
Gut Dysbiosis and Fecal Microbiota Transplantation in Autoimmune Diseases

Abstract
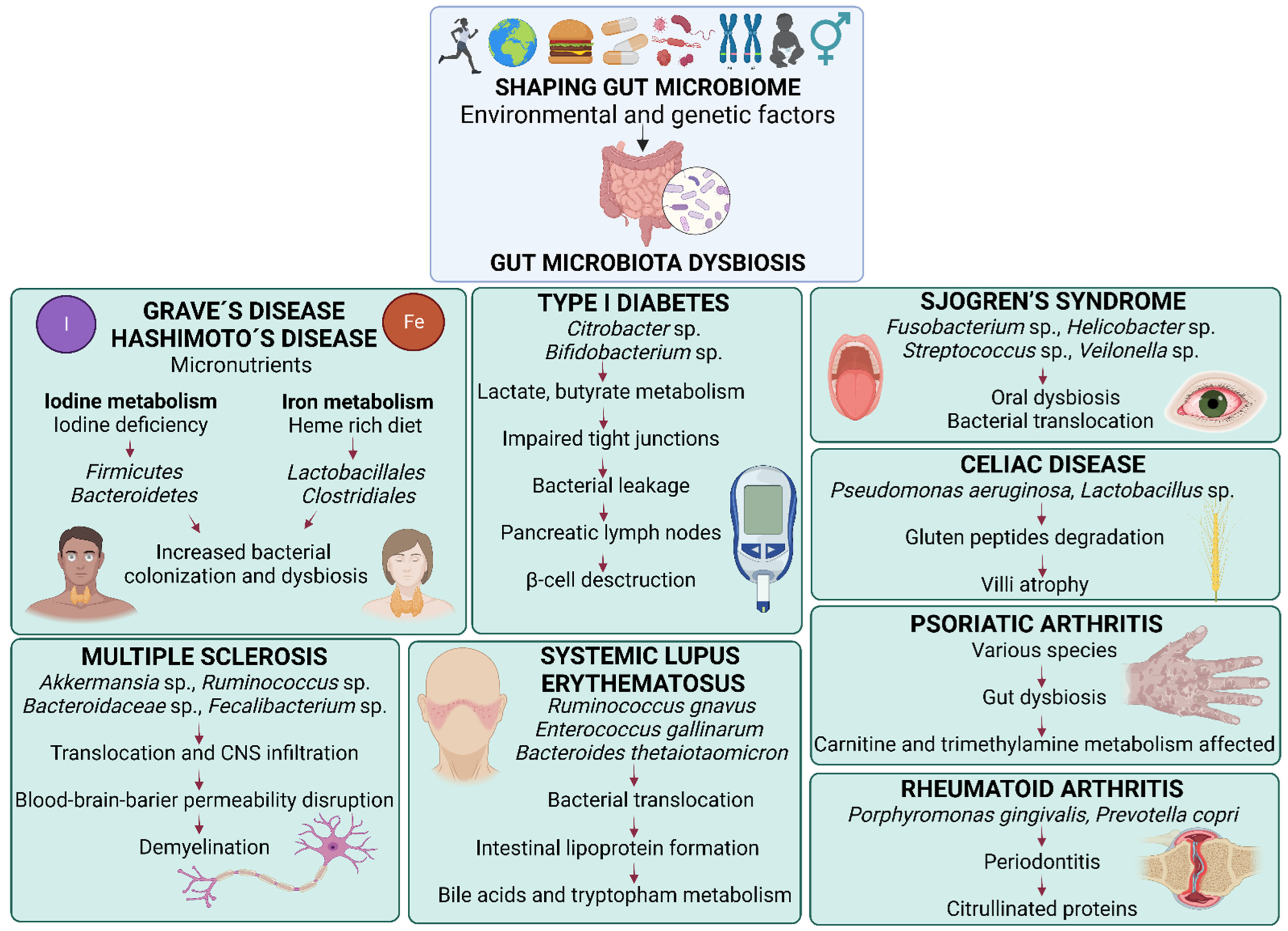
:1. Gut Microbiota and Fecal Microbiota Transplantation
2. Rheumatoid Arthritis
3. Psoriatic Arthritis
4. Multiple Sclerosis
5. Type 1 Diabetes
6. Systemic Lupus Erythematosus
7. Celiac Disease
8. Hashimoto’s Disease
9. Graves’ Disease
10. Sjogren’s Syndrome
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, Y.; Chen, Q.; Lin, P.; Xu, R.; He, D.; Ji, W.; Bian, Y.; Shen, Y.; Li, Q.; Liu, C.; et al. Characteristics of Gut Microbiota in Patients with Rheumatoid Arthritis in Shanghai, China. Front. Cell. Infect. Microbiol. 2019, 9, 369. [Google Scholar] [CrossRef] [PubMed]
- Scher, J.U.; Ubeda, C.; Artacho, A.; Attur, M.; Isaac, S.; Reddy, S.M.; Marmon, S.; Neimann, A.; Brusca, S.; Patel, T.; et al. Decreased bacterial diversity characterizes the altered gut microbiota in patients with psoriatic arthritis, resembling dysbiosis in inflammatory bowel disease. Arthritis Rheumatol. Hoboken NJ 2015, 67, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Cantoni, C.; Lin, Q.; Dorsett, Y.; Ghezzi, L.; Liu, Z.; Pan, Y.; Chen, K.; Han, Y.; Li, Z.; Xiao, H.; et al. Alterations of host-gut microbiome interactions in multiple sclerosis. eBioMedicine 2022, 76, 103798. [Google Scholar] [CrossRef] [PubMed]
- Mejía-León, M.E.; Petrosino, J.F.; Ajami, N.J.; Domínguez-Bello, M.G.; De La Barca, A.M.C. Fecal microbiota imbalance in Mexican children with type 1 diabetes. Sci. Rep. 2014, 4, 3814. [Google Scholar] [CrossRef]
- Luo, X.M.; Edwards, M.R.; Mu, Q.; Yu, Y.; Vieson, M.D.; Reilly, C.M.; Ahmed, S.A.; Bankole, A.A. Gut Microbiota in Human Systemic Lupus Erythematosus and a Mouse Model of Lupus. Appl. Environ. Microbiol. 2018, 84, e02288-17. [Google Scholar] [CrossRef]
- Collado, M.C.; Calabuig, M.; Sanz, Y. Differences between the fecal microbiota of coeliac infants and healthy controls. Curr. Issues Intest. Microbiol. 2007, 8, 9–14. [Google Scholar]
- Liu, S.; An, Y.; Cao, B.; Sun, R.; Ke, J.; Zhao, D. The Composition of Gut Microbiota in Patients Bearing Hashimoto’s Thyroiditis with Euthyroidism and Hypothyroidism. Int. J. Endocrinol. 2020, 2020, e5036959. [Google Scholar] [CrossRef]
- Zhu, Q.; Hou, Q.; Huang, S.; Ou, Q.; Huo, D.; Vázquez-Baeza, Y.; Cen, C.; Cantu, V.; Estaki, M.; Chang, H.; et al. Compositional and genetic alterations in Graves’ disease gut microbiome reveal specific diagnostic biomarkers. ISME J. 2021, 15, 3399–3411. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Lu, H.; Chen, Y.; Wang, Q.; Cao, H.; Li, T. Changes of Intestinal Microecology in Patients with Primary Sjogren’s Syndrome after Therapy of Yangyin Yiqi Huoxue Recipe (养阴益气活血方). Chin. J. Integr. Med. 2019, 25, 654–662. [Google Scholar] [CrossRef]
- Liptak, R.; Gromova, B.; Maronek, M.; Gardlik, R. Reverse phenotype transfer via fecal microbial transplantation in inflammatory bowel disease. Med. Hypotheses 2019, 122, 41–44. [Google Scholar] [CrossRef]
- Zeng, J.; Peng, L.; Zheng, W.; Huang, F.; Zhang, N.; Wu, D.; Yang, Y. Fecal microbiota transplantation for rheumatoid arthritis: A case report. Clin. Case Rep. 2021, 9, 906–909. [Google Scholar] [CrossRef] [PubMed]
- Kragsnaes, M.S.; Sødergren, S.T.; Kjeldsen, J.; Horn, H.C.; Munk, H.L.; Pedersen, J.K.; Klinkby, C.S.; de Wit, M.; Ahlmark, N.G.; Tjørnhøj-Thomsen, T.; et al. Experiences and perceptions of patients with psoriatic arthritis participating in a trial of faecal microbiota transplantation: A nested qualitative study. BMJ Open 2021, 11, e039471. [Google Scholar] [CrossRef] [PubMed]
- Al, K.F.; Craven, L.J.; Gibbons, S.; Parvathy, S.N.; Wing, A.C.; Graf, C.; Parham, K.A.; Kerfoot, S.M.; Wilcox, H.; Burton, J.P.; et al. Fecal microbiota transplantation is safe and tolerable in patients with multiple sclerosis: A pilot randomized controlled trial. Mult. Scler. J.-Exp. Transl. Clin. 2022, 8, 20552173221086664. [Google Scholar] [CrossRef]
- de Groot, P.F.; Frissen, M.N.; de Clercq, N.C.; Nieuwdorp, M. Fecal microbiota transplantation in metabolic syndrome: History, present and future. Gut Microbes 2017, 8, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Eiseman, B.; Silen, W.; Bascom, G.S.; Kauvar, A.J. Fecal enema as an adjunct in the treatment of pseudomembranous enterocolitis. Surgery 1958, 44, 854–859. [Google Scholar] [PubMed]
- Hvas, C.L.; Dahl Jørgensen, S.M.; Jørgensen, S.P.; Storgaard, M.; Lemming, L.; Hansen, M.M.; Erikstrup, C.; Dahlerup, J.F. Fecal Microbiota Transplantation Is Superior to Fidaxomicin for Treatment of Recurrent Clostridium difficile Infection. Gastroenterology 2019, 156, 1324–1332.e3. [Google Scholar] [CrossRef] [PubMed]
- Friedman-Korn, T.; Livovsky, D.M.; Maharshak, N.; Aviv Cohen, N.; Paz, K.; Bar-Gil Shitrit, A.; Goldin, E.; Koslowsky, B. Fecal Transplantation for Treatment of Clostridium Difficile Infection in Elderly and Debilitated Patients. Dig. Dis. Sci. 2018, 63, 198–203. [Google Scholar] [CrossRef]
- Ianiro, G.; Bibbò, S.; Porcari, S.; Settanni, C.R.; Giambò, F.; Curta, A.R.; Quaranta, G.; Scaldaferri, F.; Masucci, L.; Sanguinetti, M.; et al. Fecal microbiota transplantation for recurrent C. difficile infection in patients with inflammatory bowel disease: Experience of a large-volume European FMT center. Gut Microbes 2021, 13, 1994834. [Google Scholar] [CrossRef]
- Sędzikowska, A.; Szablewski, L. Human Gut Microbiota in Health and Selected Cancers. Int. J. Mol. Sci. 2021, 22, 13440. [Google Scholar] [CrossRef]
- Liptak, R.; Gromova, B.; Gardlik, R. Fecal Microbiota Transplantation as a Tool for Therapeutic Modulation of Non-gastrointestinal Disorders. Front. Med. 2021, 8, 665520. [Google Scholar] [CrossRef]
- Paray, B.A.; Albeshr, M.F.; Jan, A.T.; Rather, I.A. Leaky Gut and Autoimmunity: An Intricate Balance in Individuals Health and the Diseased State. Int. J. Mol. Sci. 2020, 21, 9770. [Google Scholar] [CrossRef] [PubMed]
- Stewart, L.; Edgar, J.D.M.; Blakely, G.; Patrick, S. Antigenic mimicry of ubiquitin by the gut bacterium Bacteroides fragilis: A potential link with autoimmune disease. Clin. Exp. Immunol. 2018, 194, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Selmi, C.; Tang, R.; Gershwin, M.E.; Ma, X. The microbiome and autoimmunity: A paradigm from the gut–liver axis. Cell. Mol. Immunol. 2018, 15, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Zheng, L.; Qing, P.; Zhao, H.; Li, Y.; Su, L.; Zhang, Q.; Zhao, Y.; Luo, Y.; Liu, Y. Oral Microbiota Perturbations Are Linked to High Risk for Rheumatoid Arthritis. Front. Cell. Infect. Microbiol. 2020, 9, 475. [Google Scholar] [CrossRef] [PubMed]
- van Beurden, Y.H.; van Gils, T.; van Gils, N.A.; Kassam, Z.; Mulder, C.J.J.; Aparicio-Pagés, N. Serendipity in Refractory Celiac Disease: Full Recovery of Duodenal Villi and Clinical Symptoms after Fecal Microbiota Transfer. J. Gastrointest. Liver Dis. JGLD 2016, 25, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Engen, P.A.; Zaferiou, A.; Rasmussen, H.; Naqib, A.; Green, S.J.; Fogg, L.F.; Forsyth, C.B.; Raeisi, S.; Hamaker, B.; Keshavarzian, A. Single-Arm, Non-randomized, Time Series, Single-Subject Study of Fecal Microbiota Transplantation in Multiple Sclerosis. Front. Neurol. 2020, 11, 978. [Google Scholar] [CrossRef]
- Kragsnaes, M.S.; Kjeldsen, J.; Horn, H.C.; Munk, H.L.; Pedersen, J.K.; Just, S.A.; Ahlquist, P.; Pedersen, F.M.; de Wit, M.; Möller, S.; et al. Safety and efficacy of faecal microbiota transplantation for active peripheral psoriatic arthritis: An exploratory randomised placebo-controlled trial. Ann. Rheum. Dis. 2021, 80, 1158–1167. [Google Scholar] [CrossRef]
- Watane, A.; Cavuoto, K.M.; Rojas, M.; Dermer, H.; Day, J.O.; Banerjee, S.; Galor, A. Fecal Microbial Transplant in Individuals with Immune-Mediated Dry Eye. Am. J. Ophthalmol. 2022, 233, 90–100. [Google Scholar] [CrossRef]
- De Groot, P.; Nikolic, T.; Pellegrini, S.; Sordi, V.; Imangaliyev, S.; Rampanelli, E.; Hanssen, N.; Attaye, I.; Bakker, G.; Duinkerken, G.; et al. Faecal microbiota transplantation halts progression of human new-onset type 1 diabetes in a randomised controlled trial. Gut 2021, 70, 92–105. [Google Scholar] [CrossRef]
- Keshavarzian, A. Single-Arm, Non-Randomized, Time Series, Single-Subject Study: Fecal Microbiota Transplantation (FMT) in Multiple Sclerosis NCT03975413. 2020. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- Gelfand, J. Fecal Microbiota Transplantation (FMT) of FMP30 in Relapsing-Remitting Multiple Sclerosis: A Phase 1b Clinical Trial to Evaluate Feasibility, Safety, Tolerability and Effects on Immune Function NCT03594487. 2022. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- Moiseev, I.S. Allogeneic Fecal Microbiota Transplantation as a Consolidation Treatment After Autologous Hematopoietic Stem Cell Transplantation in Patients with Multiple Sclerosis NCT04203017. 2022. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- Lawson Health Research Institute. Fecal Microbial Transplantation in Relapsing Multiple Sclerosis Patients NCT03183869. 2019. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- Griffin Hospital. A Pilot Study of Oral FMT (Fecal Microbial Transplant) in Subjects with Multiple Sclerosis NCT04096443. 2022. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- MD, M.S. Fecal Microbial Transplantation for Relapsing Multiple Sclerosis Patients—A Placebo-Controlled, Double-Blinded, Randomized Trial NCT04150549. 2020. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- NG, S.C. Safety and Efficacy of Fecal Microbiota Transplantation: A Pilot Study NCT04014413. 2022. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- Ellingsen, T. Efficacy and Safety of Fecal Microbiota Transplantation (FMT) in Patients with Peripheral Psoriatic Arthritis: A 6-month, Double-Blind, Randomized, Placebo-Controlled Trial NCT03058900. 2020. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- Zhang, X. Efficacy and Safety of Faecal Microbiota Transplantation in Patients with Rheumatoid Arthritis Refractory to Methotrexate: A 24-Week, Double-Blind, Randomised Trial NCT03944096. 2019. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- Ellingsen, T. Safety and Clinical Efficacy Associated with Faecal Microbiota Transplantation Performed in Treatment-Naïve Patients with Newly Diagnosed Rheumatoid Arthritis, Reactive Arthritis, Ankylosing Spondylitis, Psoriatic Arthritis, Gouty Arthritis, Psoriasis, Hidradenitis Suppurativa, Pulmonary Sarcoidosis, Crohn’s Disease, and Ulcerative Colitis: A 52-Week, Double-Blind, Randomised, Placebo-Controlled, Exploratory Trial NCT04924270. 2021. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- Galor, A. Fecal Microbial Transplant for Sjogrens Syndrome NCT03926286. 2020. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- University of Aarhus. Faecal Microbiota Transplantation for Patients with Diabetes Mellitus Type 1 and Severe Gastrointestinal Neuropathy: A Randomised, Double-Blinded Safety and Pilot-Efficacy Study NCT04749030. 2022. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- Hanssen, N. Encapsulated Faecal Microbiota Transplantation to Preserve Residual Beta Cell Function in Patients with Recently-Diagnosed Type 1 Diabetes Mellitus NCT05323162. 2022. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- Shen, J. Fecal Microbiome Transplantation (FMT) for Type 1 Diabetes NCT04124211. 2019. Available online: https://clinicaltrials.gov/ (accessed on 7 September 2022).
- Cignarella, F.; Cantoni, C.; Ghezzi, L.; Salter, A.; Dorsett, Y.; Chen, L.; Fontana, L.; Weinstock, G.M.; Cross, A.H.; Zhou, Y.; et al. Intermittent fasting confers protection in CNS autoimmunity by altering the gut microbiota. Cell Metab. 2018, 27, 1222–1235.e6. [Google Scholar] [CrossRef]
- Li, K.; Wei, S.; Hu, L.; Yin, X.; Mai, Y.; Jiang, C.; Peng, X.; Cao, X.; Huang, Z.; Zhou, H.; et al. Protection of Fecal Microbiota Transplantation in a Mouse Model of Multiple Sclerosis. Mediators Inflamm. 2020, 2020, e2058272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wan, H.; Han, J.; Sun, X.; Yu, R.; Liu, B.; Lu, C.; Zhou, J.; Su, X. Ameliorative effect of tuna elastin peptides on AIA mice by regulating the composition of intestinal microorganisms and SCFAs. J. Funct. Foods 2022, 92, 105076. [Google Scholar] [CrossRef]
- Wang, C.; Zaheer, M.; Bian, F.; Quach, D.; Swennes, A.G.; Britton, R.A.; Pflugfelder, S.C.; de Paiva, C.S. Sjögren-Like Lacrimal Keratoconjunctivitis in Germ-Free Mice. Int. J. Mol. Sci. 2018, 19, 565. [Google Scholar] [CrossRef] [PubMed]
- Zaheer, M.; Wang, C.; Bian, F.; Yu, Z.; Hernandez, H.; de Souza, R.G.; Simmons, K.T.; Schady, D.; Swennes, A.G.; Pflugfelder, S.C.; et al. Protective Role of Commensal Bacteria in Sjögren Syndrome. J. Autoimmun. 2018, 93, 45–56. [Google Scholar] [CrossRef]
- Wang, M.; Zhu, Z.; Lin, X.; Li, H.; Wen, C.; Bao, J.; He, Z. Gut microbiota mediated the therapeutic efficacies and the side effects of prednisone in the treatment of MRL/lpr mice. Arthritis Res. Ther. 2021, 23, 240. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Q.; Yu, Y.; Wang, M.; Wen, C.; He, Z. Early and Short-Term Interventions in the Gut Microbiota Affects Lupus Severity, Progression, and Treatment in MRL/lpr Mice. Front. Microbiol. 2020, 11, 628. [Google Scholar] [CrossRef]
- Khakisahneh, S.; Zhang, X.-Y.; Nouri, Z.; Wang, D.-H. Cecal microbial transplantation attenuates hyperthyroid-induced thermogenesis in Mongolian gerbils. Microb. Biotechnol. 2022, 15, 817–831. [Google Scholar] [CrossRef]
- Moshkelgosha, S.; Verhasselt, H.L.; Masetti, G.; Covelli, D.; Biscarini, F.; Horstmann, M.; Daser, A.; Westendorf, A.M.; Jesenek, C.; Philipp, S.; et al. Modulating gut microbiota in a mouse model of Graves’ orbitopathy and its impact on induced disease. Microbiome 2021, 9, 45. [Google Scholar] [CrossRef]
- Su, X.; Yin, X.; Liu, Y.; Yan, X.; Zhang, S.; Wang, X.; Lin, Z.; Zhou, X.; Gao, J.; Wang, Z.; et al. Gut Dysbiosis Contributes to the Imbalance of Treg and Th17 Cells in Graves’ Disease Patients by Propionic Acid. J. Clin. Endocrinol. Metab. 2020, 105, 3526–3547. [Google Scholar] [CrossRef]
- Su, X.; Zhao, Y.; Li, Y.; Ma, S.; Wang, Z. Gut dysbiosis is associated with primary hypothyroidism with interaction on gut-thyroid axis. Clin. Sci. Lond. Engl. 1979 2020, 134, 1521–1535. [Google Scholar] [CrossRef]
- Edwards, V.; Smith, D.L.; Meylan, F.; Tiffany, L.; Poncet, S.; Wu, W.W.; Phue, J.-N.; Santana-Quintero, L.; Clouse, K.A.; Gabay, O. Analyzing the Role of Gut Microbiota on the Onset of Autoimmune Diseases Using TNFΔARE Murine Model. Microorganisms 2022, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Pu, Y.; Zhang, Q.; Tang, Z.; Lu, C.; Wu, L.; Zhong, Y.; Chen, Y.; Hashimoto, K.; Luo, Y.; Liu, Y. Fecal microbiota transplantation from patients with rheumatoid arthritis causes depression-like behaviors in mice through abnormal T cells activation. Transl. Psychiatry 2022, 12, 223. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, Y.; Ouhara, K.; Munenaga, S.; Shoji, M.; Ozawa, T.; Hisatsune, J.; Kado, I.; Kajiya, M.; Matsuda, S.; Kawai, T.; et al. Effect of Porphyromonas gingivalis infection on gut dysbiosis and resultant arthritis exacerbation in mouse model. Arthritis Res. Ther. 2020, 22, 249. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.J.; Wenndorf, K.; Ebbers, M.; Volzke, J.; Müller, M.; Strübing, J.; Kriebel, K.; Kneitz, S.; Kreikemeyer, B.; Müller-Hilke, B. Infection with Clostridioides difficile Attenuated Collagen-Induced Arthritis in Mice and Involved Mesenteric Treg and Th2 Polarization. Front. Immunol. 2020, 11, 571049. [Google Scholar] [CrossRef]
- Schaefer, L.; Trujillo-Vargas, C.M.; Midani, F.S.; Pflugfelder, S.C.; Britton, R.A.; de Paiva, C.S. Gut Microbiota From Sjögren syndrome Patients Causes Decreased T Regulatory Cells in the Lymphoid Organs and Desiccation-Induced Corneal Barrier Disruption in Mice. Front. Med. 2022, 9, 555. [Google Scholar] [CrossRef]
- Ma, Y.; Xu, X.; Li, M.; Cai, J.; Wei, Q.; Niu, H. Gut microbiota promote the inflammatory response in the pathogenesis of systemic lupus erythematosus. Mol. Med. Camb. Mass 2019, 25, 35. [Google Scholar] [CrossRef]
- Ma, Y.; Guo, R.; Sun, Y.; Li, X.; He, L.; Li, Z.; Silverman, G.J.; Chen, G.; Gao, F.; Yuan, J.; et al. Lupus gut microbiota transplants cause autoimmunity and inflammation. Clin. Immunol. Orlando Fla 2021, 233, 108892. [Google Scholar] [CrossRef]
- Neuman, V.; Cinek, O.; Funda, D.P.; Hudcovic, T.; Golias, J.; Kramna, L.; Petruzelkova, L.; Pruhova, S.; Sumnik, Z. Human gut microbiota transferred to germ-free NOD mice modulate the progression towards type 1 diabetes regardless of the pace of beta cell function loss in the donor. Diabetologia 2019, 62, 1291–1296. [Google Scholar] [CrossRef]
- Scherer, H.U.; Häupl, T.; Burmester, G.R. The etiology of rheumatoid arthritis. J. Autoimmun. 2020, 110, 102400. [Google Scholar] [CrossRef]
- Vaahtovuo, J.; Munukka, E.; Korkeamäki, M.; Luukkainen, R.; Toivanen, P. Fecal microbiota in early rheumatoid arthritis. J. Rheumatol. 2008, 35, 1500–1505. [Google Scholar]
- Esberg, A.; Johansson, L.; Johansson, I.; Dahlqvist, S.R. Oral Microbiota Identifies Patients in Early Onset Rheumatoid Arthritis. Microorganisms 2021, 9, 1657. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Mannaa, M.; Kim, Y.; Kim, J.; Kim, G.-T.; Seo, Y.-S. Comparative Analysis of Fecal Microbiota Composition Between Rheumatoid Arthritis and Osteoarthritis Patients. Genes 2019, 10, 748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wright, K.; Davis, J.M.; Jeraldo, P.; Marietta, E.V.; Murray, J.; Nelson, H.; Matteson, E.L.; Taneja, V. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med. 2016, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Chiang, H.-I.; Li, J.-R.; Liu, C.-C.; Liu, P.-Y.; Chen, H.-H.; Chen, Y.-M.; Lan, J.-L.; Chen, D.-Y. An Association of Gut Microbiota with Different Phenotypes in Chinese Patients with Rheumatoid Arthritis. J. Clin. Med. 2019, 8, 1770. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.; Yang, Z.; Zhang, X.; Liu, Z.; Wang, M.; Wu, X.; Chen, X.; Huang, Q.; Huang, R. Sustained Drug Treatment Alters the Gut Microbiota in Rheumatoid Arthritis. Front. Immunol. 2021, 12, 4238. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Jia, H.; Feng, Q.; Wang, D.; Liang, D.; Wu, X.; Li, J.; Tang, L.; Li, Y.; et al. The oral and gut microbiomes are perturbed in rheumatoid arthritis and partly normalized after treatment. Nat. Med. 2015, 21, 895–905. [Google Scholar] [CrossRef]
- Kishikawa, T.; Maeda, Y.; Nii, T.; Motooka, D.; Matsumoto, Y.; Matsushita, M.; Matsuoka, H.; Yoshimura, M.; Kawada, S.; Teshigawara, S.; et al. Metagenome-wide association study of gut microbiome revealed novel aetiology of rheumatoid arthritis in the Japanese population. Ann. Rheum. Dis. 2020, 79, 103–111. [Google Scholar] [CrossRef]
- Kitamura, K.; Shionoya, H.; Suzuki, S.; Fukai, R.; Uda, S.; Abe, C.; Takemori, H.; Nishimura, K.; Baba, H.; Katayama, K.; et al. Oral and Intestinal Bacterial Substances Associated with Disease Activities in Patients with Rheumatoid Arthritis: A Cross-Sectional Clinical Study. J. Immunol. Res. 2022, 2022, 6839356. [Google Scholar] [CrossRef]
- Xu, H.; Cao, J.; Li, X.; Lu, X.; Xia, Y.; Fan, D.; Zhao, H.; Ju, D.; Xiao, C. Regional Differences in the Gut Microbiota and Gut-Associated Immunologic Factors in the Ileum and Cecum of Rats with Collagen-Induced Arthritis. Front. Pharmacol. 2020, 11, 587534. [Google Scholar] [CrossRef]
- Maier, L.; Pruteanu, M.; Kuhn, M.; Zeller, G.; Telzerow, A.; Anderson, E.E.; Brochado, A.R.; Fernandez, K.C.; Dose, H.; Mori, H.; et al. Extensive impact of non-antibiotic drugs on human gut bacteria. Nature 2018, 555, 623–628. [Google Scholar] [CrossRef]
- Hao, X.; Shang, X.; Liu, J.; Chi, R.; Zhang, J.; Xu, T. The gut microbiota in osteoarthritis: Where do we stand and what can we do? Arthritis Res. Ther. 2021, 23, 42. [Google Scholar] [CrossRef] [PubMed]
- Ricciuto, A.; Sherman, P.M.; Laxer, R.M. Gut microbiota in chronic inflammatory disorders: A focus on pediatric inflammatory bowel diseases and juvenile idiopathic arthritis. Clin. Immunol. Orlando Fla 2020, 215, 108415. [Google Scholar] [CrossRef] [PubMed]
- Eppinga, H.; Konstantinov, S.R.; Peppelenbosch, M.P.; Thio, H.B. The Microbiome and Psoriatic Arthritis. Curr. Rheumatol. Rep. 2014, 16, 407. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Hsu, C.-Y.; He, H.-R.; Chiang, W.-Y.; Lin, S.-H.; Huang, Y.-L.; Kuo, Y.-H.; Su, Y.-J. Gut microbiota differences between psoriatic arthritis and other undifferentiated arthritis: A pilot study. Medicine (Baltimore) 2022, 101, e29870. [Google Scholar] [CrossRef]
- FitzGerald, O.; Ogdie, A.; Chandran, V.; Coates, L.C.; Kavanaugh, A.; Tillett, W.; Leung, Y.Y.; deWit, M.; Scher, J.U.; Mease, P.J. Psoriatic arthritis. Nat. Rev. Dis. Primer 2021, 7, 59. [Google Scholar] [CrossRef] [PubMed]
- Myers, B.; Brownstone, N.; Reddy, V.; Chan, S.; Thibodeaux, Q.; Truong, A.; Bhutani, T.; Chang, H.-W.; Liao, W. The gut microbiome in psoriasis and psoriatic arthritis. Best Pract. Res. Clin. Rheumatol. 2019, 33, 101494. [Google Scholar] [CrossRef]
- Olejniczak-Staruch, I.; Ciążyńska, M.; Sobolewska-Sztychny, D.; Narbutt, J.; Skibińska, M.; Lesiak, A. Alterations of the Skin and Gut Microbiome in Psoriasis and Psoriatic Arthritis. Int. J. Mol. Sci. 2021, 22, 3998. [Google Scholar] [CrossRef]
- Axisa, P.-P.; Hafler, D.A. Multiple sclerosis: Genetics, biomarkers, treatments. Curr. Opin. Neurol. 2016, 29, 345–353. [Google Scholar] [CrossRef]
- Galluzzo, P.; Capri, F.C.; Vecchioni, L.; Realmuto, S.; Scalisi, L.; Cottone, S.; Nuzzo, D.; Alduina, R. Comparison of the Intestinal Microbiome of Italian Patients with Multiple Sclerosis and Their Household Relatives. Life 2021, 11, 620. [Google Scholar] [CrossRef]
- Waubant, E.; Lucas, R.; Mowry, E.; Graves, J.; Olsson, T.; Alfredsson, L.; Langer-Gould, A. Environmental and genetic risk factors for MS: An integrated review. Ann. Clin. Transl. Neurol. 2019, 6, 1905–1922. [Google Scholar] [CrossRef]
- Navarro-López, V.; Méndez-Miralles, M.Á.; Vela-Yebra, R.; Fríes-Ramos, A.; Sánchez-Pellicer, P.; Ruzafa-Costas, B.; Núñez-Delegido, E.; Gómez-Gómez, H.; Chumillas-Lidón, S.; Picó-Monllor, J.A.; et al. Gut Microbiota as a Potential Predictive Biomarker in Relapsing-Remitting Multiple Sclerosis. Genes 2022, 13, 930. [Google Scholar] [CrossRef] [PubMed]
- Pellizoni, F.P.; Leite, A.Z.; Rodrigues, N.d.C.; Ubaiz, M.J.; Gonzaga, M.I.; Takaoka, N.N.C.; Mariano, V.S.; Omori, W.P.; Pinheiro, D.G.; Matheucci Junior, E.; et al. Detection of Dysbiosis and Increased Intestinal Permeability in Brazilian Patients with Relapsing–Remitting Multiple Sclerosis. Int. J. Environ. Res. Public. Health 2021, 18, 4621. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.M.; Maghzi, A.H.; Liu, S.; Tankou, S.K.; Dhang, F.H.; Willocq, V.; Song, A.; Wasén, C.; Tauhid, S.; Chu, R.; et al. Gut Microbiome in Progressive Multiple Sclerosis. Ann. Neurol. 2021, 89, 1195–1211. [Google Scholar] [CrossRef] [PubMed]
- Horton, M.K.; McCauley, K.; Fadrosh, D.; Fujimura, K.; Graves, J.; Ness, J.; Wheeler, Y.; Gorman, M.P.; Benson, L.A.; Weinstock-Guttman, B.; et al. Gut microbiome is associated with multiple sclerosis activity in children. Ann. Clin. Transl. Neurol. 2021, 8, 1867–1883. [Google Scholar] [CrossRef]
- Mekky, J.; Wani, R.; Said, S.M.; Ashry, M.; Ibrahim, A.E.; Ahmed, S.M. Molecular characterization of the gut microbiome in egyptian patients with remitting relapsing multiple sclerosis. Mult. Scler. Relat. Disord. 2022, 57, 103354. [Google Scholar] [CrossRef]
- Mirza, A.I.; Zhu, F.; Knox, N.; Forbes, J.D.; Bonner, C.; Van Domselaar, G.; Bernstein, C.N.; Graham, M.; Marrie, R.A.; Hart, J.; et al. The metabolic potential of the paediatric-onset multiple sclerosis gut microbiome. Mult. Scler. Relat. Disord. 2022, 63, 103829. [Google Scholar] [CrossRef]
- Ochoa-Repáraz, J.; Mielcarz, D.W.; Ditrio, L.E.; Burroughs, A.R.; Foureau, D.M.; Haque-Begum, S.; Kasper, L.H. Role of Gut Commensal Microflora in the Development of Experimental Autoimmune Encephalomyelitis. J. Immunol. 2009, 183, 6041–6050. [Google Scholar] [CrossRef]
- Giongo, A.; Gano, K.A.; Crabb, D.B.; Mukherjee, N.; Novelo, L.L.; Casella, G.; Drew, J.C.; Ilonen, J.; Knip, M.; Hyöty, H.; et al. Toward defining the autoimmune microbiome for type 1 diabetes. ISME J. 2011, 5, 82–91. [Google Scholar] [CrossRef]
- Murri, M.; Leiva, I.; Gomez-Zumaquero, J.M.; Tinahones, F.J.; Cardona, F.; Soriguer, F.; Queipo-Ortuño, M.I. Gut microbiota in children with type 1 diabetes differs from that in healthy children: A case-control study. BMC Med. 2013, 11, 46. [Google Scholar] [CrossRef]
- Pellegrini, S.; Sordi, V.; Bolla, A.M.; Saita, D.; Ferrarese, R.; Canducci, F.; Clementi, M.; Invernizzi, F.; Mariani, A.; Bonfanti, R.; et al. Duodenal Mucosa of Patients with Type 1 Diabetes Shows Distinctive Inflammatory Profile and Microbiota. J. Clin. Endocrinol. Metab. 2017, 102, 1468–1477. [Google Scholar] [CrossRef]
- Liu, X.; Cheng, Y.W.; Shao, L.; Sun, S.H.; Wu, J.; Song, Q.H.; Zou, H.S.; Ling, Z.X. Gut microbiota dysbiosis in Chinese children with type 1 diabetes mellitus: An observational study. World J. Gastroenterol. 2021, 27, 2394–2414. [Google Scholar] [CrossRef] [PubMed]
- Qi, C.J.; Zhang, Q.; Yu, M.; Xu, J.P.; Zheng, J.; Wang, T.; Xiao, X.H. Imbalance of Fecal Microbiota at Newly Diagnosed Type 1 Diabetes in Chinese Children. Chin. Med. J. 2016, 129, 1298–1304. [Google Scholar] [CrossRef] [PubMed]
- Maffeis, C.; Martina, A.; Corradi, M.; Quarella, S.; Nori, N.; Torriani, S.; Plebani, M.; Contreas, G.; Felis, G.E. Association between intestinal permeability and faecal microbiota composition in Italian children with beta cell autoimmunity at risk for type 1 diabetes. Diabetes Metab. Res. Rev. 2016, 32, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Alkanani, A.K.; Hara, N.; Gottlieb, P.A.; Ir, D.; Robertson, C.E.; Wagner, B.D.; Frank, D.N.; Zipris, D. Alterations in intestinal microbiota correlate with susceptibility to type 1 diabetes. Diabetes 2015, 64, 3510–3520. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.C.G.; Oliveira, R.P.; Torres, L.; Aguiar, S.L.F.; Pinheiro-Rosa, N.; Lemos, L.; Guimarães, M.A.; Reis, D.; Silveira, T.; Ferreira, Ê.; et al. Frontline Science: Abnormalities in the gut mucosa of non-obese diabetic mice precede the onset of type 1 diabetes. J. Leukoc. Biol. 2019, 106, 513–529. [Google Scholar] [CrossRef]
- Mullaney, J.A.; Stephens, J.E.; Geeling, B.E.; Hamilton-Williams, E.E. Early-life exposure to gut microbiota from disease-protected mice does not impact disease outcome in type 1 diabetes susceptible NOD mice. Immunol. Cell Biol. 2019, 97, 97–103. [Google Scholar] [CrossRef]
- McLeod, K.; Mason, L.; Mariño, E. Transplantation of Fecal Microbiota Shaped by Diet. Bio-Protoc. 2018, 8, e2683. [Google Scholar] [CrossRef]
- Zhang, H.; Liao, X.; Sparks, J.B.; Luo, X.M. Dynamics of gut microbiota in autoimmune lupus. Appl. Environ. Microbiol. 2014, 80, 7551–7560. [Google Scholar] [CrossRef]
- Mu, Q.; Tavella, V.J.; Kirby, J.L.; Cecere, T.E.; Chung, M.; Lee, J.; Li, S.; Ahmed, S.A.; Eden, K.; Allen, I.C.; et al. Antibiotics ameliorate lupus-like symptoms in mice. Sci. Rep. 2017, 7, 13675. [Google Scholar] [CrossRef]
- Edwards, M.R.; Dai, R.; Heid, B.; Cecere, T.E.; Khan, D.; Mu, Q.; Cowan, C.; Luo, X.M.; Ahmed, S.A. Commercial rodent diets differentially regulate autoimmune glomerulonephritis, epigenetics and microbiota in MRL/lpr mice. Int. Immunol. 2017, 29, 263–276. [Google Scholar] [CrossRef]
- Wang, H.; Wang, G.; Banerjee, N.; Liang, Y.; Du, X.; Boor, P.J.; Hoffman, K.L.; Khan, M.F. Aberrant Gut Microbiome Contributes to Intestinal Oxidative Stress, Barrier Dysfunction, Inflammation and Systemic Autoimmune Responses in MRL/lpr Mice. Front. Immunol. 2021, 12, 651191. [Google Scholar] [CrossRef] [PubMed]
- Hevia, A.; Milani, C.; López, P.; Cuervo, A.; Arboleya, S.; Duranti, S.; Turroni, F.; González, S.; Suárez, A.; Gueimonde, M.; et al. Intestinal dysbiosis associated with systemic lupus erythematosus. mBio 2014, 5, e01548-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Z.; Shao, T.; Li, H.; Xie, Z.; Wen, C. Alterations of the gut microbiome in Chinese patients with systemic lupus erythematosus. Gut Pathog. 2016, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Gerges, M.A.; Esmaeel, N.E.; Makram, W.K.; Sharaf, D.M.; Gebriel, M.G. Altered Profile of Fecal Microbiota in Newly Diagnosed Systemic Lupus Erythematosus Egyptian Patients. Int. J. Microbiol. 2021, 2021, 9934533. [Google Scholar] [CrossRef]
- López, P.; De Paz, B.; Rodríguez-Carrio, J.; Hevia, A.; Sánchez, B.; Margolles, A.; Suárez, A. Th17 responses and natural IgM antibodies are related to gut microbiota composition in systemic lupus erythematosus patients. Sci. Rep. 2016, 6, 24072. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wan, X.; Wu, X.; Zhang, C.; Liu, J.; Hou, S. Eubacterium rectale contributes to colorectal cancer initiation via promoting colitis. Gut Pathog. 2021, 13, 2. [Google Scholar] [CrossRef] [PubMed]
- Golfetto, L.; de Senna, F.D.; Hermes, J.; Beserra, B.T.S.; da Silva França, F.; Martinello, F. Lower bifidobacteria counts in adult patients with celiac disease on a gluten-free diet. Arq. Gastroenterol. 2014, 51, 139–143. [Google Scholar] [CrossRef]
- Bibbò, S.; Abbondio, M.; Sau, R.; Tanca, A.; Pira, G.; Errigo, A.; Manetti, R.; Pes, G.M.; Dore, M.P.; Uzzau, S. Fecal Microbiota Signatures in Celiac Disease Patients with Poly-Autoimmunity. Front. Cell. Infect. Microbiol. 2020, 10, 349. [Google Scholar] [CrossRef]
- Olivares, M.; Walker, A.W.; Capilla, A.; Benítez-Páez, A.; Palau, F.; Parkhill, J.; Castillejo, G.; Sanz, Y. Gut microbiota trajectory in early life may predict development of celiac disease. Microbiome 2018, 6, 36. [Google Scholar] [CrossRef]
- Rintala, A.; Riikonen, I.; Toivonen, A.; Pietilä, S.; Munukka, E.; Pursiheimo, J.P.; Elo, L.L.; Arikoski, P.; Luopajärvi, K.; Schwab, U.; et al. Early fecal microbiota composition in children who later develop celiac disease and associated autoimmunity. Scand. J. Gastroenterol. 2018, 53, 403–409. [Google Scholar] [CrossRef]
- Panelli, S.; Capelli, E.; Lupo, G.F.D.; Schiepatti, A.; Betti, E.; Sauta, E.; Marini, S.; Bellazzi, R.; Vanoli, A.; Pasi, A.; et al. Comparative Study of Salivary, Duodenal, and Fecal Microbiota Composition Across Adult Celiac Disease. J. Clin. Med. 2020, 9, 1109. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Vannini, L.; Di Cagno, R.; Cavallo, N.; Minervini, F.; Francavilla, R.; Ercolini, D.; Gobbetti, M. Salivary and fecal microbiota and metabolome of celiac children under gluten-free diet. Int. J. Food Microbiol. 2016, 239, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Bodkhe, R.; Shetty, S.A.; Dhotre, D.P.; Verma, A.K.; Bhatia, K.; Mishra, A.; Kaur, G.; Pande, P.; Bangarusamy, D.K.; Santosh, B.P.; et al. Comparison of Small Gut and Whole Gut Microbiota of First-Degree Relatives with Adult Celiac Disease Patients and Controls. Front. Microbiol. 2019, 10, 137–140. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Rivera-Gutierrez, X.; Cobos-Quevedo, O.D.J.; Grube-Pagola, P.; Meixueiro-Daza, A.; Hernandez-Flores, K.; Cabrera-Jorge, F.J.; Vivanco-Cid, H.; Dowd, S.E.; Remes-Troche, J.M. First Insights into the Gut Microbiota of Mexican Patients with Celiac Disease and Non-Celiac Gluten Sensitivity. Nutrients 2018, 10, 1641. [Google Scholar] [CrossRef]
- Nylund, L.; Hakkola, S.; Lahti, L.; Salminen, S.; Kalliomäki, M.; Yang, B.; Linderborg, K.M. Diet, Perceived Intestinal Well-Being and Compositions of Fecal Microbiota and Short Chain Fatty Acids in Oat-Using Subjects with Celiac Disease or Gluten Sensitivity. Nutrients 2020, 12, 2570. [Google Scholar] [CrossRef]
- Nobel, Y.R.; Rozenberg, F.; Park, H.; Freedberg, D.E.; Blaser, M.J.; Green, P.H.R.; Uhlemann, A.C.; Lebwohl, B. Lack of Effect of Gluten Challenge on Fecal Microbiome in Patients with Celiac Disease and Non-Celiac Gluten Sensitivity. Clin. Transl. Gastroenterol. 2021, 12, e00441. [Google Scholar] [CrossRef] [PubMed]
- Lamas, B.; Hernandez-Galan, L.; Galipeau, H.J.; Constante, M.; Clarizio, A.; Jury, J.; Breyner, N.M.; Caminero, A.; Rueda, G.; Hayes, C.L.; et al. Aryl hydrocarbon receptor ligand production by the gut microbiota is decreased in celiac disease leading to intestinal inflammation. Sci. Transl. Med. 2020, 12, eaba0624. [Google Scholar] [CrossRef] [PubMed]
- Abdukhakimova, D.; Dossybayeva, K.; Poddighe, D. Fecal and Duodenal Microbiota in Pediatric Celiac Disease. Front. Pediatr. 2021, 9, 652208. [Google Scholar] [CrossRef]
- Abadie, V.; Kim, S.M.; Lejeune, T.; Palanski, B.A.; Ernest, J.D.; Tastet, O.; Voisine, J.; Discepolo, V.; Marietta, E.V.; Hawash, M.B.F.; et al. IL-15, gluten and HLA-DQ8 drive tissue destruction in coeliac disease. Nature 2020, 578, 600–604. [Google Scholar] [CrossRef]
- Mori, K.; Nakagawa, Y.; Ozaki, H. Does the Gut Microbiota Trigger Hashimoto’s Thyroiditis? Discov. Med. 2012, 14, 321–326. [Google Scholar]
- Caturegli, P.; De Remigis, A.; Rose, N.R. Hashimoto thyroiditis: Clinical and diagnostic criteria. Autoimmun. Rev. 2014, 13, 391–397. [Google Scholar] [CrossRef] [PubMed]
- de Freitas Cayres, L.C.; de Salis, L.V.V.; Rodrigues, G.S.P.; van Helvoort Lengert, A.; Biondi, A.P.C.; Sargentini, L.D.B.; Brisotti, J.L.; Gomes, E.; de Oliveira, G.L.V. Detection of Alterations in the Gut Microbiota and Intestinal Permeability in Patients with Hashimoto Thyroiditis. Front. Immunol. 2021, 12, 579140. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Feng, J.; Li, J.; Zhao, L.; Liu, Y.; Chen, H.; Jin, Y.; Zhu, B.; Wei, Y. Alterations of the Gut Microbiota in Hashimoto’s Thyroiditis Patients. Thyroid 2018, 28, 175–186. [Google Scholar] [CrossRef]
- Virili, C.; Fallahi, P.; Antonelli, A.; Benvenga, S.; Centanni, M. Gut microbiota and Hashimoto’s thyroiditis. Rev. Endocr. Metab. Disord. 2018, 19, 293–300. [Google Scholar] [CrossRef]
- Zhao, H.; Yuan, L.; Zhu, D.; Sun, B.; Du, J.; Wang, J. Alterations and Mechanism of Gut Microbiota in Graves’ Disease and Hashimoto’s Thyroiditis. Pol. J. Microbiol. 2022, 71, 173–189. [Google Scholar] [CrossRef]
- Ishaq, H.M.; Mohammad, I.S.; Guo, H.; Shahzad, M.; Hou, Y.J.; Ma, C.; Naseem, Z.; Wu, X.; Shi, P.; Xu, J. Molecular estimation of alteration in intestinal microbial composition in Hashimoto’s thyroiditis patients. Biomed. Pharmacother. 2017, 95, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Wang, C.; Meng, F.; Wang, H.; Song, B.; Yang, Y.; Shan, Z. Association Between Gut Microbiota and Autoimmune Thyroid Disease: A Systematic Review and Meta-Analysis. Front. Endocrinol. 2021, 12, 1544. [Google Scholar] [CrossRef]
- Hou, J.; Tang, Y.; Chen, Y.; Chen, D. The Role of the Microbiota in Graves’ Disease and Graves’ Orbitopathy. Front. Cell. Infect. Microbiol. 2021, 11, 1301. [Google Scholar] [CrossRef]
- Antonelli, A.; Ferrari, S.M.; Ragusa, F.; Elia, G.; Paparo, S.R.; Ruffilli, I.; Patrizio, A.; Giusti, C.; Gonnella, D.; Cristaudo, A.; et al. Graves’ disease: Epidemiology, genetic and environmental risk factors and viruses. Best Pract. Res. Clin. Endocrinol. Metab. 2020, 34, 101387. [Google Scholar] [CrossRef]
- Yan, H.-X.; An, W.-C.; Chen, F.; An, B.; Pan, Y.; Jin, J.; Xia, X.-P.; Cui, Z.-J.; Jiang, L.; Zhou, S.-J.; et al. Intestinal microbiota changes in Graves’ disease: A prospective clinical study. Biosci. Rep. 2020, 40, BSR20191242. [Google Scholar] [CrossRef]
- Jiang, W.; Yu, X.; Kosik, R.O.; Song, Y.; Qiao, T.; Tong, J.; Liu, S.; Fan, S.; Luo, Q.; Chai, L.; et al. Gut Microbiota May Play a Significant Role in the Pathogenesis of Graves’ Disease. Thyroid Off. J. Am. Thyroid Assoc. 2021, 31, 810–820. [Google Scholar] [CrossRef] [PubMed]
- Holdgate, N.; St.Clair, E.W. Recent advances in primary Sjogren’s syndrome. F1000Research 2016, 5, F1000 Faculty Rev-1412. [Google Scholar] [CrossRef]
- Rasmussen, A.; Ice, J.A.; Li, H.; Grundahl, K.; Kelly, J.A.; Radfar, L.; Stone, D.U.; Hefner, K.S.; Anaya, J.-M.; Rohrer, M.; et al. Comparison of the American-European Consensus Group Sjögren’s syndrome classification criteria to newly proposed American College of Rheumatology criteria in a large, carefully characterized sicca cohort. Ann. Rheum. Dis. 2014, 73, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Mendez, R.; Watane, A.; Farhangi, M.; Cavuoto, K.M.; Leith, T.; Budree, S.; Galor, A.; Banerjee, S. Gut microbial dysbiosis in individuals with Sjögren’s syndrome. Microb. Cell Factories 2020, 19, 90. [Google Scholar] [CrossRef] [PubMed]
- Cano-Ortiz, A.; Laborda-Illanes, A.; Plaza-Andrades, I.; Membrillo del Pozo, A.; Villarrubia Cuadrado, A.; Rodríguez Calvo de Mora, M.; Leiva-Gea, I.; Sanchez-Alcoholado, L.; Queipo-Ortuño, M.I. Connection between the Gut Microbiome, Systemic Inflammation, Gut Permeability and FOXP3 Expression in Patients with Primary Sjögren’s Syndrome. Int. J. Mol. Sci. 2020, 21, 8733. [Google Scholar] [CrossRef]
- D’Haens, G.R.; Jobin, C. Fecal Microbial Transplantation for Diseases Beyond Recurrent Clostridium Difficile Infection. Gastroenterology 2019, 157, 624–636. [Google Scholar] [CrossRef]
- Sonali, S.; Ray, B.; Ahmed Tousif, H.; Rathipriya, A.G.; Sunanda, T.; Mahalakshmi, A.M.; Rungratanawanich, W.; Essa, M.M.; Qoronfleh, M.W.; Chidambaram, S.B.; et al. Mechanistic Insights into the Link between Gut Dysbiosis and Major Depression: An Extensive Review. Cells 2022, 11, 1362. [Google Scholar] [CrossRef]

{kind=link}
| Disorder | Main Outcome | Outcome Details | References |
|---|---|---|---|
| Celiac disease | Successful | Cured Clostridioides difficile infection. Mitigation of symptoms of celiac disease. | [25] |
| Multiple sclerosis | Potential but more research is necessary | A donor-specific alteration of gut microbiota. No statistically significant changes of pro-inflammatory regulatory cytokines. | [13] |
| Multiple sclerosis | Potential but more research is necessary | An increase in short chain fatty acids (SCFA) genomic pathways post-FMT. A positive correlation between the abundance of microbial SCFA pathway gene content and serum brain-derived neurotrophic factor. Species Faecalibacterium prausnitzii elevated. Butyrate, propionate, total SCFA and total-butyrate-to-total SCFA ratio concentrations increased in 2 out of 5 post-FMT measurements. | [26] |
| Psoriatic arthritis | Potential but more research is necessary | Acceptable and safe FMT application. No life-threatening effects. | [12] |
| Psoriatic arthritis | Failed | Health assessment questionnaire disability index improved more in the placebo group compared with the FMT group. | [27] |
| Rheumatoid arthritis | Successful | Successfully cured with FMT. A decrease in rheumatoid factor, disease activity score-28 and improvement of the health assessment questionnaire Disability Index. | [11] |
| Sjogren’s syndrome | Failed | A donor-specific alterations of gut microbiota. However, microbiota of recipients still significantly different from donors. No significant changes before and after the treatment within recipient samples. Despite this, improved symptoms in 50% of study respondents. | [28] |
| Type 1 diabetes | Potential but more research is necessary | Type I diabetes progression slowed down. Stimulated C peptide levels were preserved in the autologous FMT group compared with healthy donor FMT group. Small intestinal Prevotella was inversely related to residual beta cell function. | [29] |
| Disorder | Current Status | Identifier and Full Name of the Study | References |
|---|---|---|---|
| Multiple sclerosis | Completed | NCT03975413 Single-arm, non-randomized, time series, single-subject study: fecal microbiota transplantation (FMT) in multiple sclerosis | [30] |
| Multiple sclerosis | Active, not recruiting | NCT03594487 Fecal microbiota transplantation (FMT) of FMP30 in relapsing-remitting multiple sclerosis: a phase 1b clinical trial to evaluate feasibility, safety, tolerability and effects on immune function | [31] |
| Multiple sclerosis | Recruiting | NCT04203017 Allogeneic fecal microbiota transplantation as a consolidation treatment after autologous hematopoietic stem cell transplantation in patients with multiple sclerosis | [32] |
| Multiple sclerosis | Terminated (primary investigator passed away) | NCT03183869 Fecal microbial transplantation in relapsing multiple sclerosis patients | [33] |
| Multiple sclerosis | Recruiting | NCT04096443 A pilot study of oral FMT (fecal microbial transplant) in subjects with multiple sclerosis | [34] |
| Multiple sclerosis | Not yet recruiting | NCT04150549 Fecal microbial transplantation for relapsing multiple sclerosis patients—a placebo-controlled, double-blinded, randomized trial | [35] |
| Multiple sclerosis, Psoriatic arthritis, Celiac disease | Recruiting | NCT04014413 Safety and efficacy of fecal microbiota transplantation: a pilot study | [36] |
| Psoriatic arthritis | Completed | NCT03058900 Efficacy and safety of fecal microbiota transplantation (FMT) in patients with peripheral psoriatic arthritis: a 6-month, double-blind, randomized, placebo-controlled trial | [37] |
| Rheumatoid arthritis and Psoriatic arthritis | Unknown | NCT03944096 Efficacy and safety of fecal microbiota transplantation in patients with rheumatoid arthritis refractory to methotrexate: a 24-week, double-blind, randomized trial | [38] |
| Rheumatoid arthritis, Psoriatic arthritis | Not yet recruiting | NCT04924270 Safety and clinical efficacy associated with fecal microbiota transplantation performed in treatment-naïve patients with newly diagnosed rheumatoid arthritis, reactive arthritis, ankylosing spondylitis, psoriatic arthritis, gouty arthritis, psoriasis, hidradenitis suppurativa, pulmonary sarcoidosis, Crohn’s disease and ulcerative colitis: a 52-week, double-blind, randomized, placebo-controlled, exploratory trial | [39] |
| Sjogren’s syndrome | Completed | NCT03926286 Fecal microbial transplant for Sjogrens syndrome | [40] |
| Type 1 diabetes | Recruiting | NCT04749030 Fecal microbiota transplantation for patients with diabetes mellitus type 1 and severe gastrointestinal neuropathy: a randomized, double-blinded safety and pilot-efficacy study | [41] |
| Type 1 diabetes | Recruiting | NCT05323162 Encapsulated fecal microbiota transplantation to preserve residual beta cell function in patients with recently diagnosed type 1 diabetes mellitus | [42] |
| Type 1 diabetes | Unknown | NCT04124211 Fecal microbiome transplantation (FMT) for type 1 diabetes | [43] |
| Disorder | Main Outcome | Outcome Details | References |
|---|---|---|---|
| Multiple sclerosis | Successful | A reduction of cord pathology and ameliorated disease severity. | [44] |
| Multiple sclerosis | Successful | The onset and amelioration of the disease slowed down post-FMT. A decrease of 13 bacterial taxa, an increase of 17 taxa. Numerous bacteria-specific negative and positive correlations described. Reduced clinical scores and cumulative disease scores. Increased expression of a tight junction protein. Decreased expression of neurofilament light chain protein. | [45] |
| Rheumatoid arthritis | Successful | A positive effect of tuna elastin peptides transmitted by FMT. | [46] |
| Sjogren’s syndrome | Successful | A donor-specific alteration of gut microbiota. Minimalized disruption of the corneal epithelial barrier. Improved density of the goblet cells. Improved autoreactive CD4+ T cells. Increase of conjunctival goblet cells. | [47] |
| Sjogren’s syndrome | Successful | Improved corneal barrier function and Sjogren’s syndrome-like phenotype. A decrease of CD4+IFNγ+ cells. | [48] |
| Systemic lupus erythematosus | Successful | Alleviated systemic lupus erythematosus symptoms after prednisone-regulated gut microbiota FMT. Decreased relative abundance of genera Ruminococcus and Alistipes. Retained relative abundance of Lactobacillus. | [49] |
| Systemic lupus erythematosus | Successful | Alleviated gut dysbiosis caused by prior antibiotics administration and suppressed SLE progression. | [50] |
| Disorder | FMT Transfer | Main Outcome | Outcome Details | References |
|---|---|---|---|---|
| Graves’ disease | A-to-A | Successful | Gut microbiota alternations post-FMT. A greater decrease of T3 and T4 hormone concentrations, increased liver expression of type 2 deiodinase and better recovery of hypothyroid-induced resting metabolic rate back to normal. | [51] |
| Graves’ disease | H-to-A | Successful | A donor-specific alteration of gut microbiota. Increased disease severity, reduction in Shannon diversity, increased richness indices. Decreased abundance of Bacteroides compared with control mice. | [52] |
| Graves’ disease | H-to-A | Successful | Increased incidence of Graves’ disease. An increase of serum total thyroxine concentrations, thyroglobulin antibodies and IL-17A. Decreased serum concentrations of IL-10. | [53] |
| Hashimoto’s disease | H-to-A | Successful | A decrease of serum total thyroxine concentrations, mRNA expression of occludin, junctional adhesion molecule-A and zonula occludens-3211. | [54] |
| Rheumatoid arthritis | A-to-A | Successful | Physical changes, e.g., cartilage alterations, paw deformities present. Increased concentration of tissue inflammatory mediators. Activation of CD4/CD8+ T-lymphocytes. Behavioral modifications. Occult bleeding with gut tissue disruption. | [55] |
| Rheumatoid arthritis | H-to-A | Successful | Depression-like phenotypes. Alterations of gut microbiota composition. Increased percentage of CD3e+ and CD4+ T-lymphocytes in Peyer’s plaques and spleen. Increased Th1/Th2 index and decreased CD25+ and FOX3+ Treg cells. Downregulation of synaptic proteins. Negative correlation of Bacteroides, Phascolarctobacterium with the Th1/Th2 index and positive correlations with a decreased percentage of Treg cells in Peyer’s plaques and spleen. Twelve promising rheumatoid bacterial biomarkers proposed. | [56] |
| Rheumatoid arthritis | A-to-A | Successful | Severe joint swelling. Maximum arthritis score observed in FMT transplanted mice compared to non-FMT mice. | [57] |
| Rheumatoid arthritis | A-to-A | Failed | Attenuation of experimental arthritis more efficient without antibiotic treatment and FMT administration. | [58] |
| Sjogren’s syndrome | H-to-A | Potential but more research is necessary | Decreased corneal epithelial barrier integrity and decreased concentrations of CD45+, CD4+, FOXP3+ in cervical lymph nodes cells. Decreased CD4+, FOXP3+ cells in cervical lymph nodes tissue and spleen in offspring of FMT-transplanted mice. | [59] |
| Systemic lupus erythematosus | A-to-A | Successful | Changes in immune cell distribution in recipients. Upregulated expression of lupus susceptibility genes. | [60] |
| Systemic lupus erythematosus | H-to-A | Successful | A lupus-like phenotypic features in FMT transplanted mice. Increased serum autoimmune antibodies, imbalanced cytokines, altered distribution of immune cells in mucosal and peripheral immune response and upregulated expression of genes related to systemic lupus erythematosus. Metabolism of histidine modified. | [61] |
| Type 1 diabetes | H-to-A | Potential but more research is necessary | Delayed onset of Type I diabetes. The pace of beta cell loss not transferable to the mouse model. | [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belvoncikova, P.; Maronek, M.; Gardlik, R. Gut Dysbiosis and Fecal Microbiota Transplantation in Autoimmune Diseases. Int. J. Mol. Sci. 2022, 23, 10729. https://doi.org/10.3390/ijms231810729
Belvoncikova P, Maronek M, Gardlik R. Gut Dysbiosis and Fecal Microbiota Transplantation in Autoimmune Diseases. International Journal of Molecular Sciences. 2022; 23(18):10729. https://doi.org/10.3390/ijms231810729
Chicago/Turabian StyleBelvoncikova, Paulina, Martin Maronek, and Roman Gardlik. 2022. "Gut Dysbiosis and Fecal Microbiota Transplantation in Autoimmune Diseases" International Journal of Molecular Sciences 23, no. 18: 10729. https://doi.org/10.3390/ijms231810729
APA StyleBelvoncikova, P., Maronek, M., & Gardlik, R. (2022). Gut Dysbiosis and Fecal Microbiota Transplantation in Autoimmune Diseases. International Journal of Molecular Sciences, 23(18), 10729. https://doi.org/10.3390/ijms231810729