
Evaluation of Plant Ceramide Species-Induced Exosome Release from Neuronal Cells and Exosome Loading Using Deuterium Chemistry
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
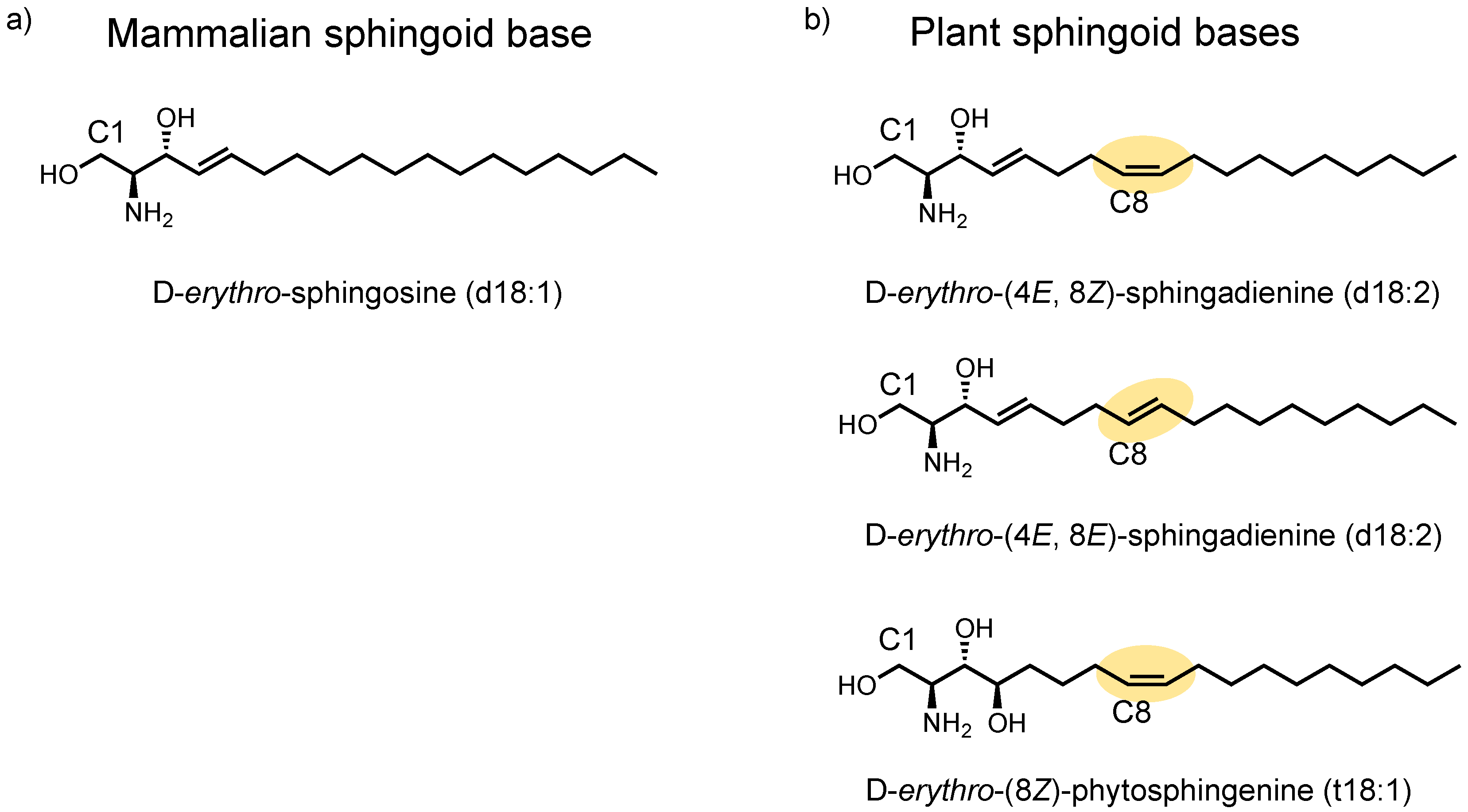
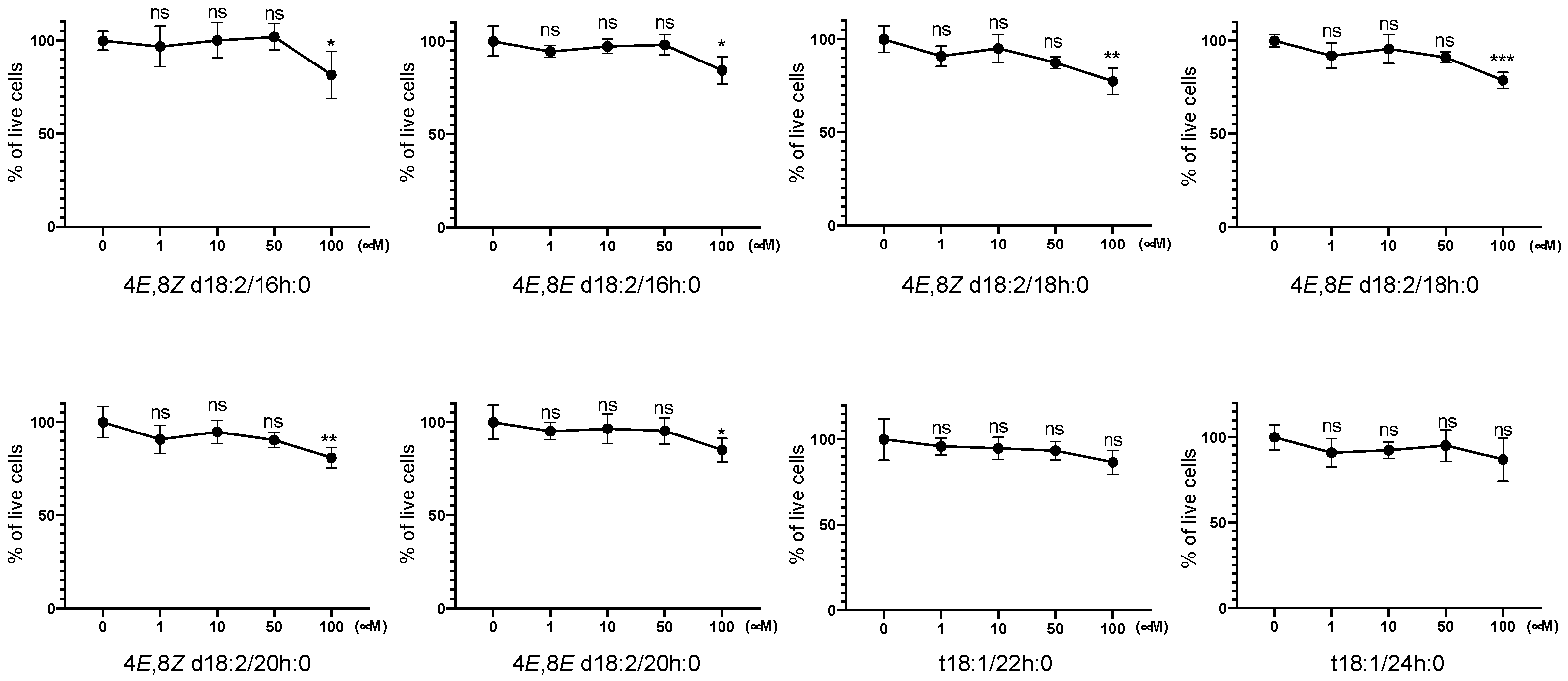
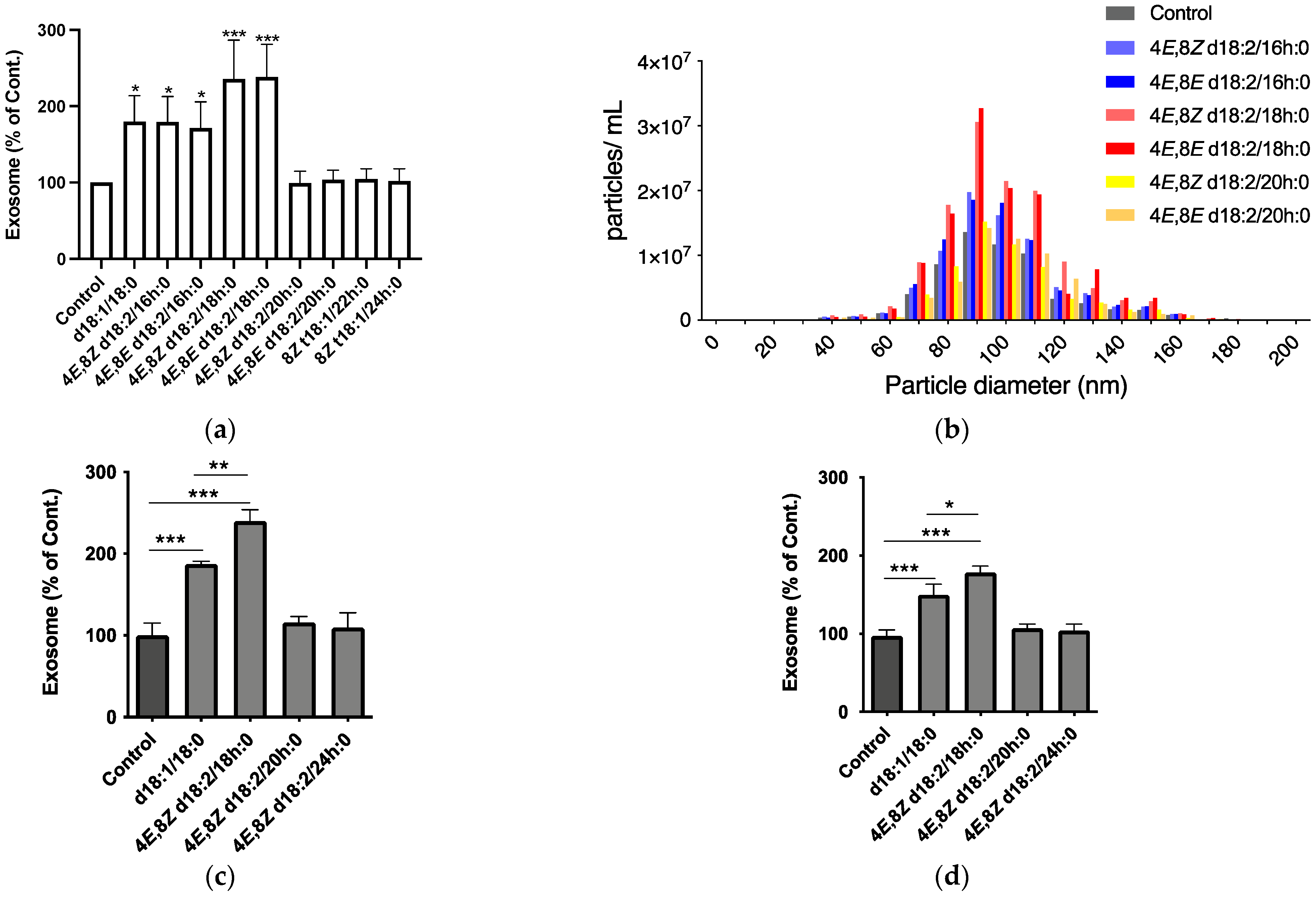
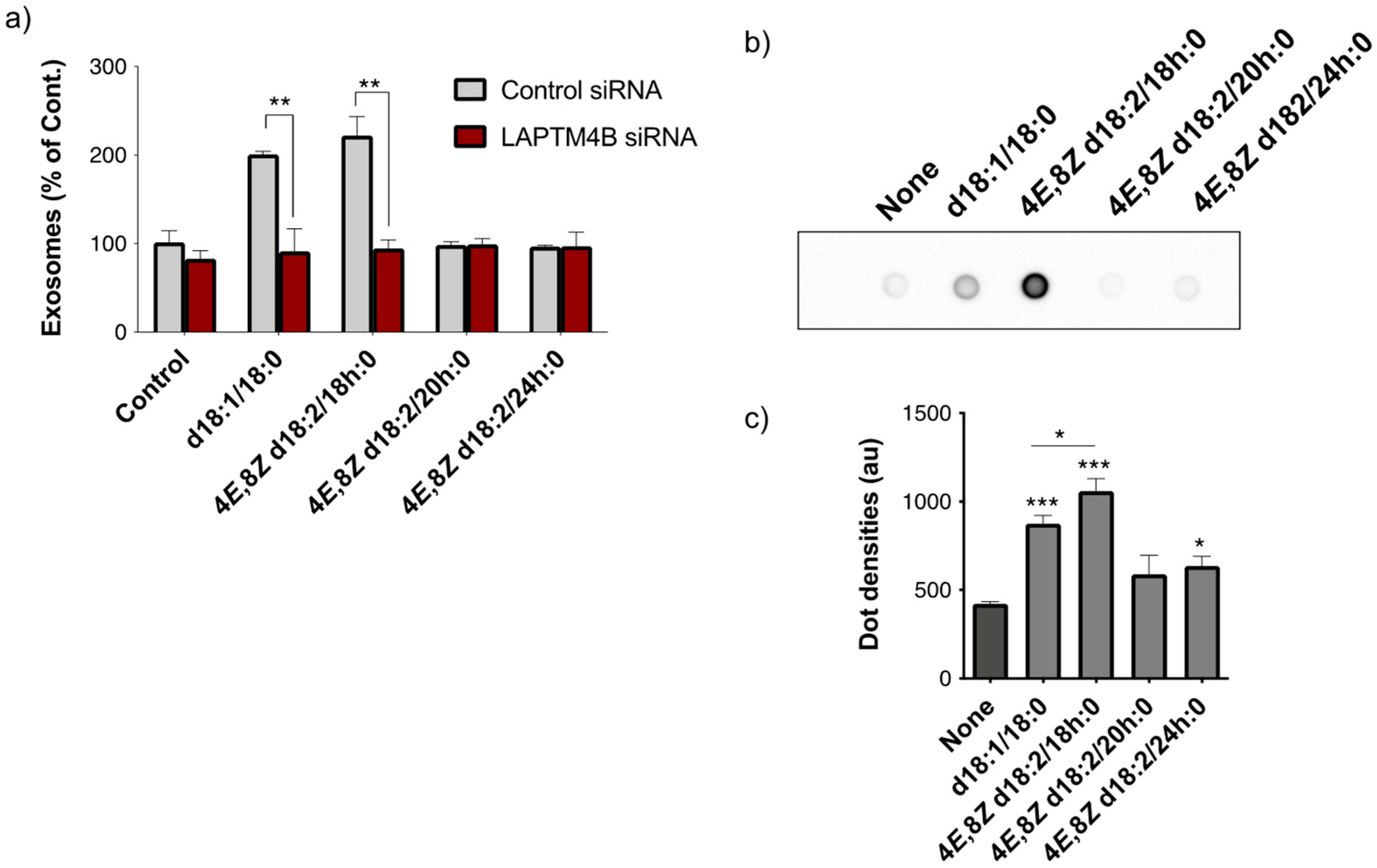
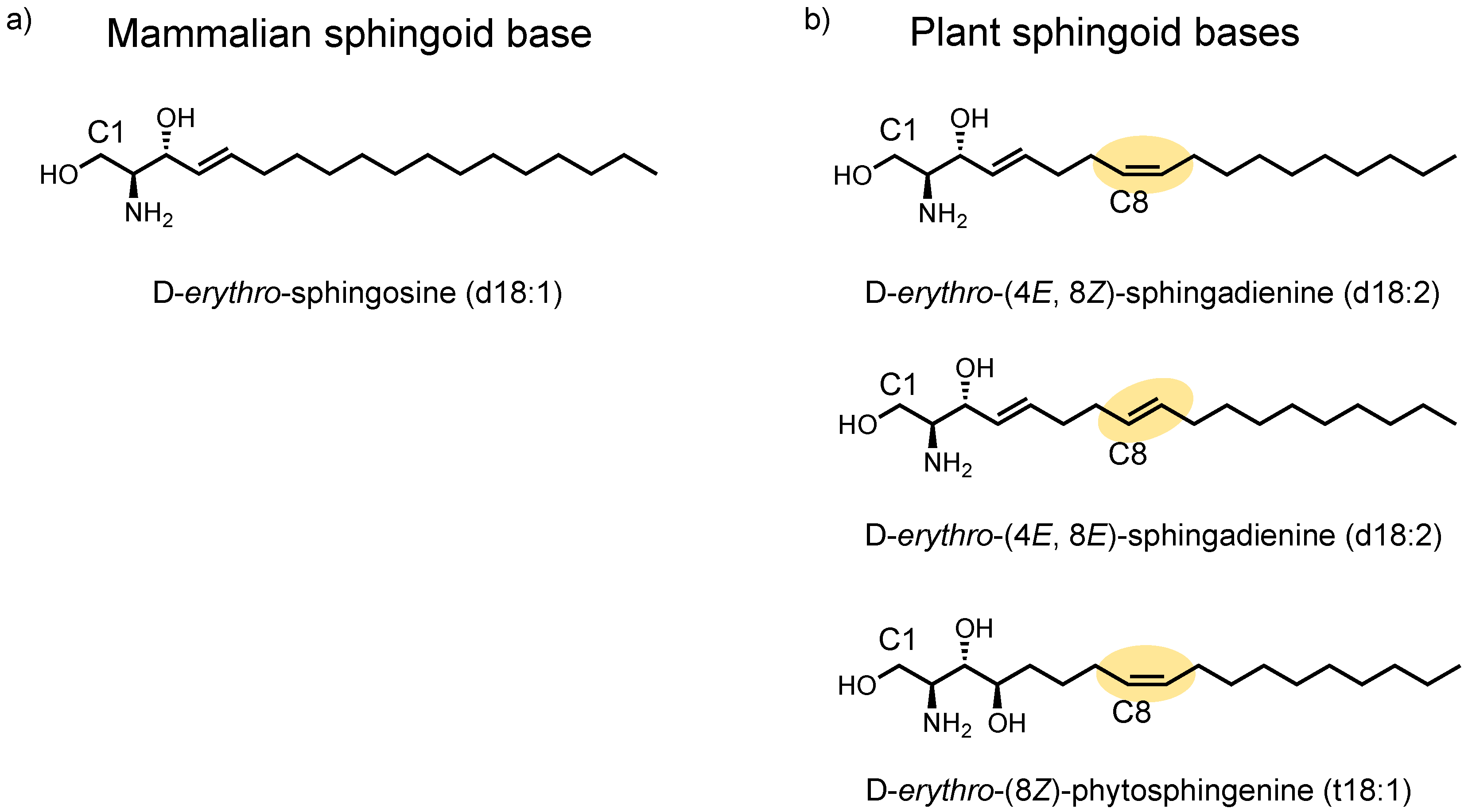
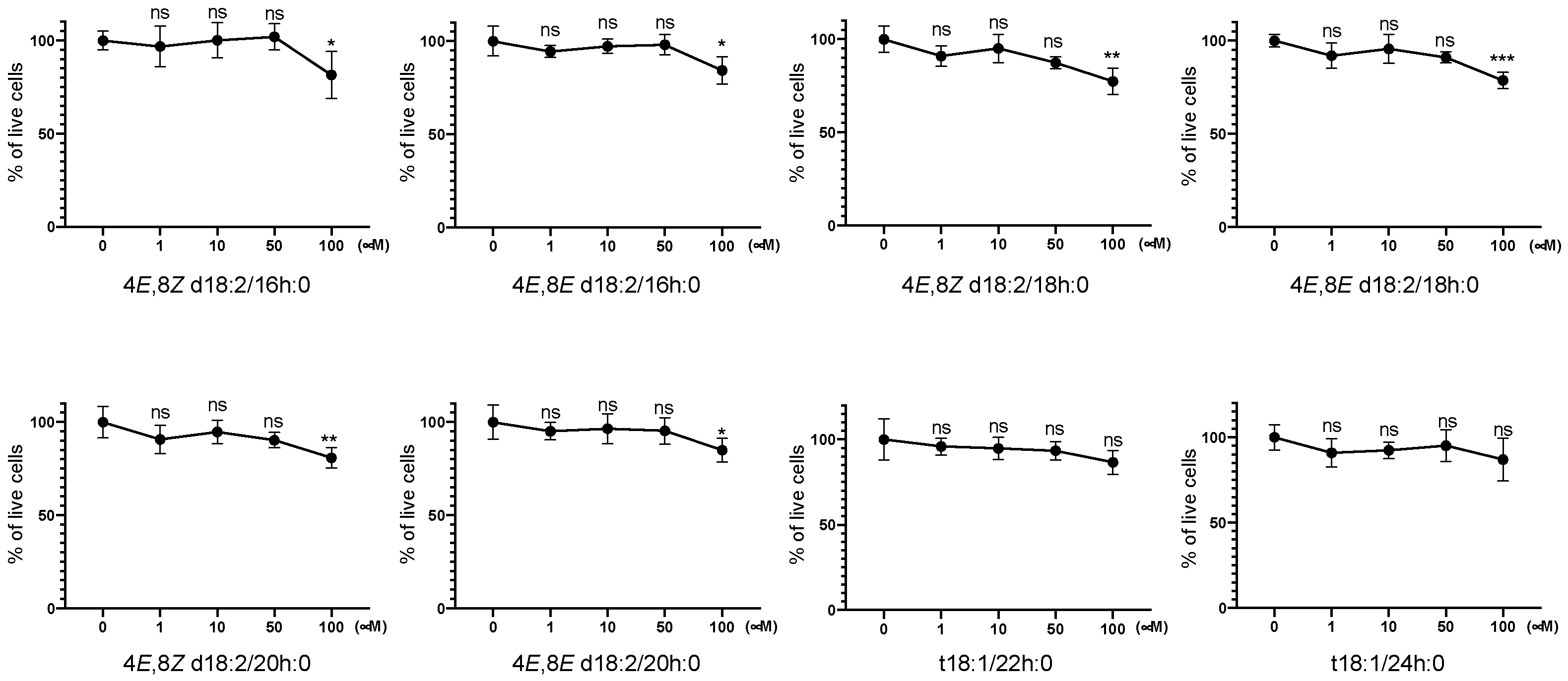
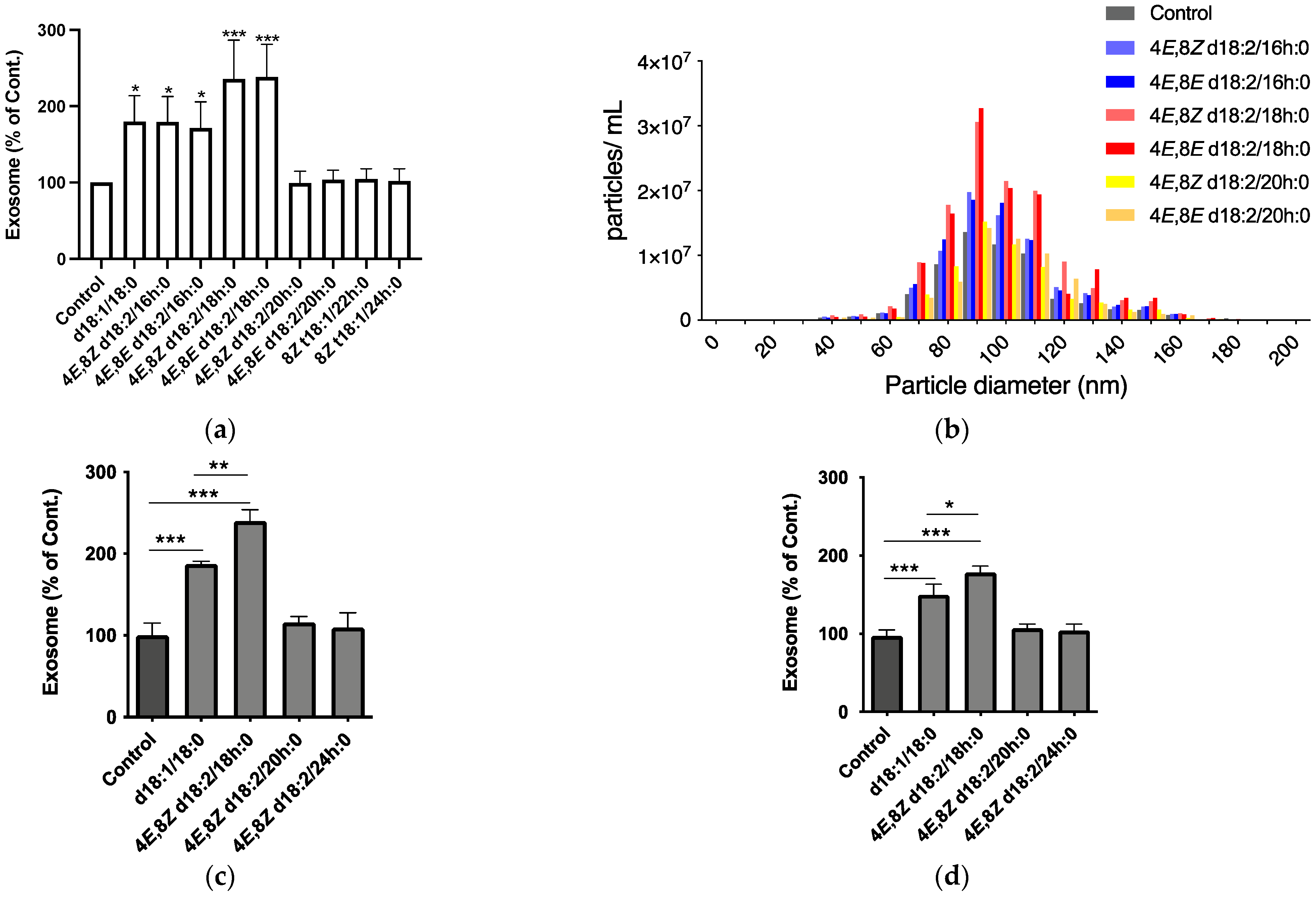
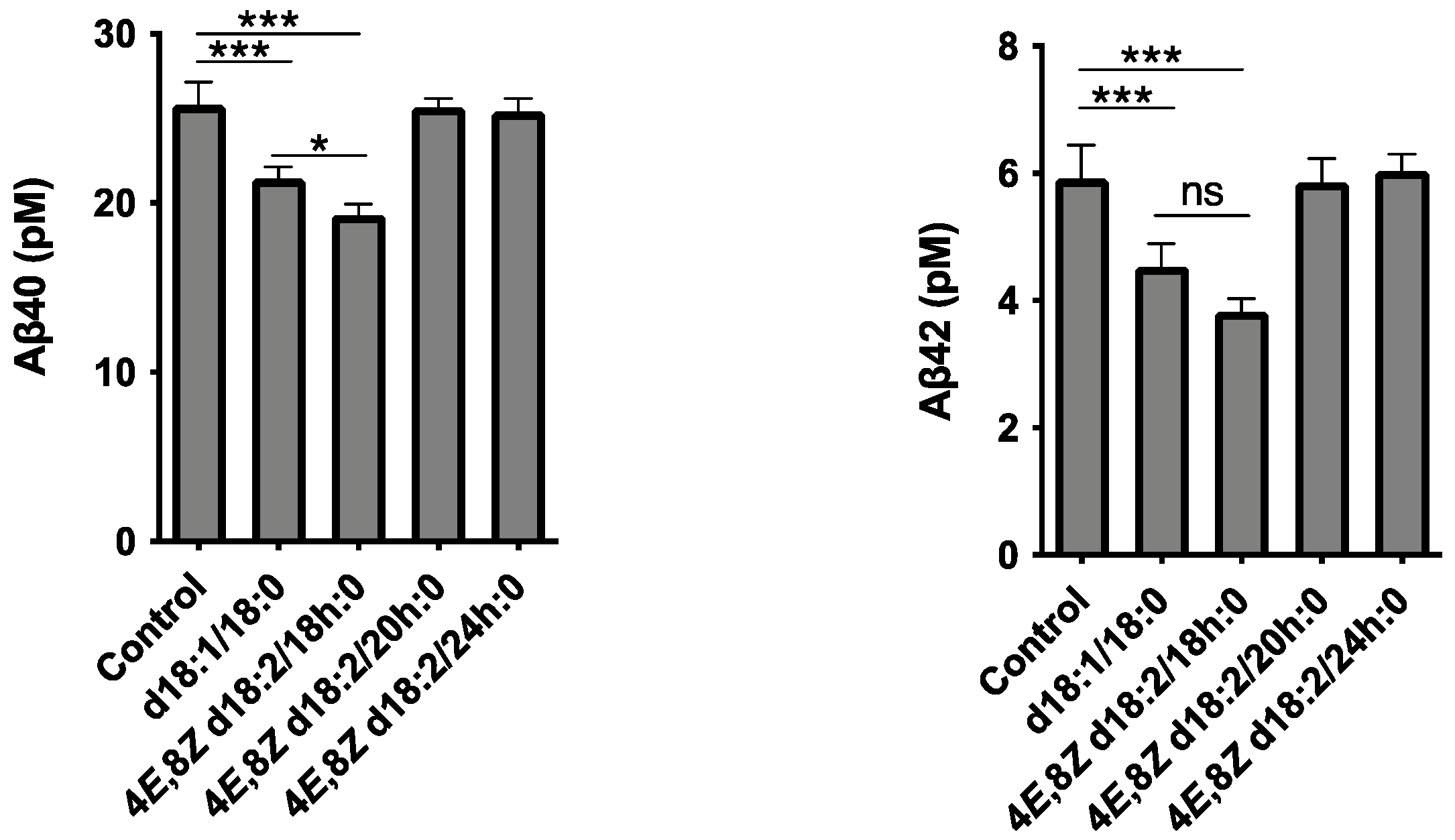
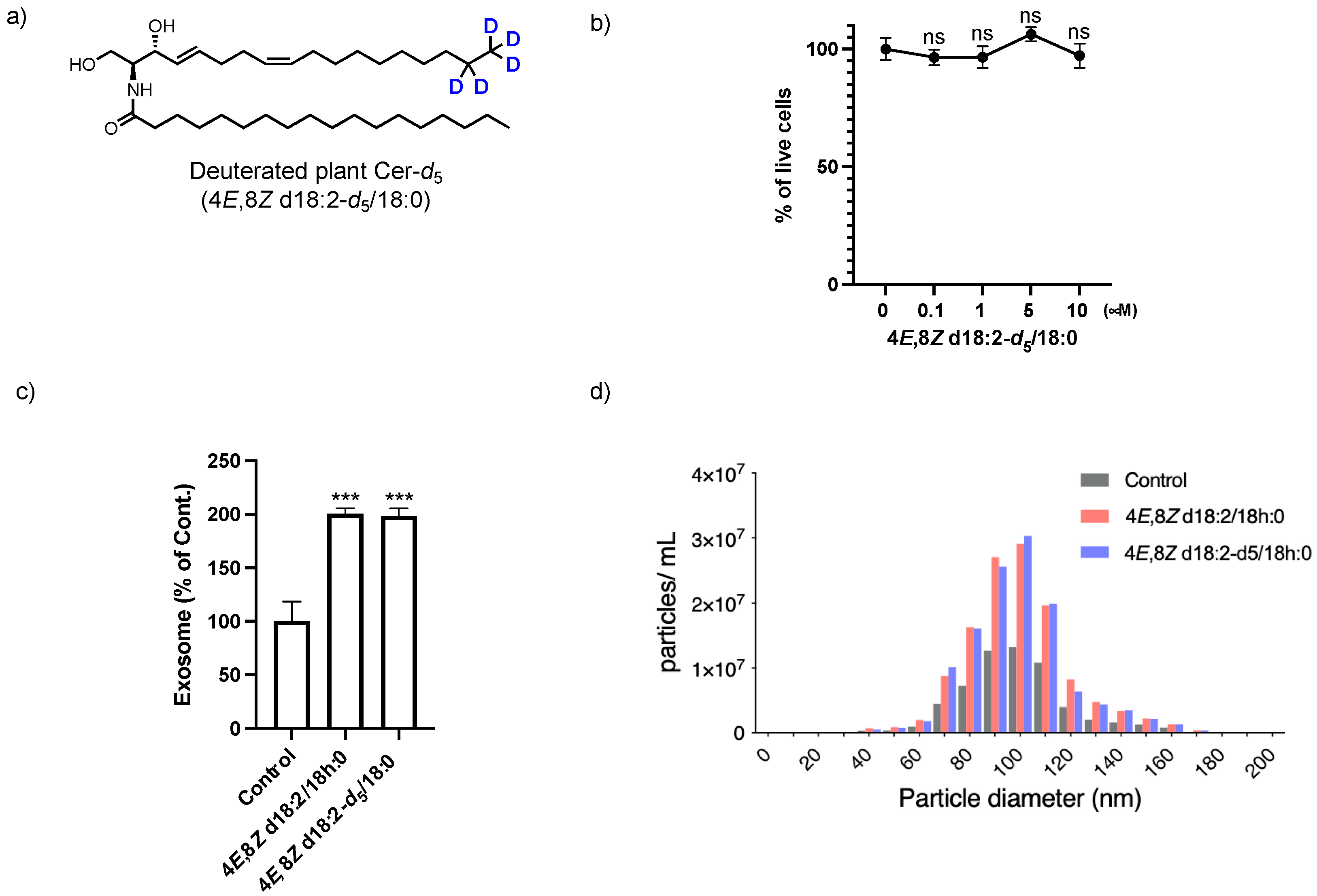
2.1. Identification of Plant Cer Types Promoting Exosome Release
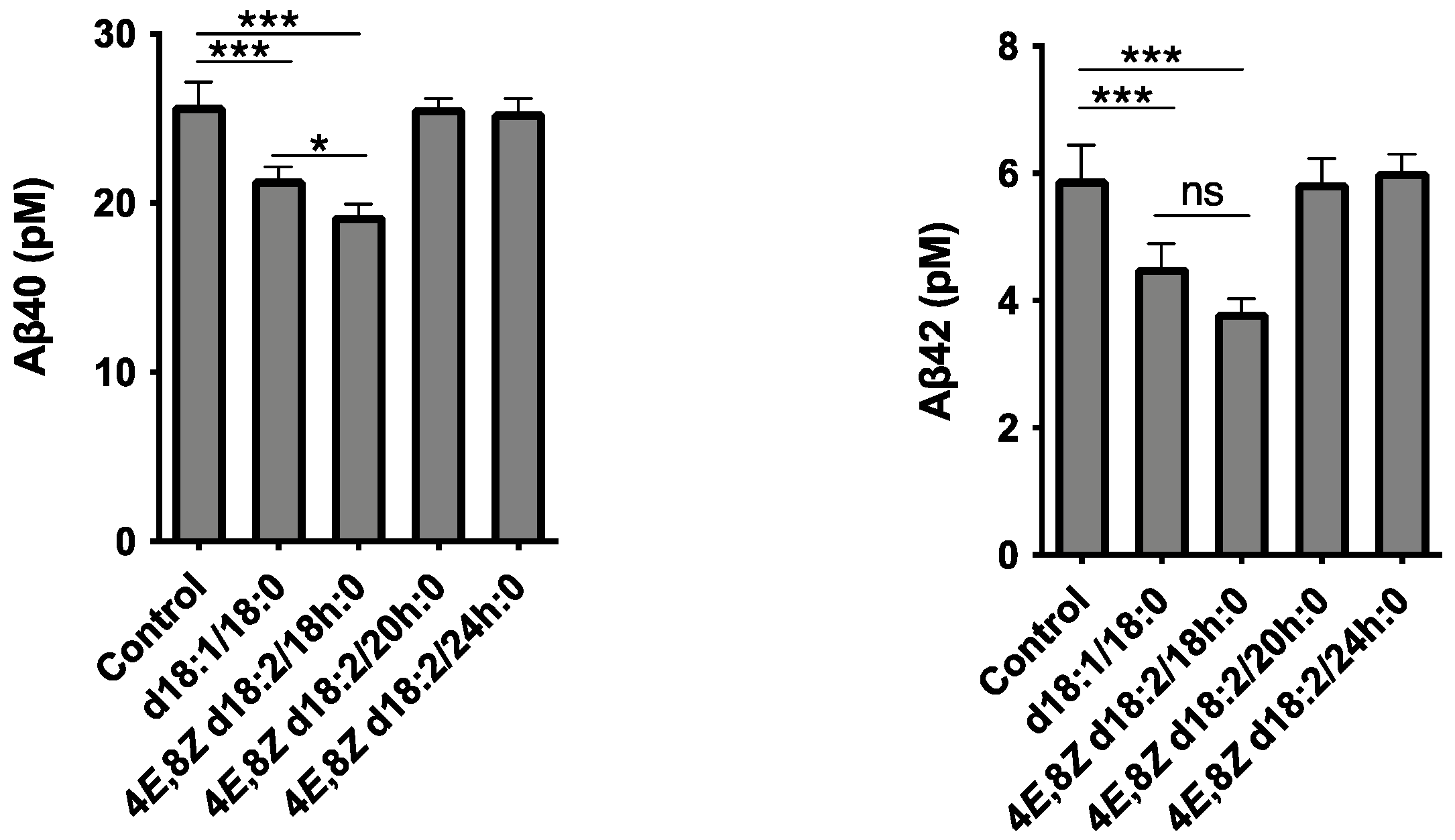
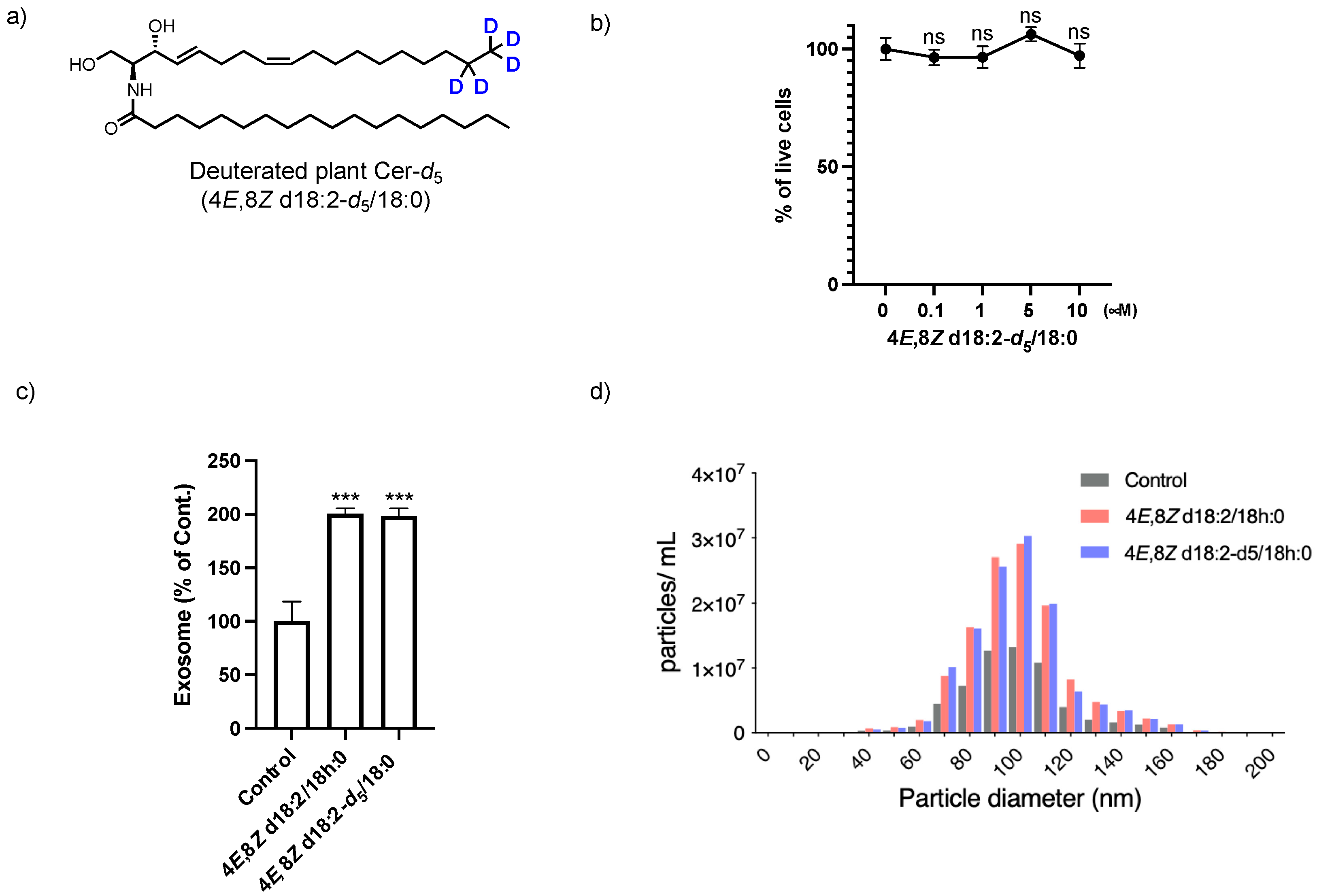
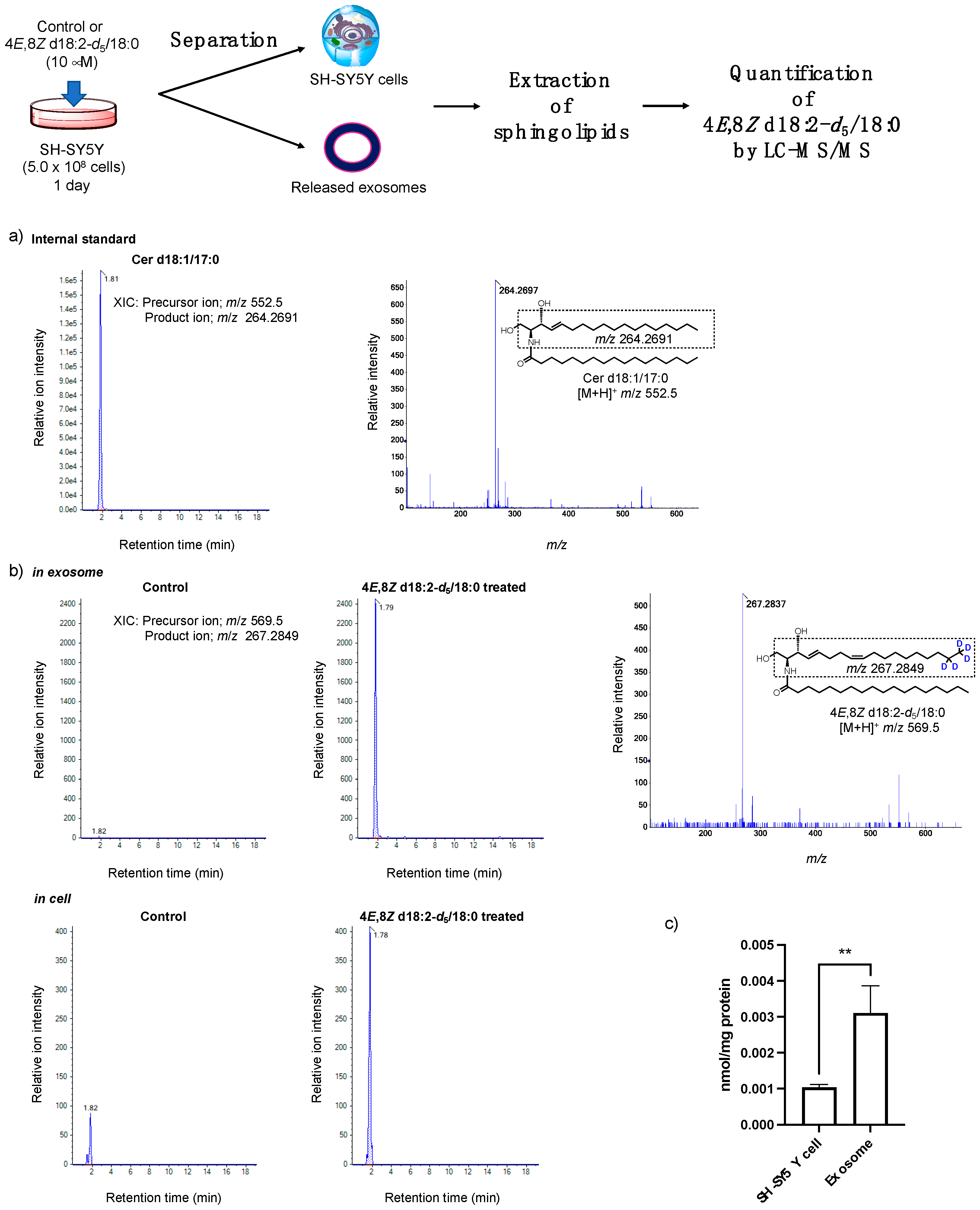
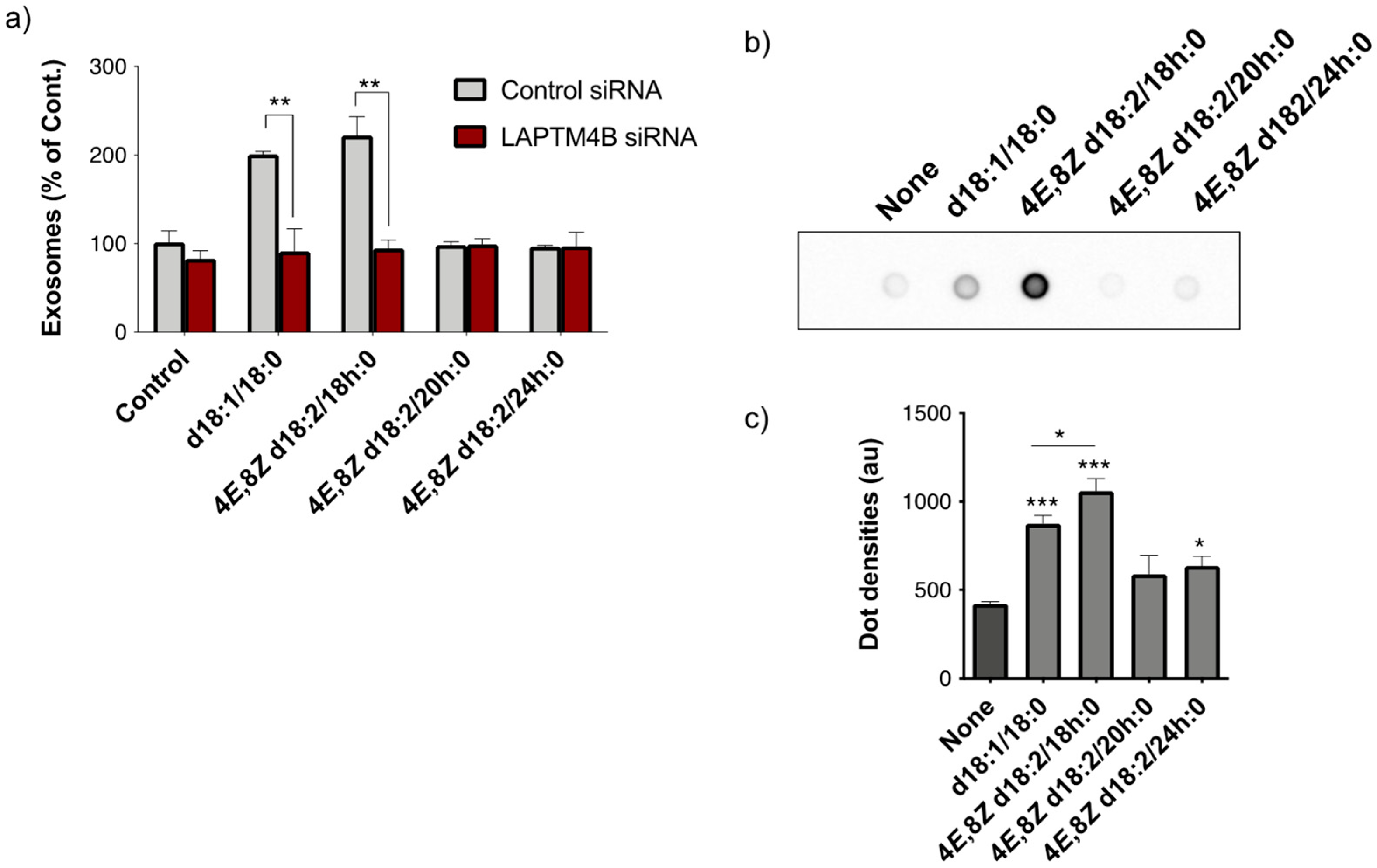
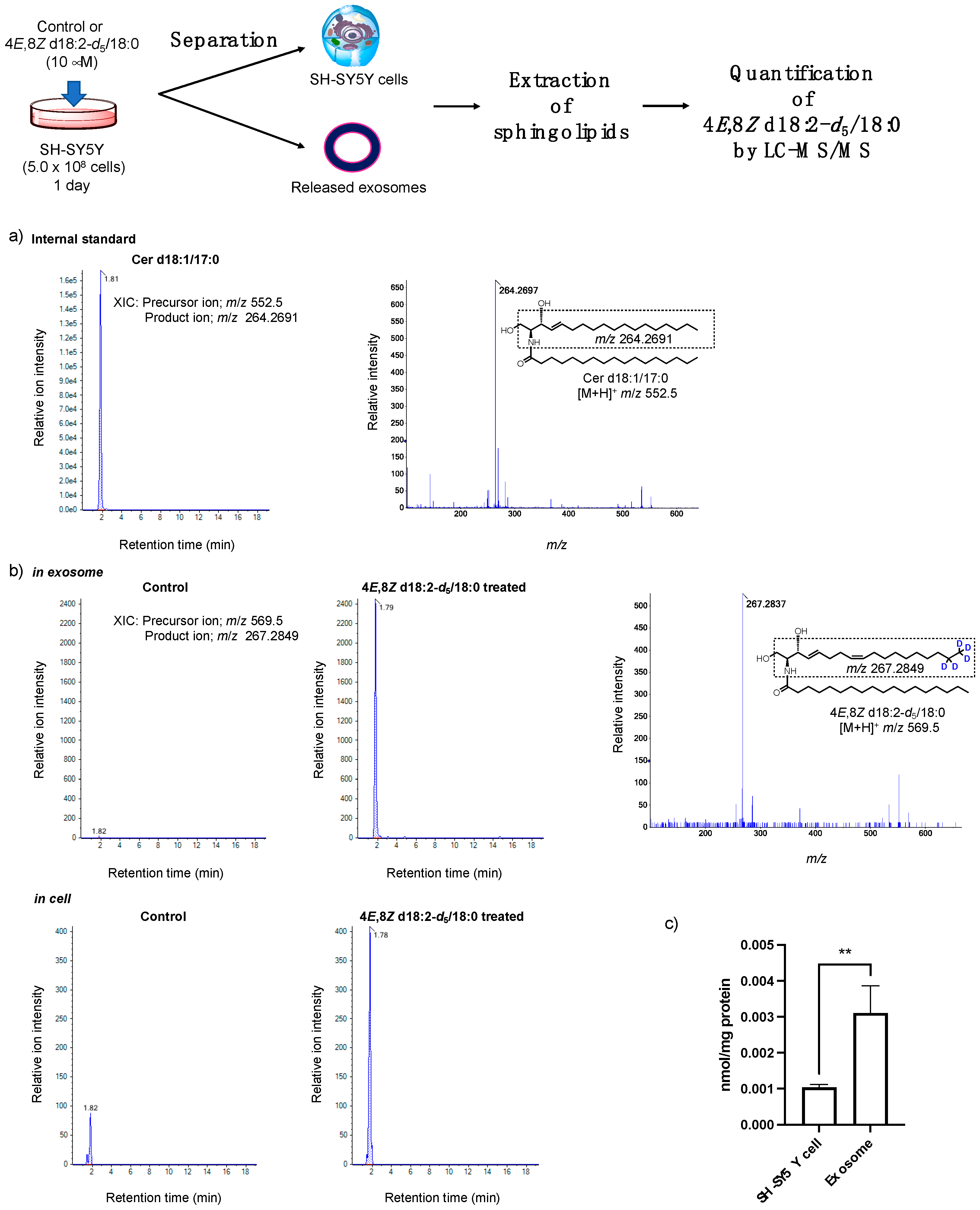
2.2. Analysis of the Loading of Exogenous Plant Cers into Exosomes Using Deuterium Chemistry and Mass Spectrometry
3. Discussion
4. Materials and Methods
4.1. Synthesis of N-((2S,3R,4E,8Z)-1,3-dihydroxyoctadeca-4,8-dien-2-yl-17,17,18,18,18-d5)stearamide (Deuterated Plant Cer-d5)
4.2. Preparation of Plant Cers
4.3. Cell Culture and Treatments
4.4. Exosome Collection and Quantification
4.5. Ceramide Overlay Assay
4.6. Deuterated Plant Cer-d5 Extraction and Quantification by LC-MS/MS
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Mawuenyega, K.G.; Sigurdson, W.; Ovod, V.; Munsell, L.; Kasten, T.; Morris, J.C.; Yarasheski, K.E.; Bateman, R.J. Decreased clearance of CNS beta-amyloid in Alzheimer’s disease. Science 2010, 330, 1774. [Google Scholar] [CrossRef] [PubMed]
- Sperling, R.A.; Aisen, P.S.; Beckett, L.A.; Bennett, D.A.; Craft, S.; Fagan, A.M.; Iwatsubo, T.; Jack, C.R., Jr.; Kaye, J.; Montine, T.J.; et al. Toward defining the preclinical stages of Alzheimer’s disease: Recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimer’s Dement. 2011, 7, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Martha, C.M.; Denis, A.E.; Julia, L.B.; Christine, C.T.; David, A.B.; Robert, S.W.; Neelum, A.; Julie, S. Consumption of fish and n-3 fatty acids and risk of incident Alzheimer disease. Arch Neurol. 2003, 60, 940–946. [Google Scholar]
- Yuyama, K.; Takahashi, K.; Usuki, S.; Mikami, D.; Sun, H.; Hanamatsu, H.; Furukawa, J.; Mukai, K.; Igarashi, Y. Plant sphingolipids promote extracellular vesicle release and alleviate amyloid-β pathologies in a mouse model of Alzheimer’s disease. Sci. Rep. 2019, 9, 16827. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, K.; Igarahsi, Y. Linking glycosphingolipids to Alzheimer’s amyloid-β: Extracellular vesicles and functional plant materials. Glycoconjugate J. 2022; in press. [Google Scholar]
- Yuyama, K.; Sun, H.; Mitsutake, S.; Igarashi, Y. Sphingolipid-modulated exosome secretion promotes clearance of amyloid-β by microglia. J. Biol. Chem. 2012, 287, 10977–10989. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, K.; Sun, H.; Sakai, S.; Mitsutake, S.; Okada, M.; Tahara, H.; Furukawa, J.; Fujitani, N.; Shinohara, Y.; Igarashi, Y. Decreased amyloid-β pathologies by intracerebral loading of glycosphingolipid-enriched exosomes in Alzheimer model mice. J. Biol. Chem. 2014, 289, 24488–24498. [Google Scholar] [CrossRef]
- Yuyama, K.; Sun, H.; Usuki, S.; Sakai, S.; Hanamatsu, H.; Mioka, T.; Kimura, N.; Okada, M.; Tahara, H.; Furukawa, J.; et al. A potential function for neuronal exosomes: Sequestering intracerebral amyloid-β peptide. FEBS Lett. 2015, 589, 84–88. [Google Scholar] [CrossRef]
- Yuyama, K.; Igarashi, Y. Exosomes as Carriers of Alzheimer’s Amyloid-β. Front. Neurosci. 2017, 11, 229. [Google Scholar] [CrossRef]
- Yuyama, K.; Sun, H.; Mikami, D.; Mioka, T.; Mukai, K.; Igarashi, Y. Lysosomal-associated transmembrane protein 4B regulates ceramide-induced exosome release. FASEB J. 2020, 34, 16022–16033. [Google Scholar] [CrossRef]
- Yunoki, K.; Ogawa, T.; Ono, J.; Miyashita, R.; Aida, K.; Oda, Y.; Ohnishi, M. Analysis of sphingolipid classes and their contents in meals. Biosci. Biotechnol. Biochem. 2008, 72, 222–225. [Google Scholar] [CrossRef]
- Usuki, S.; Tamura, N.; Sakai, S.; Tamura, T.; Mukai, K.; Igarashi, Y. Chemoenzymatically prepared konjac ceramide inhibits NGF-induced neurite outgrowth by a semaphoring 3A-like action. Biochem. Biophys. Rep. 2016, 5, 160–167. [Google Scholar] [PubMed] [Green Version]
- Nakai, W.; Yoshida, T.; Diez, D.; Miyatake, Y.; Nishibu, T.; Imawaka, N.; Naruse, K.; Sadamura, Y.; Hanayama, R. A novel affinity-based method for the isolation of highly purified extracellular vesicles. Sci. Rep. 2016, 6, 33935. [Google Scholar] [CrossRef] [PubMed]
- Murai, Y.; Yuyama, K.; Mikami, D.; Igarashi, Y.; Monde, K. Penta-deuterium-labeled 4E,8Z-sphingadienine for rapid analysis in sphingolipidomics study. Chem. Phys. Lipids 2022, 245, 105202. [Google Scholar] [CrossRef] [PubMed]
- Blom, T.; Li, S.; Dichlberger, A.; Bäck, N.; Kim, Y.A.; Loizides-Mangold, U.; Riezman, H.; Bittman, R.; Ikonen, E. LAPTM4B facilitates late endosomal ceramide export to control cell death pathways. Nat. Chem. Biol. 2015, 11, 799–806. [Google Scholar] [CrossRef]
- Zhou, K.; Dichlberger, A.; Martinez-Seara, H.; Nyholm, T.K.M.; Li, S.; Kim, Y.A.; Vattulainen, I.; Ikonen, E.; Blom, T. A ceramide-regulated element in the late endosomal protein LAPTM4B controls amino acid transporter interaction. ACS Central Sci. 2018, 4, 548–558. [Google Scholar] [CrossRef]
- Koolath, S.; Murai, Y.; Suga, Y.; Monde, K. Chiral combinatorial preparation and biological evaluation of unique ceramides for inhibition of sphingomyelin synthase. Chirality 2020, 32, 308–313. [Google Scholar] [CrossRef]
- Hogue, D.L.; Nash, C.; Ling, V.; Hobman, T.C. Lysosome-associated protein transmembrane 4 alpha (LAPTM4 alpha) requires two tandemly arranged tyrosine-based signals for sorting to lysosomes. Biochem. J. 2002, 365, 721–730. [Google Scholar] [CrossRef]
- Shao, G.Z.; Zhou, R.L.; Zhang, Q.Y.; Zhang, Y.; Liu, J.J.; Rui, J.A.; Wei, X.; Ye, D.X. Molecular cloning and characterization of LAPTM4B, a novel gene upregulated in hepatocellular carcinoma. Oncogene 2003, 22, 5060–5069. [Google Scholar] [CrossRef]
- Tsuji, K.; Mitsutake, S.; Ishikawa, J.; Takagi, Y.; Akiyama, M.; Shimizu, H.; Tomiyama, T.; Igarashi, Y. Dietary glucosylceramide improves skin barrier function in hairless mice. J. Dermatol. Sci. 2006, 44, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A. Metabolism of cerebroside in the intestinal tract of the rat. Biochim. Biophys. Acta 1969, 187, 113–121. [Google Scholar] [CrossRef]
- Mikami, D.; Sakai, S.; Nishimukai, M.; Yuyama, K.; Mukai, K.; Igarashi, Y. Structure-dependent absorption of atypical sphingoid long-chain bases from digestive tract into lymph. Lipids Health Dis. 2021, 20, 24. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, E.M.; Crall, K.J.; Larocque, R.; Dillehay, D.L.; Merrill, A.H. Uptake and metabolism of sphingolipids in isolated intestinal loops of mice. J. Nutr. 1994, 124, 702–712. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, J.; Takada, S.; Hashizume, K.; Takagi, Y.; Hotta, M.; Masukawa, Y.; Kitahara, T.; Mizutani, Y.; Igarashi, Y. Dietary glucosylceramide is absorbed into the lymph and increases levels of epidermal sphingolipids. J. Dermatol. Sci. 2009, 56, 220–222. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, K.; Mikami, D.; Sun, H.; Tsumita, T.; Takahashi, K.; Mukai, K.; Yuyama, K.; Igarashi, Y. Blood-brain barrier permeability analysis of plant ceramides. PLoS ONE 2020, 15, e0241640. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murai, Y.; Honda, T.; Yuyama, K.; Mikami, D.; Eguchi, K.; Ukawa, Y.; Usuki, S.; Igarashi, Y.; Monde, K. Evaluation of Plant Ceramide Species-Induced Exosome Release from Neuronal Cells and Exosome Loading Using Deuterium Chemistry. Int. J. Mol. Sci. 2022, 23, 10751. https://doi.org/10.3390/ijms231810751
Murai Y, Honda T, Yuyama K, Mikami D, Eguchi K, Ukawa Y, Usuki S, Igarashi Y, Monde K. Evaluation of Plant Ceramide Species-Induced Exosome Release from Neuronal Cells and Exosome Loading Using Deuterium Chemistry. International Journal of Molecular Sciences. 2022; 23(18):10751. https://doi.org/10.3390/ijms231810751
Chicago/Turabian StyleMurai, Yuta, Takumi Honda, Kohei Yuyama, Daisuke Mikami, Koichi Eguchi, Yuichi Ukawa, Seigo Usuki, Yasuyuki Igarashi, and Kenji Monde. 2022. "Evaluation of Plant Ceramide Species-Induced Exosome Release from Neuronal Cells and Exosome Loading Using Deuterium Chemistry" International Journal of Molecular Sciences 23, no. 18: 10751. https://doi.org/10.3390/ijms231810751
APA StyleMurai, Y., Honda, T., Yuyama, K., Mikami, D., Eguchi, K., Ukawa, Y., Usuki, S., Igarashi, Y., & Monde, K. (2022). Evaluation of Plant Ceramide Species-Induced Exosome Release from Neuronal Cells and Exosome Loading Using Deuterium Chemistry. International Journal of Molecular Sciences, 23(18), 10751. https://doi.org/10.3390/ijms231810751