
Golgi Phosphoprotein 3 Regulates the Physical Association of Glycolipid Glycosyltransferases †

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
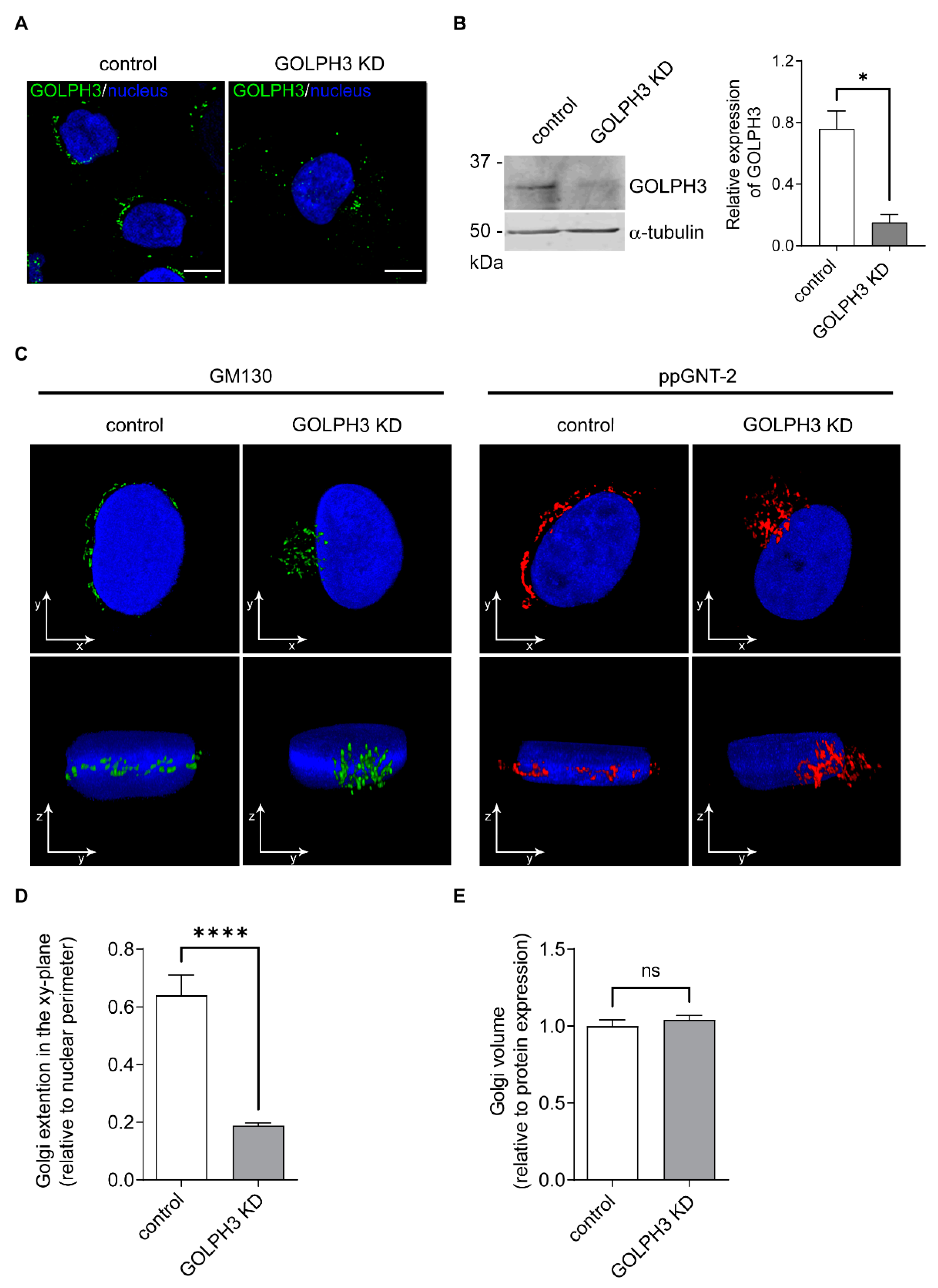
2.1. Knockdown of GOLPH3 Is Associated with Changes in the Distribution of the Golgi Complex
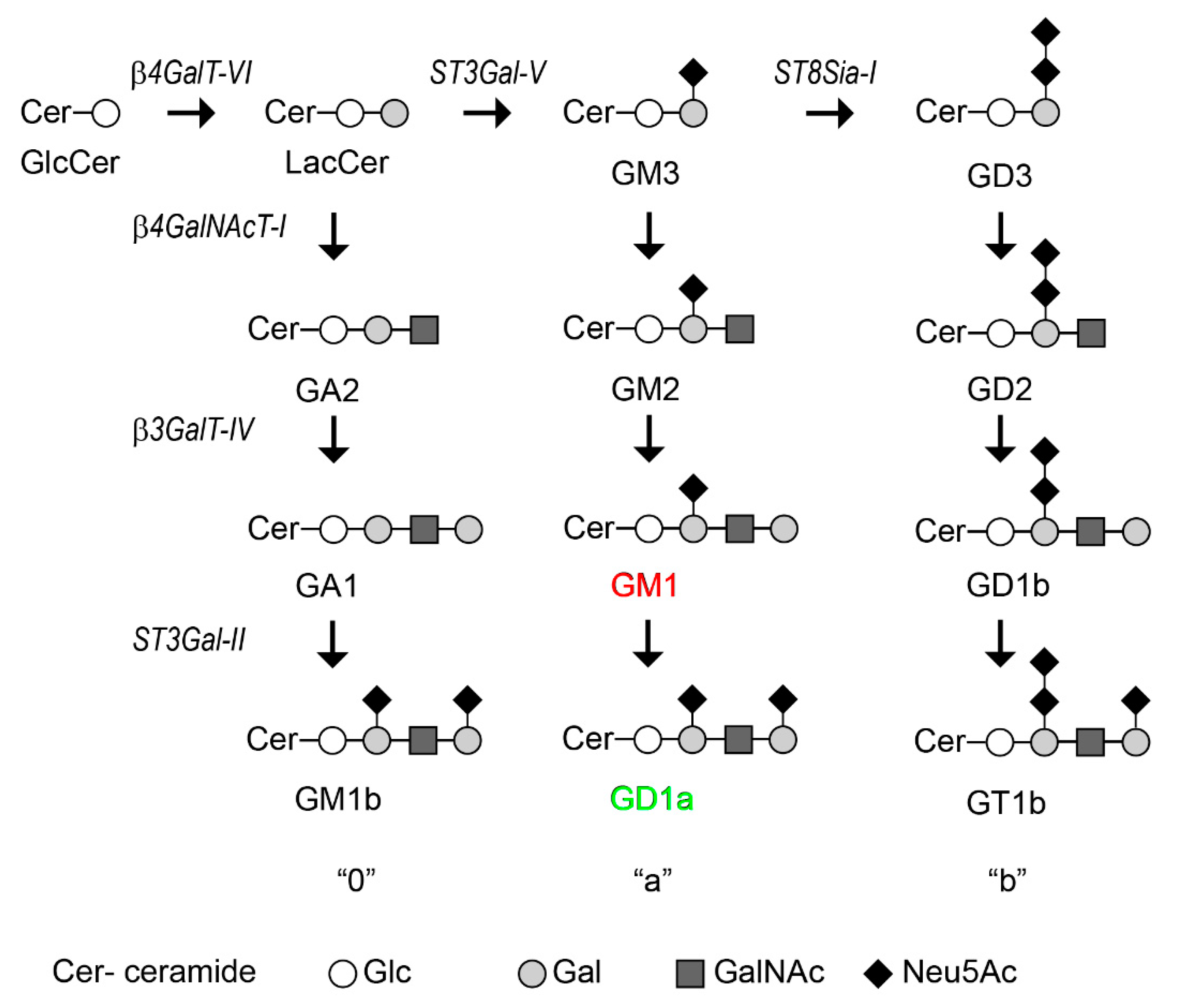
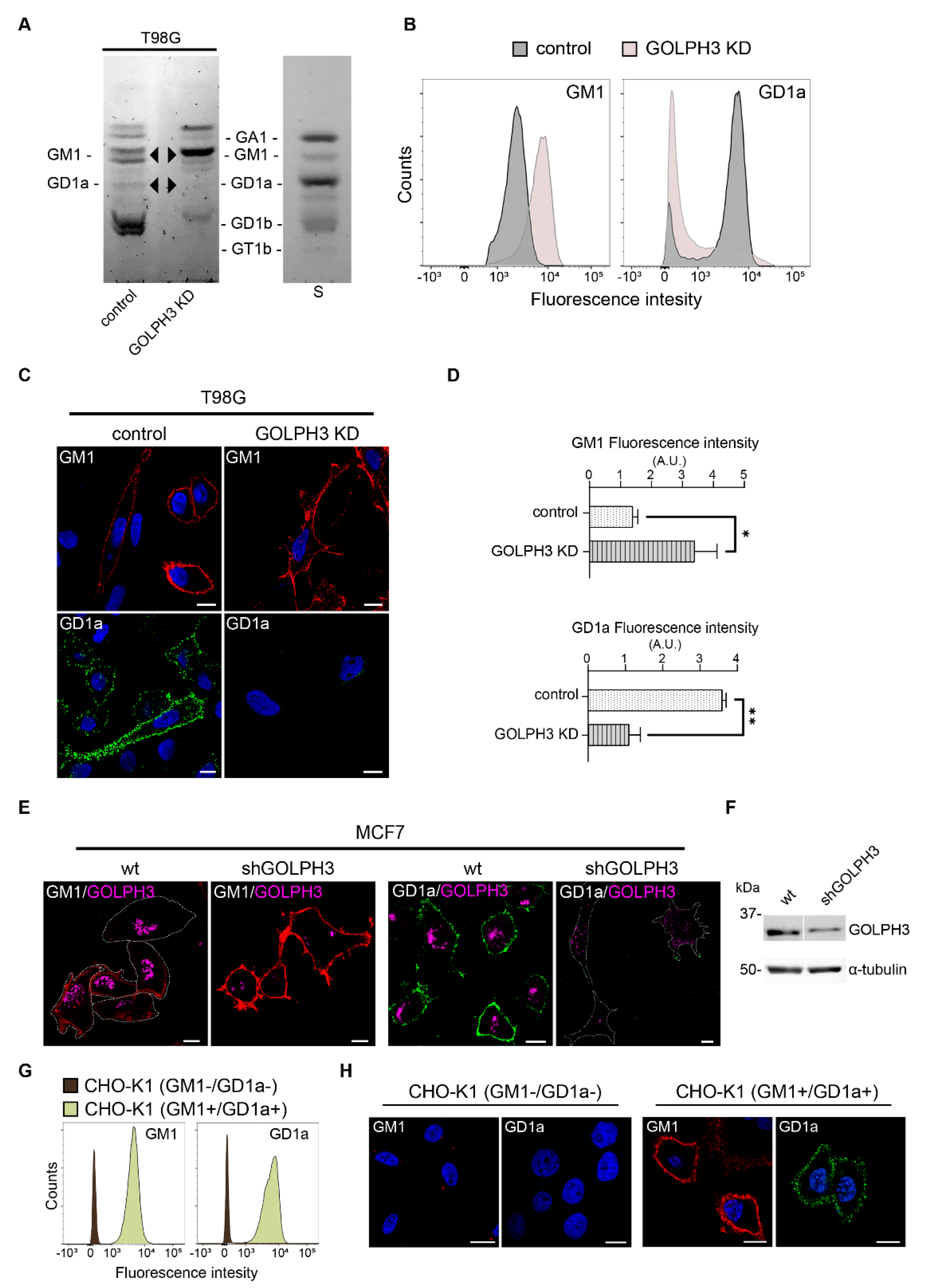
2.2. GOLPH3 Knockdown Is Associated with Changes in Glycolipid Expression
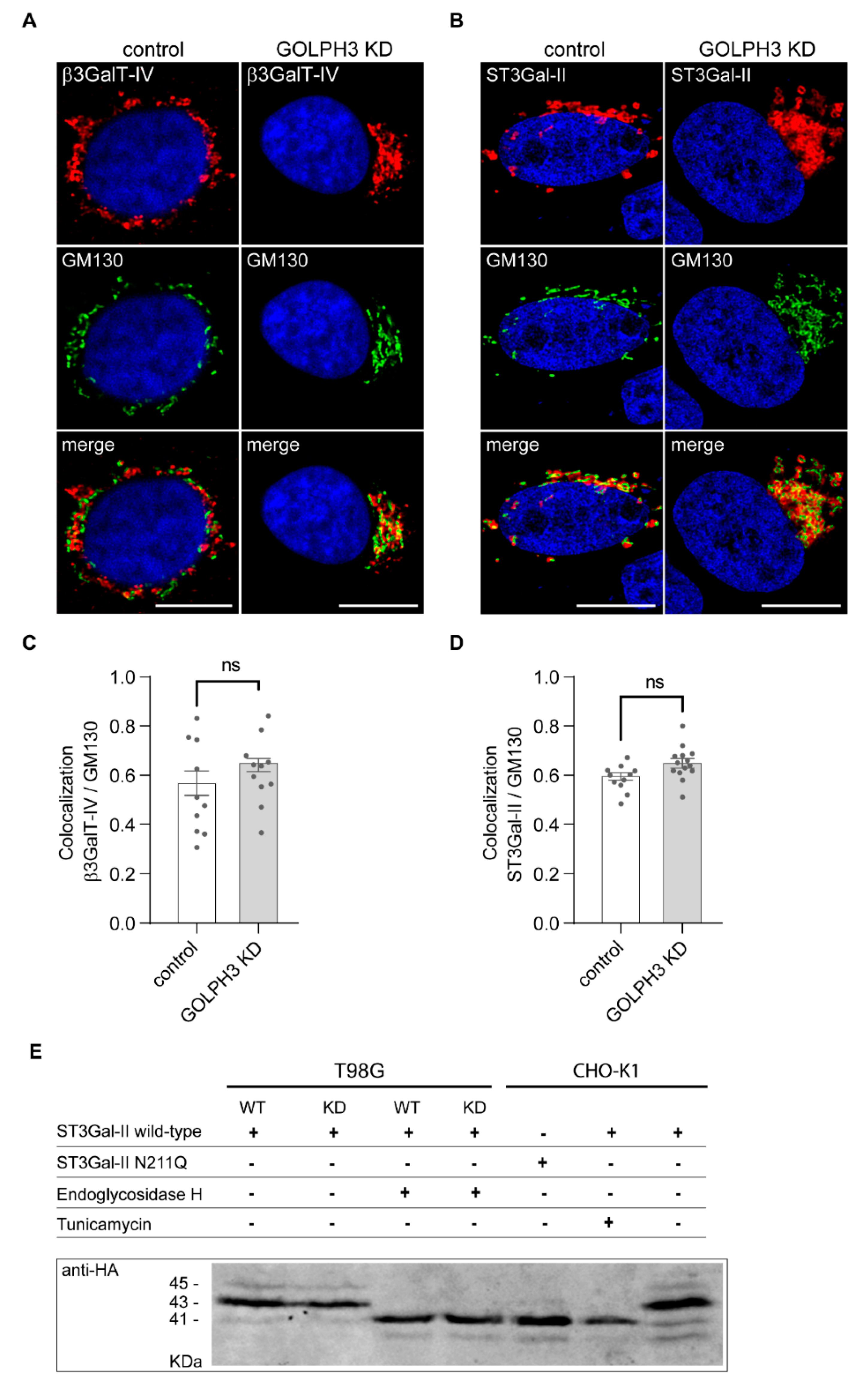
2.3. GOLPH3 Expression Does Not Affect the Subcellular Localization or N-Glycosylation of ST3Gal-II Sialyltransferase
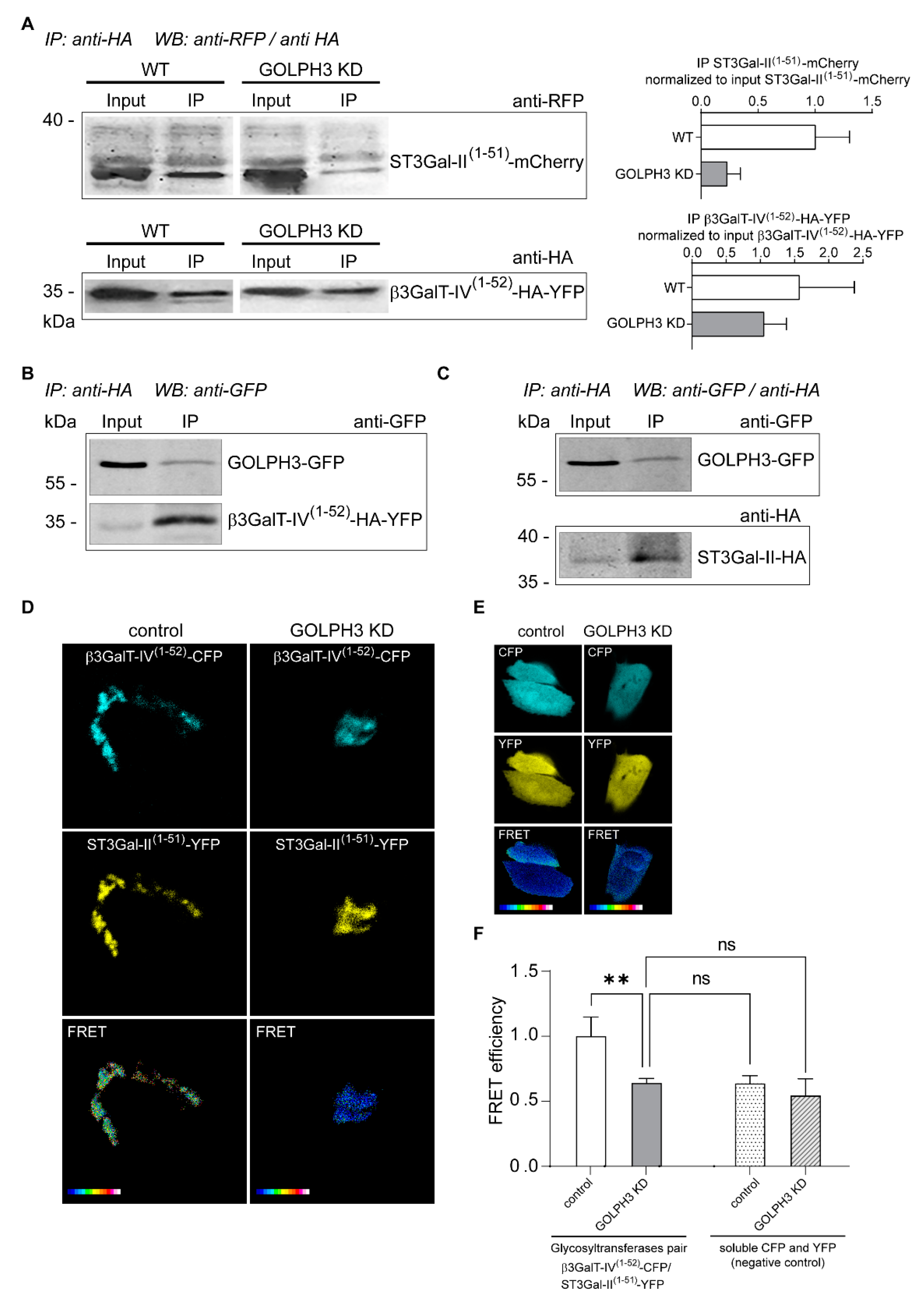
2.4. Role of GOLPH3 in the Physical Association of ST3Gal-II and β3GalT-IV Ganglioside Glycosyltransferases
3. Discussion
4. Materials and Methods
4.1. Cell Lines, Transfection and Electroporation
4.2. Cloning
4.3. Immunoprecipitation and Western Blot
4.4. Confocal Immunofluorescence Microscopy
4.5. FRET Analysis
4.6. Flow Cytometry
4.7. Plasmids and Antibodies
4.8. Thin Layer Chromatography (TLC)
4.9. Statistical Analyses
4.10. Image Processing
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Daniotti, J.L.; Vilcaes, A.A.; Rodriguez-Walker, M.; Ruggiero, F.M. Chapter 4: Metabolism of Glycolipids at the Cell Surface. In Sphingolipids: Biology, Synthesis and Functions; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2015; pp. 67–90. ISBN 978-1-63483-708-8. [Google Scholar]
- Daniotti, J.L.; Vilcaes, A.A.; Demichelis, V.T.; Ruggiero, F.M.; Rodriguez-Walker, M. Glycosylation of Glycolipids in Cancer: Basis for Development of Novel Therapeutic Approaches. Front. Oncol. 2013, 3, 306. [Google Scholar] [CrossRef] [PubMed]
- Groux-Degroote, S.; Delannoy, P. Cancer-Associated Glycosphingolipids as Tumor Markers and Targets for Cancer Immunotherapy. Int. J. Mol. Sci. 2021, 22, 6145. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Ohmi, Y.; Ohkawa, Y.; Bhuiyan, R.H.; Zhang, P.; Tajima, O.; Hashimoto, N.; Hamamura, K.; Furukawa, K. New era of research on cancer-associated glycosphingolipids. Cancer Sci. 2019, 110, 1544–1551. [Google Scholar] [CrossRef]
- Ruggiero, F.M.; Rodríguez-Walker, M.; Daniotti, J.L. Exploiting the internalization feature of an antibody against the glycosphingolipid SSEA-4 to deliver immunotoxins in breast cancer cells. Immunol. Cell Biol. 2020, 98, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Maccioni, H.J.; Quiroga, R.; Spessott, W. Organization of the synthesis of glycolipid oligosaccharides in the Golgi complex. FEBS Lett. 2011, 585, 1691–1698. [Google Scholar] [CrossRef] [PubMed]
- Sandhoff, R.; Sandhoff, K. Emerging concepts of ganglioside metabolism. FEBS Lett. 2018, 592, 3835–3864. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, S.C.; Ruggiero, F.M.; Daniotti, J.L.; Taubas, J.V. Ganglioside glycosyltransferases are S-acylated at conserved cysteine residues involved in homodimerisation. Biochem. J. 2017, 474, 2803–2816. [Google Scholar] [CrossRef]
- Ferrari, M.L.; Gomez, G.A.; Maccioni, H.J.F. Spatial Organization and Stoichiometry of N-Terminal Domain-Mediated Glycosyltransferase Complexes in Golgi Membranes Determined by Fret Microscopy. Neurochem. Res. 2012, 37, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, F.M.; Vilcaes, A.A.; Iglesias-Bartolome, R.; Daniotti, J.L. Critical role of evolutionarily conserved glycosylation at Asn211 in the intracellular trafficking and activity of sialyltransferase ST3Gal-II. Biochem. J. 2015, 469, 83–95. [Google Scholar] [CrossRef]
- Quiroga, R.; Trenchi, A.; Montoro, A.G.; Taubas, J.V.; Maccioni, H.J.F. Short length transmembrane domains high-volume exoplasmic halves determine retention of Type II membrane proteins in the Golgi complex. J. Cell Sci. 2013, 126, 5344–5349. [Google Scholar] [CrossRef] [PubMed]
- Cosson, P.; Perrin, J.; Bonifacino, J.S. Anchors aweigh: Protein localization and transport mediated by transmembrane domains. Trends Cell Biol. 2013, 23, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Spessott, W.; Crespo, P.M.; Daniotti, J.L.; Maccioni, H.J. Glycosyltransferase complexes improve glycolipid synthesis. FEBS Lett. 2012, 586, 2346–2350. [Google Scholar] [CrossRef] [PubMed]
- Bieberich, E.; MacKinnon, S.; Silva, J.; Li, D.D.; Tencomnao, T.; Irwin, L.; Kapitonov, D.; Yu, R.K. Regulation of Ganglioside Biosynthesis by Enzyme Complex Formation of Glycosyltransferases. Biochemistry 2002, 41, 11479–11487. [Google Scholar] [CrossRef] [PubMed]
- Martina, J.; Daniotti, J.L.; Maccioni, H.J.F. GM1 synthase depends on N-glycosylation for enzyme activity and trafficking to the Golgi complex. Neurochem. Res. 2000, 25, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Beki, M.E.; Wang, L.; Li, J.; Huang, H.; Tang, D.; Zhang, X.; Wang, Y. Knockout of the Golgi stacking proteins GRASP55 and GRASP65 impairs Golgi structure and function. Mol. Biol. Cell 2017, 28, 2833–2842. [Google Scholar] [CrossRef]
- Hassinen, A.; Pujol, F.M.; Kokkonen, N.; Pieters, C.; Kihlström, M.; Korhonen, K.; Kellokumpu, S. Functional Organization of Golgi N- and O-Glycosylation Pathways Involves pH-dependent Complex Formation That Is Impaired in Cancer Cells*. J. Biol. Chem. 2011, 286, 38329–38340. [Google Scholar] [CrossRef]
- Giraudo, C.G.; Maccioni, H.J. Ganglioside Glycosyltransferases Organize in Distinct Multienzyme Complexes in CHO-K1 Cells. J. Biol. Chem. 2003, 278, 40262–40271. [Google Scholar] [CrossRef] [PubMed]
- Opazo, J.C.; Vandewege, M.W.; Gutierrez, J.; Zavala, K.; Vargas-Chacoff, L.; Morera, F.J.; Mardones, G.A. Independent duplications of the Golgi phosphoprotein 3 oncogene in birds. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sechi, S.; Frappaolo, A.; Karimpour-Ghahnavieh, A.; Piergentili, R.; Giansanti, M.G. Oncogenic Roles of GOLPH3 in the Physiopathology of Cancer. Int. J. Mol. Sci. 2020, 21, 933. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.L.; Kabbarah, O.; Liang, M.-C.; Ivanova, E.; Anagnostou, V.; Wu, J.; Dhakal, S.; Wu, M.; Chen, S.; Feinberg, T.; et al. GOLPH3 modulates mTOR signalling and rapamycin sensitivity in cancer. Nature 2009, 459, 1085–1090. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.L.; Chin, L. Signaling From the Golgi: Mechanisms and Models for Golgi Phosphoprotein 3–Mediated Oncogenesis. Clin. Cancer Res. 2010, 16, 2229–2234. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Lin, H.; Zhao, X.; Liu, G.; Wang, X.; Xu, R.; Chen, K.; Li, J.; Song, L. Overexpression of GOLPH3 Promotes Proliferation and Tumorigenicity in Breast Cancer via Suppression of the FOXO1 Transcription Factor. Clin. Cancer Res. 2012, 18, 4059–4069. [Google Scholar] [CrossRef]
- Jiang, Y.; Su, Y.; Zhao, Y.; Pan, C.; Chen, L. Golgi phosphoprotein3 overexpression is associated with poor survival in patients with solid tumors: A meta-analysis. Int. J. Clin. Exp. Pathol. 2015, 8, 10615–10624. [Google Scholar]
- Sechi, S.; Frappaolo, A.; Belloni, G.; Colotti, G.; Giansanti, M.G. The multiple cellular functions of the oncoprotein Golgi phosphoprotein 3. Oncotarget 2015, 6, 3493–3506. [Google Scholar] [CrossRef]
- Tang, S.; Pan, H.; Wei, W.; Yang, H.; Liu, J.; Yang, R. GOLPH3: A novel biomarker that correlates with poor survival and resistance to chemotherapy in breast cancer. Oncotarget 2017, 8, 105155–105169. [Google Scholar] [CrossRef]
- Dippold, H.C.; Ng, M.M.; Farber-Katz, S.E.; Lee, S.-K.; Kerr, M.L.; Peterman, M.C.; Sim, R.; Wiharto, P.A.; Galbraith, K.A.; Madhavarapu, S.; et al. GOLPH3 Bridges Phosphatidylinositol-4- Phosphate and Actomyosin to Stretch and Shape the Golgi to Promote Budding. Cell 2009, 139, 337–351. [Google Scholar] [CrossRef]
- Kuna, R.S.; Field, S.J. GOLPH3: A Golgi phosphatidylinositol(4)phosphate effector that directs vesicle trafficking and drives cancer. J. Lipid Res. 2019, 60, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Sechi, S.; Colotti, G.; Belloni, G.; Mattei, V.; Frappaolo, A.; Raffa, G.D.; Fuller, M.T.; Giansanti, M.G. GOLPH3 Is Essential for Contractile Ring Formation and Rab11 Localization to the Cleavage Site during Cytokinesis in Drosophila melanogaster. PLoS Genet. 2014, 10, e1004305. [Google Scholar] [CrossRef] [PubMed]
- Farber-Katz, S.E.; Dippold, H.C.; Buschman, M.D.; Peterman, M.C.; Xing, M.; Noakes, C.J.; Tat, J.; Ng, M.M.; Rahajeng, J.; Cowan, D.M.; et al. DNA Damage Triggers Golgi Dispersal via DNA-PK and GOLPH3. Cell 2014, 156, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.M.; Dippold, H.C.; Buschman, M.D.; Noakes, C.J.; Field, S.J. GOLPH3L antagonizes GOLPH3 to determine Golgi morphology. Mol. Biol. Cell 2013, 24, 796–808. [Google Scholar] [CrossRef]
- Liu, L.; Doray, B.; Kornfeld, S. Recycling of Golgi glycosyltransferases requires direct binding to coatomer. Proc. Natl. Acad. Sci. USA 2018, 115, 8984–8989. [Google Scholar] [CrossRef] [PubMed]
- Buschman, M.D.; Exing, M.; Field, S.J. The GOLPH3 pathway regulates Golgi shape and function and is activated by DNA damage. Front. Neurosci. 2015, 9, 362. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, R.; Russo, D.; Kurokawa, K.; Sahu, P.; Lombardi, B.; Supino, D.; Zhukovsky, M.A.; Vocat, A.; Pothukuchi, P.; Kunnathully, V.; et al. Golgi maturation-dependent glycoenzyme recycling controls glycosphingolipid biosynthesis and cell growth via GOLPH3. EMBO J. 2021, 40, e107238. [Google Scholar] [CrossRef] [PubMed]
- Iwasawa, T.; Zhang, P.; Ohkawa, Y.; Momota, H.; Wakabayashi, T.; Ohmi, Y.; Bhuiyan, R.H.; Furukawa, K.; Furukawa, K. Enhancement of malignant properties of human glioma cells by ganglioside GD3/GD2. Int. J. Oncol. 2018, 52, 1255–1266. [Google Scholar] [CrossRef]
- Serafim, R.B.; da Silva, P.; Cardoso, C.; Di Cristofaro, L.F.M.; Netto, R.P.; de Almeida, R.; Navegante, G.; Storti, C.B.; de Sousa, J.F.; de Souza, F.C.; et al. Expression Profiling of Glioblastoma Cell Lines Reveals Novel Extracellular Matrix-Receptor Genes Correlated With the Responsiveness of Glioma Patients to Ionizing Radiation. Front. Oncol. 2021, 11, 668090. [Google Scholar] [CrossRef] [PubMed]
- Arriagada, C.; Cavieres, V.; Luchsinger, C.; González, A.; Muñoz, V.; Cancino, J.; Burgos, P.; Mardones, G. GOLPH3 Regulates EGFR in T98G Glioblastoma Cells by Modulating Its Glycosylation and Ubiquitylation. Int. J. Mol. Sci. 2020, 21, 8880. [Google Scholar] [CrossRef] [PubMed]
- Arriagada, C.; Luchsinger, C.; González, A.E.; Schwenke, T.; Arriagada, G.; Folch, H.; Ehrenfeld, I.; Burgos, P.V.; Mardones, G.A. The knocking down of the oncoprotein Golgi phosphoprotein 3 in T98G cells of glioblastoma multiforme disrupts cell migration by affecting focal adhesion dynamics in a focal adhesion kinase-dependent manner. PLoS ONE 2019, 14, e0212321. [Google Scholar] [CrossRef]
- Tenorio, M.J.; Ross, B.H.; Luchsinger, C.; Rivera-Dictter, A.; Arriagada, C.; Acuña, D.; Aguilar, M.; Cavieres, V.; Burgos, P.V.; Ehrenfeld, P.; et al. Distinct Biochemical Pools of Golgi Phosphoprotein 3 in the Human Breast Cancer Cell Lines MCF7 and MDA-MB-231. PLoS ONE 2016, 11, e0154719. [Google Scholar] [CrossRef]
- Rahajeng, J.; Kuna, R.S.; Makowski, S.L.; Tran, T.T.; Buschman, M.D.; Li, S.; Cheng, N.; Ng, M.M.; Field, S.J. Efficient Golgi Forward Trafficking Requires GOLPH3-Driven, PI4P-Dependent Membrane Curvature. Dev. Cell 2019, 50, 573–585.e5. [Google Scholar] [CrossRef] [PubMed]
- Halberg, N.; Sengelaub, C.A.; Navrazhina, K.; Molina, H.; Uryu, K.; Tavazoie, S.F. PITPNC1 Recruits RAB1B to the Golgi Network to Drive Malignant Secretion. Cancer Cell 2016, 29, 339–353. [Google Scholar] [CrossRef]
- Giraudo, C.G.; Rosales Fritz, V.M.; Maccioni, H.J. GA2/GM2/GD2 synthase localizes to the trans-golgi network of CHO-K1 cells. Biochem. J. 1999, 342, 633–640. [Google Scholar] [CrossRef]
- Fritz, V.M.R.; Maxzud, M.K.; Maccioni, H.J.F. GT3 Synthesis in the Proximal Golgi Occurs in a Compartment Different from Those for GD3 and GM3 Synthesis. J. Neurochem. 2002, 67, 1393–1400. [Google Scholar] [CrossRef]
- Uliana, A.S.; Crespo, P.M.; Martina, J.A.; Daniotti, J.L.; Maccioni, H.J.F. Modulation of GalT1 and SialT1 Sub-Golgi Localization by SialT2 Expression Reveals an Organellar Level of Glycolipid Synthesis Control. J. Biol. Chem. 2006, 281, 32852–32860. [Google Scholar] [CrossRef]
- Ruggiero, F.M.; Vilcaes, A.A.; Yuki, N.; Daniotti, J.L. Membrane binding, endocytic trafficking and intracellular fate of high-affinity antibodies to gangliosides GD1a and GM1. Biochim. Et Biophys. Acta Biomembr. 2017, 1859, 80–93. [Google Scholar] [CrossRef]
- Sturgill, E.R.; Aoki, K.; Lopez, P.H.; Colacurcio, D.; Vajn, K.; Lorenzini, I.; Majić, S.; Yang, W.H.; Heffer, M.; Tiemeyer, M.; et al. Biosynthesis of the major brain gangliosides GD1a and GT1b. Glycobiology 2012, 22, 1289–1301. [Google Scholar] [CrossRef]
- Tu, L.; Tai, W.C.S.; Chen, L.; Banfield, D.K. Signal-Mediated Dynamic Retention of Glycosyltransferases in the Golgi. Science 2008, 321, 404–407. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, K.; Liu, J.; Li, S.; Setty, T.G.; Wood, C.S.; Burd, C.G.; Ferguson, K.M. Golgi Localization of Glycosyltransferases Requires a Vps74p Oligomer. Dev. Cell 2008, 14, 523–534. [Google Scholar] [CrossRef]
- Ali, M.F.; Chachadi, V.B.; Petrosyan, A.; Cheng, P.-W. Golgi Phosphoprotein 3 Determines Cell Binding Properties under Dynamic Flow by Controlling Golgi Localization of Core 2 N-Acetylglucosaminyltransferase 1. J. Biol. Chem. 2012, 287, 39564–39577. [Google Scholar] [CrossRef] [PubMed]
- Pereira, N.A.; Pu, H.X.; Goh, H.; Song, Z. Golgi Phosphoprotein 3 Mediates the Golgi Localization and Function of Protein O-Linked Mannose β-1,2-N-Acetlyglucosaminyltransferase 1. J. Biol. Chem. 2014, 289, 14762–14770. [Google Scholar] [CrossRef] [PubMed]
- Giraudo, C.G.; Daniotti, J.L.; Maccioni, H.J.F. Physical and functional association of glycolipid N -acetyl-galactosaminyl and galactosyl transferases in the Golgi apparatus. Proc. Natl. Acad. Sci. USA 2001, 98, 1625–1630. [Google Scholar] [CrossRef] [PubMed]
- Frappaolo, A.; Karimpour-Ghahnavieh, A.; Sechi, S.; Giansanti, M.G. The Close Relationship between the Golgi Trafficking Machinery and Protein Glycosylation. Cells 2020, 9, 2652. [Google Scholar] [CrossRef]
- Eckert, E.S.; Reckmann, I.; Hellwig, A.; Röhling, S.; El-Battari, A.; Wieland, F.T.; Popoff, V. Golgi Phosphoprotein 3 Triggers Signal-mediated Incorporation of Glycosyltransferases into Coatomer-coated (COPI) Vesicles. J. Biol. Chem. 2014, 289, 31319–31329. [Google Scholar] [CrossRef] [PubMed]
- Giraudo, C.G.; Maccioni, H.J.F. Endoplasmic Reticulum Export of Glycosyltransferases Depends on Interaction of a Cytoplasmic Dibasic Motif with Sar1. Mol. Biol. Cell 2003, 14, 3753–3766. [Google Scholar] [CrossRef]
- Isaji, T.; Im, S.; Gu, W.; Wang, Y.; Hang, Q.; Lu, J.; Fukuda, T.; Hashii, N.; Takakura, D.; Kawasaki, N.; et al. An Oncogenic Protein Golgi Phosphoprotein 3 Up-regulates Cell Migration via Sialylation. J. Biol. Chem. 2014, 289, 20694–20705. [Google Scholar] [CrossRef] [PubMed]
- Kellokumpu, S.; Hassinen, A.; Glumoff, T. Glycosyltransferase complexes in eukaryotes: Long-known, prevalent but still unrecognized. Cell. Mol. Life Sci. 2016, 73, 305–325. [Google Scholar] [CrossRef]
- Vilcaes, A.A.; Garbarino-Pico, E.; Demichelis, V.T.; Daniotti, J.L. Ganglioside Synthesis by Plasma Membrane-Associated Sialyltransferase in Macrophages. Int. J. Mol. Sci. 2020, 21, 1063. [Google Scholar] [CrossRef]
- Donizy, P.; Kaczorowski, M.; Biecek, P.; Halon, A.; Szkudlarek, T.; Matkowski, R. Golgi-Related Proteins GOLPH2 (GP73/GOLM1) and GOLPH3 (GOPP1/MIDAS) in Cutaneous Melanoma Patterns of Expression and Prognostic Significance. Int. J. Mol. Sci. 2016, 17, 1619. [Google Scholar] [CrossRef]
- Maccioni, H.J.F.; Quiroga, R.; Ferrari, M.L. Cellular and molecular biology of glycosphingolipid glycosylation. J. Neurochem. 2011, 117, 589–602. [Google Scholar] [CrossRef] [PubMed]
- Gordon, G.W.; Berry, G.; Liang, X.H.; Levine, B.; Herman, B. Quantitative Fluorescence Resonance Energy Transfer Measurements Using Fluorescence Microscopy. Biophys. J. 1998, 74, 2702–2713. [Google Scholar] [CrossRef]
- Schnaar, R.L.; Fromholt, S.E.; Gong, Y.; Vyas, A.A.; Laroy, W.; Wayman, D.M.; Heffer, M.; Ito, H.; Ishida, H.; Kiso, M.; et al. Immunoglobulin G-Class Mouse Monoclonal Antibodies to Major Brain Gangliosides. Anal. Biochem. 2002, 302, 276–284. [Google Scholar] [CrossRef]
- Müthing, J.; Peter-Katalinić, J.; Hanisch, F.-G.; Neumann, U. Structural studies of gangliosides from the YAC-1 mouse lymphoma cell line by immunological detection and fast atom bombardment mass spectrometry. Glycoconj. J. 1991, 8, 414–423. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruggiero, F.M.; Martínez-Koteski, N.; Cavieres, V.A.; Mardones, G.A.; Fidelio, G.D.; Vilcaes, A.A.; Daniotti, J.L. Golgi Phosphoprotein 3 Regulates the Physical Association of Glycolipid Glycosyltransferases. Int. J. Mol. Sci. 2022, 23, 10354. https://doi.org/10.3390/ijms231810354
Ruggiero FM, Martínez-Koteski N, Cavieres VA, Mardones GA, Fidelio GD, Vilcaes AA, Daniotti JL. Golgi Phosphoprotein 3 Regulates the Physical Association of Glycolipid Glycosyltransferases. International Journal of Molecular Sciences. 2022; 23(18):10354. https://doi.org/10.3390/ijms231810354
Chicago/Turabian StyleRuggiero, Fernando M., Natalia Martínez-Koteski, Viviana A. Cavieres, Gonzalo A. Mardones, Gerardo D. Fidelio, Aldo A. Vilcaes, and Jose L. Daniotti. 2022. "Golgi Phosphoprotein 3 Regulates the Physical Association of Glycolipid Glycosyltransferases" International Journal of Molecular Sciences 23, no. 18: 10354. https://doi.org/10.3390/ijms231810354
APA StyleRuggiero, F. M., Martínez-Koteski, N., Cavieres, V. A., Mardones, G. A., Fidelio, G. D., Vilcaes, A. A., & Daniotti, J. L. (2022). Golgi Phosphoprotein 3 Regulates the Physical Association of Glycolipid Glycosyltransferases. International Journal of Molecular Sciences, 23(18), 10354. https://doi.org/10.3390/ijms231810354