
Identification of Candidate Chemosensory Gene Families by Head Transcriptomes Analysis in the Mexican Fruit Fly, Anastrepha ludens Loew (Diptera: Tephritidae)
 , , , ,
, , , ,
Abstract
1. Introduction
2. Results
2.1. Transcriptome Assembly and Annotation
2.2. Mining of Chemosensory Genes
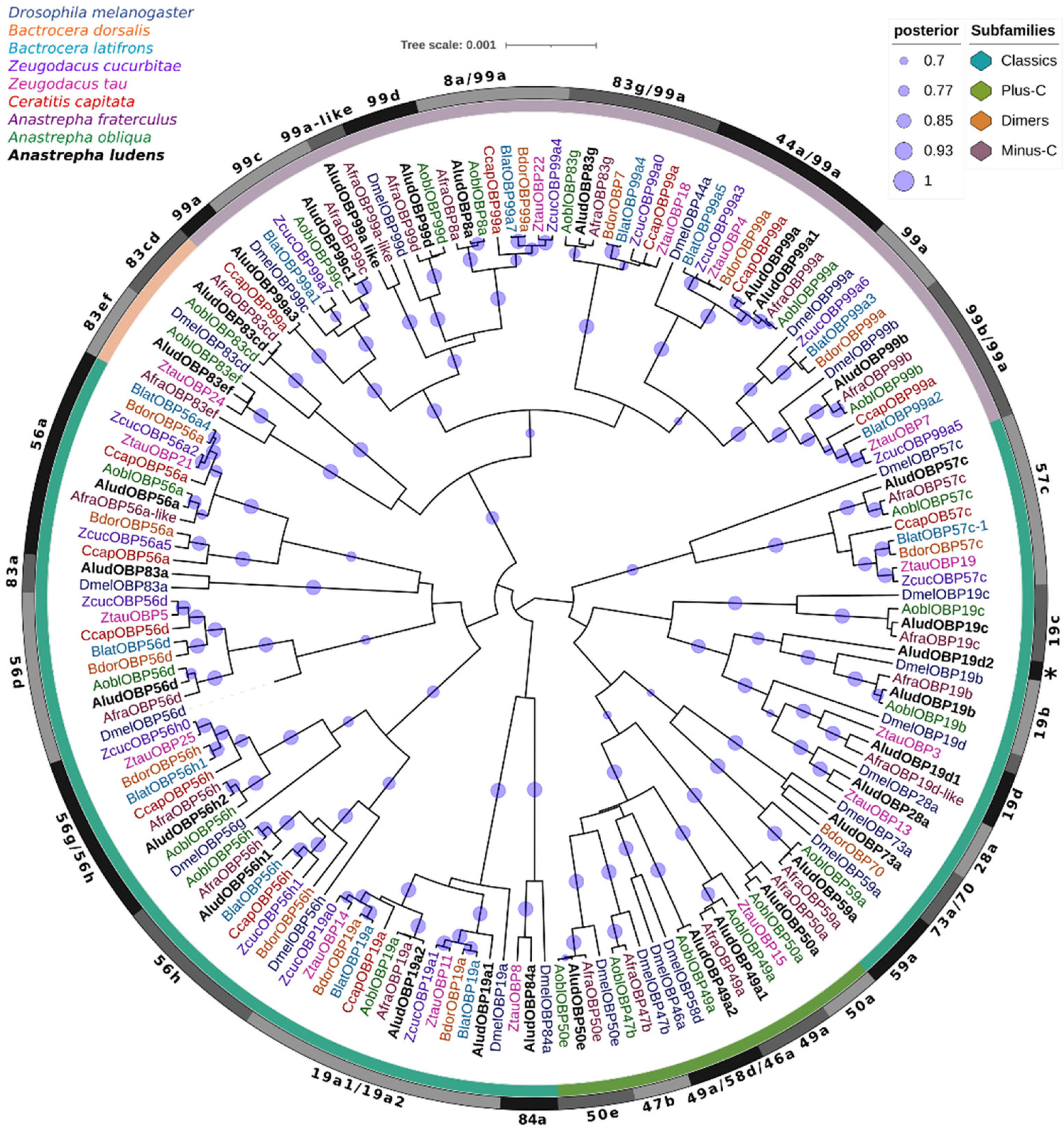
2.3. Odorant Binding Proteins (OBPs)
2.4. Chemosensory Proteins (CSPs)
2.5. Sensory Neuron Membrane Proteins (SNMPs)
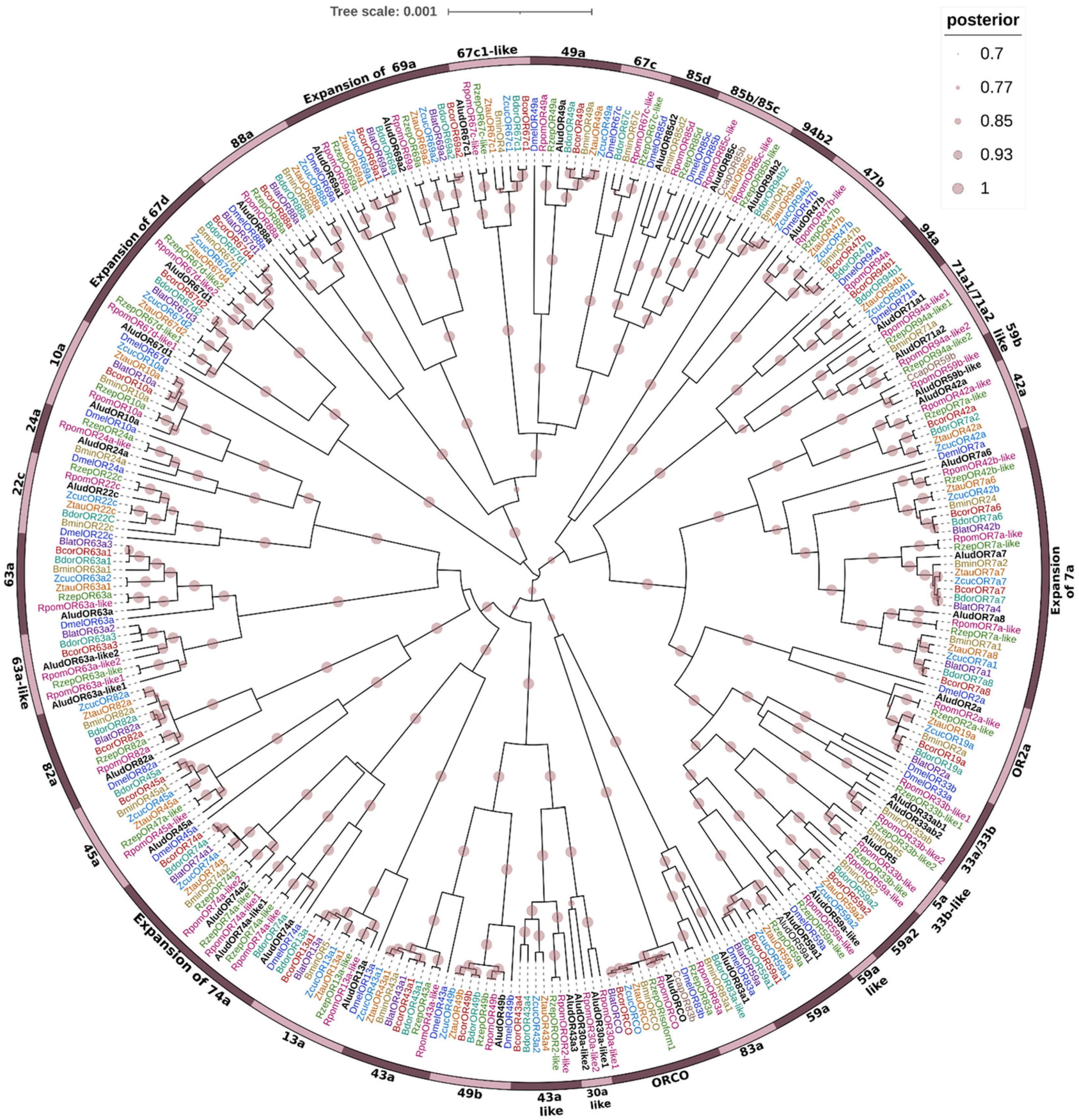
2.6. Odorant Receptors (ORs)
2.7. Gustatory Receptors (GRs)
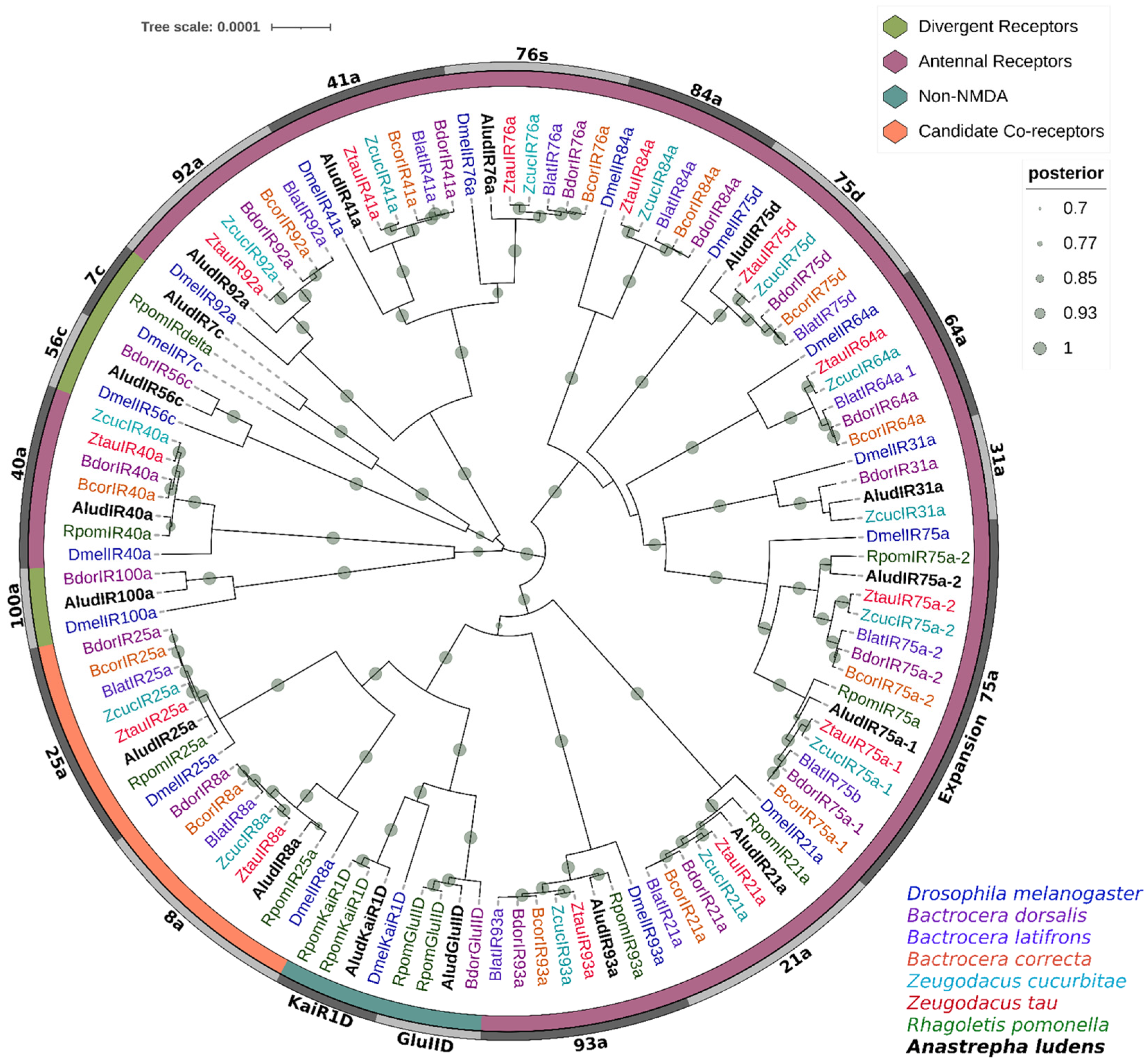
2.8. Ionotropic Receptors (IRs)
3. Discussion
4. Materials and Methods
4.1. Collection of Biological Material
4.2. Preparation and Sequencing of cDNA Libraries
4.3. Cleaning, Assembly, and Annotation of the De Novo Transcriptome
4.4. Mining and Functional Analysis of Chemosensory Genes
4.5. Sequence Comparison and Phylogenetic Analysis
4.6. Sequence Information
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hansson, B.S.; Stensmyr, M.C. Evolution of insect olfaction. Neuron 2011, 72, 698–711. [Google Scholar] [CrossRef] [PubMed]
- Sachse, S.; Krieger, J. Olfaction in insects—The primary processes of odor recognition and coding. E-Neuroforum 2011, 2, 49–60. [Google Scholar] [CrossRef]
- Tunstall, N.E.; Warr, C.G. Chemical communication in insects: The peripheral odour coding system of Drosophila melanogaster. Adv. Exp. Med. Biol. 2012, 739, 59–77. [Google Scholar]
- Leal, W.S. Odorant Reception in Insects: Roles of Receptors, Binding Proteins, and Degrading Enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef]
- Benton, R.; Vannice, K.S.; Vosshall, L.B. An essential role for a CD36-related receptor in pheromone detection in Drosophila. Nature 2007, 450, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Diaz, C.; Bargeton, B.; Abuin, L.; Bukar, N.; Reina, J.H.; Bartoi, T.; Graf, M.; Ong, H.; Ulbrich, M.H.; Masson, J.-F.; et al. A CD36 ectodomain mediates insect pheromone detection via a putative tunnelling mechanism. Nat. Commun. 2016, 7, 11866. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Iovinella, I.; Felicioli, A.; Dani, F.R. Soluble proteins of chemical communication: An overview across arthropods. Front. Physiol. 2014, 5, 320. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Challenges of Pest Management in the Twenty First Century: New Tools and Strategies to Combat Old and New Foes Alike. Front. Agron. 2019, 1, 2. [Google Scholar] [CrossRef]
- Bosa, C.F.; Cruz-López, L.; Zepeda-Cisneros, C.S.; Valle-Mora, J.; Guillén-Navarro, K.; Liedo, P. Sexual behavior and male volatile compounds in wild and mass-reared strains of the Mexican fruit fly Anastrepha ludens (Diptera: Tephritidae) held under different colony management regimes. Insect Sci. 2016, 23, 105–116. [Google Scholar] [CrossRef]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. 2017, 93, 184–200. [Google Scholar] [CrossRef]
- Dimitratos, S.D.; Hommel, A.S.; Konrad, K.D.; Simpson, L.M.; Wu-Woods, J.J.; Woods, D.F. Biosensors to monitor water quality utilizing insect odorant-binding proteins as detector elements. Biosensors 2019, 9, 62. [Google Scholar] [CrossRef] [PubMed]
- Ingham, V.A.; Anthousi, A.; Douris, V.; Harding, N.J.; Lycett, G.; Morris, M.; Vontas, J.; Ranson, H. A sensory appendage protein protects malaria vectors from pyrethroids. Nature 2019, 577, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Portilla-Pulido, J.S.; Castillo-Morales, R.M.; Barón-Rodríguez, M.A.; Duque, J.E.; Mendez-Sanchez, S.C. Design of a repellent against Aedes aegypti (Diptera: Culicidae) using in silico simulations with AaegOBP1 protein. J. Med. Entomol. 2019, 57, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Cheema, J.A.; Carraher, C.; Plank, N.O.V.; Travas-Sejdic, J.; Kralicek, A. Insect odorant receptor-based biosensors: Current status and prospects. Biotechnol. Adv. 2021, 53, 107840. [Google Scholar] [CrossRef]
- Chen, X.; Lei, Y.; Li, H.; Xu, L.; Yang, H.; Wang, J.; Jiang, H. CRISPR/Cas9 mutagenesis abolishes odorant-binding protein BdorOBP56f-2 and impairs the perception of methyl eugenol in Bactrocera dorsalis (Hendel). Insect Biochem. Mol. Biol. 2021, 139, 103656. [Google Scholar] [CrossRef]
- Koutroumpa, F.; Monsempès, C.; Anton, S.; François, M.-C.; Montagné, N.; Jacquin-Joly, E. Pheromone Receptor Knock-Out Affects Pheromone Detection and Brain Structure in a Moth. Biomolecules 2022, 12, 341. [Google Scholar] [CrossRef]
- Martinazzo, J.; Ballen, C.S.; Steffens, J.; Steffens, C. Sensing of pheromones from Euschistus heros (F.) stink bugs by nanosensors. Sens. Actuators Rep. 2022, 4, 100071. [Google Scholar] [CrossRef]
- Trombik, J.; Ward, S.F.; Norrbom, A.L.; Liebhold, A.M. Global drivers of historical true fruit fly (Diptera: Tephritidae) invasions. J. Pest. Sci. 2022, 1–13. [Google Scholar] [CrossRef]
- Malacrida, A.R.; Gomulski, L.M.; Bonizzoni, M.; Bertin, S.; Gasperi, G.; Guglielmino, C.R. Globalization and fruitfly invasion and expansion: The medfly paradigm. Genetica 2007, 131, 1–9. [Google Scholar] [CrossRef]
- Vera, M.T.; Cáceres, C.; Wornoayporn, V.; Islam, A.; Robinson, A.S.; De La Vega, M.H.; Hendrichs, J.; Cayol, J.-P. Mating Incompatibility Among Populations of the South American Fruit Fly Anastrepha fraterculus (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2006, 99, 387–397. [Google Scholar] [CrossRef]
- Caceres, C.; Segura, D.F.; Vera, M.T.; Wornoayporn, V.; Cladera, J.L.; Teal, P.; Sapountzis, P.; Bourtzis, K.; Zacharopoulou, A.; Robinson, S.A. Incipient speciation revealed in Anastrepha fraterculus (Diptera: Tephritidae) by studies on mating compatibility, sex pheromones, hybridization and cytology. Biol. J. Linn. Soc. 2009, 97, 152–165. [Google Scholar] [CrossRef]
- Hernandez-Ortíz, V. Distribución y biogeografía del género Anastrepha en México. In Moscas de la Fruta en Latinoamérica, Diversidad, Biología y Manejo; Hernández-Ortiz, V., Ed.; S y G Editores: Mexico City, Mexico, 2007; pp. 53–76. ISBN 9687623284. [Google Scholar]
- Aluja, M.; Mangan, R.L. Fruit Fly (Diptera: Tephritidae) Host Status Determination: Critical Conceptual, Methodological, and Regulatory Considerations. Annu. Rev. Entomol. 2008, 53, 473–502. [Google Scholar] [CrossRef] [PubMed]
- Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria (SENASICA). Available online: https://www.gob.mx/senasica/documentos/ficha-tecnica (accessed on 14 June 2022).
- Gazit, Y.; Rossler, Y.; Wang, S.; Tang, J.; Lurie, S. Thermal Death Kinetics of Egg and Third Instar Mediterranean Fruit Fly (Diptera: Tephritidae). J. Econ. Entomol. 2004, 97, 1540–1546. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, P.; He, X.L.; Woodcock, C.; Pickett, J.A.; Field, L.M.; Birkett, M.A.; Kalinova, B.; Gomulski, L.M.; Scolari, F.; Gasperi, G.; et al. Identification of pheromone components and their binding affinity to the odorant binding protein CcapOBP83a-2 of the Mediterranean fruit fly, Ceratitis capitata. Insect Biochem. Mol. Biol. 2014, 48, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Campanini, E.B.; de Brito, R.A. Molecular evolution of Odorant-binding proteins gene family in two closely related Anastrepha fruit flies. BMC Evol. Biol. 2016, 16, 198. [Google Scholar] [CrossRef]
- Elfekih, S.; Chen, C.-Y.; Hsu, J.-C.; Belcaid, M.; Haymer, D. Identification and preliminary characterization of chemosensory perception-associated proteins in the melon fly Bactrocera cucurbitae using RNA-seq. Sci. Rep. 2016, 6, 19112. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Qi, Z.; Chen, J.; Wang, F.; Gui, L.; Zhang, G. Molecular characterization of chemosensory protein genes in Bactrocera minax (Diptera: Tephritidae). Entomol. Res. 2021, 51, 349–356. [Google Scholar] [CrossRef]
- Wu, Z.; Kang, C.; Qu, M.; Chen, J.; Chen, M.; Bin, S.; Lin, J. Candidates for chemosensory genes identified in the Chinese citrus fly, Bactrocera minax, through a transcriptomic analysis. BMC Genom. 2019, 20. [Google Scholar] [CrossRef]
- Cheng, J.-F.; Yu, T.; Chen, Z.-J.; Chen, S.; Chen, Y.-P.; Gao, L.; Zhang, W.-H.; Jiang, B.; Bai, X.; Walker, E.D.; et al. Comparative genomic and transcriptomic analyses of chemosensory genes in the citrus fruit fly Bactrocera (Tetradacus) minax. Sci. Rep. 2020, 10, 18068. [Google Scholar] [CrossRef]
- Ono, H.; Hee, A.K.W.; Jiang, H. Recent advancements in studies on chemosensory mechanisms underlying detection of semiochemicals in Dacini fruit flies of economic importance (Diptera: Tephritidae). Insects 2021, 12, 106. [Google Scholar] [CrossRef]
- Wachi, N.; Matsubayashi, K.W.; Maeto, K. Application of next-generation sequencing to the study of non-model insects. Entomol. Sci. 2017, 21, 3–11. [Google Scholar] [CrossRef]
- Ruiz-May, E.; Altúzar-Molina, A.; Elizalde-Contreras, J.M.; Arellano-de los Santos, J.; Monribot-Villanueva, J.; Guillén, L.; Vázquez-Rosas-Landa, M.; Ibarra-Laclette, E.; Ramírez-Vázquez, M.; Ortega, R.; et al. A first glimpse of the mexican fruit fly Anastrepha ludens (Diptera: Tephritidae) antenna morphology and proteome in response to a proteinaceous Attractant. Int. J. Mol. Sci. 2020, 21, 8086. [Google Scholar] [CrossRef] [PubMed]
- Perre, P.; Jorge, L.R.; Lewinsohn, T.M.; Zucchi, R.A. Morphometric differentiation of fruit fly pest species of the Anastrepha fraterculus group (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2014, 107, 490–495. [Google Scholar] [CrossRef]
- Li, X.-M.; Zhu, X.-Y.; He, P.; Xu, L.; Sun, L.; Chen, L.; Wang, Z.-Q.; Deng, D.-G.; Zhang, Y.-N. Molecular characterization and sex distribution of chemosensory receptor gene family based on transcriptome analysis of Scaeva pyrastri. PLoS ONE 2016, 11, e0155323. [Google Scholar] [CrossRef]
- Jia, X.; Zhang, X.; Liu, H.; Wang, R.; Zhang, T. Identification of chemosensory genes from the antennal transcriptome of Indian meal moth Plodia interpunctella. PLoS ONE 2018, 13, e0189889. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.-H.; Yang, R.-N.; Guo, M.-B.; Wang, G.-R.; Wu, K.-M.; Guo, Y.-Y.; Zhou, J.-J.; Zhang, Y.-J. Molecular identification and differential expression of sensory neuron membrane proteins in the antennae of the black cutworm moth Agrotis ipsilon. J. Insect Physiol. 2013, 59, 430–443. [Google Scholar] [CrossRef]
- Wang, B.; Liu, Y.; Wang, G.-R. Chemosensory genes in the antennal transcriptome of two syrphid species, Episyrphus balteatus and Eupeodes corollae (Diptera: Syrphidae). BMC Genom. 2017, 18, 586. [Google Scholar] [CrossRef]
- Torres-Huerta, B.; Segura-León, O.L.; Aragón-Magadan, M.A.; González-Hernández, H. Identification and motif analyses of candidate nonreceptor olfactory genes of Dendroctonus adjunctus Blandford (Coleoptera: Curculionidae) from the head transcriptome. Sci. Rep. 2020, 10, 20695. [Google Scholar] [CrossRef]
- Archunan, G. Odorant Binding Proteins: A key player in the sense of smell. Bioinformation 2018, 14, 36–37. [Google Scholar] [CrossRef]
- Chen, X.-F.; Xu, L.; Zhang, Y.-X.; Wei, D.; Wang, J.-J.; Jiang, H.-B. Genome-wide identification and expression profiling of odorant-binding proteins in the oriental fruit fly, Bactrocera dorsalis. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 31, 100605. [Google Scholar] [CrossRef]
- Liu, Z.; Liang, X.-F.; Xu, L.; Keesey, I.W.; Lei, Z.-R.; Smagghe, G.; Wang, J.-J. An antennae-specific odorant-binding protein is involved in Bactrocera dorsalis olfaction. Front. Ecol. Evol. 2020, 8, 63. [Google Scholar] [CrossRef]
- Liu, Y.; Cui, Z.; Si, P.; Liu, Y.; Zhou, Q.; Wang, G. Characterization of a specific odorant receptor for linalool in the Chinese citrus fly Bactrocera minax (Diptera: Tephritidae). Insect Biochem. Mol. Biol. 2020, 122, 103389. [Google Scholar] [CrossRef] [PubMed]
- Garczynski, S.F.; Martin, J.A.; Griste, M.; Willett, L.S.; Cooper, W.R.; Swisher, K.D.; Unruh, T.R. CRISPR/Cas9 Editing of the Codling Moth (Lepidoptera: Tortricidae) CpomOR1 Gene Affects Egg Production and Viability. J. Econ. Entomol. 2017, 110, 1847–1855. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, L.; Tian, X.-L.; Gui, L.-Y.; Wang, F.-L.; Zhang, G.-H. Functional characterization of two antenna-enriched odorant-binding proteins from Bactrocera minax (Diptera: Tephritidae). J. Econ. Entomol. 2021, 144, 2361–2369. [Google Scholar] [CrossRef]
- Yao, R.; Zhao, M.; Zhong, L.; Li, Y.; Li, D.; Deng, Z.; Ma, X. Characterization of the binding ability of the odorant binding protein BminOBP9 of Bactrocera minax to citrus volatiles. Pest Manag. Sci. 2021, 77, 1214–1225. [Google Scholar] [CrossRef]
- Hekmat-Scafe, D.S.; Scafe, C.R.; McKinney, A.J.; Tanouye, M.A. Genome-wide analysis of the odorant-binding protein gene family in Drosophila melanogaster. Genome Res. 2002, 12, 1357–1369. [Google Scholar] [CrossRef]
- Vieira, F.G.; Rozas, J. Comparative genomics of the odorant-binding and chemosensory protein gene families across the Arthropoda: Origin and evolutionary history of the chemosensory system. Genome Biol. Evol. 2011, 3, 476–490. [Google Scholar] [CrossRef]
- Nakamura, A.M.; Chahad-Ehlers, S.; Lima, A.L.A.; Taniguti, C.H.; Sobrinho, I.; Torres, F.R.; de Brito, R.A. Reference genes for accessing differential expression among developmental stages and analysis of differential expression of OBP genes in Anastrepha obliqua. Sci. Rep. 2016, 6, 17480. [Google Scholar] [CrossRef]
- Campanini, E.B.; Congrains, C.; Torres, F.R.; de Brito, R.A. Odorant-binding proteins expression patterns in recently diverged species of Anastrepha fruit flies. Sci. Rep. 2017, 7, 2194. [Google Scholar] [CrossRef]
- Cheng, J.-F.; Yu, T.; Chen, Z.-J.; Chen, Y.-P.; Gao, L.; Zhang, W.-H.; Jiang, B.; Bai, X.; Liu, J.; Lu, Y. Identification and expression analysis of chemosensory genes in the citrus fruit fly Bactrocera (Tetradacus) minax. PeerJ Prepr. 2018, 6, e27297v1. [Google Scholar] [CrossRef]
- Wu, Z.; Cui, Y.; Ma, J.; Qu, M.; Lin, J. Analyses of chemosensory genes provide insight into the evolution of behavioral differences to phytochemicals in Bactrocera species. Mol. Phylogenet. Evol. 2020, 151, 106858. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Zhou, J.-J.; Ban, L.P.; Calvello, M. Soluble proteins in insect chemical communication. Cell Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef] [PubMed]
- Vieira, F.G.; Sánchez-Gracia, A.; Rozas, J. Comparative genomic analysis of the odorant-binding protein family in 12 Drosophila genomes: Purifying selection and birth-and-death evolution. Genome Biol. 2007, 8, R235. [Google Scholar] [CrossRef]
- Rihani, K.; Ferveur, J.-F.; Briand, L. The 40-Year Mystery of Insect Odorant-Binding Proteins. Biomolecules 2021, 11, 509. [Google Scholar] [CrossRef]
- Missbach, C.; Vogel, H.; Hansson, B.S.; Groβe-Wilde, E. Identification of odorant binding proteins and chemosensory proteins in antennal transcriptomes of the jumping bristletail Lepismachilis y-signata and the firebrat Thermobia domestica: Evidence for an independent OBP–OR origin. Chem. Senses 2015, 40, 615–626. [Google Scholar] [CrossRef] [PubMed]
- He, X.; He, Z.-B.; Zhang, Y.-J.; Zhou, Y.; Xian, P.-J.; Qiao, L.; Chen, B. Genome-wide identification and characterization of odorant-binding protein (OBP) genes in the malaria vector Anopheles sinensis (Diptera: Culicidae). Insect Sci. 2016, 23, 366–376. [Google Scholar] [CrossRef]
- Sun, J.S.; Xiao, S.; Carlson, J.R. The diverse small proteins called odorant-binding proteins. Open Biol. 2018, 8, 180208. [Google Scholar] [CrossRef]
- Simpson, M.G. Phylogenetic Systematics. In Plant Systematics; Elsevier-Academic Press: San Diego, CA, USA, 2010; pp. 17–52. [Google Scholar] [CrossRef]
- Pelosi, P.; Calvello, M.; Ban, L. Diversity of odorant-binding proteins and chemosensory proteins in insects. Chem. Senses 2005, 30, i291–i292. [Google Scholar] [CrossRef]
- Forêt, S.; Maleszka, R. Function and evolution of a gene family encoding odorant binding-like proteins in a social insect, the honey bee (Apis mellifera). Genome Res. 2006, 16, 1404–1413. [Google Scholar] [CrossRef]
- Forêt, S.; Wanner, K.W.; Maleszka, R. Chemosensory proteins in the honey bee: Insights from the annotated genome, comparative analyses and expressional profiling. Insect Biochem. Mol. Biol. 2007, 37, 19–28. [Google Scholar] [CrossRef]
- Gong, D.-P.; Zhang, H.-J.; Zhao, P.; Lin, Y.; Xia, Q.-Y.; Xiang, Z.-H. Identification and expression pattern of the chemosensory protein gene family in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2007, 37, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.-P.; Zhang, H.-J.; Zhao, P.; Xia, Q.-Y.; Xiang, Z.-H. The odorant binding protein gene family from the genome of silkworm, Bombyx mori. BMC Genom. 2009, 10, 332. [Google Scholar] [CrossRef] [PubMed]
- Dippel, S.; Oberhofer, G.; Kahnt, J.; Gerischer, L.; Opitz, L.; Schachtner, J.; Stanke, M.; Schütz, S.; Wimmer, E.A.; Angeli, S. Tissue-specific transcriptomics, chromosomal localization, and phylogeny of chemosensory and odorant binding proteins from the red flour beetle Tribolium castaneum reveal subgroup specificities for olfaction or more general functions. BMC Genom. 2014, 15, 1141. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, H.; Wang, Z.; Bin, S.; He, H.; Lin, J. Discovery of chemosensory genes in the oriental fruit fly, Bactrocera dorsalis. PLoS ONE 2015, 10, e0129794. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Smagghe, G.; Lei, Z.; Wang, J.-J. Identification of male- and female-specific olfaction genes in antennae of the oriental fruit fly (Bactrocera dorsalis). PLoS ONE 2016, 11, e0147783. [Google Scholar] [CrossRef]
- Xu, P.; Wang, Y.; Akami, M.; Niu, C.-Y. Identification of olfactory genes and functional analysis of BminCSP and BminOBP21 in Bactrocera minax. PLoS ONE 2019, 14, e0222193. [Google Scholar] [CrossRef]
- Yi, X.; Zhao, H.; Dong, X.; Wang, P.; Hu, M.; Zhong, G. BdorCSP2 Is Important for Antifeed and Oviposition-Deterring Activities Induced by Rhodojaponin-III against Bactrocera dorsalis. PLoS ONE 2013, 8, e77295. [Google Scholar] [CrossRef]
- Vogt, R.G. Biochemical diversity of odor detection: OBPs, ODEs and SNMPs. In Insect Pheromone Biochemistry and Molecular Biology: The Biosynthesis and Detection of Pheromones and Plant Volatiles; Blomquist, G., Vogt, R.G., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2003; pp. 391–445. ISBN 9780121071516. [Google Scholar]
- Nichols, Z.; Vogt, R.G. The SNMP/CD36 gene family in Diptera, Hymenoptera and Coleoptera: Drosophila melanogaster, D. pseudoobscura, Anopheles gambiae, Aedes aegypti, Apis mellifera, and Tribolium castaneum. Insect Biochem. Mol. Biol. 2008, 38, 398–415. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, J.; Liu, Y.; Wang, G.; Dong, S. Expression of SNMP1 and SNMP2 genes in antennal sensilla of Spodoptera exigua (Hübner). Arch. Insect Biochem. Physiol. 2014, 85, 114–126. [Google Scholar] [CrossRef]
- Isono, K.; Morita, H. Molecular and cellular designs of insect taste receptor system. Front. Cell Neurosci. 2010, 4, 20. [Google Scholar] [CrossRef]
- Rytz, R.; Croset, V.; Benton, R. Ionotropic receptors (IRs): Chemosensory ionotropic glutamate receptors in Drosophila and beyond. Insect Biochem. Mol. Biol. 2013, 43, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Wicher, D. Tuning insect odorant receptors. Front. Cell Neurosci. 2018, 12, 94. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Zhou, X.; Gu, F.; Zhong, G.; Yi, X. Olfactory plasticity: Variation in the expression of chemosensory receptors in Bactrocera dorsalis in different physiological states. Front. Physiol. 2017, 8, 672. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Zhu, C.; Peng, T.; Zhang, H. Odorant receptor co-receptor Orco is upregulated by methyl eugenol in male Bactrocera dorsalis (Diptera: Tephritidae). J. Insect Physiol. 2012, 58, 1122–1127. [Google Scholar] [CrossRef]
- Lin, C.-C.; Prokop-Prigge, K.A.; Preti, G.; Potter, C.J. Food odors trigger Drosophila males to deposit a pheromone that guides aggregation and female oviposition decisions. Neuroscience 2015, 4, e08688. [Google Scholar] [CrossRef]
- Miyazaki, H.; Otake, J.; Mitsuno, H.; Ozaki, K.; Kanzaki, R.; Chieng, A.C.-T.; Hee, A.K.-W.; Nishida, R.; Ono, H. Functional characterization of olfactory receptors in the oriental fruit fly Bactrocera dorsalis that respond to plant volatiles. Insect Biochem. Mol. Biol. 2018, 101, 32–46. [Google Scholar] [CrossRef]
- Hill, C.A.; Fox, A.N.; Pitts, R.J.; Kent, L.B.; Tan, P.L.; Chrystal, M.A.; Cravchik, A.; Collins, F.H.; Robertson, H.M.; Zwiebel, L.J. G protein-coupled receptors in Anopheles gambiae. Science 2002, 298, 176–178. [Google Scholar] [CrossRef]
- Robertson, H.M.; Warr, C.G.; Carlson, J.R. Molecular evolution of the insect chemoreceptor gene superfamily in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2003, 100, 14537–14542. [Google Scholar] [CrossRef]
- Robertson, H.M.; Wanner, K.W. The chemoreceptor superfamily in the honey bee, Apis mellifera: Expansion of the odorant, but not gustatory, receptor family. Genome Res. 2006, 16, 1395–1403. [Google Scholar] [CrossRef]
- Sánchez-Gracia, A.; Vieira, F.G.; Rozas, J. Molecular evolution of the major chemosensory gene families in insects. Heredity 2009, 103, 208–216. [Google Scholar] [CrossRef]
- Gouin, A.; Bretaudeau, A.; Nam, K.; Gimenez, S.; Aury, J.-M.; Duvic, B.; Hilliou, F.; Durand, N.; Montagné, N.; Darboux, I.; et al. Two genomes of highly polyphagous lepidopteran pests (Spodoptera frugiperda, Noctuidae) with different host-plant ranges. Sci. Rep. 2017, 7, 11816. [Google Scholar] [CrossRef] [PubMed]
- Pearce, S.L.; Clarke, D.F.; East, P.D.; Elfekih, S.; Gordon, K.H.J.; Jermiin, L.S.; McGaughran, A.; Oakeshott, J.G.; Papanikolaou, A.; Perera, O.P.; et al. Genomic innovations, transcriptional plasticity and gene loss underlying the evolution and divergence of two highly polyphagous and invasive Helicoverpa pest species. BMC Biol. 2017, 15, 63. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.Y.; Dahanukar, A.; Weiss, L.A.; Carlson, J.R. The molecular basis of CO2 reception in Drosophila. Proc. Natl. Acad. Sci. USA 2007, 104, 3574–3578. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.; Lee, Y.; Jeong, Y.T.; Kim, Y.; Lee, M.G.; Montell, C.; Moon, S.J. The full repertoire of Drosophila gustatory receptors for detecting an aversive compound. Nat. Commun. 2015, 6, 8867. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.Y.; Jeong, Y.T.; Lim, J.Y.; Kim, H.; Oh, S.M.; Hwang, S.W.; Oh, S.M.; Hwang, S.W.; Kwon, J.Y.; Moon, S.J. Heterogeneity in the Drosophila gustatory receptor complexes that detect aversive compounds. Nat. Commun. 2017, 8, 1484. [Google Scholar] [CrossRef]
- Aryal, B.; Lee, Y. Histamine gustatory aversion in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2021, 134, 103586. [Google Scholar] [CrossRef]
- Benton, R.; Vannice, K.S.; Gomez-Diaz, C.; Vosshall, L.B. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell 2009, 136, 149–162. [Google Scholar] [CrossRef]
- Croset, V.; Rytz, R.; Cummins, S.F.; Budd, A.; Brawand, D.; Kaessmann, H.; Gibson, T.J.; Benton, R. Ancient protostome origin of chemosensory ionotropic glutamate receptors and the evolution of insect taste and olfaction. PLoS Genet. 2010, 6, e1001064. [Google Scholar] [CrossRef]
- Ai, M.; Min, S.; Grosjean, Y.; Leblanc, C.; Bell, R.; Benton, R.; Suh, G.S.B. Acid sensing by the Drosophila olfactory system. Nature 2010, 468, 691–695. [Google Scholar] [CrossRef]
- Gereau, R.W.; Swanson, G.T. The Glutamate Receptors; Humana Press: Totawa, NJ, USA, 2008; ISBN 9781588297921. [Google Scholar]
- Abuin, L.; Bargeton, B.; Ulbrich, M.H.; Isacoff, E.Y.; Kellenberger, S.; Benton, R. Functional architecture of olfactory ionotropic glutamate receptors. Neuron 2011, 69, 44–60. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Troughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 14 June 2022).
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Bushmanova, E.; Antipov, D.; Lapidus, A.; Suvorov, V.; Prjibelski, A.D. rnaQUAST: A quality assessment tool forde novotranscriptome assemblies. Bioinformatics 2016, 32, 2210–2212. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef]
- Almagro, A.J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef]
- Kazutaka, K.; Misakwa, K.; Kei-ichi, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A new and scalable tool for the selection of DNA and protein evolutionary models. Mol. Biol. Evol. 2020, 37, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, 242–245. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily | Number | Length (aa) | Protein Core Region | Similarity (%) |
|---|---|---|---|---|
| Classic | 17 | 110–306 | C1-X36–171-C2-X3-C3-X32–50-C4-X8–21-C5-X8-C6 | 26.41 |
| Minus-C | 8 | 102–173 | C1-X16–30-C2-X38–39-C3-X18–19-C4 and C1-X28-C2-X3-C3-X39–40-C4-X10-C5-X8-C6 | 39.26 |
| Plus-C | 4 | 126–232 | C1a-X10–13-C1b-C1c-X13–19-C1-X9–48-C2-X3-C3-X43–44-C4-X19–33-C5-X9-C6-X8-C7-X10–11-C8-X9-C9 | 28.87 |
| Dimer | 2 | 271–282 | C1-X28–34-C2-X3-C3-X31-C4-X10–14-C5-X8-C6-X17–28-C1′-X24–25-C2′-X3-C3′-X35-C4′-X17–21-C5′-X8-C6′ | 35.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Segura-León, O.L.; Torres-Huerta, B.; Estrada-Pérez, A.R.; Cibrián-Tovar, J.; Hernandez-Hernandez, F.d.l.C.; Cruz-Jaramillo, J.L.; Meza-Hernández, J.S.; Sánchez-Galicia, F. Identification of Candidate Chemosensory Gene Families by Head Transcriptomes Analysis in the Mexican Fruit Fly, Anastrepha ludens Loew (Diptera: Tephritidae). Int. J. Mol. Sci. 2022, 23, 10531. https://doi.org/10.3390/ijms231810531
Segura-León OL, Torres-Huerta B, Estrada-Pérez AR, Cibrián-Tovar J, Hernandez-Hernandez FdlC, Cruz-Jaramillo JL, Meza-Hernández JS, Sánchez-Galicia F. Identification of Candidate Chemosensory Gene Families by Head Transcriptomes Analysis in the Mexican Fruit Fly, Anastrepha ludens Loew (Diptera: Tephritidae). International Journal of Molecular Sciences. 2022; 23(18):10531. https://doi.org/10.3390/ijms231810531
Chicago/Turabian StyleSegura-León, Obdulia L., Brenda Torres-Huerta, Alan Rubén Estrada-Pérez, Juan Cibrián-Tovar, Fidel de la Cruz Hernandez-Hernandez, José Luis Cruz-Jaramillo, José Salvador Meza-Hernández, and Fabian Sánchez-Galicia. 2022. "Identification of Candidate Chemosensory Gene Families by Head Transcriptomes Analysis in the Mexican Fruit Fly, Anastrepha ludens Loew (Diptera: Tephritidae)" International Journal of Molecular Sciences 23, no. 18: 10531. https://doi.org/10.3390/ijms231810531
APA StyleSegura-León, O. L., Torres-Huerta, B., Estrada-Pérez, A. R., Cibrián-Tovar, J., Hernandez-Hernandez, F. d. l. C., Cruz-Jaramillo, J. L., Meza-Hernández, J. S., & Sánchez-Galicia, F. (2022). Identification of Candidate Chemosensory Gene Families by Head Transcriptomes Analysis in the Mexican Fruit Fly, Anastrepha ludens Loew (Diptera: Tephritidae). International Journal of Molecular Sciences, 23(18), 10531. https://doi.org/10.3390/ijms231810531