
Plants in Microgravity: Molecular and Technological Perspectives

 , ,
, , 
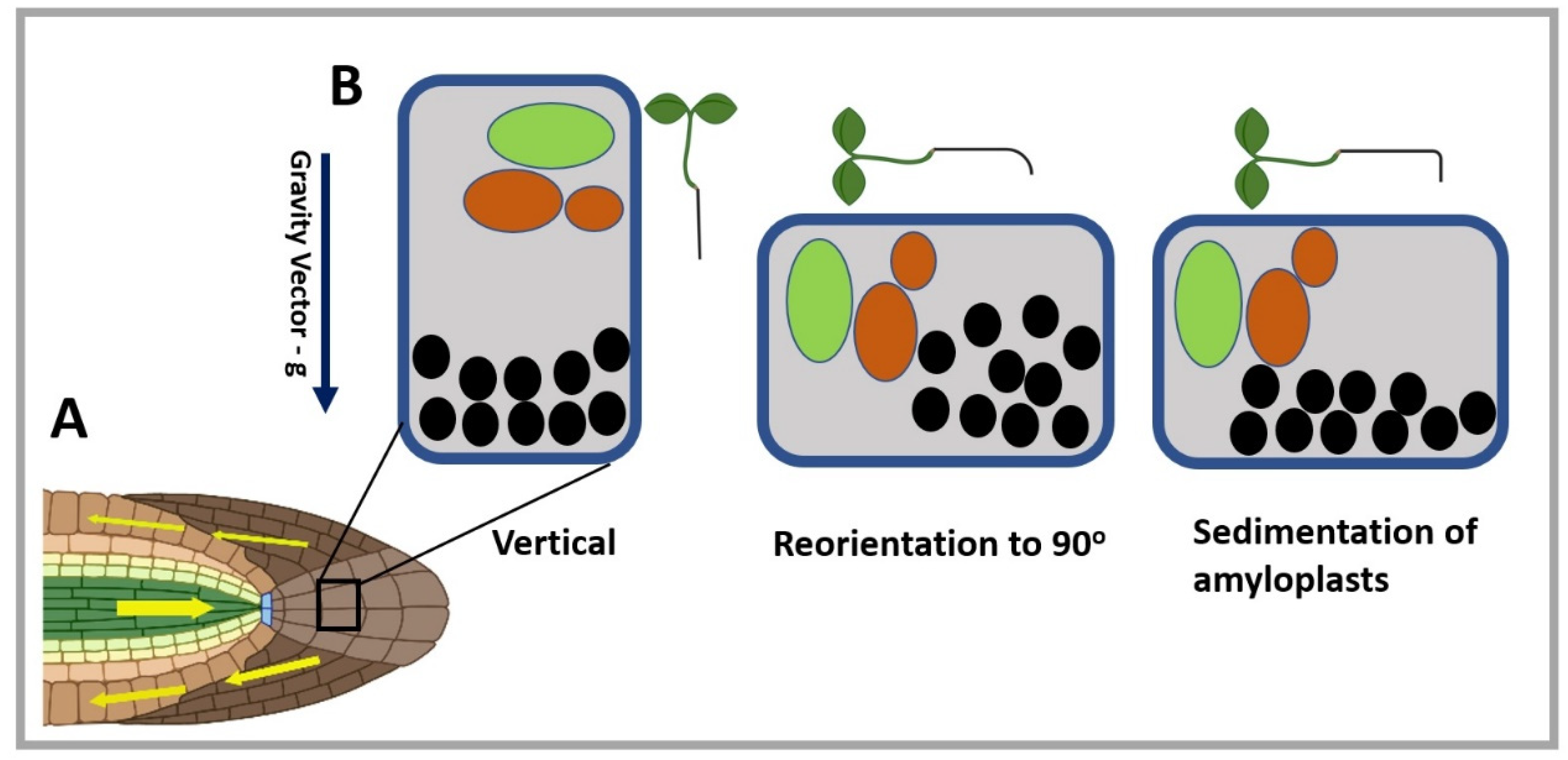{kind=link}
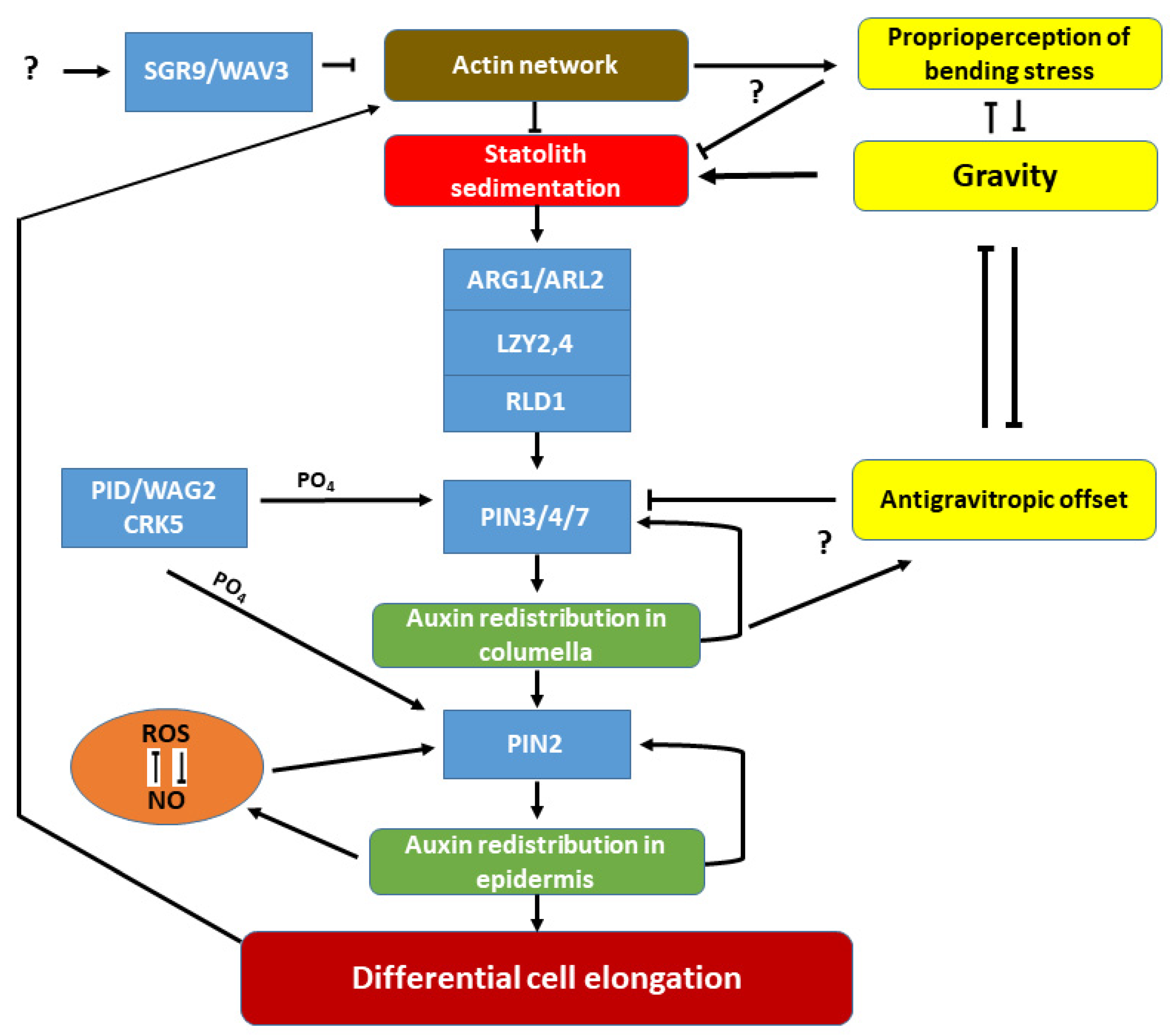{kind=link}
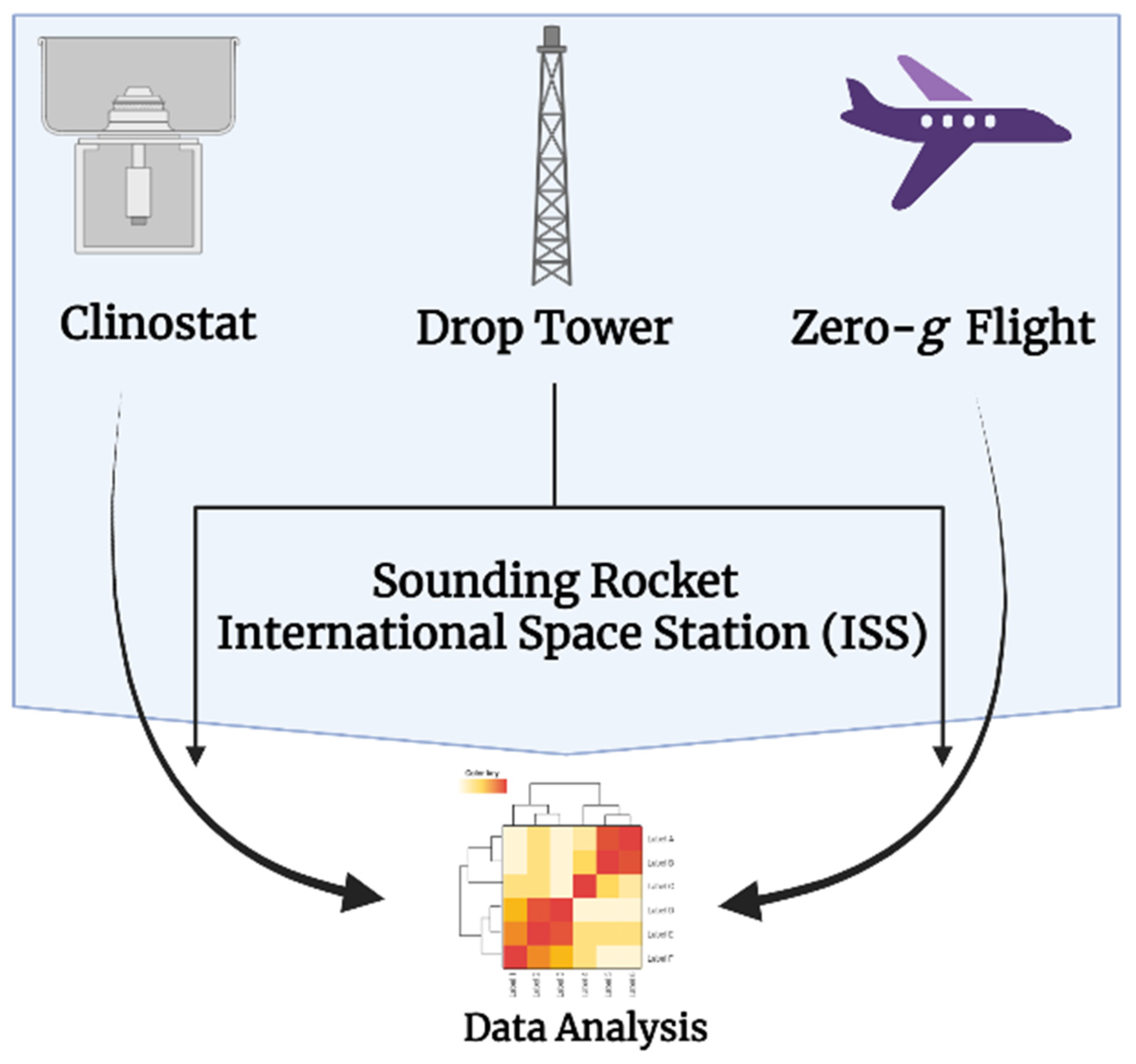{kind=link}
Abstract
:1. Introduction
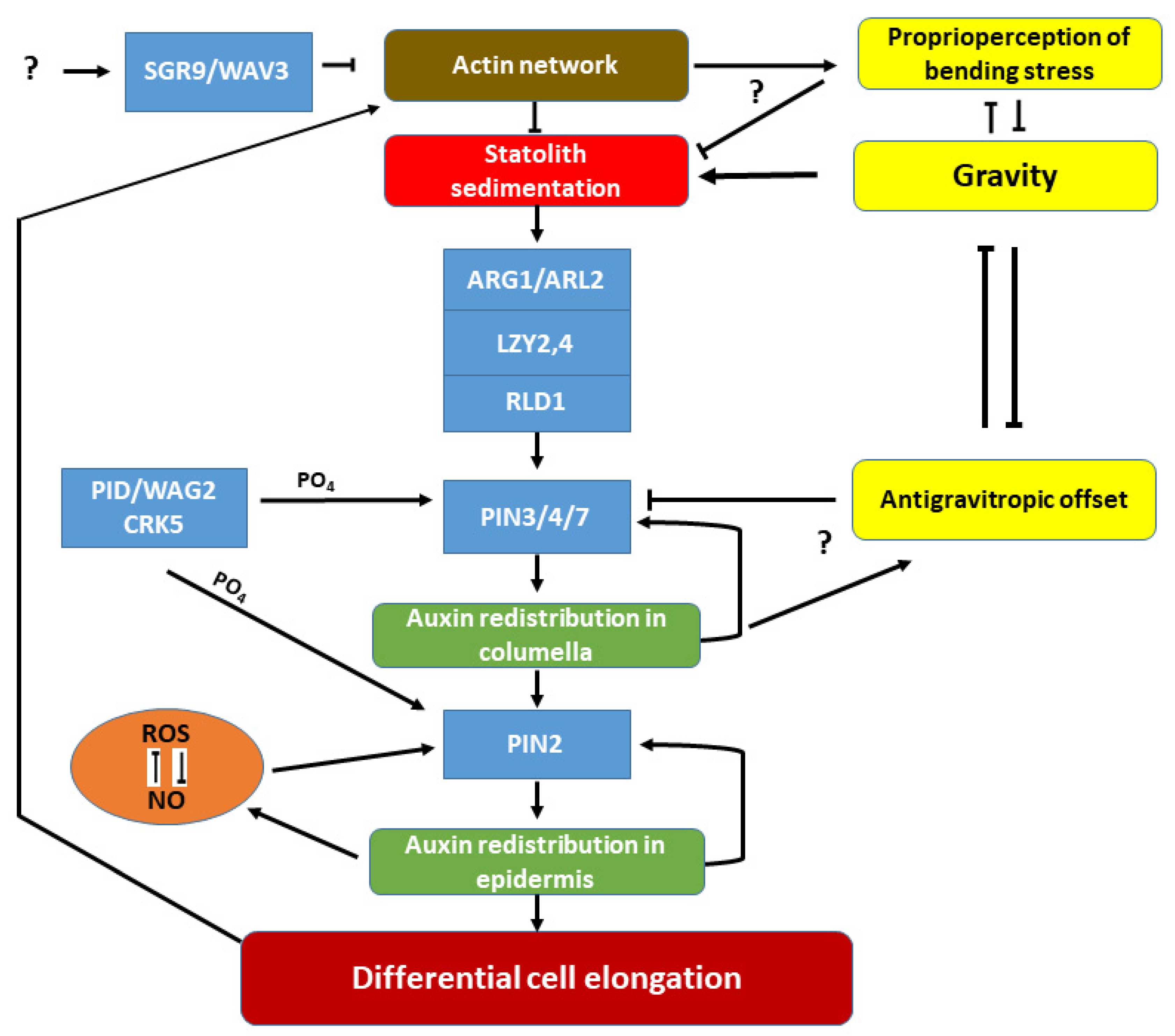
2. Molecular Aspects of Plant Gravity Responses on Earth
3. Plants in Simulated or Real Microgravity Conditions
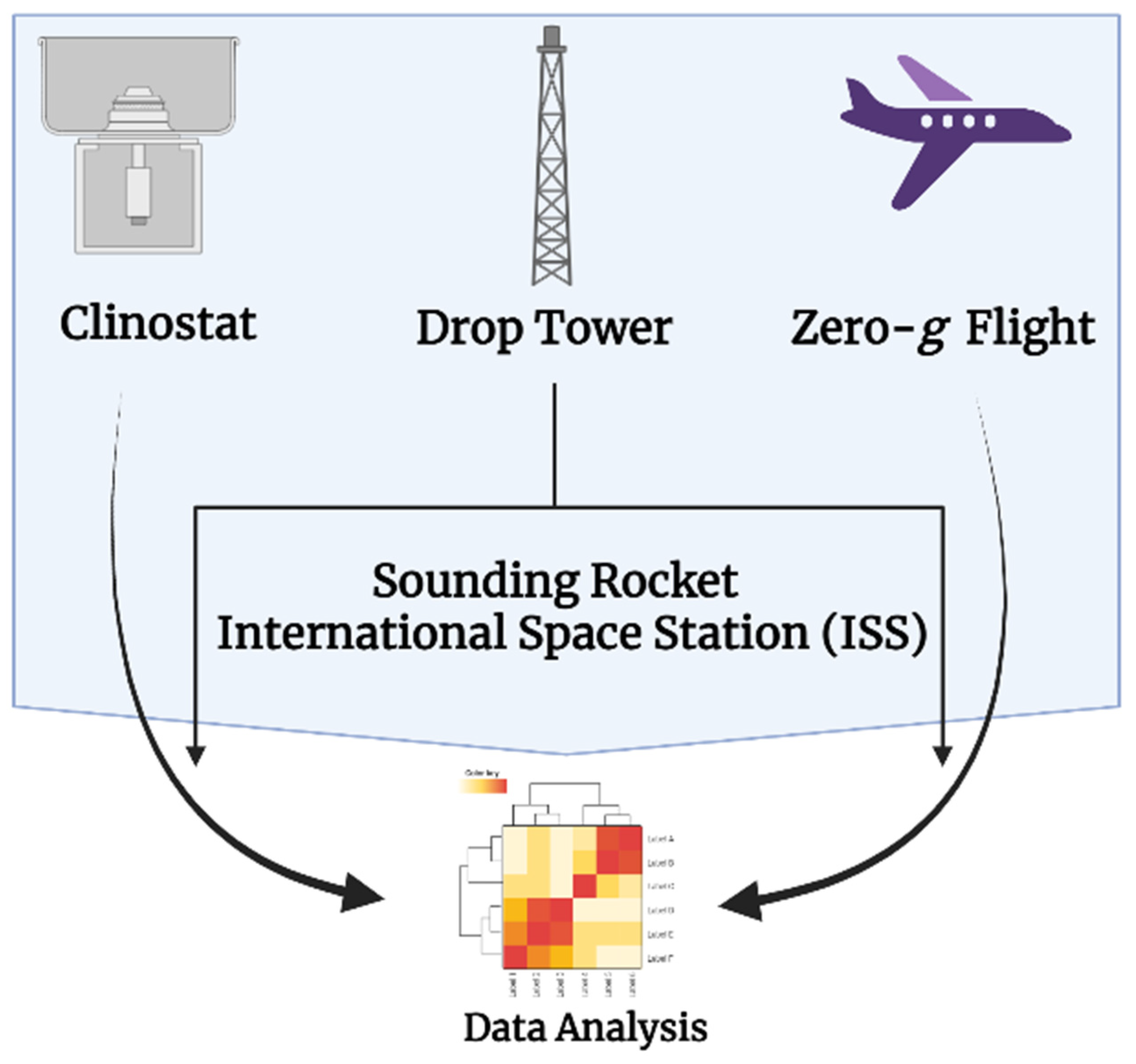
3.1. Experimental Platforms
3.1.1. Clinostats
3.1.2. Drop Towers
3.1.3. Parabolic Flights and Sounding Rockets
3.1.4. Space Flights and the International Space Station
4. Advancements in Sample and Data Analysis
5. The Reaction of Plants to Microgravity
6. Concluding Remarks and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferl, R.; Wheeler, R.; Levine, H.G.; Paul, A.-L. Plants in space. Curr. Opin. Plant Biol. 2002, 5, 258–263. [Google Scholar] [CrossRef]
- Fu, Y.; Li, L.; Xie, B.; Dong, C.; Wang, M.; Jia, B.; Shao, L.; Dong, Y.; Deng, S.; Liu, H.; et al. How to establish a bioregenerativelLife support system for long-term crewed missions to the moon or mars. Astrobiology 2016, 16, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Medina, F.J.; Manzano, A.; Villacampa, A.; Ciska, M.; Herranz, R. Understanding reduced gravity effects on early plant development before attempting life-support farming in the moon and mars. Front. Astron. Space Sci. 2021, 8, 729154. [Google Scholar] [CrossRef]
- Wheeler, R.M. Plants for human life support in space: From Myers to Mars. Gravit. Space Biol. 2010, 23, 25. [Google Scholar]
- Evans, M.L. Gravitropism: Interaction of sensitivity modulation and effector redistribution. Plant Physiol. 1991, 95, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C. The Power of Movement in Plants; John Murray Publishers: London, UK, 1880. [Google Scholar]
- Su, S.-H.; Gibbs, N.M.; Jancewicz, A.L.; Masson, P.H. Molecular mechanisms of root gravitropism. Curr. Biol. 2017, 27, R964–R972. [Google Scholar] [CrossRef] [PubMed]
- Chebli, Y.; Geitmann, A. Gravity Research on Plants: Use of single-cell experimental models. Front. Plant Sci. 2011, 28, 56. [Google Scholar] [CrossRef] [PubMed]
- Kiss, J.Z.; Wolverton, C.; Wyatt, S.E.; Hasenstein, K.H.; van Loon, J.J.W.A. Comparison of microgravity analogs to spaceflight in studies of plant growth and development. Front. Plant Sci. 2019, 10, 1577. [Google Scholar] [CrossRef]
- Simmons, C.; Migliaccio, F.; Masson, P.; Caspar, T.; Soll, D. A novel root gravitropism mutant of Arabidopsis thaliana exhibiting altered auxin Physiology. Physiol. Plant 1995, 93, 790–798. [Google Scholar] [CrossRef]
- Sato, E.M.; Hijazi, H.; Bennett, M.J.; Vissenberg, K.; Swarup, R. New insights into root gravitropic signalling. J. Exp. Bot. 2015, 66, 2155–2165. [Google Scholar] [CrossRef]
- Kiss, J.Z.; Millar, K.D.L.; Edelmann, R.E. Phototropism of Arabidopsis thaliana in microgravity and fractional gravity on the International Space Station. Planta 2012, 236, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Rosen, E.; Masson, P.H. Gravitropism in higher plants. Plant Physiol. 1999, 120, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Estelle, M. Plant Tropisms: The Ins and outs of auxin. Curr. Biol. 1996, 6, 1589–1591. [Google Scholar] [CrossRef]
- Digby, J.; Firn, R.D. The gravitropic set-point angle (GSA): The identification of an important developmentally controlled variable governing plant architecture. Plant Cell Environ. 1995, 18, 1434–1440. [Google Scholar] [CrossRef]
- Roychoudhry, S.; Kieffer, M.; Del Bianco, M.; Liao, C.Y.; Weijers, D.; Kepinski, S. The developmental and environmental regulation of gravitropic setpoint angle in Arabidopsis and bean. Sci. Rep. 2017, 7, 42664. [Google Scholar] [CrossRef]
- Sack, F.D. Plant gravity sensing. Intern. Rev. Cytol. 1991, 127, 193–252. [Google Scholar] [CrossRef]
- Sack, F.D. Plastids and gravitropic sensing. Planta 1997, 203, S63–S68. [Google Scholar] [CrossRef]
- Kiss, J.Z. Mechanisms of the early phases of plant gravitropism. Crit. Rev. Plant Sci. 2000, 19, 551–573. [Google Scholar] [CrossRef]
- Wayne, R.; Staves, M.P. A down to earth model of gravisensing or Newton’s law of gravitation from the apple’s perspective. Physiol. Plant 1996, 98, 917–921. [Google Scholar] [CrossRef]
- Casper, T.; Pickard, B.G. Gravitropism in a starchless mutant of Arabidopsis: Implications for the starch-statolith theory of gravity sensing. Planta 1989, 177, 185–197. [Google Scholar] [CrossRef]
- Moulia, B.; Douady, S.; Hamant, O. Fluctuations shape plants through proprioception. Science 2021, 372, eabc6868. [Google Scholar] [CrossRef] [PubMed]
- Cholodny, N. Wuchshormone und tropismen bei den pflanzen. Biol. Zent. 1927, 47, 604–626. [Google Scholar]
- Went, F. On growth-accelerating susbtances in the coleoptile of Avena sativa. Proc. K. Ned. Acad. Wet. 1926, 30, 10–19. [Google Scholar]
- Young, L.M.; Evans, M.L.; Hertel, R. Correlations between gravitropic curvature and auxin movement across gravistimulated roots of Zea Mays. Plant Physiol. 1990, 92, 792–796. [Google Scholar] [CrossRef]
- Rashotte, A.M.; Brady, S.R.; Reed, R.C.; Ante, S.J.; Muday, G.K. Basipetal auxin transport is required for gravitropism in roots of Arabidopsis. Plant Physiol. 2000, 122, 481–490. [Google Scholar] [CrossRef]
- Brown, D.E.; Rashotte, A.M.; Murphy, A.S.; Normanly, J.; Tague, B.W.; Peer, W.A.; Taiz, L.; Muday, G.K. Flavonoids act as negative regulators of auxin transport in vivo in Arabidopsis. Plant Physiol. 2001, 126, 524–535. [Google Scholar] [CrossRef]
- Buer, C.S.; Muday, G.K. The Transparent testa4 mutation prevents flavonoid synthesis and alters auxin transport and the response of Arabidopsis roots to gravity and light. Plant Cell 2004, 16, 1191–1205. [Google Scholar] [CrossRef]
- Lewis, D.R.; Ramirez, M.V.; Miller, N.D.; Vallabhaneni, P.; Ray, W.K.; Helm, R.F.; Winkel, B.S.J.; Muday, G.K. Auxin and ethylene induce flavonol accumulation through distinct transcriptional networks. Plant Physiol. 2011, 156, 144–164. [Google Scholar] [CrossRef]
- Blakeslee, J.J.; Peer, W.A.; Murphy, A.S. Auxin transport. Curr. Opin. Plant Biol. 2005, 8, 494–500. [Google Scholar] [CrossRef]
- Konstantinova, N.; Korbei, B.; Luschnig, C. Auxin and root gravitropism: Addressing basic cellular processes by exploiting a defined growth response. Int. J. Mol. Sci. 2021, 22, 2749. [Google Scholar] [CrossRef]
- Adamowski, M.; Friml, J. PIN-dependent auxin transport: Action, regulation, and evolution. Plant Cell 2015, 27, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Michniewicz, M.; Brewer, P.B.; Friml, J.Í. Polar auxin transport and asymmetric auxin distribution. Arab. Book 2007, 5, e0108. [Google Scholar] [CrossRef]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samakovli, D.; Roka, L.; Dimopoulou, A.; Plitsi, P.K.; Žukauskaitė, A.; Georgopoulou, P.; Novák, O.; Milioni, D.; Hatzopoulos, P. HSP90 affects root growth in Arabidopsis by regulating the polar distribution of PIN1. New Phytol. 2021, 231, 1814–1831. [Google Scholar] [CrossRef]
- Wang, H.Z.; Yang, K.Z.; Zou, J.J.; Zhu, L.L.; Xie, Z.D.; Morita, M.T.; Tasaka, M.; Friml, J.; Grotewold, E.; Beeckman, T.; et al. Transcriptional regulation of PIN genes by FOUR LIPS and MYB88 during Arabidopsis root gravitropism. Nat. Commun. 2015, 6, 8822. [Google Scholar] [CrossRef]
- Rigó, G.; Ayaydin, F.; Tietz, O.; Zsigmond, L.; Kovács, H.; Páy, A.; Salchert, K.; Darula, Z.; Medzihradszky, K.F.; Szabados, L.; et al. Inactivation of plasma membrane–localized CDPK-RELATED KINASE5 decelerates PIN2 exocytosis and root gravitropic response in Arabidopsis. Plant Cell 2013, 25, 1592–1608. [Google Scholar] [CrossRef]
- Cséplő, Á.; Zsigmond, L.; Andrási, N.; Baba, A.I.; Labhane, N.M.; Pető, A.; Kolbert, Z.; Kovács, H.E.; Steinbach, G.; Szabados, L.; et al. The AtCRK5 protein kinase is required to maintain the ROS NO balance affecting the PIN2-mediated root gravitropic response in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 5979. [Google Scholar] [CrossRef]
- Kleine-Vehn, J.; Friml, J. Polar Targeting and endocytic recycling in auxin-dependent plant development. Annu. Rev. Cell Dev. Biol. 2008, 24, 447–473. [Google Scholar] [CrossRef]
- Baster, P.; Robert, S.; Kleine-Vehn, J.; Vanneste, S.; Kania, U.; Grunewald, W.; De Rybel, B.; Beeckman, T.; Friml, J. SCF(TIR1/AFB)-auxin signalling regulates PIN vacuolar trafficking and auxin fluxes during root gravitropism. EMBO J. 2013, 32, 260–274. [Google Scholar] [CrossRef]
- Gibson, C.L.; Isley, J.W.; Falbel, T.G.; Mattox, C.T.; Lewis, D.R.; Metcalf, K.E.; Muday, G.K. A conditional mutation in SCD1 reveals linkage between PIN protein trafficking, auxin transport, gravitropism, and lateral root initiation. Front. Plant Sci. 2020, 11, 910. [Google Scholar] [CrossRef]
- Kleine-Vehn, J.; Ding, Z.; Jones, A.R.; Tasaka, M.; Morita, M.T.; Friml, J. Gravity-induced PIN transcytosis for polarization of auxin fluxes in gravity-sensing root cells. Proc. Natl. Acad. Sci. USA 2010, 107, 22344–22349. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, I.C.R.; Schwechheimer, C. Dynamic control of auxin transport-dependent growth by AGCVIII protein kinases. Curr. Opin. Plant Biol. 2014, 22, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, I.C.R.; Hammes, U.Z.; Schwechheimer, C. Activation and polarity control of PINFORMED auxin transporters by phosphorylation. Trends Plant Sci. 2018, 23, 523–538. [Google Scholar] [CrossRef]
- Baba, A.I.; Rigó, G.; Ayaydin, F.; Rehman, A.U.; Andrási, N.; Zsigmond, L.; Valkai, I.; Urbancsok, J.; Vass, I.; Pasternak, T.; et al. Functional Analysis of the Arabidopsis thaliana CDPK-Related Kinase Family: AtCRK1 Regulates Responses to Continuous Light. Int. J. Mol. Sci. 2018, 19, 1282. [Google Scholar] [CrossRef]
- Baba, A.I.; Andrási, N.; Valkai, I.; Gorcsa, T.; Koczka, L.; Darula, Z.; Medzihradszky, K.F.; Szabados, L.; Fehér, A.; Rigó, G.; et al. AtCRK5 protein kinase exhibits a regulatory role in hypocotyl hook development during skotomorphogenesis. Int. J. Mol. Sci. 2019, 20, 3432. [Google Scholar] [CrossRef]
- Baba, A.I.; Rigó, G.; Andrási, N.; Tietz, O.; Palme, K.; Szabados, L.; Cséplő, Á. Striving Towards Abiotic Stresses: Role of the Plant CDPK Superfamily Members. In International Climate Protection; Palocz-Andresen, M., Szalay, D., Gosztom, A., Sípos, L., Taligás, T., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 99–105. [Google Scholar] [CrossRef]
- Baba, A.I.; Valkai, I.; Labhane, N.; Koczka, L.; Andrási, N.; Klement, E.; Darula, Z.; Medzihradszky, K.; Szabados, L.; Fehér, A.; et al. CRK5 protein kinase contributes to the progression of embryogenesis of Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 6120. [Google Scholar] [CrossRef]
- Grones, P.; Abas, M.; Hajný, J.; Jones, A.; Waidmann, S.; Kleine-Vehn, J.; Friml, J. (2018). PID/WAG-mediated phosphorylation of the Arabidopsis PIN3 auxin transporter mediates polarity switches during gravitropism. Sci. Rep. 2018, 8, 10279. [Google Scholar] [CrossRef]
- Nakamura, M.; Nishimura, T.; Morita, M.T. Bridging the gap between amyloplasts and directional auxin transport in plant gravitropism. Curr. Opin. Plant Biol. 2019, 52, 54–60. [Google Scholar] [CrossRef]
- Sukumar, P.; Edwards, K.S.; Rahman, A.; Delong, A.; Muday, G.K. PINOID kinase regulates root gravitropism through modulation of PIN2-dependent basipetal auxin transport in Arabidopsis. Plant Physiol. 2009, 150, 722–735. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Neill, S.J.; Tang, Z.; Cai, W. Nitric oxide mediates gravitropic bending in soybean roots. Plant Physiol. 2005, 137, 663–670. [Google Scholar] [CrossRef]
- Joo, J.H.; Bae, Y.S.; Lee, J.S. Role of auxin-induced reactive oxygen species in root gravitropism. Plant Physiol. 2001, 126, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- París, R.; Vazquez, M.M.; Graziano, M.; Terrile, M.C.; Miller, N.D.; Spalding, E.P.; Otegui, M.S.; Casalongué, C.A. Distribution of endogenous NO regulates early gravitropic response and PIN2 localization in Arabidopsis roots. Front. Plant Sci. 2018, 9, 495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwiewka, M.; Bielach, A.; Tamizhselvan, P.; Madhavan, S.; Ryad, E.E.; Tan, S.; Hrtyan, M.; Dobrev, P.; Vankovï, R.; Friml, J.; et al. Root adaptation to H2O2-induced oxidative stress by ARF-GEF BEN1- and cytoskeleton-mediated PIN2 trafficking. Plant Cell Physiol. 2019, 60, 255–273. [Google Scholar] [CrossRef] [PubMed]
- Gadalla, D.S.; Braun, M.; Böhmer, M. Gravitropism in Higher Plants: Cellular Aspects. In Gravitational Biology I. SpringerBriefs in Space Life Sciences; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Hensel, W. Gravi- and Phototropism of higher plants. In Progress in Botany; Behnke, H.-D., Esser, K., Kubitzki, K., Runge, M., Ziegler, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1986; pp. 205–214. [Google Scholar] [CrossRef]
- Wendt, M.; Kuo-Huang, L.-L.; Sievers, A. Gravitropic bending of cress roots without contact between amyloplasts and complexes of endoplasmic reticulum. Planta 1987, 172, 321–329. [Google Scholar] [CrossRef] [PubMed]
- White, R.G.; Sack, F.D. Actin microfilaments in presumptive statocytes of root caps and coleoptiles. Am. J. Bot. 1990, 77, 17–26. [Google Scholar] [CrossRef]
- Baluska, F.; Hasenstein, K.H. Root cytoskeleton: Its role in perception of and response to gravity. Planta 1997, 203, S69–S78. [Google Scholar] [CrossRef]
- Blancaflor, E.B.; Hasenstein, K.H. The Organization of the actin cytoskeleton in vertical and graviresponding primary roots of maize. Plant Physiol. 1997, 113, 1447–1455. [Google Scholar] [CrossRef]
- Collings, D.A.; Zsuppan, G.; Allen, N.S.; Blancaflor, E.B. Demonstration of prominent actin filaments in the root columella. Planta 2001, 212, 392–403. [Google Scholar] [CrossRef]
- Friedman, H.; Meir, S.; Halevy, A.H.; Philosoph-Hadas, S. Inhibition of the gravitropic bending response of flowering shoots by salicylic acid. Plant Sci. 2003, 165, 905–911. [Google Scholar] [CrossRef]
- Livanos, P.; Galatis, B.; Apostolakos, P. The interplay between ROS and tubulin cytoskeleton in plants. Plant Signal. Behav. 2014, 9, e28069. [Google Scholar] [CrossRef]
- Blancaflor, E.B. Regulation of plant gravity sensing and signaling by the actin cytoskeleton. Am. J. Bot. 2013, 100, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Toyota, M.; Tasaka, M.; Morita, M.T. An Arabidopsis E3 ligase, SHOOT GRAVITROPISM9, modulates the interaction between statoliths and F-actin in gravity sensing. Plant Cell 2011, 23, 1830–1848. [Google Scholar] [CrossRef] [PubMed]
- Sakai, T.; Mochizuki, S.; Haga, K.; Uehara, Y.; Suzuki, A.; Harada, A.; Wada, T.; Ishiguro, S.; Okada, K. The WAVY GROWTH 3 E3 ligase family controls the gravitropic response in Arabidopsis roots: WAV3 family role in root gravitropism. Plant J. 2012, 70, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, M.; Furutani, M.; Nishimura, T.; Nakamura, M.; Fushita, T.; Iijima, K.; Baba, K.; Tanaka, H.; Toyota, M.; Tasaka, M.; et al. The Arabidopsis LAZY1 family plays a key role in gravity signaling within statocytes and in branch angle control of roots and shoots. Plant Cell 2017, 29, 1984–1999. [Google Scholar] [CrossRef] [PubMed]
- Furutani, M.; Morita, M.T. LAZY1-LIKE-mediated gravity signaling pathway in root gravitropic set-point angle control. Plant Physiol. 2021, 187, 1087–1095. [Google Scholar] [CrossRef]
- Mullen, J.L.; Hangarter, R.P. Genetic analysis of the gravitropic set-point angle in lateral roots of Arabidopsis. Adv. Space Res. 2003, 31, 2229–2236. [Google Scholar] [CrossRef]
- Roychoudhry, S.; Del Bianco, M.; Kieffer, M.; Kepinski, S. Auxin controls gravitropic setpoint angle in higher plant lateral branches. Curr. Biol. 2013, 23, 1497–1504. [Google Scholar] [CrossRef]
- Roychoudhry, S.; Kepinski, S. Shoot and root branch growth angle control-the wonderfulness of lateralness. Curr. Opin. Plant Biol. 2015, 23, 124–131. [Google Scholar] [CrossRef]
- Rosquete, M.R.; von Wangenheim, D.; Marhavý, P.; Barbez, E.; Stelzer, E.H.; Benková, E.; Maizel, A.; Kleine-Vehn, J. An auxin transport mechanism restricts positive orthogravitropism in lateral roots. Curr. Biol. 2013, 23, 817–822. [Google Scholar] [CrossRef]
- Furutani, M.; Hirano, Y.; Nishimura, T.; Nakamura, M.; Taniguchi, M.; Suzuki, K.; Oshida, R.; Kondo, C.; Sun, S.; Kato, K.; et al. Polar recruitment of RLD by LAZY1-like protein during gravity signaling in root branch angle control. Nat. Commun. 2020, 11, 76. [Google Scholar] [CrossRef] [Green Version]
- Harrison, B.R.; Masson, P.H. ARL2, ARG1 and PIN3 define a gravity signal transduction pathway in root statocytes: ARL2 and ARG1 modulate gravity signal transduction. Plant J. 2007, 53, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, K.L.; Strohm, A.K.; Masson, P.H. Gravity sensing and signal transduction in vascular plant primary roots. Am. J. Bot. 2013, 100, 126–142. [Google Scholar] [CrossRef] [PubMed]
- Sedbrook, J.; Boonsirichai, K.; Chen, R.; Hilson, P.; Pearlman, R.; Rosen, E.; Rutherford, R.; Batiza, A.; Carroll, K.; Schulz, T.; et al. Molecular genetics of root gravitropism and waving in Arabidopsis thaliana. Gravit Space Biol Bull. 1998, 11, 71–78. [Google Scholar] [PubMed]
- Porterfield, D.M.; Neichitailo, G.S.; Mashinski, A.L.; Musgrave, M.E. Spaceflight hardware for conducting plant growth experiments in space: The early years 1960–2000. Adv. Space Res. 2003, 31, 183–193. [Google Scholar] [CrossRef]
- Harvey, B.; Zakutnyaya, O. Russian Space Probes: Scientific Discoveries and Future Missions; Springer-Praxis books in space exploration; Springer: New York, NY, USA, 2011; pp. 301–374. [Google Scholar]
- Vandenbrink, J.P.; Kiss, J.Z. Space, the Final Frontier: A critical review of recent experiments performed in microgravity. Plant Sci. 2016, 243, 115–119. [Google Scholar] [CrossRef]
- Sathasivam, M.; Hosamani, R.; Swamy, B.K.; Kumaran, G.S. Plant responses to real and simulated microgravity. Life Sci. Space Res. 2021, 28, 74–86. [Google Scholar] [CrossRef]
- Wolff, S.; Coelho, L.; Karoliussen, I.; Jost, A.I. Effects of the extraterrestrial environment on plants: Recommendations for future space experiments for the MELiSSA higher plant compartment. Life 2014, 4, 189–204. [Google Scholar] [CrossRef]
- Kiss, J.Z.; Aanes, G.; Schiefloe, M.; Coelho, L.H.F.; Millar, K.D.L.; Edelmann, R.E. Changes in operational procedures to improve spaceflight experiments in plant biology in the European modular cultivation system. Adv. Space Res. 2014, 53, 818–827. [Google Scholar] [CrossRef]
- Braun, M.; Böhmer, M.; Häderc, D.P.; Hemmersbach, R.; Palme, K. Gravitational Biology I: Gravity Sensing and Graviorientation in Microorganisms and Plants, 1st ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2018. [Google Scholar]
- Vandenbrink, J.P.; Kiss, J.Z.; Herranz, R.; Medina, F.J. Light and gravity signals synergize in modulating plant development. Front. Plant Sci. 2014, 5, 563. [Google Scholar] [CrossRef]
- Sachs, J. Über orthotrope und plagiotrope Pflanzenteile. Arb. Bot. Inst. Wurzburg. 1882, 2, 226–284. [Google Scholar]
- Kamal, K.Y.; Herranz, R.; van Loon, J.J.W.A.; Christianen, P.C.M.; Medina, F.J. Evaluation of simulated microgravity environments induced by diamagnetic levitation of plant cell suspension cultures. Microgravity Sci. Technol. 2016, 28, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Hasenstein, K.H.; van Loon, J.J.W.A. Clinostats and other rotating systems—design, function, and limitations. In Generation and Applications of Extra-Terrestrial Environments on Earth; Beysens, D.A., van Loon, J.J.W.A., Eds.; River Publishers: Aalborg, Denmark, 2015; pp. 147–156. [Google Scholar]
- Hemmersbach, R.; von der Wiesche, M.; Seibt, D. Ground-based experimental platforms in gravitational biology and human physiology. Signal Transduct. 2006, 6, 381–387. [Google Scholar] [CrossRef]
- Hoson, T.; Kamisaka, S.; Masuda, Y.; Yamashita, M.; Buchen, B. Evaluation of the three-dimensional clinostat as a simulator of weightlessness. Planta 1997, 203, S187–S197. [Google Scholar] [CrossRef] [PubMed]
- Kraft, T.F.B.; van Loon, J.J.W.A.; Kiss, J.Z. Plastid position in Arabidopsis columella cells is similar in microgravity and on a Random-Positioning Machine. Planta 2000, 211, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.M.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.A.; Lebert, M.; et al. Ground-based facilities for simulation of microgravity: Organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef]
- Huang, B.; Li, D.G.; Huang, Y. Effects of spaceflight and simulated microgravity on microbial growth and secondary metabolism. Military Med Res. 2018, 5, 18. [Google Scholar] [CrossRef]
- Dedolph, R.R.; Dipert, M.H. The physical basis of gravity stimulus nullification by clinostat rotation. Plant Physiol. 1971, 47, 756–764. [Google Scholar] [CrossRef]
- John, S.P.; Hasenstein, K.H. Effects of mechanostimulation on gravitropism and signal persistence in Flax roots. Plant Signal. Behav. 2011, 6, 1365–1370. [Google Scholar] [CrossRef]
- Böhmer, M.; Schleiff, E. Microgravity research in plants A range of platforms and options allow research on plants in zero or low gravity that can yield important insights into plant physiology. EMBO Rep. 2019, 20, e48541. [Google Scholar] [CrossRef]
- Pletser, V.; Russomano, T. Research in Microgravity in Physical and Life Sciences: An Introduction to Means and Methods. In Preparation of Space Experiments; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Limbach, C.; Hauslage, J.; Schäfer, C.; Braun, M. How to activate a plant gravireceptor. Early mechanisms of gravity sensing studied in Characean rhizoids during parabolic flights. Plant Physiol. 2005, 139, 1030–1040. [Google Scholar] [CrossRef] [Green Version]
- Halstead, T.W.; Dutcher, F.R. Plants in space. Annu. Rev. Plant. Physiol. 1987, 38, 317–345. [Google Scholar] [CrossRef] [PubMed]
- Kiss, J.Z. Conducting plant experiments in space. In Plant Gravitropism; Methods in Molecular Biology; Blancaflor, E.B., Ed.; Springer: New York, NY, USA, 2015; Volume 1309, pp. 255–283. [Google Scholar] [CrossRef]
- Carillo, P.; Morrone, B.; Fusco, G.M.; De Pascale, S.; Rouphael, Y. Challenges for a sustainable food production system on board of the International Space Station: A technical review. Agronomy 2020, 10, 687. [Google Scholar] [CrossRef]
- Zabel, P.; Bamsey, M.; Schubert, D.; Tajmar, M. Review and analysis of over 40 years of space plant growth systems. Life Sci. Space Res. 2016, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Khodadad, C.L.M.; Hummerick, M.E.; Spencer, L.E.; Dixit, A.R.; Richards, J.T.; Romeyn, M.W.; Smith, T.M.; Wheeler, R.M.; Massa, G.D. Microbiological and nutritional analysis of lettuce crops grown on the International Space Station. Front. Plant Sci. 2020, 11, 199. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.L.; Popp, M.P.; Gurley, W.B.; Guy, C.; Norwood, K.L.; Ferl, R.J. Arabidopsis gene expression patterns are altered during spaceflight. Adv. Space Res. 2005, 36, 1175–1181. [Google Scholar] [CrossRef]
- Paul, A.L.; Zupanska, A.K.; Ostrow, D.T.; Zhang, Y.; Sun, Y.; Li, J.-L.; Shanker, S.; Farmerie, W.G.; Amalfitano, C.E.; Ferl, R.J. Spaceflight transcriptomes: Unique responses to a novel environment. Astrobiology 2012, 12, 40–56. [Google Scholar] [CrossRef]
- Paul, A.L.; Zupanska, A.K.; Schultz, E.R.; Ferl, R.J. Organ-specific remodeling of the Arabidopsis transcriptome in response to spaceflight. BMC Plant Biol. 2013, 13, 112. [Google Scholar] [CrossRef]
- Kwon, T.; Sparks, J.A.; Nakashima, J.; Allen, S.N.; Tang, Y.; Blancaflor, E.B. Transcriptional response of Arabidopsis seedlings during spaceflight reveals peroxidase and cell wall remodeling genes associated with root hair development. Am. J. Bot. 2015, 102, 21–35. [Google Scholar] [CrossRef]
- Kruse, C.P.S.; Meyers, A.D.; Basu, P.; Hutchinson, S.; Luesse, D.R.; Wyatt, S.E. Spaceflight induces novel regulatory responses in Arabidopsis seedling as revealed by combined proteomic and transcriptomic analyses. BMC Plant Bio. 2020, 20, 237. [Google Scholar] [CrossRef]
- Zeng, D.; Cui, J.; Yin, Y.; Xiong, Y.; Liu, M.; Guan, S.; Cheng, D.; Sun, Y.; Lu, W. Metabolomics analysis in different development stages on SP0 generation of rice seeds after spaceflight. Front. Plant Sci. 2021, 12, 700267. [Google Scholar] [CrossRef]
- Paul, A.L.; Levine, H.G.; McLamb, W.; Norwood, K.L.; Reed, D.; Stutte, G.W.; William Wells, H.; Ferl, R.J. Plant molecular biology in the space station era: Utilization of KSC fixation tubes with RNAlater. Acta Astronaut. 2005, 56, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Ferl, R.J.; Zupanska, A.; Spinale, A.; Reed, D.; Manning-Roach, S.; Guerra, G.; Cox, D.R.; Paul, A.-L. The performance of KSC fixation tubes with RNALater for orbital experiments: A case study in ISS operations for molecular biology. Adv. Space Res. 2011, 4, 199–206. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Mazzucato, A.; Olimpieri, I. Growth, yield and reproduction of Dwarf tomato grown under simulated microgravity conditions. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2007, 141, 75–81. [Google Scholar] [CrossRef]
- Zheng, H.Q.; Han, F.; Le, J. Higher plants in space: Microgravity perception, response, and adaptation. Microgravity Sci. Technol. 2015, 27, 377–386. [Google Scholar] [CrossRef]
- Merkys, A.J.; Laurinavičius, R.S.; Kenstavičien, P.F.; Nečitailo, G.S. formation and growth of callus tissue of Arabidopsis under changed gravity. Adv. Space Res. 1989, 9, 37–40. [Google Scholar] [CrossRef]
- Levine, H.G.; Krikorian, A.D. Shoot growth in aseptically cultivated Daylily and Haplopappus plantlets after a 5-day spaceflight. Physiol Plant 1992, 86, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Salmi, M.L.; Roux, S.J. Gene expression changes induced by space flight in single-cells of the fern Ceratopteris richardii. Planta 2008, 229, 151–159. [Google Scholar] [CrossRef]
- Hauslage, J.; Görög, M.; Krause, L.; Schüler, O.; Schäfer, M.; Witten, A.; Kesseler, L.; Böhmer, M.; Hemmersbach, R. ARABIDOMICS—A new experimental platform for molecular analyses of plants in drop towers, on parabolic flights, and sounding rockets. Rev. Sci. Instrum. 2020, 91, 034504. [Google Scholar] [CrossRef]
- Watanabe, C.; Fujii, N.; Yanai, K.; Hotta, T.; Kim, D.-H.; Kamada, M.; Sasagawa-Saito, Y.; Nishimura, T.; Koshiba, T.; Miyazawa, Y.; et al. Gravistimulation changes the accumulation pattern of the CsPIN1 auxin efflux facilitator in the endodermis of the transition zone in cucumber seedlings. Plant Physiol. 2012, 158, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, C.; Fujii, N.; Miyazawa, Y.; Kamada, M.; Kasahara, H.; Osada, I.; Shimazu, T.; Fusejima, Y.; Higashibata, A.; Yamazaki, T.; et al. The gravity-induced re-localization of auxin efflux carrier CsPIN1 in cucumber seedlings: Spaceflight experiments for immunohistochemical microscopy. npj Microgravity 2016, 2, 16030. [Google Scholar] [CrossRef]
- Oka, M.; Kamada, M.; Inoue, R.; Miyamoto, K.; Uheda, E.; Yamazaki, C.; Shimazu, T.; Sano, H.; Kasahara, H.; Suzuki, T.; et al. Altered localisation of ZmPIN1a proteins in plasma membranes responsible for enhanced-polar auxin transport in etiolated maize seedlings under microgravity conditions in space. Funct. Plant Biol. FPB 2020, 47, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Matía, I.; González-Camacho, F.; Herranz, R.; Kiss, J.Z.; Gasset, G.; van Loon, J.J.W.A.; Marco, R.; Javier Medina, F. Plant cell proliferation and growth are altered by microgravity conditions in spaceflight. J. Plant Physiol. 2010, 167, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.M.; Subramanian, A.; Edelmann, R.E.; Kiss, J.Z. Morphometric analyses of petioles of seedlings grown in a spaceflight experiment. J Plant Res. 2015, 128, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Correll, M.J.; Pyle, T.P.; Millar, K.D.L.; Sun, Y.; Yao, J.; Edelmann, R.E.; Kiss, J.Z. Transcriptome analyses of Arabidopsis thaliana seedlings grown in space: Implications for gravity-responsive genes. Planta 2013, 238, 519–533. [Google Scholar] [CrossRef]
- Paul, A.L.; Sng, N.J.; Zupanska, A.K.; Krishnamurthy, A.; Schultz, E.R.; Ferl, R.J. Genetic dissection of the Arabidopsis spaceflight transcriptome: Are some responses dispensable for the physiological adaptation of plants to spaceflight? PLoS ONE 2017, 12, e0180186. [Google Scholar] [CrossRef]
- Choi, W.G.; Barker, R.J.; Kim, S.-H.; Swanson, S.J.; Gilroy, S. Variation in the transcriptome of different ecotypes of Arabidopsis thaliana reveals signatures of oxidative stress in plant responses to spaceflight. Am. J. Bot. 2019, 106, 123–136. [Google Scholar] [CrossRef]
- Paul, A.L.; Amalfitano, C.E.; Ferl, R.J. Plant Growth strategies are remodeled by spaceflight. BMC Plant Biol. 2012, 12, 232. [Google Scholar] [CrossRef]
- Kozeko, L.Y.; Buy, D.D.; Pirko, Y.V.; Blume, Y.B.; Kordyum, E.L. Clinorotation affects induction of the heat shock response in Arabidopsis thaliana seedlings. Gravit. Space Res. 2018, 6, 2–9. [Google Scholar] [CrossRef]
- Aubry-Hivet, D.; Nziengui, H.; Rapp, K.; Oliveira, O.; Paponov, I.A.; Li, Y.; Hauslage, J.; Vagt, N.; Braun, M.; Ditengou, F.A.; et al. Analysis of gene expression during parabolic flights reveals distinct early gravity responses in Arabidopsis roots. Plant Biol. 2014, 16, 129–141. [Google Scholar] [CrossRef]
- Beisel, N.S.; Noble, J.; Barbazuk, W.B.; Paul, A.L.; Ferl, R.J. Spaceflight-induced alternative splicing during seedling development in Arabidopsis thaliana. npj Microgravity 2019, 5, 9. [Google Scholar] [CrossRef]
- Ferl, R.J.; Paul, A.L. The effect of spaceflight on the gravity-sensing auxin gradient of roots: GFP reporter gene microscopy on orbit. npj Microgravity 2016, 2, 15023. [Google Scholar] [CrossRef] [PubMed]
- Babbick, M.; Dijkstra, C.; Larkin, O.J.; Anthony, P.; Davey, M.R.; Power, J.B.; Lowe, K.C.; Cogoli-Greuterd, M.; Hamp, R. Expression of Transcription Factors after Short-Term Exposure of Arabidopsis thaliana Cell Cultures to Hypergravity and Simulated Microgravity (2-D/3-D Clinorotation, Magnetic Levitation). Adv. Space Res. 2007, 39, 1182–1189. [Google Scholar] [CrossRef]
- Sheppard, J.; Land, E.S.; Toennisson, T.A.; Doherty, C.J.; Perera, I.Y. 2021. Uncovering transcriptional responses to fractional gravity in Arabidopsis roots. Life 2021, 11, 1010. [Google Scholar] [CrossRef] [PubMed]
- Manzano, A.; Herranz, R.; den Toom, L.A.; te Slaa, S.; Borst, G.; Visser, M.; Medina, J.; van Loon, J.J. Novel, Moon and Mars, partial gravity simulation paradigms and their effects on the balance between cell growth and cell proliferation during early plant development. npj Microgravity 2018, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Kamal, K.Y.; Herranz, R.; van Loon, J.J.; Medina, F.J. Cell cycle acceleration and changes in essential nuclear functions induced by simulated microgravity in a synchronized Arabidopsis cell culture. Plant Cell Environ. 2019, 42, 480–494. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.; Hilaire, E.; Paulsen, A.Q.; Guikema, J.A. Brassica rapa plants adapted to microgravity with reduced photosystem I and its photochemical activity. Physiol. Plant. 2004, 122, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Jagtap, S.S.; Awhad, R.B.; Santosh, B.; Vidyasagar, P.B. Effects of Clinorotation on Growth and Chlorophyll Content of Rice Seeds. Microgravity Sci. Technol. 2011, 23, 41–48. [Google Scholar] [CrossRef]
- Kitaya, Y.; Kawai, M.; Tsuruyama, J.; Takahashi, H.; Tani, A.; Goto, E.; Saito, T.; Kiyota, M. The effect of gravity on surface temperature and net photosynthetic rate of plant leaves. Adv. Space Res. 2001, 28, 659–664. [Google Scholar] [CrossRef]
- Faraoni, P.; Sereni, E.; Gnerucci, A.; Cialdai, F.; Monici, M.; Ranaldi, F. Glyoxylate cycle activity in Pinus pinea seeds during germination in altered gravity conditions. Plant Physiol. Biochem. 2019, 139, 389–394. [Google Scholar] [CrossRef]
- Ranaldi, F.; Vanni, P.; Giachetti, E. Enzyme catalysis in microgravity: Steady-state kinetic analysis of the isocitrate lyase reaction. Biophys. Chem. 2003, 103, 169–177. [Google Scholar] [CrossRef]
- Chandler, J.O.; Haas, F.B.; Khan, S.; Bowden, L.; Ignatz, M.; Enfissi, E.M.; Gawthrop, F.; Griffiths, A.; Fraser, P.D.; Rensing, S.A.; et al. Rocket science: The effect of spaceflight on germination physiology, ageing, and transcriptome of Eruca sativa seeds. Life 2020, 10, 49. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Cui, J.; Yin, Y.; Zhang, M.; Shan, S.; yao Liu, M.; Cheng, D.; Lu, W.; Sun, Y. Proteomic analysis in different development stages on SP0 generation of rice seeds after space flight. Life Sci. Space Res. 2020, 26, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Villacampa, A.; Sora, L.; Herranz, R.; Medina, F.J.; Ciska, M. Analysis of gravi-response and biological effects of vertical and horizontal clinorotation in Arabidopsis thaliana root tip. Plants 2021, 10, 734. [Google Scholar] [CrossRef] [PubMed]
- Villacampa, A.; Fañanás-Pueyo, I.; Medina, F.J.; Ciska, M. Root growth direction in simulated microgravity is modulated by a light avoidance mechanism mediated by flavonols. Physiol. Plant. 2022, 174, e13722. [Google Scholar] [CrossRef] [PubMed]
- Aronne, G.; Muthert, L.W.F.; Izzo, L.G.; Romano, L.E.; Iovane, M.; Capozzi, F.; Manzano, A.; Ciska, M.; Herranz, R.; Medina, F.J.; et al. A novel device to study altered gravity and light interactions in seedling tropisms. Life Sci. Space Res. 2022, 32, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Oluwafemi, F.A.; Akpu, S.U.; Akomolafe, C.B.; Billyok, B.J.; Okhuelegbe, E.O.; Doherty, K.B.; Olubiyi, R.; Adeleke, O.; Oluwafemi, L.; Agboola, O.A. Microgravity-simulation of plant growth and its implications to the Sustainable Development Goals. Int. J. Biomed. Health Sci. 2022, 17. [Google Scholar]
- Xu, P.; Chen, H.; Hu, J.; Pang, X.; Jin, J.; Cai, W. Pectin methylesterase gene AtPMEPCRA contributes to physiological adaptation to simulated and spaceflight micro-gravity in Arabidopsis. iScience 2022, 25, 104331. [Google Scholar] [CrossRef]
- Mohammadalikhani, S.; Ghanati, F.; Hajebrahimi, Z.; Sharifi, M. Molecular and biochemical modifications of suspension-cultured tobacco cell walls after exposure to alternative gravity. Plant Physiol. Biochem. 2022, 176, 1–7. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baba, A.I.; Mir, M.Y.; Riyazuddin, R.; Cséplő, Á.; Rigó, G.; Fehér, A. Plants in Microgravity: Molecular and Technological Perspectives. Int. J. Mol. Sci. 2022, 23, 10548. https://doi.org/10.3390/ijms231810548
Baba AI, Mir MY, Riyazuddin R, Cséplő Á, Rigó G, Fehér A. Plants in Microgravity: Molecular and Technological Perspectives. International Journal of Molecular Sciences. 2022; 23(18):10548. https://doi.org/10.3390/ijms231810548
Chicago/Turabian StyleBaba, Abu Imran, Mohd Yaqub Mir, Riyazuddin Riyazuddin, Ágnes Cséplő, Gábor Rigó, and Attila Fehér. 2022. "Plants in Microgravity: Molecular and Technological Perspectives" International Journal of Molecular Sciences 23, no. 18: 10548. https://doi.org/10.3390/ijms231810548
APA StyleBaba, A. I., Mir, M. Y., Riyazuddin, R., Cséplő, Á., Rigó, G., & Fehér, A. (2022). Plants in Microgravity: Molecular and Technological Perspectives. International Journal of Molecular Sciences, 23(18), 10548. https://doi.org/10.3390/ijms231810548