; Villa, R.F. The neurobiology of depression: An integrated overview from biological theories to clinical evidence. Mol. Neurobiol. 2017, 54, 4847–4865. [Google Scholar] [CrossRef]
- Bus, B.A.; Molendijk, M.L. The neurotrophic hypothesis of depression. Tijdschr. Psychiatr. 2016, 58, 215–222. [Google Scholar]
- Fung, J.; Gelaye, B.; Zhong, Q.Y.; Rondon, M.B.; Sanchez, S.E.; Barrios, Y.V.; Williams, M.A. Association of decreased serum brain-derived neurotrophic factor (BDNF) concentrations in early pregnancy with antepartum depression. BMC Psychiatry 2015, 15, 43. [Google Scholar] [CrossRef]
- Polyakova, M.; Stuke, K.; Schuemberg, K.; Mueller, K.; Schoenknecht, P.; Schroeter, M.L. BDNF as a biomarker for successful treatment of mood disorders: A systematic & quantitative meta-analysis. J. Affect. Disord. 2015, 174, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Rana, T.; Behl, T.; Sehgal, A.; Srivastava, P.; Bungau, S. Unfolding the role of BDNF as a biomarker for treatment of depression. J. Mol. Neurosci. 2021, 71, 2008–2021. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhong, J.; Zou, B.; Fang, L.; Chen, J.; Deng, X.; Lei, T. Meta-analyses of comparative efficacy of antidepressant medications on peripheral BDNF concentration in patients with depression. PLoS ONE 2017, 12, e0172270. [Google Scholar] [CrossRef] [PubMed]
- Kerling, A.; Kück, M.; Tegtbur, U.; Grams, L.; Weber-Spickschen, S.; Hanke, A.; Kahl, K.G. Exercise increases serum brain-derived neurotrophic factor in patients with major depressive disorder. J. Affect. Disord. 2017, 215, 152–155. [Google Scholar] [CrossRef]
- Sheldrick, A.; Camara, S.; Ilieva, M.; Riederer, P.; Michel, T.M. Brain-derived neurotrophic factor (BDNF) and neurotrophin 3 (NT3) levels in post-mortem brain tissue from patients with depression compared to healthy individuals—A proof of concept study. Eur. Psychiatry 2017, 46, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Levy, G.A.; Rocha, L.; Rodríguez-Pineda, F.; Alonso-Vanegas, M.A.; Nani, A.; Buentello-García, R.M.; Cruz-Fuentes, C.S. Increased expression of brain-derived neurotrophic factor transcripts I and VI, cAMP response element binding, and glucocorticoid receptor in the cortex of patients with temporal lobe epilepsy. Mol. Neurobiol. 2018, 55, 3698–3708. [Google Scholar] [CrossRef] [PubMed]
- Kraus, C.; Castrén, E.; Kasper, S.; Lanzenberger, R. Serotonin and neuroplasticity–links between molecular, functional and structural pathophysiology in depression. Neurosci. Biobehav. Rev. 2017, 77, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Chakrapani, S.; Eskander, N.; Lorenzo, A.; Omisore, B.A.; Mostafa, J.A. Neuroplasticity and the biological role of brain derived neurotrophic factor in the pathophysiology and management of depression. Cureus 2020, 12, e11396. [Google Scholar] [CrossRef]
- Ikram, H.; Haleem, D.J. Repeated treatment with reserpine as a progressive animal model of depression. Pak. J. Pharm. Sci. 2017, 30, 897–902. [Google Scholar]
- Fakhoury, M. Revisiting the Serotonin Hypothesis: Implications for Major Depressive Disorders. Mol. Neurobiol. 2016, 53, 2778–2786. [Google Scholar] [CrossRef]
- Yang, T.; Nie, Z.; Shu, H.; Kuang, Y.; Chen, X.; Cheng, J.; Liu, H. The role of BDNF on neural plasticity in depression. Front. Cell. Neurosci. 2020, 14, 82. [Google Scholar] [CrossRef] [PubMed]
- Soria-Fregozo, C.; Pérez-Vega, M.I.; Rodríguez-Landa, J.F.; Germán-Ponciano, L.J.; García-Ríos, R.I.; Mora-Perez, A. Association of 5-HT1A Receptors with Affective Disorders. In Serotonin-A Chemical Messenger between All Types of Living Cells; IntechOpen: London, UK, 2017. [Google Scholar]
- Björkholm, C.; Monteggia, L.M. BDNF–a key transducer of antidepressant effects. Neuropharmacology 2016, 102, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Kreinin, A.; Lisson, S.; Nesher, E.; Schneider, J.; Bergman, J.; Farhat, K.; Pinhasov, A. Blood BDNF level is gender specific in severe depression. PLoS ONE 2015, 10, e0127643. [Google Scholar] [CrossRef]
- Vahdati Hassani, F.; Naseri, V.; Razavi, B.M.; Mehri, S.; Abnous, K.; Hosseinzadeh, H. Antidepressant effects of crocin and its effects on transcript and protein levels of CREB, BDNF, and VGF in rat hippocampus. Daru 2014, 22, 16. [Google Scholar] [CrossRef] [PubMed]
- Cao, K.; Shen, C.; Yuan, Y.; Bai, S.; Yang, L.; Guo, L.; Shi, Y. SiNiSan ameliorates the depression-like behavior of rats that experienced maternal separation through 5-HT1A receptor/CREB/BDNF pathway. Front. Psychiatry 2019, 10, 160. [Google Scholar] [CrossRef] [PubMed]
- Popova, N.K.; Naumenko, V.S. Neuronal and behavioral plasticity: The role of serotonin and BDNF systems tandem. Expert Opin. Ther. Targets 2019, 23, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lan, N.; Ren, J.; Wu, Y.; Wang, S.T.; Huang, X.F.; Yu, Y. Orientin improves depression-like behavior and BDNF in chronic stressed mice. Mol. Nutr. Food Res. 2015, 59, 1130–1142. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, X.; Zhao, Y.; Liu, A.; Zhao, T.; Zhang, Y.; Teng, W. Elevated thyroid peroxidase antibody increases risk of post-partum depression by decreasing prefrontal cortex BDNF and 5-HT levels in mice. Front. Cell. Neurosci. 2017, 10, 307. [Google Scholar] [CrossRef]
- Yu, H.; Lv, D.; Shen, M.; Zhang, Y.; Zhou, D.; Chen, Z.; Wang, C. BDNF mediates the protective effects of scopolamine in reserpine-induced depression-like behaviors via up-regulation of 5-HTT and TPH1. Psychiatry Res. 2019, 271, 328–334. [Google Scholar] [CrossRef]
- Diniz, D.M.; Calabrese, F.; Brivio, P.; Riva, M.A.; Grandjean, J.; Homberg, J.R. BDNF overexpression in the ventral hippocampus promotes antidepressant-and anxiolytic-like activity in serotonin transporter knockout rats. Int. J. Mol. Sci. 2021, 22, 5040. [Google Scholar] [CrossRef]
- Mössner, R.; Daniel, S.; Albert, D.; Heils, A.; Okladnova, O.; Schmitt, A.; Lesch, K.P. Serotonin transporter function is modulated by brain-derived neurotrophic factor (BDNF) but not nerve growth factor (NGF). Neurochem. Int. 2000, 36, 197–202. [Google Scholar] [CrossRef]
- Chu, A.; Wadhwa, R. Selective serotonin reuptake inhibitors. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Saadati, H.; Sadegzadeh, F.; Sakhaie, N.; Panahpour, H.; Sagha, M. Serotonin depletion during the postnatal developmental period causes behavioral and cognitive alterations and decreases BDNF level in the brain of rats. Int. J. Dev. Neurosci. 2021, 81, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Dahlstroem, A.; Fuxe, K. Evidence for the existence of monoamine-containing neurons in the central nervous system. I. Demonstration of monoamines in the cell bodies of brain stem neurons. Acta Physiol. 1964, 232, 1–55. [Google Scholar]
- Törk, I. Raphe nuclei and serotonin containing systems. In The Rat Nervous System; Paxinos, G., Ed.; Academic Press: Sydney, Australia, 1964; Volume 2, pp. 43–78. [Google Scholar]
- Kawashima, T. The role of the serotonergic system in motor control. Neurosci. Res. 2018, 129, 32–39. [Google Scholar] [CrossRef]
- Lesch, K.P.; Waider, J. Serotonin in the modulation of neural plasticity and networks: Implications for neurodevelopmental disorders. Neuron 2012, 76, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Barnes, N.M.; Ahern, G.P.; Becamel, C.; Bockaert, J.; Camilleri, M.; Chaumont-Dubel, S. International union of basic and clinical pharmacology. CX Classification of receptors for 5-hydroxytryptamine; pharmacology and function. Pharmacol. Rev. 2021, 73, 310–520. [Google Scholar] [CrossRef]
- Anderson, I.M.; Tomenson, B.M. The efficacy of selective serotonin re-uptake inhibitors in depression: A meta-analysis of studies against tricyclic antidepressants. J. Psychopharmacol. 1994, 8, 238–249. [Google Scholar] [CrossRef]
- Nabeshima, T.; Kim, H.C. Involvement of genetic and environmental factors in the onset of depression. Exp. Neurobiol. 2013, 22, 235. [Google Scholar] [CrossRef]
- Duman, R.S.; Heninger, G.R.; Nestler, E.J. A molecular and cellular theory of depression. Arch. Gen. Psychiatry 1997, 54, 597–606. [Google Scholar] [CrossRef]
- Jacobsen, J.P.R.; Siesser, W.B.; Sachs, B.D.; Peterson, S.; Cools, M.J.; Setola, V.; Caron, M.G. Deficient serotonin neurotransmission and depression-like serotonin biomarker alterations in tryptophan hydroxylase 2 (Tph2) loss-of-function mice. Mol. Psychiatry 2012, 17, 694–704. [Google Scholar] [CrossRef]
- Albert, P.R.; Benkelfat, C.; Descarries, L. The neurobiology of depression—revisiting the serotonin hypothesis. I. Cellular and molecular mechanisms. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2012, 367, 2378–2381. [Google Scholar] [CrossRef] [PubMed]
- Mulinari, S. Monoamine theories of depression: Historical impact on biomedical research. J. Hist. Neurosci. 2012, 21, 366–392. [Google Scholar] [CrossRef] [PubMed]
- Malhi, G.S.; Mann, J.J. Depression. Lancet 2018, 392, 2299–2312. [Google Scholar] [CrossRef]
- Taylor, W.D.; Steffens, D.C.; Payne, M.E.; MacFall, J.R.; Marchuk, D.A.; Svenson, I.K.; Krishnan, K.R.R. Influence of serotonin transporter promoter region polymorphisms on hippocampal volumes in late-life depression. Arch. Gen. Psychiatry 2015, 62, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Perez-Caballero, L.; Torres-Sanchez, S.; Romero-López-Alberca, C.; González-Saiz, F.; Mico, J.A.; Berrocoso, E. Monoaminergic system and depression. Cell Tissue Res. 2019, 377, 107–113. [Google Scholar] [CrossRef]
- Racagni, G.; Popoli, M. Cellular and molecular mechanisms in the long-term action of antidepressants. Dialogues Clin. Neurosci. 2022, 10, 385–400. [Google Scholar] [CrossRef]
- Blier, P.; De Montigny, C. Current advances, and trends in the treatment of depression. Trends Pharmacol. Sci. 1994, 15, 220–226. [Google Scholar] [CrossRef]
- Hyman, S.E.; Nestler, E.J. Initiation and adaptation: A paradigm for understanding psychotropic drug action. Am. J. Psychiatry 1996, 153, 151–162. [Google Scholar] [CrossRef]
- Carlezon, W.A., Jr.; Duman, R.S.; Nestler, E.J. The many faces of CREB. Trends Neurosci. 2005, 28, 436–445. [Google Scholar] [CrossRef]
- Levada, O.A.; Cherednichenko, N.V. Brain-derived neurotrophic factor (BDNF): Neurobiology and marker value in neuropsychiatry. Lik. Sprava 2015, 15–25. [Google Scholar]
- Hollon, N.G.; Burgeno, L.M.; Phillips, P.E. Stress effects on the neural substrates of motivated behavior. Nat. Neurosci. 2015, 18, 1405–1412. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.W.; Chaudhury, D.; Han, M.H.; Nestler, E.J. Role of mesolimbic brain-derived neurotrophic factor in depression. Biol. Psychiatry 2019, 86, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Martinowich, K.; Lee, F. BDNF at the synapse: Why location matters. Mol. Psychiatry 2017, 22, 1370–1375. [Google Scholar] [CrossRef] [PubMed]
- Hing, B.; Sathyaputri, L.; Potash, J.B. A comprehensive review of genetic and epigenetic mechanisms that regulate BDNF expression and function with relevance to major depressive disorder. Am. J. Med. Genet B Neuropsychiatr. Genet. 2018, 177, 143–167. [Google Scholar] [CrossRef] [PubMed]
- Nibuya, M.; Morinobu, S.; Duman, R.S. Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. J. Neurosci. 1995, 15, 7539–7547. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, Y. Involvement of brain-derived neurotrophic factor in late-life depression. Am. J. Geriatr. Psychiatry 2013, 21, 433–449. [Google Scholar] [CrossRef]
- Ray, M.T.; Shannon-Weickert, C.; Webster, M.J. Decreased BDNF and TrkB mRNA expression in multiple cortical areas of patients with schizophrenia and mood disorders. Transl. Psychiatry 2014, 4, e389. [Google Scholar] [CrossRef]
- Guilloux, J.P.; Douillard-Guilloux, G.; Kota, R.; Wang, X.; Gardier, A.M.; Martinowich, K. Molecular evidence for BDNF- and GABA-related dysfunctions in the amygdala of female subjects with major depression. Mol. Psychiatry 2017, 17, 1130–1142. [Google Scholar] [CrossRef]
- Youssef, M.M.; Underwood, M.D.; Huang, Y.Y.; Hsiung, S.C.; Liu, Y.; Simpson, N.R. Association of BDNF Val66Met polymorphism and brain BDNF levels with major depression and suicide. Int. J. Neuropsychopharmacol. 2018, 21, 528–538. [Google Scholar] [CrossRef]
- Schroter, K.; Brum, M.; Brunkhorst-Kanaan, N.; Tole, F.; Ziegler, C.; Domschke, K. Longitudinal multi-level biomarker analysis of BDNF in major depression and bipolar disorder. Eur. Arch. Psychiatry Clin. Neurosci. 2020, 270, 169–181. [Google Scholar] [CrossRef]
- Zheleznyakova, G.Y.; Cao, H.; Schiöth, H.B. BDNF DNA methylation changes as a biomarker of psychiatric disorders: Literature review and open access database analysis. Behav. Brain Funct. 2016, 12, 17. [Google Scholar] [CrossRef] [PubMed]
- Carlino, D.; De Vanna, M.; Tongiorgi, E. Is altered BDNF biosynthesis a general feature in patients with cognitive dysfunctions. Neuroscientist 2013, 19, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Michalski, B.; Corrada, M.M.; Kawas, C.H.; Fahnestock, M. Brain derived neurotrophic factor and TrkB expression in the “oldest-old,” the 901 Study: Correlation with cognitive status and levels of soluble amyloid-beta. Neurobiol. Aging 2015, 36, 3130–3139. [Google Scholar] [CrossRef] [PubMed]
- Berry, A.; Panetta, P.; Luoni, A.; Bellisario, V.; Capoccia, S.; Riva, M.A.; Cirulli, F. Decreased BDNF expression and reduced social behavior in periadolescent rats following prenatal stress. Dev. Psychobiol. 2015, 57, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Murínová, J.; Hlaváčová, N.; Chmelová, M.; Riečanský, I. The evidence for altered BDNF expression in the brain of rats reared or housed in social isolation: A systematic review. Front. Behav. Neurosci. 2017, 11, 101. [Google Scholar] [CrossRef]
- Ashokan, A.; Hegde, A.; Mitra, R. Short-term environmental enrichment is sufficient to counter stress-induced anxiety and associated structural and molecular plasticity in basolateral amygdala. Psychoneuroendocrinology 2016, 69, 189–196. [Google Scholar] [CrossRef]
- Castrén, E.; Monteggia, L.M. Brain-derived neurotrophic factor signaling in depression and antidepressant action. Biol. Psychiatry 2021, 90, 128–136. [Google Scholar] [CrossRef]
- Polacchini, A.; Metelli, G.; Francavilla, R.; Baj, G.; Florean, M.; Mascaretti, L.G. A method for reproducible measurements of serum BDNF: Comparison of the performance of six commercial assays. Sci. Rep. 2015, 5, 17989. [Google Scholar] [CrossRef]
- Rantamäki, T. TrkB neurotrophin receptor at the core of antidepressant effects, but how? Cell Tissue Res. 2019, 377, 115–124. [Google Scholar] [CrossRef]
- Qi, X.; Zhao, J.; Liu, J.; Fang, H.; Swaab, D.F.; Zhou, J. Abnormal retinoid and TrkB signaling in the prefrontal cortex in mood disorders. Cereb. Cortex 2015, 25, 75–83. [Google Scholar] [CrossRef]
- Dwivedi, Y.; Rizavi, H.S.; Zhang, H.; Mondal, A.C.; Roberts, R.C.; Conley, R.R. Neurotrophin receptor activation and expression in human postmortem brain: Effect of suicide. Biol. Psychiatry 2009, 65, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.C.; Wu, J.; Fujita, Y.; Yao, W.; Ren, Q.; Yang, C.; Hashimoto, K. Antidepressant effects of TrkB ligands on depression-like behavior and dendritic changes in mice after inflammation. Int. J. Neuropsychopharmacol. 2015, 18, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Adachi, M.; Autry, A.E.; Mahgoub, M.; Suzuki, K.; Monteggia, L.M. TrkB signaling in dorsal raphe nucleus is essential for antidepressant efficacy and normal aggression behavior. Neuropsychopharmacology 2017, 42, 886–894. [Google Scholar] [CrossRef] [PubMed]
- De Vry, J.; Vanmierlo, T.; Martínez-Martínez, P.; Losen, M.; Temel, Y.; Boere, J.; Prickaerts, J. TrkB in the hippocampus and nucleus accumbens differentially modulates depression-like behavior in mice. Behav. Brain Res. 2016, 296, 15–25. [Google Scholar] [CrossRef]
- Casarotto, P.C.; Girych, M.; Fred, S.M.; Kovaleva, V.; Moliner, R.; Enkavi, G. Antidepressant drugs act by directly binding to TRKB neurotrophin receptors. Cell 2021, 184, 1299–1313. [Google Scholar] [CrossRef] [PubMed]
- Karson, C.N.; Newton, J.E.; Livingston, R.; Jolly, J.B.; Cooper, T.B.; Sprigg, J. Human brain fluoxetine concentrations. J. Neuropsychiatry Clin. Neurosci. 1993, 5, 322–329. [Google Scholar] [CrossRef]
- Pradier, M.F.; McCoy, T.H., Jr.; Hughes, M. Predicting treatment dropout after antidepressant initiation. Transl. Psychiatry 2020, 10, 60. [Google Scholar] [CrossRef]
- Simon, G.E.; Savarino, J.; Operskalski, B.; Wang, P.S. Suicide risk during antidepressant treatment. Am. J. Psychiatry 2006, 163, 41–47. [Google Scholar] [CrossRef]
- Alli, S.R.; Gorbovskaya, I.; Liu, J.; Kolla, N.J.; Brown, L.; Müller, D.J. The gut microbiome in depression and potential benefit of prebiotics, probiotics and synbiotics: A systematic review of clinical trials and observational studies. Int. J. Mol. Sci. 2022, 23, 4494. [Google Scholar] [CrossRef]
- Kang, M.; Hawken, E.; Vazquez, G.H. The mechanisms behind rapid antidepressant effects of ketamine: A systematic review with a focus on molecular neuroplasticity. Front. Psychiatry 2022, 13, 860882. [Google Scholar] [CrossRef]
- Pehrson, A.L.; Roberts, D.; Khawaja, A.; McNair, R. The role of serotonin neurotransmission in rapid antidepressant actions. Psychopharmacology 2022, 239, 1823–1838. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.F.; Pogačnik, L. Polyphenols from food and natural products: Neuroprotection and safety. Antioxidants 2020, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Galanakis, C.M. (Ed.) Polyphenols: Properties, Recovery, and Applications; Woodhead Publishing: Sawston, UK, 2018. [Google Scholar]
- Guan, L.P.; Liu, B.Y. Antidepressant-like effects and mechanisms of flavonoids and related analogues. Eur. J. Med. Chem. 2016, 121, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.; Najmi, A.K.; Akhtar, M.A. Natural phenolic compound quercetin showed the usefulness by targeting inflammatory, oxidative stress markers and augment 5-HT levels in one of the animal models of depression in mice. Drug Res. 2019, 69, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Vuolo, M.M.; Lima, V.S.; Junior, M.M. Phenolic compounds: Structure, classification, and antioxidant power. In Bioactive Compounds; Campos, M.R.S., Ed.; Woodhead Publishing: Sawston, UK, 2019; pp. 33–50. [Google Scholar]
- De Luna, S.L.; Ramírez-Garza, R.E.; Saldívar, S.O.S. Environmentally friendly methods for flavonoid extraction from plant material: Impact of their operating conditions on yield and antioxidant properties. Sci. World J. 2020, 2020, 6792069. [Google Scholar] [CrossRef]
- Hole, K.L.; Williams, R.J. Flavonoids as an Intervention for Alzheimer’s disease: Progress and hurdles towards defining a mechanism of action. Brain Plast. 2020, 6, 167–192. [Google Scholar] [CrossRef]
- Garg, S.K.; Shukla, A.; Choudhury, S. Nutraceuticals in Veterinary Medicine; Polyphenols and Flavonoids; Springer: Berlin, Germany, 2019; pp. 187–204. [Google Scholar]
- Saini, N.; Gahlawat, S.K.; Lather, V. Flavonoids: A nutraceutical and its role as anti-inflammatory and anticancer agent. In Plant Biotechnology: Recent Advancements and Developments; Springer: Singapore, 2017; pp. 255–270. [Google Scholar]
- Nabavi, S.M.; Šamec, D.; Tomczyk, M.; Milella, L.; Russo, D.; Habtemariam, S.; Shirooie, S. Flavonoid biosynthetic pathways in plants: Versatile targets for metabolic engineering. Biotechnol. Adv. 2020, 38, 107316. [Google Scholar] [CrossRef]
- Williamson, J.; Hughes, C.M.; Davison, G.W. Exogenous plant-based nutraceutical supplementation and peripheral cell mononuclear DNA damage following high intensity exercise. Antioxidants 2018, 7, 70. [Google Scholar] [CrossRef]
- Magalingam, K.B.; Radhakrishnan, A.; Haleagrahara, N. Protective effects of flavonol isoquercitrin, against 6-hydroxydopamine (6-OHDA)—induced toxicity in PC12 cells. BMC Res. Notes 2014, 7, 49. [Google Scholar] [CrossRef]
- Banjarnahor, S.D.; Artanti, N. Antioxidant properties of flavonoids. Med. J. Indones. 2014, 23, 239–244. [Google Scholar] [CrossRef]
- Fraga, C.G. Plant polyphenols: How to translate their in vitro antioxidant actions to in vivo conditions. IUBMB Life 2007, 59, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.Y.; Li, Q.; Bi, K.S. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. 2018, 13, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Denisov, E.T.; Denisova, T.G. The reactivity of natural phenols. Russ. Chem. Rev. 2009, 78, 1047–1073. [Google Scholar] [CrossRef]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Bose, S.; Sarkar, D.; Bose, A.; Mandal, S.C. Natural flavonoids and its pharmaceutical importance. Pharma Rev. 2018, 94, 61–75. [Google Scholar]
- Solnier, J.; Fladerer, J.P. Flavonoids: A complementary approach to conventional therapy of COVID-19? Phytochem. Rev. 2021, 20, 773–795. [Google Scholar] [CrossRef]
- Cao, G.; Sofic, E.; Prior, R.L. Antioxidant and prooxidant behavior of flavonoids: Structure-activity relationships. Free Radic. Biol. Med. 1997, 22, 749–760. [Google Scholar] [CrossRef]
- Nowak, M.; Tryniszewski, W.; Sarniak, A.; Wlodarczyk, A.; Nowak, P.J.; Nowak, D. Concentration dependence of anti- and pro-oxidant activity of polyphenols as evaluated with a light-emitting Fe2+-Egta-H2O2 System. Molecules 2022, 27, 3453. [Google Scholar] [CrossRef]
- Laughton, M.J.; Halliwell, B.; Evans, P.J.; Robin, J.; Hoult, S. Antioxidant, and pro-oxidant actions of the plant phenolics quercetin, gossypol and myricetin: Effects on lipid peroxidation, hydroxyl radical generation and bleomycin-dependent damage to DNA. Biochem. Pharmacol. 1989, 38, 2859–2865. [Google Scholar] [CrossRef]
- Halliwell, B. Are polyphenols antioxidants or pro-oxidants? What do we learn from cell culture and in vivo studies? Arch. Biochem. Biophys. 2008, 476, 107–112. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, J.; Xie, Y. Improvement strategies for the oral bioavailability of poorly water-soluble flavonoids: An overview. Int. J. Pharm. 2019, 570, 118642. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Kensler, T.W. Role of dietary supplements/nutraceuticals in chemoprevention through induction of cytoprotective enzymes. Chem. Res. Toxicol. 2007, 20, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Kocyigit, A.; Koyuncu, I.; Dikilitas, M.; Bahadori, F.; Turkkan, B. Cytotoxic, genotoxic and apoptotic effects of naringenin-oxime relative to naringenin on normal and cancer cell lines. Asian Pac. J. Trop. Med. 2016, 6, 872–880. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Barreca, D.; Mandalari, G.; Calderaro, A.; Smeriglio, A.; Trombetta, D.; Felice, M.R.; Gattuso, G. Citrus flavones: An update on sources, biological functions, and health promoting properties. Plants 2020, 9, 288. [Google Scholar] [CrossRef]
- D’Andrea, G. Quercetin: A flavonol with multifaceted therapeutic applications? Fitoterapia 2015, 106, 256–271. [Google Scholar] [CrossRef]
- De Mendonça, C.R.; Manhães-de-Castro, R.; de Santana, B.J.R.C.; da Silva, L.O.; Toscano, A.E.; Guzmán-Quevedo, O.; Galindo, L.C.M. Effects of flavonols on emotional behavior and compounds of the serotonergic system: A preclinical systematic review. Eur. J. Pharmacol. 2021, 916, 174697. [Google Scholar] [CrossRef]
- Manach, C.; Donovan, J.L. Pharmacokinetics and metabolism of dietary flavonoids in humans. Free. Radic. Res. 2004, 38, 771–786. [Google Scholar] [CrossRef]
- Ahmad, A.; Kumari, P.; Ahmad, M. Apigenin attenuates edifenphos-induced toxicity by modulating ROS-mediated oxidative stress, mitochondrial dysfunction and caspase signal pathway in rat liver and kidney. Pestic. Biochem. Physiol. 2019, 159, 163–172. [Google Scholar] [CrossRef]
- Filho, C.B.; Jesse, C.R.; Donato, F.; Giacomeli, R.; Del Fabbro, L.; da Silva Antunes, M.; de Gomes, M.G.; Goes, A.T.; Boeira, S.P.; Prigol, M. Chronic unpredictable mild stress decreases BDNF and NGF levels and Na+,K+- ATPase activity in the hippocampus and prefrontal cortex of mice: Antidepressant effect of chrysin. Neuroscience 2015, 289, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Filho, C.B.; Jesse, C.R.; Donato, F.; Del Fabbro, L.; Gomes, M.; Rossito-Goes, A.T.; Souza, L.C.; Boeira, S.P. Chrysin promotes attenuation of depressive-like behavior and hippocampal dysfunction resulting from olfactory bulbectomy in mice. Chem. Biol. Interact. 2016, 260, 154–162. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, D.R.; Todo, A.H.; Rêgo, G.M.; Cerutti, J.M.; Cavalheiro, A.J.; Rando, D.G.G.; Cerutti, S.M. Flavones-bound in benzodiazepine site on GABAA receptor: Concomitant anxiolytic-like and cognitive-enhancing effects produced by Isovitexin and 6-C-glycoside-Diosmetin. Eur. J. Pharmacol. 2018, 831, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xu, B.; Sun, J.; Jiang, X.; Bai, W. Anthocyanin supplement as a dietary strategy in cancer prevention and management: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2021, 62, 7242–7254. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zuo, Z.; Lu, S.; Liu, A.; Liu, X. Naringin attenuates diabetic retinopathy by inhibiting inflammation, oxidative stress and NF-κB activation in vivo and in vitro. Iran J. Basic Med. Sci. 2017, 20, 813. [Google Scholar] [CrossRef]
- Jung, U.J.; Kim, S.R. Effects of naringin, a flavanone glycoside in grapefruits and citrus fruits, on the nigrostriatal dopaminergic projection in the adult brain. Neural Regen. Res. 2014, 9, 1514. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef]
- Di Pietro, N.; Baldassarre, M.P.A.; Cichelli, A.; Pandolfi, A.; Formoso, G.; Pipino, C. Role of polyphenols and carotenoids in endothelial dysfunction: An overview from classic to innovative biomarkers. Oxidative Med. Cell. Longev. 2020, 2020, 6381380. [Google Scholar] [CrossRef]
- Pons, Z.; Margalef, M.; Bravo, F.I.; Arola-Arnal, A.; Muguerza, B. Grape seed flavanols decrease blood pressure via Sirt-1 and confer a vasoprotective pattern in rats. J. Funct. Foods 2016, 24, 164–172. [Google Scholar] [CrossRef]
- Martin, M.Á.; Ramos, S. Impact of cocoa flavanols on human health. Food Chem. Toxicol. 2021, 151, 112121. [Google Scholar] [CrossRef]
- Matthies, A.; Clavel, T.; Gütschow, M.; Engst, W.; Haller, D.; Blaut, M.; Braune, A. Conversion of daidzein and genistein by an anaerobic bacterium newly isolated from the mouse intestine. Appl. Environ. Microbiol. 2008, 74, 4847–4852. [Google Scholar] [CrossRef] [PubMed]
- Desmawati, D.; Sulastri, D. Phytoestrogens and their health effect. Open Access Maced. J. Med. Sci. 2019, 7, 495. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.R.; Ko, N.Y.; Chen, K.H. Isoflavone supplements for menopausal women: A systematic review. Nutrients 2019, 11, 2649. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Birru, R.L.; Snitz, B.E.; Ihara, M.; Kakuta, C.; Lopresti, B.J.; Sekikawa, A. Effects of soy isoflavones on cognitive function: A systematic review and meta-analysis of randomized controlled trials. Nutr. Rev. 2020, 78, 134–144. [Google Scholar] [CrossRef]
- Goyal, K.; Kaur, R.; Goyal, A.; Awasthi, R. Chalcones: A review on synthesis and pharmacological activities. J. Appl. Pharm. Sci. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Zhao, D.H.; Wang, Y.C.; Zheng, L.W.; Liu, B.Y.; Guan, L.P. Antidepressant-like effect of a chalcone compound, DHIPC and its possible mechanism. Iran J. Pharm. Res. 2018, 17, 193. [Google Scholar]
- Burton-Freeman, B.; Brzeziński, M.; Park, E.; Sandhu, A.; Xiao, D.; Edirisinghe, I. A selective role of dietary anthocyanins and flavan-3-ols in reducing the risk of type 2 diabetes mellitus: A review of recent evidence. Nutrients 2019, 11, 841. [Google Scholar] [CrossRef]
- Millar, C.L.; Duclos, Q.; Blesso, C.N. Effects of dietary flavonoids on reverse cholesterol transport, HDL metabolism, and HDL function. Adv. Nutr. 2017, 8, 226–239. [Google Scholar] [CrossRef]
- Fernández-Demeneghi, R.; Vargas-Moreno, I.; Acosta-Mesa, H.G.; Puga-Olguín, A.; Campos-Uscanga, Y.; Romo-González, T.; Guzmán-Gerónimo, R.I.; Patraca-Camacho, L.; Herrera-Meza, S. Berry Supplementation and Their Beneficial Effects on Some Central Nervous System Disorders. In Behavioral Pharmacology-From Basic to Clinical Research; Rodríguez-Landa, J.F., Cueto-Escobedo, J., Eds.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Ali, S.; Corbi, G.; Maes, M.; Scapagnini, G.; Davinelli, S. Exploring the impact of flavonoids on symptoms of depression: A systematic review and meta-analysis. Antioxidants 2021, 10, 1644. [Google Scholar] [CrossRef]
- Estrada-Reyes, R.; Ubaldo-Suárez, D.; Araujo-Escalona, A.G. Los flavonoides y el sistema nervioso central. Salud Mental 2012, 35, 375–384. [Google Scholar]
- Kaeko, M.; Yoshimasa, N.; Mariko, U. Flavonoid metabolism: The interaction of metabolites and gut microbiota. Biosci. Biotechnol. Biochem. 2018, 82, 600–610. [Google Scholar] [CrossRef]
- Ren, W.; Qian, Z.; Wang, H.; Zhu, L.; Zhang, L. Flavonoids: Promising anticancer agents. Med. Res. Rev. 2003, 23, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.H.; Kim, S.K.; Lee, S.Y.; Jang, C.G. Flavonoids as therapeutic candidates for emotional disorders such as anxiety and depression. Arch. Pharm. Res. 2020, 43, 1128–1143. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Merino, C.; Lopez-Sanchez, C.; Lagoa, R.; Samhan-Arias, A.; Bueno, C.; Garcia-Martinez, V. Neuroprotective actions of flavonoids. Curr. Med. Chem. 2011, 18, 1195–1212. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Rauf, A.; Shah, Z.A.; Saeed, F.; Imran, A.; Arshad, M.U.; Mubarak, M.S. Chemo-preventive and therapeutic effect of the dietary flavonoid kaempferol: A comprehensive review. Phytother. Res. 2019, 33, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Hembrom, S. Neuroprotective effect of flavonoids: A systematic review. Int. J. Aging Res. 2019, 2, 26. [Google Scholar] [CrossRef]
- Xiao, J. Recent advances in dietary flavonoids for management of type 2 diabetes. Curr. Opin. Food Sci. 2022, 44, 100806. [Google Scholar] [CrossRef]
- Faria, A.; Mateus, N.; Calhau, C. Flavonoid transport across blood-brain barrier: Implication for their direct neuroprotective actions. Nutr. Aging 2012, 1, 89–97. [Google Scholar] [CrossRef]
- Abd El Mohsen, M.M.; Kuhnle, G.; Rechner, A.R.; Schroeter, H.; Rose, S.; Jenner, P.; Rice-Evans, C.A. Uptake and metabolism of epicatechin and its access to the brain after oral ingestion. Free Radic. Biol. Med. 2002, 33, 1693–1702. [Google Scholar] [CrossRef]
- Faria, A.; Meireles, M.; Fernandes, I.; Santos-Buelga, C.; Gonzalez-Manzano, S.; Dueñas, M.; Calhau, C. Flavonoid metabolites transport across a human BBB model. Food Chem. 2014, 149, 190–196. [Google Scholar] [CrossRef]
- Ishisaka, A.; Ichikawa, S.; Sakakibara, H.; Piskula, M.K.; Nakamura, T.; Kato, Y.; Terao, J. Accumulation of orally administered quercetin in brain tissue and its antioxidative effects in rats. Free Radic. Biol. Med. 2011, 51, 1329–1336. [Google Scholar] [CrossRef] [PubMed]
- Milbury, P.E.; Kalt, W. Xenobiotic metabolism and berry flavonoid transport across the blood-brain barrier. J. Agric. Food Chem. 2010, 58, 3950–3956. [Google Scholar] [CrossRef] [PubMed]
- Milbury, P.E.; Vita, J.A.; Blumberg, J.B. Anthocyanins are bioavailable in humans following an acute dose of cranberry juice. J Nutr. 2010, 140, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Najmanová, I.; Pourová, J.; Mladěnka, P. A Mixture of phenolic metabolites of quercetin can decrease elevated blood pressure of spontaneously hypertensive rats even in low doses. Nutrients 2020, 12, 213. [Google Scholar] [CrossRef] [PubMed]
- Abelaira, H.M.; Réus, G.Z.; Quevedo, J. Animal models as tools to study the pathophysiology of depression. Braz. J. Psychiatry 2013, 35, S112–S120. [Google Scholar] [CrossRef]
- Zhen, L.; Zhu, J.; Zhao, X.; Huang, W.; An, Y.; Li, S.; Du, X.; Lin, M.; Wang, Q.; Xu, Y.; et al. The antidepressant-like effect of fisetin involves the serotonergic and noradrenergic system. Behav. Brain Res. 2012, 228, 359–366. [Google Scholar] [CrossRef]
- Donato, F.; de Gomes, M.G.; Goes, A.T.; Filho, C.B.; Del Fabbro, L.; Antunes, M.S.; Souza, L.C.; Boeira, S.P.; Jesse, C.R. Hesperidin exerts antidepressant-like effects in acute and chronic treatments in mice: Possible role of l-arginine-NO-cGMP pathway and BDNF levels. Brain Res. Bull. 2014, 104, 19–26. [Google Scholar] [CrossRef]
- Lv, Q.Q.; Wu, W.J.; Guo, X.L.; Liu, R.L.; Yang, Y.P.; Zhou, D.S.; Zhang, J.X.; Liu, J.Y. Antidepressant activity of astilbin: Involvement of monoaminergic neurotransmitters and BDNF signal pathway. Biol. Pharm. Bull. 2014, 37, 987–995. [Google Scholar] [CrossRef]
- Souza, L.C.; de Gomes, M.G.; Goes, A.T.; Del Fabbro, L.; Filho, C.B.; Boeira, S.P.; Jesse, C.R. Evidence for the involvement of the serotonergic 5-HT(1A) receptors in the antidepressant-like effect caused by hesperidin in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 40, 103–109. [Google Scholar] [CrossRef]
- El-Marasy, S.A.; Abdallah, H.M.; El-Shenawy, S.M.; El-Khatib, A.S.; El-Shabrawy, O.A.; Kenawy, S.A. Anti-depressant effect of hesperidin in diabetic rats. Can. J. Physiol. Pharmacol. 2014, 92, 945–952. [Google Scholar] [CrossRef]
- Nadar, J.S.; Kale, P.P.; Kadu, P.K.; Prabhavalkar, K.; Dhangar, R. Potentiation of antidepressant effects of agomelatine and bupropion by hesperidin in mice. Neurol. Res. Int. 2018, 2018, 9828639. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Choi, G.M.; Sur, B. Antidepressant-like effects of hesperidin in animal model of post-traumatic stress disorder. Chin. J. Integr. Med. 2021, 27, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.G.; Bettio, L.E.; Cunha, M.P.; Santos, A.R.; Pizzolatti, M.G.; Brighente, I.M.; Rodrigues, A.L. Antidepressant-like effect of rutin isolated from the ethanolic extract from Schinus molle L. in mice: Evidence for the involvement of the serotonergic and noradrenergic systems. Eur. J. Pharmacol. 2008, 587, 163–168. [Google Scholar] [CrossRef]
- Lin, S.H.; Chang, H.C.; Chen, P.J.; Hsieh, C.L.; Su, K.P.; Sheen, L.Y. The antidepressant-like effect of ethanol extract of daylily flowers (Jīn Zhēn Huā) in rats. J. Tradit. Complement. Med. 2013, 3, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Wang, F.M.; Qiang, L.Q.; Zhang, D.M.; Kong, L.D. Icariin attenuates chronic mild stress-induced dysregulation of the LHPA stress circuit in rats. Psychoneuroendocrinology 2010, 35, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Orzelska-Górka, J.; Szewczyk, K.; Gawrońska-Grzywacz, M.; Kędzierska, E.; Głowacka, E.; Herbet, M.; Dudka, J.; Biała, G. Monoaminergic system is implicated in the antidepressant-like effect of hyperoside and protocatechuic acid isolated from Impatiens glandulifera Royle in mice. Neurochem. Int. 2019, 12, 206–214. [Google Scholar] [CrossRef]
- Can, Ö.D.; Demir-Özkay, Ü.; Üçel, U.İ. Anti-depressant-like effect of vitexin in BALB/c mice and evidence for the involvement of monoaminergic mechanisms. Eur. J. Pharmacol. 2013, 699, 250–257. [Google Scholar] [CrossRef]
- Yi, L.T.; Li, J.M.; Li, Y.C.; Pan, Y.; Xu, Q.; Kong, L.D. Antidepressant-like behavioral and neurochemical effects of the citrus-associated chemical apigenin. Life Sci. 2008, 82, 741–751. [Google Scholar] [CrossRef]
- Al-Yamani, M.J.; Mohammed-Basheeruddin, S.; Alamri, A.S.; Alsanie, W.F.; Alhomrani, M.; Alsalman, A.J.; Al-Mohaini, M.; Al-Hawaj, M.A.; Alanazi, A.A.; Alanzi, K.D.; et al. The role of serotonergic and catecholaminergic systems for possible antidepressant activity of apigenin. Saudi J. Biol. Sci. 2022, 29, 11–17. [Google Scholar] [CrossRef]
- Yi, L.T.; Li, C.F.; Zhan, X.; Cui, C.C.; Xiao, F.; Zhou, L.P.; Xie, Y. Involvement of monoaminergic system in the antidepressant-like effect of the flavonoid naringenin in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2010, 34, 1223–1228. [Google Scholar] [CrossRef]
- Bansal, Y.; Singh, R.; Saroj, P.; Sodhi, R.K.; Kuhad, A. Naringenin protects against oxido-inflammatory aberrations and altered tryptophan metabolism in olfactory bulbectomized-mice model of depression. Toxicol. Appl. Pharmacol. 2018, 355, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.J.; Tan, Y.C.; Xu, J.C.; Tang, X.P.; Zhang, C.; Zhang, P.B.; Ren, Z.Q. Protective effects of silibinin and its possible mechanism of action in mice exposed to chronic unpredictable mild stress. Biomol. Ther. 2015, 23, 245–250. [Google Scholar] [CrossRef][Green Version]
- Lee, B.; Choi, G.M.; Sur, B. Silibinin prevents depression-like behaviors in a single prolonged stress rat model: The possible role of serotonin. BMC Complement. Med. Ther. 2020, 20, 70. [Google Scholar] [CrossRef] [PubMed]
- Bortolotto, V.C.; Pinheiro, F.C.; Araujo, S.M.; Poetini, M.R.; Bertolazi, B.S.; de Paula, M.T.; Meichtry, L.B.; de Almeida, F.P.; de Freitas Couto, S.; Jesse, C.R.; et al. Chrysin reverses the depressive-like behavior induced by hypothyroidism in female mice by regulating hippocampal serotonin and dopamine. Eur. J. Pharmacol. 2018, 822, 78–84. [Google Scholar] [CrossRef] [PubMed]
- German-Ponciano, L.J.; Rosas-Sánchez, G.U.; Ortiz-Guerra, S.I.; Soria-Fregozo, C.; Rodríguez-Landa, J.F. Effects of chrysin on mRNA expression of 5-HT1A and 5-HT2A receptors in the raphe nuclei and hippocampus. Rev. Bras. Farmacogn. 2021, 31, 353–360. [Google Scholar] [CrossRef]
- Yi, L.T.; Xu, H.L.; Feng, J.; Zhan, X.; Zhou, L.P.; Cui, C.C. Involvement of monoaminergic systems in the antidepressant-like effect of nobiletin. Physiol. Behav. 2011, 102, 1–6. [Google Scholar] [CrossRef]
- Wang, W.; Hu, X.; Zhao, Z.; Liu, P.; Hu, Y.; Zhou, J.; Guo, H. Antidepressant-like effects of liquiritin and isoliquiritin from Glycyrrhiza uralensis in the forced swimming test and tail suspension test in mice. Progress in Neuro- Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 1179–1184. [Google Scholar] [CrossRef]
- Justin-Thenmozhi, A.; Dhivya Bharathi, M.; Kiruthika, R.; Manivasagam, T.; Borah, A.; Essa, M.M. Attenuation of aluminum chloride-induced neuroinflammation and caspase activation through the AKT/GSK-3β pathway by hesperidin in wistar rats. Neurotox. Res. 2018, 34, 463–476. [Google Scholar] [CrossRef]
- Herath, H.M.U.L.; Piao, M.J.; Kang, K.A.; Zhen, A.X.; Fernando, P.D.S.M.; Kang, H.K.; Yi, J.M.; Hyun, J.W. Hesperidin exhibits protective effects against PM2.5-mediated mitochondrial damage, cell cycle arrest, and cellular senescence in human HaCaT Keratinocytes. Molecules 2022, 27, 4800. [Google Scholar] [CrossRef]
- Sharma, P.; Kumar, A.; Singh, D. Dietary flavonoids interaction with CREB-BDNF pathway: An unconventional approach for comprehensive management of epilepsy. Curr. Neuropharmacol. 2019, 17, 1158–1175. [Google Scholar] [CrossRef]
- Neshatdoust, S.; Saunders, C.; Castle, S.M.; Vauzour, D.; Williams, C.; Butler, L.; Lovegrove, J.A.; Spencer, J.P. High-flavonoid intake induces cognitive improvements linked to changes in serum brain-derived neurotrophic factor: Two randomised, controlled trials. Nutr. Healthy Aging 2016, 4, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Sur, B.; Park, J.; Kim, S.-H.; Kwon, S.; Yeom, M.; Shim, I.; Lee, H.; Hahm, D.-H. Chronic administration of baicalein decreases depression-like behavior induced by repeated restraint stress in rats. Korean J. Physiol. Pharmacol. 2013, 17, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Kong, D.; Zhou, Q.; Wei, G.; Song, J.; Liang, Y.; Du, G. Baicalein alleviates depression-like behavior in rotenone-induced Parkinson’s disease model in mice through activating the BDNF/TrkB/CREB pathway. Biomed. Pharmacother. 2021, 140, 111556. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wu, J.; Xia, S.; Li, B.; Dong, J. Icaritin opposes the development of social aversion after defeat stress via increases of GR mRNA and BDNF mRNA in mice. Behav. Brain Res. 2013, 256, 602–608. [Google Scholar] [CrossRef]
- Yi, L.T.; Liu, B.B.; Li, J.; Luo, L.; Liu, Q.; Geng, D.; Tang, Y.; Xia, Y.; Wu, D. BDNF signaling is necessary for the antidepressant-like effect of naringenin. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 48, 135–141. [Google Scholar] [CrossRef]
- Zhang, M.W.; Zhang, S.F.; Li, Z.H.; Han, F. 7,8- Dihydroxyflavone reverses the depressive symptoms in mouse chronic mild stress. Neurosci. Lett. 2016, 635, 33–38. [Google Scholar] [CrossRef]
- Sawamoto, A.; Okuyama, S.; Yamamoto, K.; Amakura, Y.; Yoshimura, M.; Nakajima, M.; Furukawa, Y. 3,5,6,7,8,30,40-Heptamethoxyflavone, a citrus flavonoid, ameliorates corticosterone-induced depression-like behavior and restores brain-derived neurotrophic factor expression, neurogenesis, and neuroplasticity in the hippocampus. Molecules 2016, 21, 541. [Google Scholar] [CrossRef]
- Weng, L.; Guo, X.; Li, Y.; Yang, X.; Han, Y. Apigenin reverses depression-like behavior induced by chronic corticosterone treatment in mice. Eur. J. Pharm. 2016, 774, 50–54. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, B.; Lu, J.; Shi, H.; Gong, S.; Wang, Y.; Hamdy, R.C.; Chua, B.H.L.; Yang, L.; Xu, X. Fisetin provides antidepressant effects by activating the tropomyosin receptor kinase B signal pathway in mice. J. Neurochem. 2017, 143, 561–568. [Google Scholar] [CrossRef]
- Song, X.; Liu, B.; Cui, L.; Zhou, B.; Liu, W.; Xu, F.; Ikejima, T. Silibinin ameliorates anxiety/depression-like behaviors in amyloid β-treated rats by upregulating BDNF/TrkB pathway and attenuating autophagy in hippocampus. Physiol. Behav. 2017, 179, 487–493. [Google Scholar] [CrossRef]
- Wang, G.; Li, Y.; Lei, C.; Lei, X.; Zhu, X.; Yang, L.; Zhang, R. Quercetin exerts antidepressant and cardioprotective effects in estrogen receptor α-deficient female mice via BDNF-AKT/ERK1/2 signaling. J. Steroid. Biochem. Mol. Biol. 2021, 206, 105795. [Google Scholar] [CrossRef] [PubMed]
- Gold, S.M.; Köhler-Forsberg, O.; Moss-Morris, R. Comorbid depression in medical diseases. Nat. Rev. Dis. Primers 2020, 6, 69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.C.; Yao, W.; Dong, C. Comparison of ketamine, 7,8-dihydroxyflavone, and ANA-12 antidepressant effects in the social defeat stress model of depression. Psychopharmacology 2015, 232, 4325–4335. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.C.; Dang, Y.H.; Jia, M.; Ma, R.; Wang, F.; Wu, J.; Gao, C.G.; Hashimoto, K. Long-lasting antidepressant action of ketamine, but not glycogen synthase kinase-3 inhibitor SB216763, in the chronic mild stress model of mice. PLoS ONE 2013, 8, e56053. [Google Scholar] [CrossRef]
- Liu, W.X.; Wang, J.; Xie, Z.M.; Xu, N.; Zhang, G.F.; Jia, M.; Zhou, Z.Q.; Hashimoto, K.; Yang, J.J. Regulation of glutamate transporter 1 via BDNF-TrkB signaling plays a role in the anti-apoptotic and antidepressant effects of ketamine in chronic unpredictable stress model of depression. Psychopharmacolog 2016, 233, 405–415. [Google Scholar] [CrossRef]
- Yamamoto, S.; Ohba, H.; Nishiyama, S.; Harada, N.; Kakiuchi, T.; Tsukada, H.; Domino, E.F. Subanesthetic doses of ketamine transiently decrease serotonin transporter activity: A PET study in conscious monkeys. Neuropsychopharmacology 2013, 38, 2666–2674. [Google Scholar] [CrossRef]
- Chang, L.; Zhang, K.; Pu, Y.; Qu, Y.; Wang, S.M.; Xiong, Z.; Ren, Q.; Dong, C.; Fujita, Y.; Hashimoto, K. Comparison of antidepressant and side effects in mice after intranasal administration of (R,S)-ketamine, (R)-ketamine, and (S)-ketamine. Pharmacol. Biochem. Behav. 2019, 181, 53–59. [Google Scholar] [CrossRef]
- Domino, E.F. Taming the ketamine tiger. 1965. Anesthesiology 2010, 113, 678–684. [Google Scholar] [CrossRef]
- Albert, P.R. Why is depression more prevalent in women? J. Psychiatry Neurosci. 2015, 40, 219. [Google Scholar] [CrossRef]
- Rubinow, D.R.; Schmidt, P.J. Is there a role for reproductive steroids in the etiology and treatment of affective disorders? Dialogues Clin. Neurosci. 2022, 20, 187–196. [Google Scholar] [CrossRef]
- Zhou, N.; Gu, X.; Zhuang, T.; Xu, Y.; Yang, L.; Zhou, M. Gut microbiota: A pivotal hub for polyphenols as antidepressants. J. Agric. Food Chem. 2020, 68, 6007–6020. [Google Scholar] [CrossRef] [PubMed]

| Structure | Examples of Compounds | Food Sources | ||
|---|---|---|---|---|
 Basic Skeleton | Flavonol |  | Quercetin, kaempferol, myricetin, and rutin | Olive oil, brocoli, apples, cherries, and berries |
| Flavone |  | Chrysin, apigenin, luteolin | Red pepper, tomatoes, carrot, and chamomile | |
| Flavanone |  | Naringenin, pinocembrin, and naringin | Orange, lemon, citrus, and grapefruit juice | |
| Flavanol |  | Catechin, epicatechin, epigallocatechin, and proanthocyanin | Grapefruit, lemon, apple, red wine, nuts, and tea | |
| Isoflavone |  | Genistein, Daidzein, and biochanin A | Soybeans and legumes | |
| Chalcone |  | Naringenin-chalcone and pinocembrin-chalcone | Soy, apple, citrus, and ginger | |
| Anthocyanidin |  | Cyanidin, delphinidin, pelargonidin, malvidin, and petunidin | Blackberries, cherries, strawberries, and chagalapoli |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
German-Ponciano, L.J.; Rosas-Sánchez, G.U.; Cueto-Escobedo, J.; Fernández-Demeneghi, R.; Guillén-Ruiz, G.; Soria-Fregozo, C.; Herrera-Huerta, E.V.; Rodríguez-Landa, J.F. Participation of the Serotonergic System and Brain-Derived Neurotrophic Factor in the Antidepressant-like Effect of Flavonoids. Int. J. Mol. Sci. 2022, 23, 10896. https://doi.org/10.3390/ijms231810896
German-Ponciano LJ, Rosas-Sánchez GU, Cueto-Escobedo J, Fernández-Demeneghi R, Guillén-Ruiz G, Soria-Fregozo C, Herrera-Huerta EV, Rodríguez-Landa JF. Participation of the Serotonergic System and Brain-Derived Neurotrophic Factor in the Antidepressant-like Effect of Flavonoids. International Journal of Molecular Sciences. 2022; 23(18):10896. https://doi.org/10.3390/ijms231810896
Chicago/Turabian StyleGerman-Ponciano, León Jesús, Gilberto Uriel Rosas-Sánchez, Jonathan Cueto-Escobedo, Rafael Fernández-Demeneghi, Gabriel Guillén-Ruiz, César Soria-Fregozo, Emma Virginia Herrera-Huerta, and Juan Francisco Rodríguez-Landa. 2022. "Participation of the Serotonergic System and Brain-Derived Neurotrophic Factor in the Antidepressant-like Effect of Flavonoids" International Journal of Molecular Sciences 23, no. 18: 10896. https://doi.org/10.3390/ijms231810896
APA StyleGerman-Ponciano, L. J., Rosas-Sánchez, G. U., Cueto-Escobedo, J., Fernández-Demeneghi, R., Guillén-Ruiz, G., Soria-Fregozo, C., Herrera-Huerta, E. V., & Rodríguez-Landa, J. F. (2022). Participation of the Serotonergic System and Brain-Derived Neurotrophic Factor in the Antidepressant-like Effect of Flavonoids. International Journal of Molecular Sciences, 23(18), 10896. https://doi.org/10.3390/ijms231810896