

Isoquinoline Alkaloids from Coptis chinensis Franch: Focus on Coptisine as a Potential Therapeutic Candidate against Gastric Cancer Cells

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
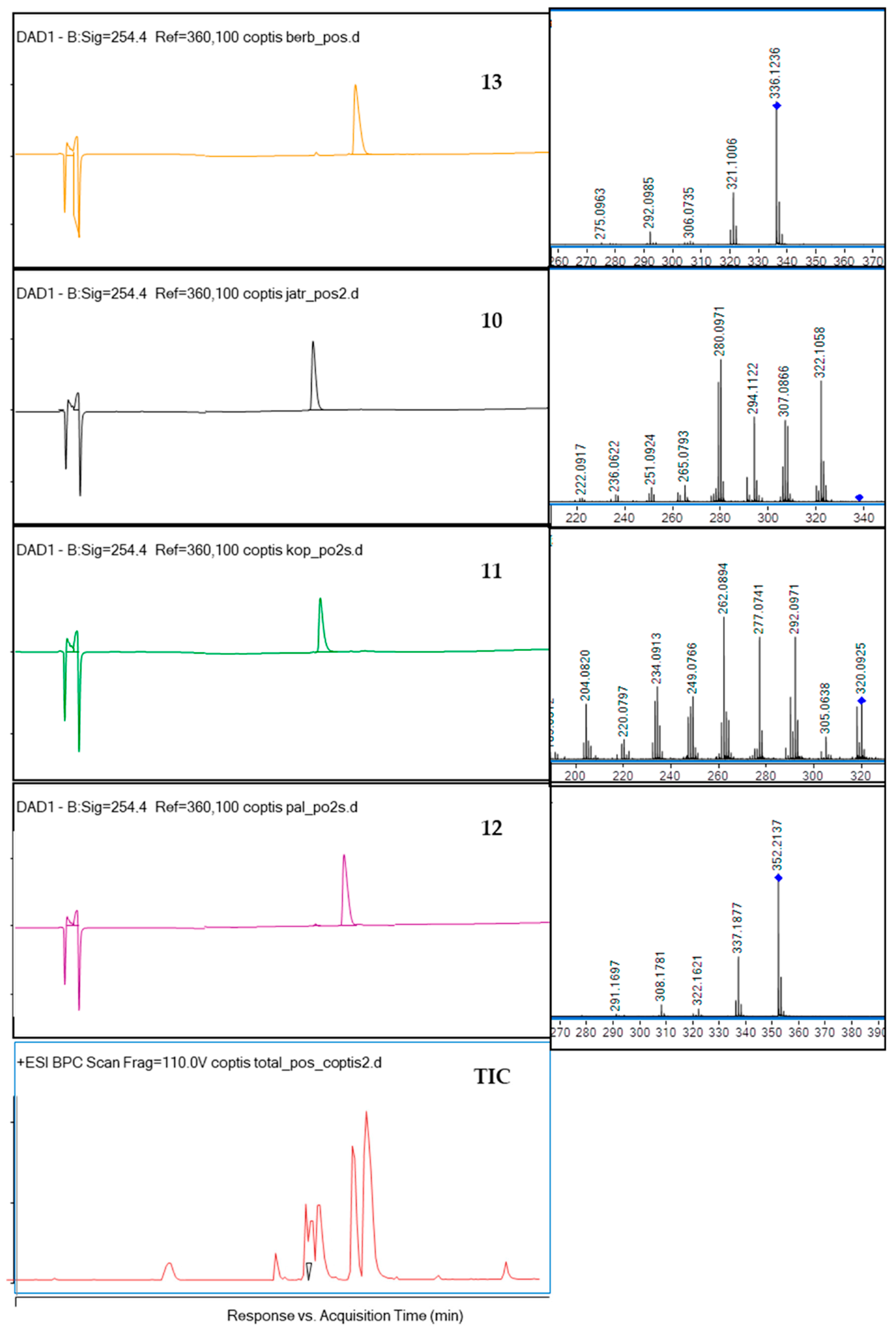
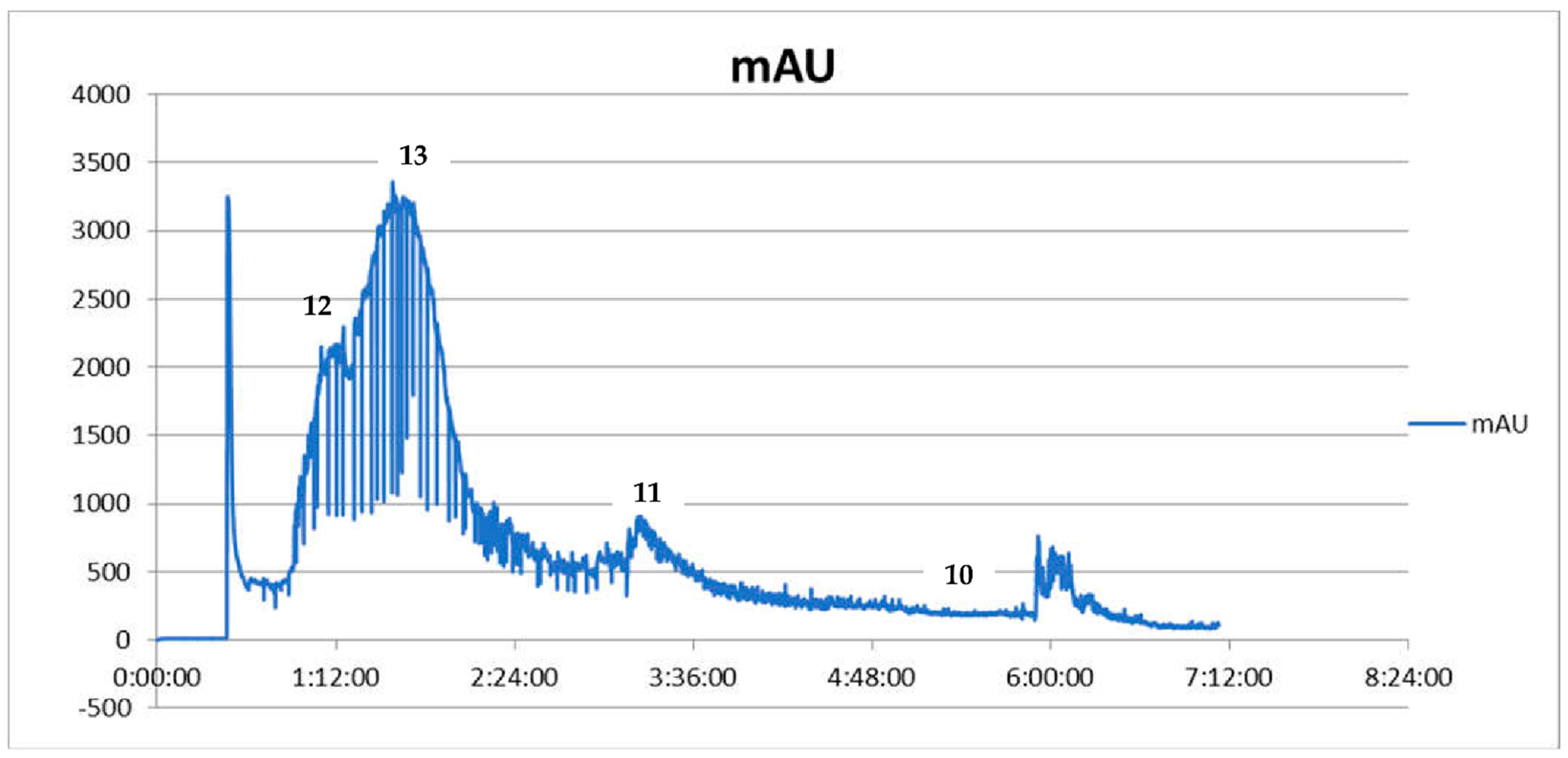
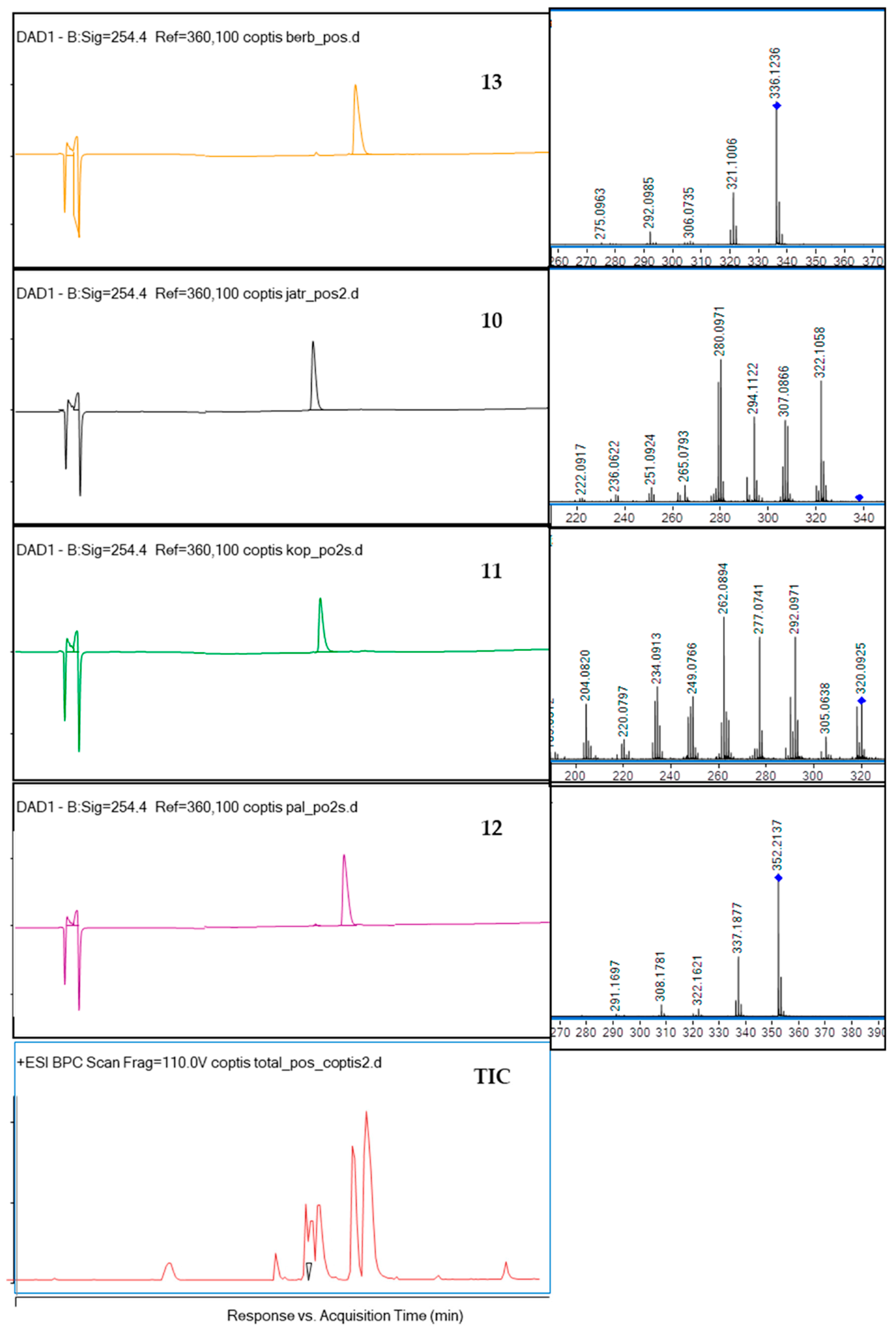
2.1. Compositional Studies and Isolation of Alkaloids from Coptis chinensis Root Methanolic Extract by CPC Chromatography
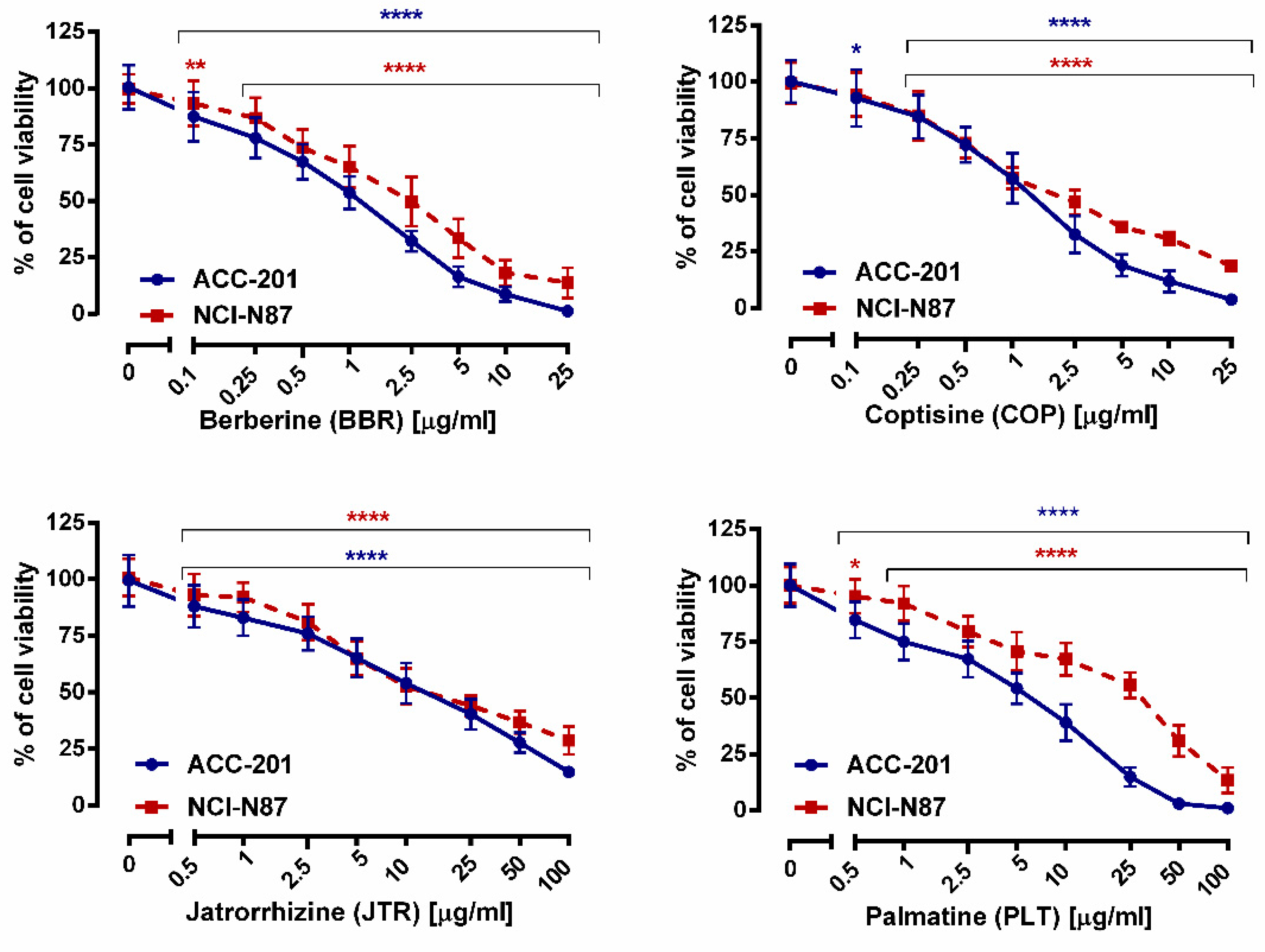
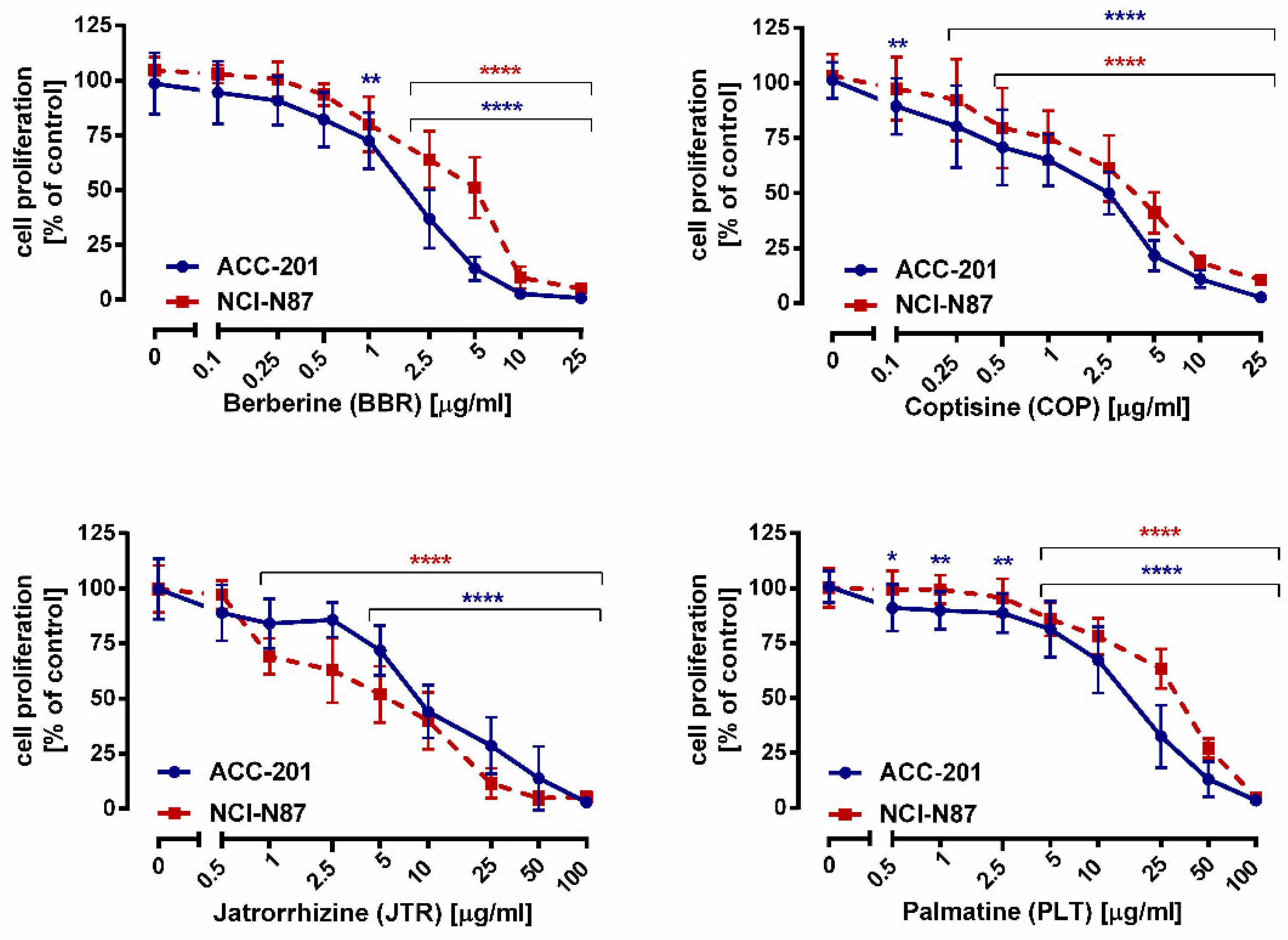
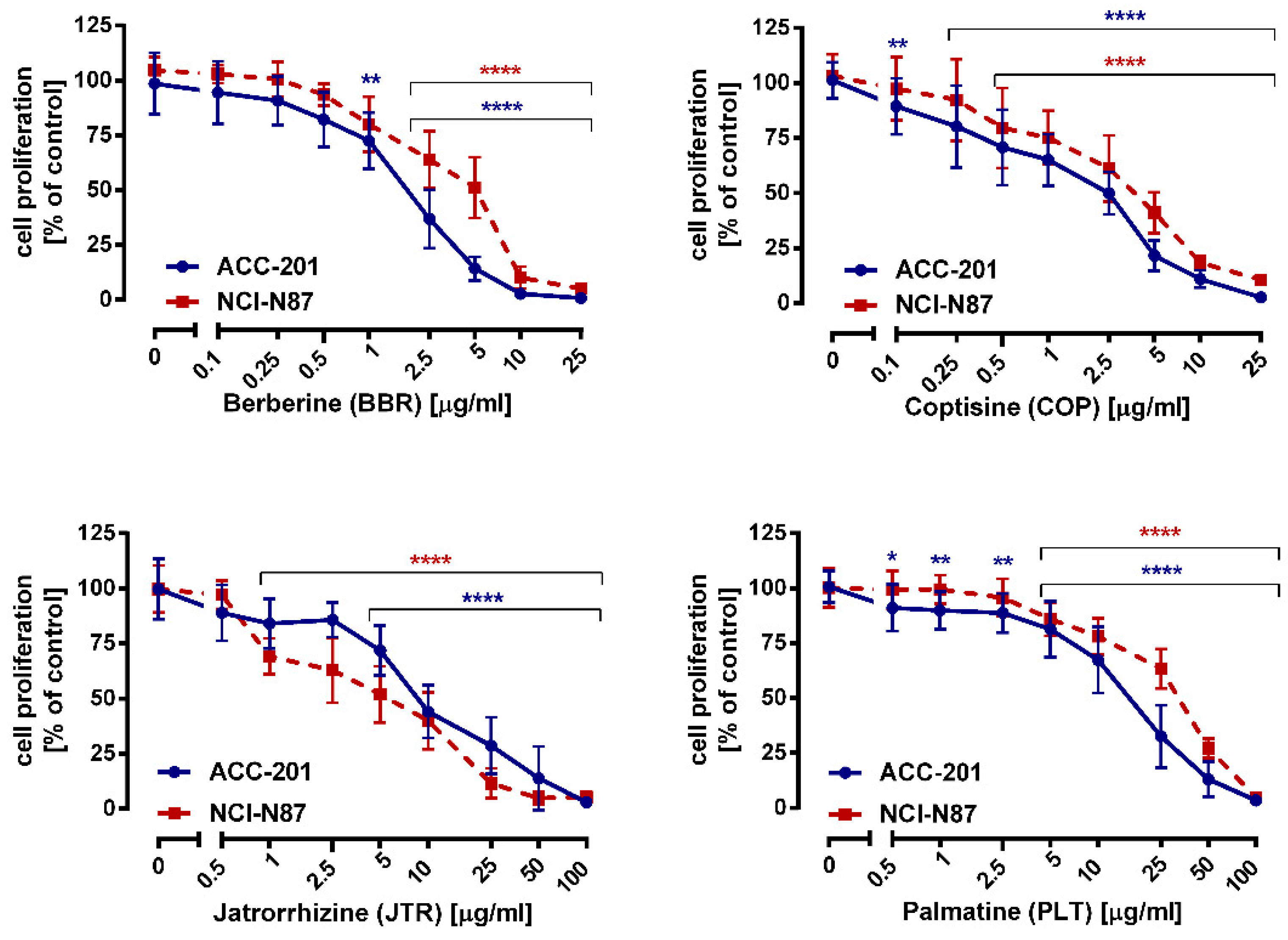
2.2. Impact of Isoquinoline Alkaloids Isolated from Coptis chinensis on the Viability and Proliferation of ACC-201 and NCI-N87 Gastric Cancer Cells
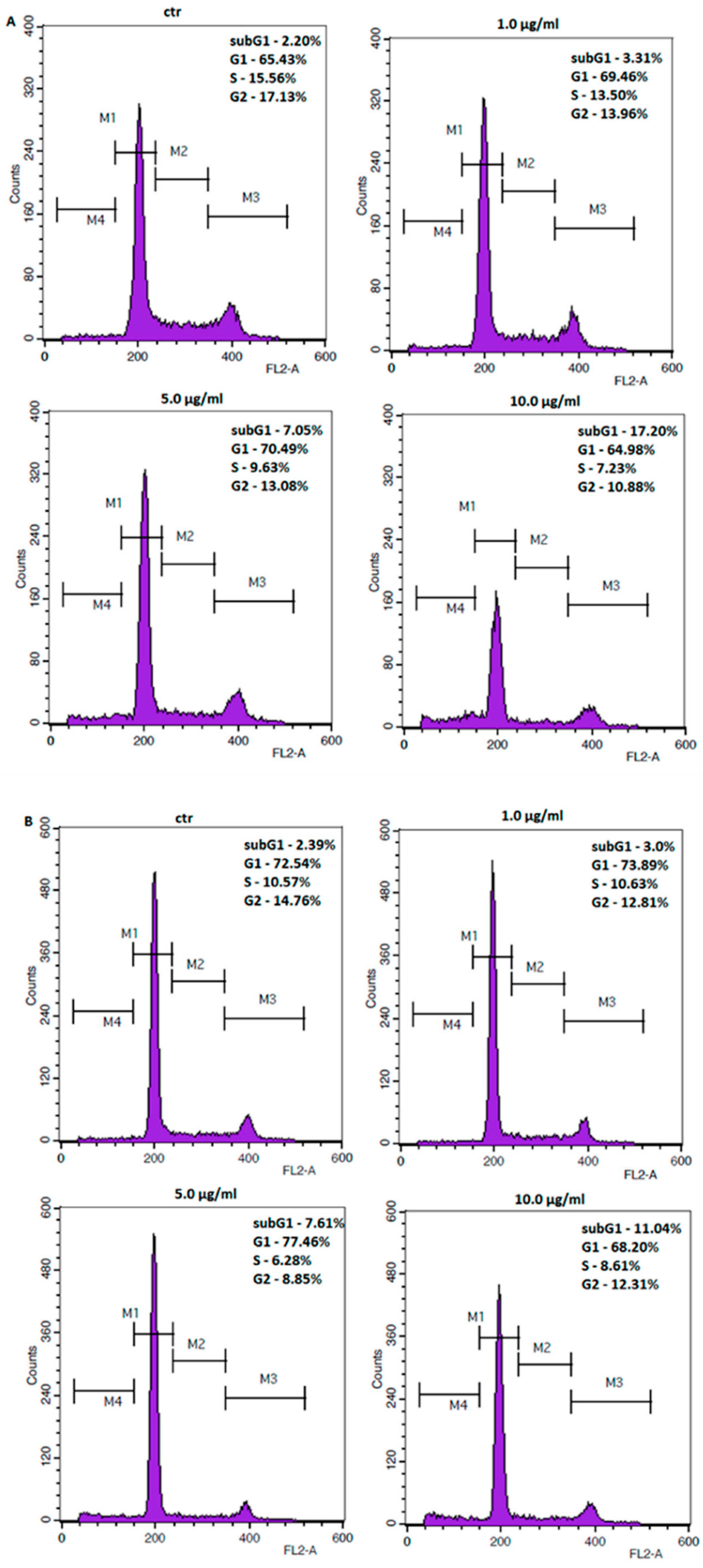
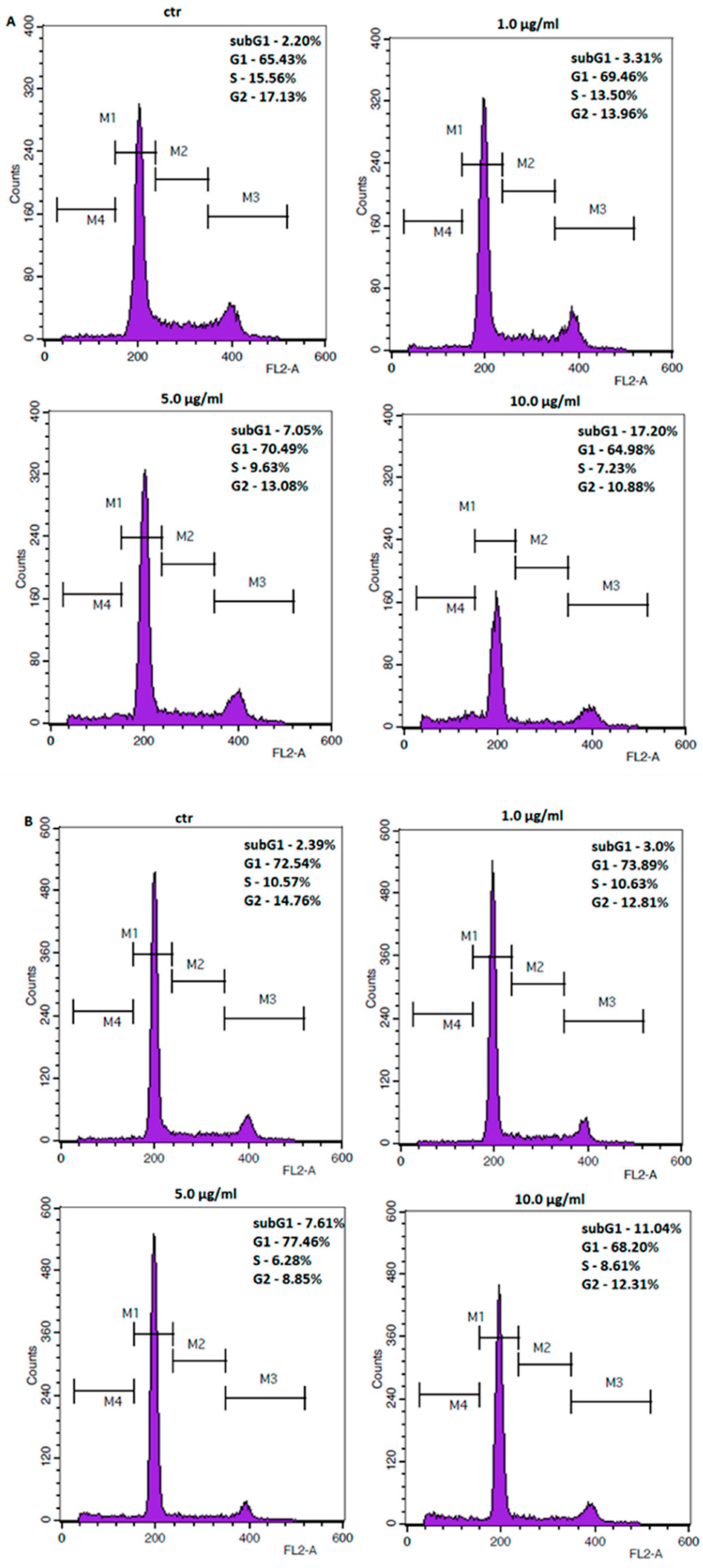
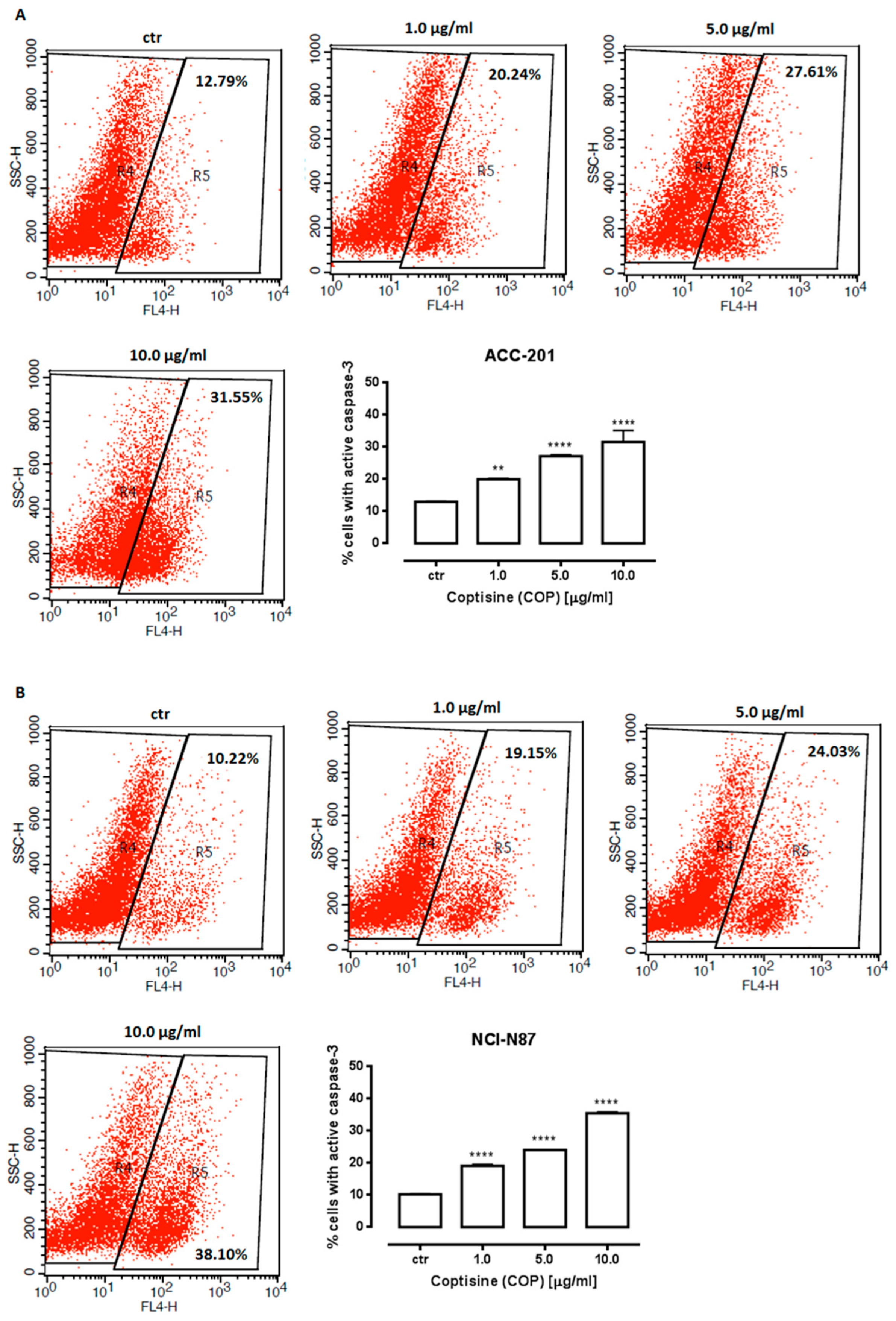
2.3. Coptisine Treatment Increases Population of Cells in Sub-G1 Phase and Induces Apoptosis in Gastric Cancer Cell Lines
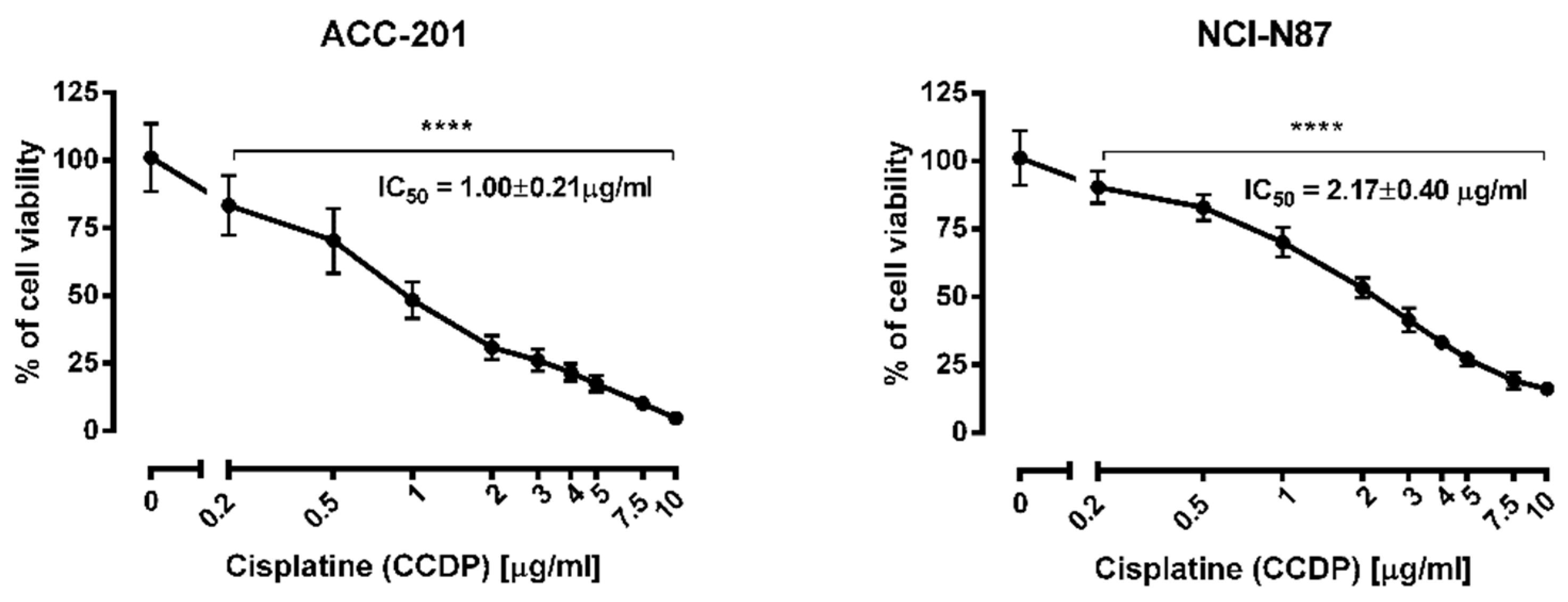
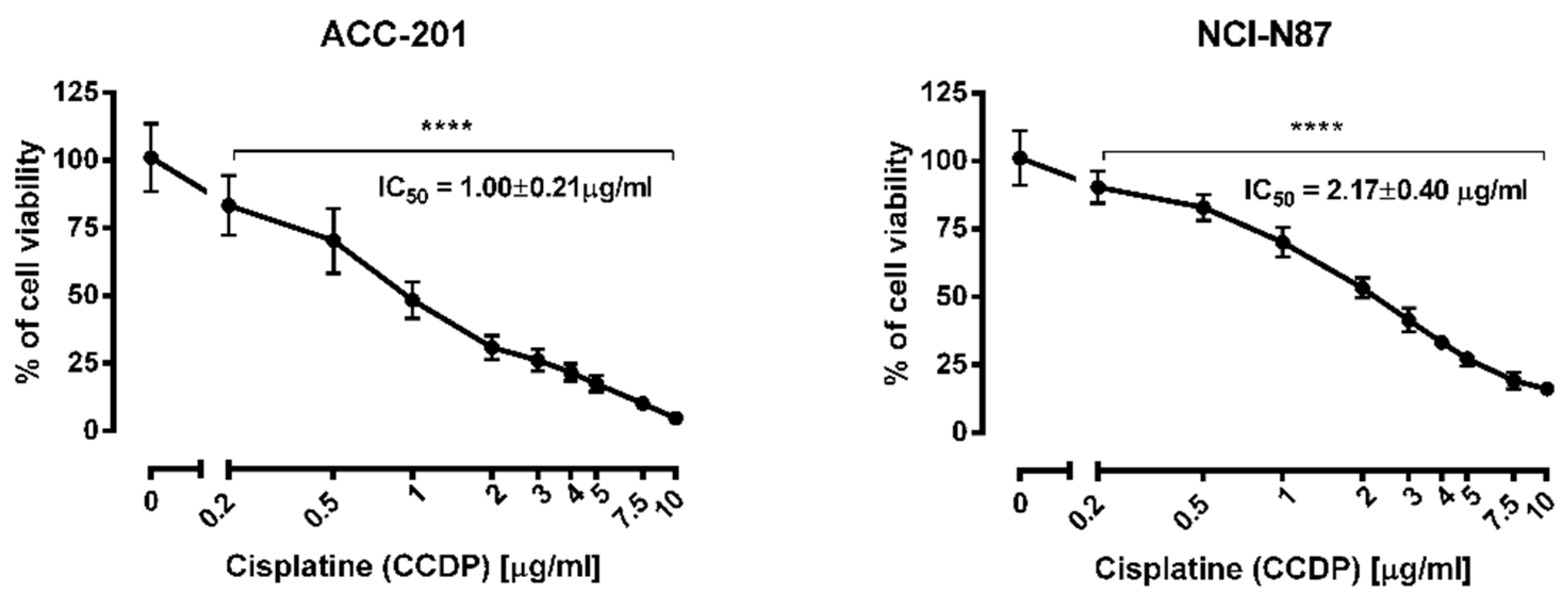
2.4. The Anti-Proliferative Effects of Coptisine Administered in Combinatin with Cisplatin
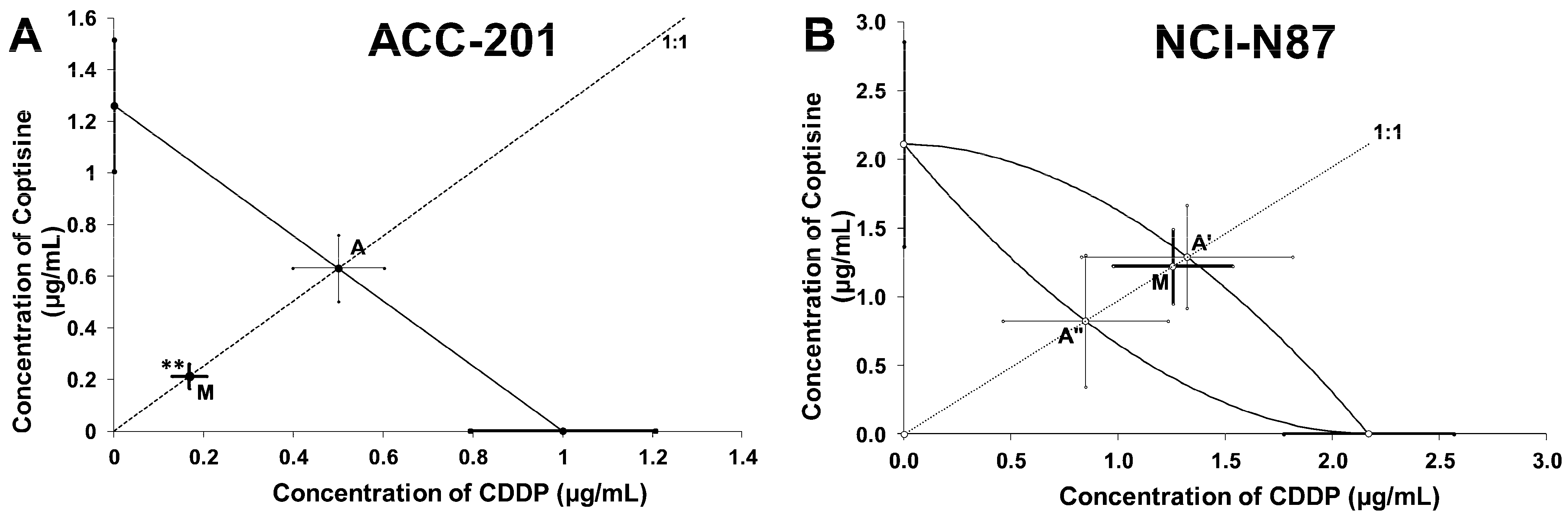
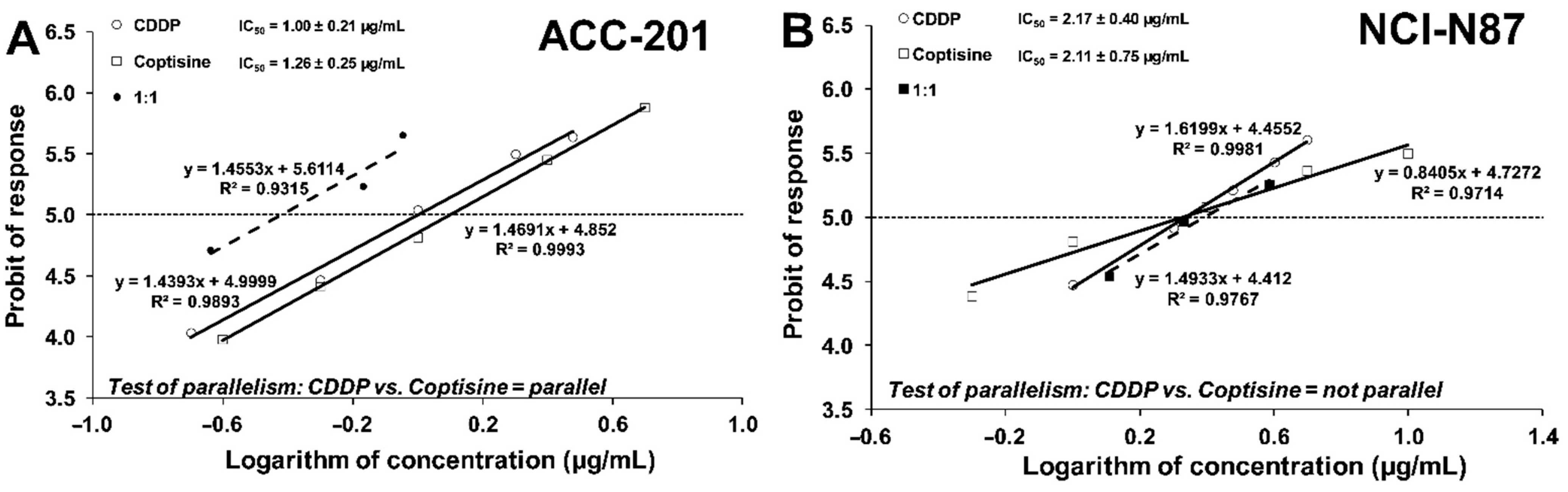
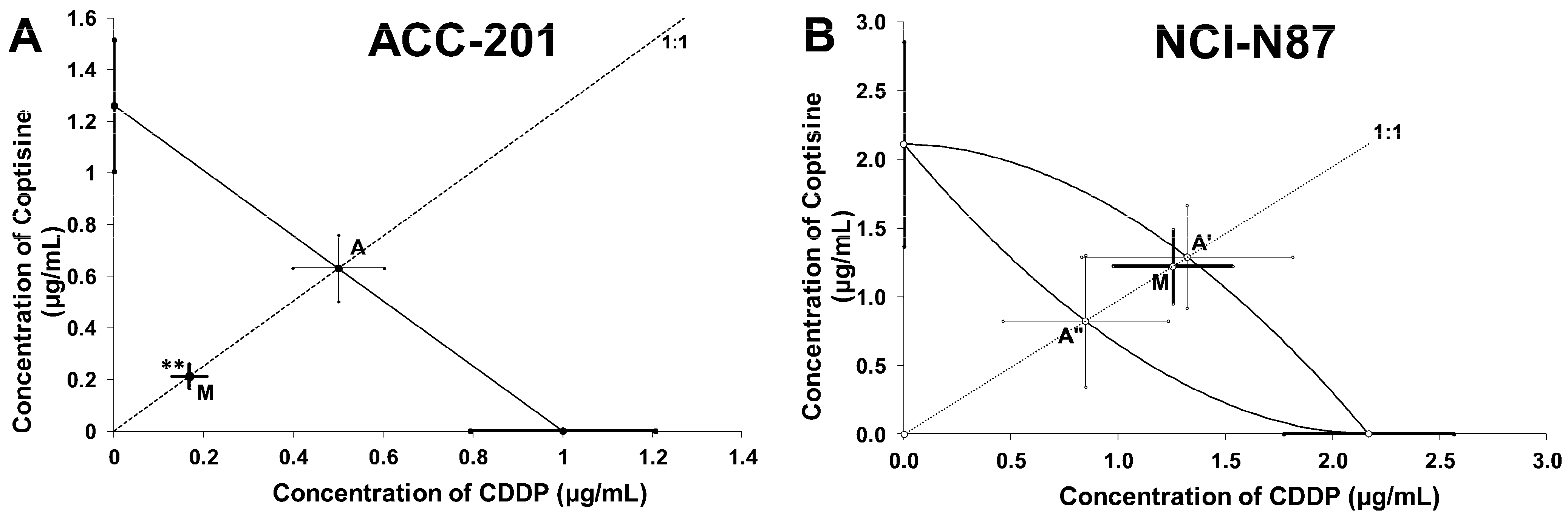
2.5. Isobolographic Analysis of the Interactions between Cisplatin and Coptisine
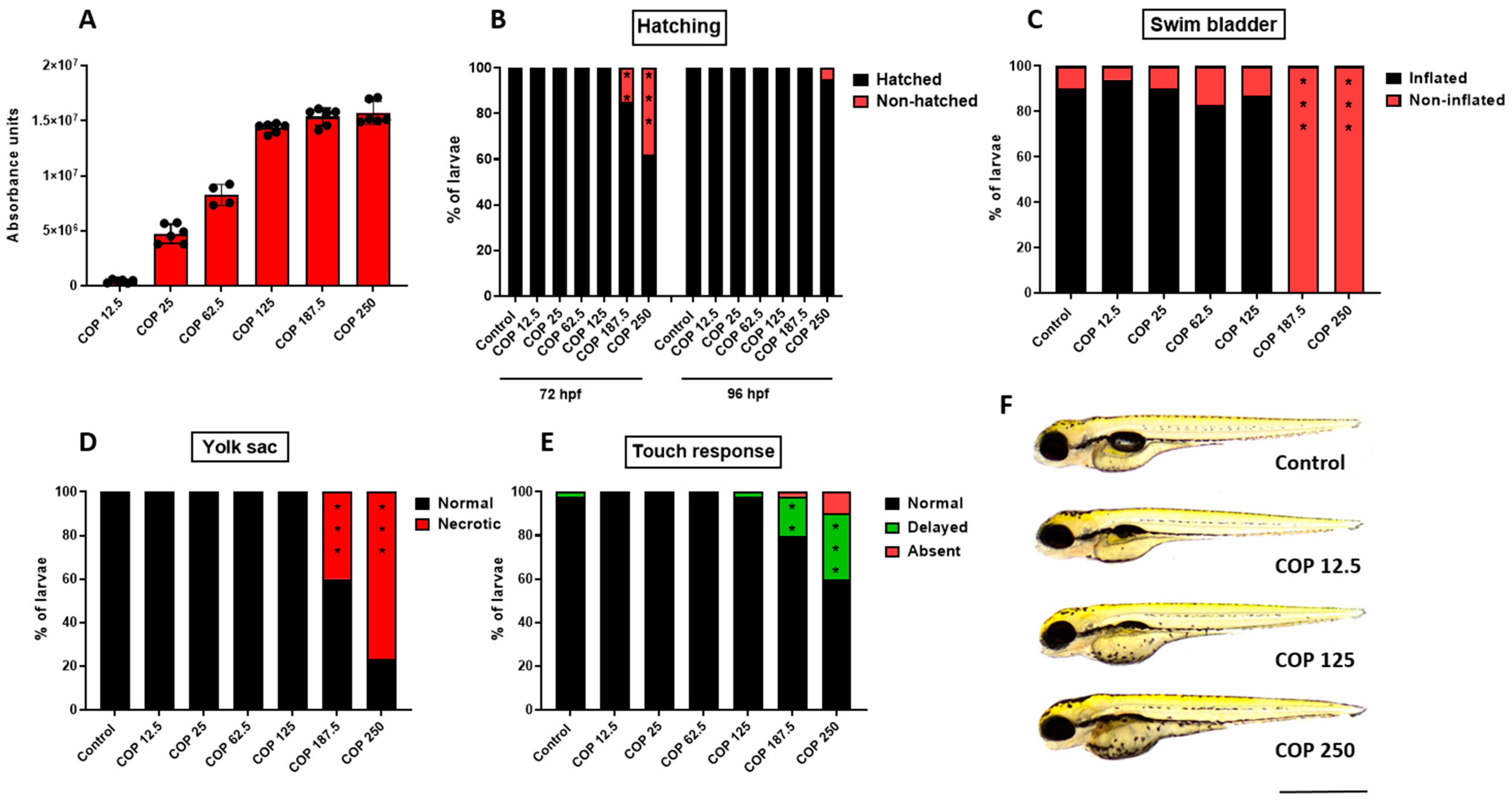
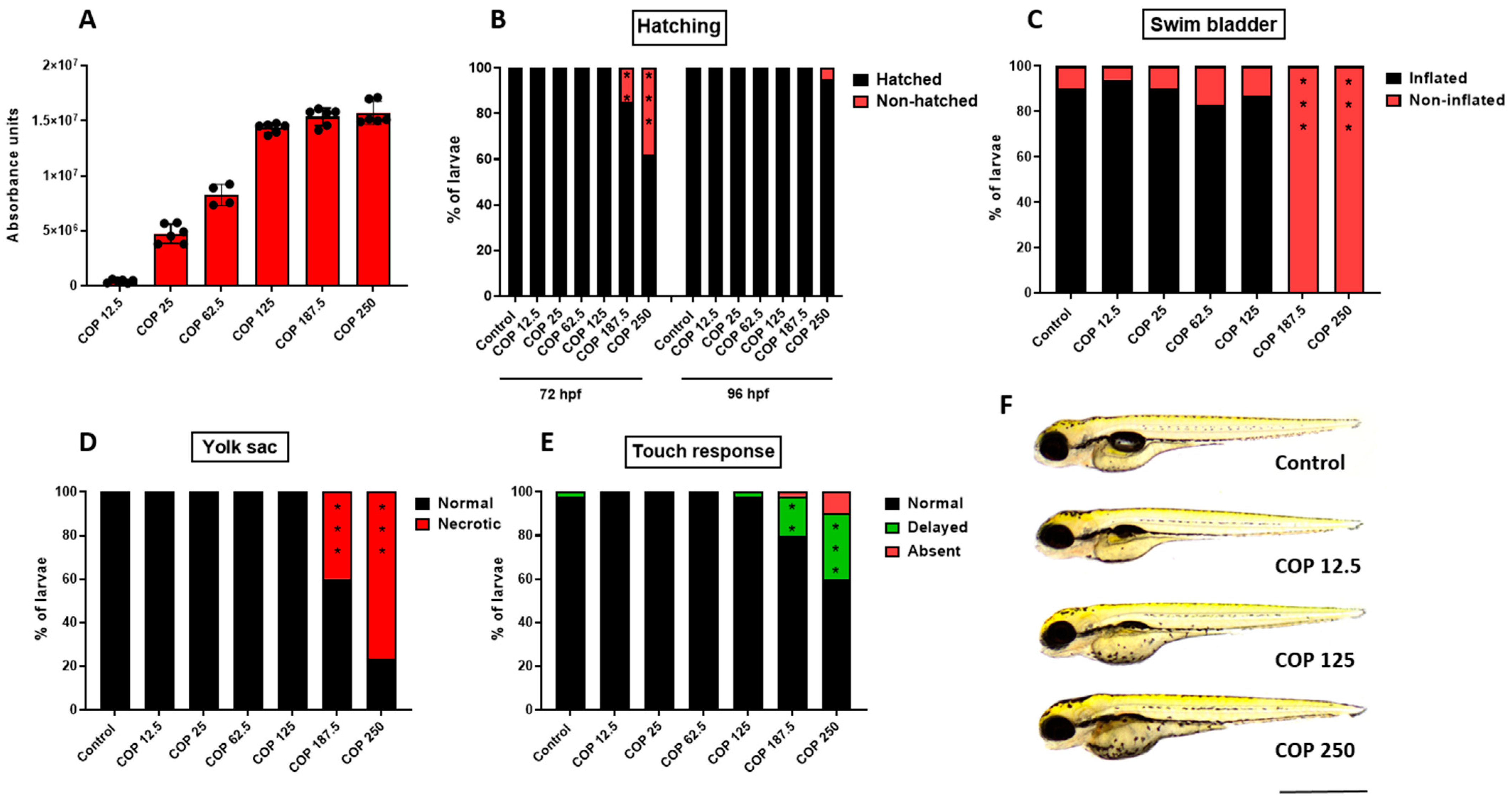
2.6. Effect of Coptisine on Developing Zebrafish
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Plant Material
4.3. Extraction
4.4. Qualitative and Quantitative HPLC-MS Analyses
4.5. Fractionation of Extract by Centrifugal Partition Chromatography
4.6. Cell Lines Culture
4.7. Cell Viability Assay
4.8. Cell Proliferation Assay
4.9. Cell Cycle Analysis
4.10. Active Caspase-3/7 Apoptosis Assay
4.11. The Pharmacological Interaction between Coptisine and CDDP with Isobolographic Analysis
4.12. Zebrafish Experiments
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, Z.; Wu, Y.; Yang, J.; Yang, D.; Fang, X. Progress in the treatment of advanced gastric cancer. Tumour Biol. 2017, 39, 1010428317714626. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Cisło, M.; Filip, A.; Arnold Offerhaus, G.; Ciseł, B.; Rawicz-Pruszyński, K.; Skierucha, M.; Polkowski, W. Distinct molecular subtypes of gastric cancer: From Laurén to molecular pathology. Oncotarget 2018, 9, 19427. [Google Scholar] [CrossRef]
- Poonyam, P.; Aumpan, N.; Vilaichone, R. Prognostic factors for survival in patients with gastric adenocarcinoma. Cancer Rep. 2021, 4, e1305. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.; Fischbach, L.; Delongchamp, R.; Faramawi, M.; Orloff, M. Association between Dietary Salt Intake and Progression in the Gastric Precancerous Process. Cancers 2019, 11, 467. [Google Scholar] [CrossRef]
- Toh, J.; Wilson, R. Pathways of Gastric Carcinogenesis, Helicobacter pylori Virulence and Interactions with Antioxidant Systems, Vitamin C and Phytochemicals. Int. J. Mol. Sci. 2020, 21, 6451. [Google Scholar] [CrossRef]
- Koessler, T.; Roth, A.; Cacheux, W. Early gastric cancer: Epidemiology, diagnostic and management. Rev. Med. Suisse 2014, 10, 1118–1122. [Google Scholar]
- Tokunaga, M.; Sato, Y.; Nakagawa, M.; Aburatani, T.; Matsuyama, T.; Nakajima, Y.; Kinugasa, Y. Perioperative chemotherapy for locally advanced gastric cancer in Japan: Current and future perspectives. Surg. Today 2020, 50, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Arai, H.; Nakajima, T. Recent Developments of Systemic Chemotherapy for Gastric Cancer. Cancers 2020, 12, 1100. [Google Scholar] [CrossRef]
- Rayan, A.; Raiyn, J.; Falah, M. Nature is the best source of anticancer drugs: Indexing natural products for their anticancer bioactivity. PloS ONE 2017, 12, e0187925. [Google Scholar] [CrossRef]
- Mathur, S.; Hoskins, C. Drug development: Lessons from nature. Biomed. Rep. 2017, 6, 612–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.; Lu, J.; Ding, J. Natural Products in Cancer Therapy: Past, Present and Future. Nat. Prod. Bioprospect. 2021, 11, 5–13. [Google Scholar] [CrossRef]
- Ali, H.; Dixit, S. Extraction optimization of Tinosporacordifolia and assessment of the anticancer activity of its alkaloid palmatine. Sci. World J. 2013, 2013, 376216. [Google Scholar] [CrossRef]
- Khazir, J.; Riley, D.; Pilcher, L.; De-Maayer, P.; Mir, B. Anticancer agents from diverse natural sources. Nat. Prod. Commun. 2014, 9, 1934578X1400901130. [Google Scholar] [CrossRef]
- Habtemariam, S.; Lentini, G. Plant-Derived Anticancer Agents: Lessons from the Pharmacology of Geniposide and Its Aglycone, Genipin. Biomedicines 2018, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Dehelean, C.; Marcovici, I.; Soica, C.; Mioc, M.; Coricovac, D.; Iurciuc, S.; Cretu, O.; Pinzaru, I. Plant-Derived Anticancer Compounds as New Perspectives in Drug Discovery and Alternative Therapy. Molecules 2021, 26, 1109. [Google Scholar] [CrossRef]
- Heinrich, M.; Mah, J.; Amirkia, V. Alkaloids Used as Medicines: Structural Phytochemistry Meets Biodiversity-An Update and Forward Look. Molecules 2021, 26, 1836. [Google Scholar] [CrossRef]
- Kukula-Koch, W.; Widelski, J. Alkaloids. In Pharmacognosy; Chapter, 9; Badal, S., Delgoda, R., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 163–198. [Google Scholar]
- Wang, J.; Wang, L.; Lou, G.; Zeng, H.; Hu, J.; Huang, Q.; Peng, W.; Yang., X. Coptidis Rhizoma: A comprehensive review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. Pharm. Biol. 2019, 57, 193–225. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia. 2022. Available online: http://wp.chp.org.cn/front/chpint/en/ (accessed on 15 May 2022).
- He, L.; Zhong, Z.; Chen, M.; Liang, Q.; Wang, Y.; Tan, W. Current Advances in Coptidis Rhizoma for Gastrointestinal and Other Cancers. Front. Pharmacol. 2022, 12, 775084. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y. Golden rules and pitfalls in selecting optimum conditions for high-speed counter-current chromatography. J. Chromatogr. A 2005, 1065, 145–168. [Google Scholar] [CrossRef]
- Kukula-Koch, W.; Kruk-Słomka, M.; Stępnik, K.; Szalak, R.; Biała, G. The Evaluation of Pro-Cognitive and Antiamnestic Properties of Berberine and Magnoflorine Isolated from Barberry Species by Centrifugal Partition Chromatography (CPC), in Relation to QSAR Modelling. Int. J. Mol. Sci. 2017, 18, 2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friesen, J.; McAlpine, J.; Chen, S.; Pauli, G. Countercurrent Separation of Natural Products: An Update. J. Nat. Prod. 2015, 78, 1765–1796. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; Zhang, X.; Wang, H.; Li, P.; Liu, Y.; Li, S. Rapid Analysis of Components in CoptischinensisFranch by Ultra-Performance Liquid Chromatography with Quadrupole Time-of-Flight Mass Spectrometry. Pharmacogn. Mag. 2017, 13, 175. [Google Scholar]
- Lin, Z.; Yang, R.; Guan, Z.; Chen, A.; Li, W. Ultra-performance LC separation and quadrupole time-of-flight MS identification of major alkaloids in PlumulaNelumbinis. Phytochem. Anal. 2014, 25, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, F.; Liu, J.; Lee, F.; Wang, X.; Yang, H. Analysis of alkaloids in CoptischinensisFranch by accelerated solvent extraction combined with ultra performance liquid chromatographic analysis with photodiode array and tandem mass spectrometry detections. Anal. Chim. Acta 2008, 613, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Liu, Y.; Cheng, H.; Xu, K.; Wang, G. Coptis chinensis and dried ginger herb combination inhibits gastric tumor growth by interfering with glucose metabolism via LDHA and SLC2A1. J. Ethnopharmacol. 2022, 284, 114771. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Qi, L.; Zhong, F.; Li, Y.; Ke, W.; Ma, Y. Integrated metabolomics and ligand fishing approaches to screen the hypoglycemic ingredients from four Coptis medicines. J. Pharm. Biomed. Anal. 2021, 192, 113655. [Google Scholar] [CrossRef]
- Lou, G.; Xiong, H.; Gan, Q.; Hu, J.; Peng, C.; Yan, Z.; Yan, H.; Huang, Q. UPLC-Q-Orbitrap HRMS Analysis of Coptischinensis Aerial Parts and Its Regulatory Activity on Glucose-lipid Metabolism. Rev. Bras. Farmacogn. 2021, 31, 24–31. [Google Scholar] [CrossRef]
- Kukula-Koch, W. The Elevation of LC-ESI-Q-TOF-MS Response in the Analysis of Isoquinoline Alkaloids from Some Papaveraceae and Berberidaceae Representatives. J. Anal. Methods Chem. 2017, 2017, 8384107. [Google Scholar] [CrossRef]
- Peng, J.; Han, X.; Xu, Y.; Qi, Y.; Xu, L.; Xu, Q. New Approach for Application of High Speed Countercurrent Chromatography Coupled with Direct Injection of the Powders of a Raw Material without any Preparation, for Isolation and Separation of Four Alkaloids with High Recoveries from Coptis chinensis Franch. J. Liq. Chromatogr. Relat. Technol. 2007, 30, 2929–2940. [Google Scholar]
- Sun, C.; Li, J.; Wang, X.; Duan, W.; Zhang, T.; Ito, Y. Preparative separation of quaternary ammonium alkaloids from CoptischinensisFranch by pH-zone-refining counter-current chromatography. J. Chromatogr. A 2014, 1370, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, T.; Zhang, R.; Ito, Y. Application of analytical and preparative high-speed counter-current chromatography for separation of alkaloids from Coptis chinensis Franch. J. Chromatogr. A 1998, 829, 137–141. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, M.; Wang, C. Preparative separation and purification of alkaloids from Rhizomacoptidis by high-speed counter-current chromatography. Sep. Purif. Technol. 2011, 76, 428–431. [Google Scholar] [CrossRef]
- Kim, J.-B. Isolation of Berberine from the Rhizome of Coptis chinensis by Centrifugal Partition Chromatography. Korean J. Food Nutr. 2011, 24, 617–621. [Google Scholar] [CrossRef]
- Sun, M.; Liu, J.; Lin, C.; Miao, L.; Lin, L. Alkaloid profiling of the traditional Chinese medicine Rhizoma corydalis using high performance liquid chromatography-tandem quadrupole time-of-flight mass spectrometry. Acta Pharm. Sin. B 2014, 4, 208–216. [Google Scholar] [CrossRef]
- Basera, I.; Girme, A.; Bhatt, V.; Saste, G.; Pawar, S.; Hingorani, L.; Shah, M. Development of validated UHPLC–PDA with ESI–MS-MS method for concurrent estimation of magnoflorine, berbamine, columbamine, jatrorrhizine, palmatine and berberine in Berberisaristata. Acta Chromatogr. 2021, 34, 412–421. [Google Scholar] [CrossRef]
- Shim, H.; Lee, J.; Kim, B.; Hong, J. General Fragmentations of Alkaloids in Electrospray Ionization Tandem Mass Spectrometry. Mass Spectrom. Lett. 2013, 4, 79–82. [Google Scholar] [CrossRef]
- Nichani, K.; Li, J.; Suzuki, M.; Houston, J. Evaluation of Caspase-3 Activity During Apoptosis with Fluorescence Lifetime-Based Cytometry Measurements and Phasor Analyses. Cytometry. Part A J. Int. Soc. Anal. Cytol. 2020, 97, 1265–1275. [Google Scholar] [CrossRef]
- Litchfield, J.; Wilcoxon, F. A simplified method of evaluating dose-effect experiments. J. Pharmacol. Exp. Ther. 1949, 96, 99–113. [Google Scholar] [PubMed]
- Test No. 236: Fish Embryo Acute Toxicity (FET) Test. [Text]. 2022. Available online: https://www.oecd-ilibrary.org/environment/test-no-236-fish-embryo-acute-toxicity-fet-test_9789264203709-en (accessed on 20 July 2022).
- Kumar, A.; Jaitak, V. Natural products as multidrug resistance modulators in cancer. Eur. J. Med. Chem. 2019, 176, 268–291. [Google Scholar] [CrossRef] [PubMed]
- Tuzimski, T.; Petruczynik, A.; Kaproń, B.; Makuch-Kocka, A.; Szultka-Młyńska, M.; Misiurek, J.; Szymczak, G.; Buszewski, B. Determination of Cytotoxic Activity of Selected Isoquinoline Alkaloids and Plant Extracts Obtained from Various Parts of Mahoniaaquifolium Collected in Various Vegetation Seasons. Molecules 2021, 26, 816. [Google Scholar] [CrossRef] [PubMed]
- Qing, Z.; Huang, J.; Yang, X.; Liu, J.; Cao, H.; Xiang, F.; Cheng, P.; Zeng, J. Anticancer and Reversing Multidrug Resistance Activities of Natural Isoquinoline Alkaloids and their Structure-activity Relationship. Curr. Med. Chem. 2018, 25, 5088–5114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, X.; Cao, S.; Sun, Y.; He, X.; Jiang, B.; Yu, Y.; Duan, J.; Qiu, F.; Kang, N. Berberine represses human gastric cancer cell growth in vitro and in vivo by inducing cytostatic autophagy via inhibition of MAPK/mTOR/p70S6K and Akt signaling pathways. Biomed. Pharmacother. Biomed. Pharmacother. 2020, 128, 110245. [Google Scholar] [CrossRef]
- Yun, D.; Yoon, S.; Park, S.; Park, Y. The Anticancer Effect of Natural Plant Alkaloid Isoquinolines. Int. J. Mol. Sci. 2021, 22, 1653. [Google Scholar] [CrossRef]
- Grabarska, A.; Wróblewska-Łuczka, P.; Kukula-Koch, W.; Łuszczki, J.; Kalpoutzakis, E.; Adamczuk, G.; Skaltsounis, A.; Stepulak, A. Palmatine, a Bioactive Protoberberine Alkaloid Isolated from Berberiscretica, Inhibits the Growth of Human Estrogen Receptor-Positive Breast Cancer Cells and Acts Synergistically and Additively with Doxorubicin. Molecules 2021, 26, 6253. [Google Scholar] [CrossRef]
- Szalak, R.; Kukula-Koch, W.; Matysek, M.; Kruk-Słomka, M.; Koch, W.; Czernicka, L.; Khurelbat, D.; Biała, G.; Arciszewski, M. Effect of Berberine Isolated from Barberry Species by Centrifugal Partition Chromatography on Memory and the Expression of Parvalbumin in the Mouse Hippocampus Proper. Int. J. Mol. Sci. 2021, 22, 4487. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Ng, L.T.; Hsu, F.F.; Shieh, D.E.; Chiang, L.C. Cytotoxic effects of Coptis chinensis and Epimediumsagittatum extracts and their major constituents (berberine, coptisine and icariin) on hepatoma and leukaemia cell growth. Clin. Exp. Pharmacol. Physiol. 2004, 31, 65–69. [Google Scholar] [CrossRef]
- Inoue, N.; Terabayashi, T.; Takiguchi-Kawashima, Y.; Fujinami, D.; Matsuoka, S.; Kawano, M.; Tanaka, K.; Tsumura, H.; Ishizaki, T.; Narahara, H.; et al. The benzylisoquinoline alkaloids, berberine and coptisine, act against camptothecin-resistant topoisomerase I mutants. Sci. Rep. 2021, 11, 7718. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, Y.; Yan, X.; Wang, W.; Tian, X.; Wang, L.; Zhu, W.; Gong, L.; Pan, G. Different structures of berberine and five other protoberberine alkaloids that affect P-glycoprotein-mediated efflux capacity. Acta Pharmacol. Sin. 2019, 40, 133–142. [Google Scholar] [CrossRef]
- Ai, X.; Yu, P.; Peng, L.; Luo, L.; Liu, J.; Li, S.; Lai, X.; Luan, F.; Meng, X. Berberine: A Review of its Pharmacokinetics Properties and Therapeutic Potentials in Diverse Vascular Diseases. Front. Pharmacol. 2021, 12, 762654. [Google Scholar] [CrossRef]
- Leyva-Peralta, M.; Robles-Zepeda, R.; Razo-Hernández, R.; Berber, L.; Lara, K.; Ruiz-Bustos, E.; Gálvez-Ruíz, J. Berberine as Source of Antiproliferative Hybrid Compounds: In Vitro Antiproliferative Activity and Quantitative Structure-activity Relationship. Anti Cancer Agents Med. Chem. 2019, 19, 1820–1834. [Google Scholar] [CrossRef]
- Xie, L.; Feng, S.; Zhang, X.; Zhao, W.; Feng, J.; Ma, C.; Wang, R.; Song, W.; Cheng, J. Biological Response Profiling Reveals the Functional Differences of Main Alkaloids in RhizomaCoptidis. Molecules 2021, 26, 7389. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ma, L.; Yan, D.; Zhang, C.; Gao, D.; Xiong, Y.; Sheng, F.; Dong, X.; Xiao, X. Dynamic monitoring of the cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence system. Chin. J. Nat. Med. 2014, 12, 428–435. [Google Scholar] [PubMed]
- Guo, Y.; Pan, W.; Liu, S.; Shen, Z.; Xu, Y.; Hu, L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef]
- Yu, D.; Fu, S.; Cao, Z.; Bao, M.; Zhang, G.; Pan, Y.; Liu, W.; Zhou, Q. Unraveling the novel anti-osteosarcoma function of coptisine and its mechanisms. Toxicol. Lett. 2014, 226, 328–336. [Google Scholar] [CrossRef]
- Rao, P.; Begum, S.; Sahai, M.; Sriram, D. Coptisine-induced cell cycle arrest at G2/M phase and reactive oxygen species-dependent mitochondria-mediated apoptosis in non-small-cell lung cancer A549 cells. Tumour Biol. 2017, 39, 1010428317694565. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Zhang, X.; Miao, X.Z.; Yuan, Y.Y.; Gao, J.; Li, X.; Liu, Y.G.; Tan, P. Coptisine suppresses proliferation and inhibits metastasis in human pancreatic cancer PANC-1 cells. J. Asian Nat. Prod. Res. 2020, 22, 452–463. [Google Scholar] [CrossRef]
- Han, B.; Jiang, P.; Li, Z.; Yu, Y.; Huang, T.; Ye, X.; Li, X. Coptisine-induced apoptosis in human colon cancer cells (HCT-116) is mediated by PI3K/Akt and mitochondrial-associated apoptotic pathway. Phytomed. Int. J. Phytother. Phytopharm. 2018, 48, 152–160. [Google Scholar] [CrossRef]
- Kim, S.; Hwangbo, H.; Lee, H.; Park, C.; Kim, G.; Moon, S.; Yun, S.; Kim, W.; Cheong, J.; Choi, Y. Induction of Apoptosis by Coptisine in Hep3B Hepatocellular Carcinoma Cells through Activation of the ROS-Mediated JNK Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 5502. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, F.; Li, G.; Huang, J.; Liu, Y.; Zhang, Q.; Tang, Q.; Hu, C.; Zhang, R. Coptisine Induces Apoptosis in Human Hepatoma Cells Through Activating 67-kDa Laminin Receptor/cGMP Signaling. Front. Pharmacol. 2018, 9, 517. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Hong, S.; Li, Y.; Chen, H.; Shen, Y.; Shao, K.; Lu, M.; Dai, H.; Ma, S.; Dai, G. Coptisine suppresses tumor growth and progression by down-regulating MFG-E8 in colorectal cancer. RSC Adv. 2018, 8, 30937–30945. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Ye, X.; Wu, H.; Wang, Y.; Zou, Z.; Ning, N.; Hu, Y.; Chen, B.; Fang, X.; Li, X. The safety and anti-hypercholesterolemic effect of coptisine in Syrian golden hamsters. Lipids 2015, 50, 185–194. [Google Scholar] [CrossRef]
- Yi, J.; Ye, X.; Wang, D.; He, K.; Yang, Y.; Liu, X.; Li, X. Safety evaluation of main alkaloids from Rhizoma Coptidis. J. Ethnopharmacol. 2013, 145, 303–310. [Google Scholar] [CrossRef]
- Li, Y.; Wang, H.; Si, N.; Ren, W.; Han, L.; Xin, S.; Zuo, R.; Wei, X.; Yang, J.; Zhao, H.; et al. Metabolic profiling analysis of berberine, palmatine, jatrorrhizine, coptisine and epiberberine in zebrafish by ultra-high performance liquid chromatography coupled with LTQ Orbitrap mass spectrometer. Xenobiotica 2015, 45, 302–311. [Google Scholar] [CrossRef]
- Hu, Y.; Ma, H.; Zou, Z.; He, K.; Xiao, Y.; Wang, Y.; Feng, M.; Ye, X.; Li, X. Activation of Akt and JNK/Nrf2/NQO1 pathway contributes to the protective effect of coptisine against AAPH-induced oxidative stress. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 85, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, Z.; Sun, J.; Lv, H.; Wang, Y.; Ni, Y.; Chen, S.; Hu, C.; Wang, L.; Chen, W.; et al. Cisplatin resistance in gastric cancer cells is involved with GPR30-mediated epithelial-mesenchymal transition. J. Cell. Mol. Med. 2020, 24, 3625–3633. [Google Scholar] [CrossRef]
- Hussain, Y.; Islam, L.; Khan, H.; Filosa, R.; Aschner, M.; Javed, S. Curcumin-cisplatin chemotherapy: A novel strategy in promoting chemotherapy efficacy and reducing side effects. Phytother. Res. 2021, 35, 6514–6529. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhang, Y.; Zhou, Y.; Lian, X.; Yan, L.; Pan, T.; Jin, T.; Xie, H.; Liang, Z.; Qiu, W.; et al. NPCDR: Natural product-based drug combination and its disease-specific molecular regulation. Nucleic Acids Res. 2022, 50, D1324–D1333. [Google Scholar] [CrossRef]
- Wróblewska-Łuczka, P.; Grabarska, A.; Florek-Łuszczki, M.; Plewa, Z.; Łuszczki, J. Synergy, Additivity, and Antagonism between Cisplatin and Selected Coumarins in Human Melanoma Cells. Int. J. Mol. Sci. 2021, 22, 537. [Google Scholar] [CrossRef]
- Okon, E.; Luszczki, J.; Kukula-Koch, W.; Halasa, M.; Jarzab, A.; Khurelbat, D.; Stepulak, A.; Wawruszak, A. Synergistic or Additive Pharmacological Interactions between Magnoflorine and Cisplatin in Human Cancer Cells of Different Histological Origin. Int. J. Mol. Sci. 2020, 21, 2848. [Google Scholar] [CrossRef]
- Jarząb, A.; Łuszczki, J.; Guz, M.; Skalicka-Woźniak, K.; Hałasa, M.; Smok-Kalwat, J.; Polberg, K.; Stepulak, A. Combination of Osthole and CisplatinAgainstRhabdomyosarcoma TE671 Cells Yielded Additive Pharmacologic Interaction by Means of Isobolographic Analysis. Anticancer Res. 2018, 38, 205–210. [Google Scholar]
- Castañeda, A.; Meléndez, C.; Uribe, D.; Pedroza-Díaz, J. Synergistic effects of natural compounds and conventional chemotherapeutic agents: Recent insights for the development of cancer treatment strategies. Heliyon 2022, 8, e09519. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Jing, Z.; Li, Y.; Mao, W. Berberine in combination with cisplatin suppresses breast cancer cell growth through induction of DNA breaks and caspase-3-dependent apoptosis. Oncol. Rep. 2016, 36, 567–572. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P. Analysis of Cell Viability by the MTT Assay. Cold Spring Harb. Protoc. 2018. [Google Scholar] [CrossRef]
- Grabarska, A.; Skalicka-Woźniak, K.; Kiełbus, M.; Dmoszyńska-Graniczka, M.; Miziak, P.; Szumiło, J.; Nowosadzka, E.; Kowalczuk, K.; Khalifa, S.; Smok-Kalwat, J.; et al. Imperatorin as a Promising Chemotherapeutic Agent Against Human Larynx Cancer and Rhabdomyosarcoma Cells. Molecules 2020, 25, 2046. [Google Scholar] [CrossRef] [PubMed]
- Luszczki, J.; Czuczwar, S. Isobolographic profile of interactions between tiagabine and gabapentin: A preclinical study. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2004, 369, 434–446. [Google Scholar] [CrossRef]
- Luszczki, J.; Czuczwar, S. Biphasic characteristic of interactions between stiripentol and carbamazepine in the mouse maximal electroshock-induced seizure model: A three-dimensional isobolographic analysis. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2006, 374, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Luszczki, J. Isobolographic analysis of interaction between drugs with nonparallel dose-response relationship curves: A practical application. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2007, 375, 105–114. [Google Scholar] [CrossRef]
- Grabovsky, Y.; Tallarida, R. Isobolographic analysis for combinations of a full and partial agonist: Curved isoboles. J. Pharmacol. Exp. Ther. 2004, 310, 981–986. [Google Scholar] [CrossRef]
- Tallarida, R.J. An overview of drug combination analysis with isobolograms. J. Pharmacol. Exp. Ther. 2006, 319, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tallarida, R.J. Interactions between drugs and occupied receptors. Pharmacol. Ther. 2007, 113, 197–209. [Google Scholar] [CrossRef]
- Gawel, K.; Kukula-Koch, W.; Banono, N.; Nieoczym, D.; Targowska-Duda, K.; Czernicka, L.; Parada-Turska, J.; Esguerra, C. 6-Gingerol, a Major Constituent of ZingiberofficinaleRhizoma, Exerts Anticonvulsant Activity in the Pentylenetetrazole-Induced Seizure Model in Larval Zebrafish. Int. J. Mol. Sci. 2021, 22, 7745. [Google Scholar] [CrossRef]
- Gawel, K.; Turski, W.; van der Ent, W.; Mathai, B.; Kirstein-Smardzewska, K.; Simonsen, A.; Esguerra, C. Phenotypic Characterization of Larval Zebrafish (Daniorerio) with Partial Knockdown of the cacna1a Gene. Mol. Neurobiol. 2020, 57, 1904–1916. [Google Scholar] [CrossRef]
- Sztal, T.; Ruparelia, A.; Williams, C.; Bryson-Richardson, R. Using Touch-evoked Response and Locomotion Assays to Assess Muscle Performance and Function in Zebrafish. JoVE J. Vis. Exp. 2022, 116, e54431. [Google Scholar] [CrossRef] [Green Version]
- Tallarida, R. Drug Combinations: Tests and Analysis with Isoboles. Curr. Protoc. Pharmacol. 2016, 72, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Ion (+/−) | Rt (min) | Molecular Formula | m/z Calculated | m/z Experimental | Error (ppm) | DBE | MS/MS Fragments | Proposed Compound | References |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | [M + H]+ | 6.6 | C20H24NO4+ | 342.1700 | 342.1733 | −9.72 | 10 | 297, 265, 237 | Magnoflorine | This study, [25] |
| 2 | [M + H]+ | 8.9 | C19H23NO3 | 314.1751 | 314.1777 | −8.4 | 9 | 269, 237 | 4′-O-Methyl-N-methylcoclaurine | This study, [26] |
| 3 | [M + H]+ | 10.1 | C21H26NO4 | 356.1856 | 356.1888 | −8.9 | 10 | 206, 175 | Menisperine | This study, [25] |
| 4 | [M + H]+ | 10.5 | C21H25NO5 | 372.1805 | 372.1833 | −7.41 | 10 | 357, 222, 162 | Stecepharine | This study, [25] |
| 5 | [M + H]+ | 10.7 | C19H17NO4 | 324.1230 | 324.1245 | −4.54 | 12 | 309, 294, 266 | Demethyleneberberine/isomer | This study, [25] |
| 6 | [M + H]+ | 10.85 | C19H16NO4+ | 322.1074 | 322.1100 | −8.15 | 13 | 250, 192 | Berberrubine/Thalifendine | [27] |
| 7 | [M + H]+ | 11.4 | C20H17NO5+ | 352.1153 | 352.1190 | −2.99 | 13 | 336, 322, 308, 294 | 13-methyljatrorrhizine/13-methylcolumbamine | This study, [25] |
| 8 | [M + H]+ | 11.7 | C20H20NO4 | 338.1413 | 338.1387 | −7.76 | 12 | 322, 308, 294, 280 | Columbamine | This study, [25] |
| 9 | [M + H]+ | 11.95 | C20H18NO4 | 336.1230 | 336.1239 | −2.58 | 13 | 320, 292, 280, 262 | Epiberberine | This study, [25,28] |
| 10 | [M + H]+ | 12.2 | C20H19NO4 | 338.1387 | 338.1415 | −8.35 | 12 | 322, 307, 294, 280 | Jatrorrhizine | This study, [25] |
| 11 | [M + H]+ | 12.6 | C19H13NO4 | 320.0917 | 320.0942 | −7.73 | 14 | 320, 292, 277, 262 | Coptisine | This study, [25] |
| 12 | [M + H]+ | 13.1 | C21H21NO4 | 352.1543 | 352.1575 | −9.01 | 12 | 337, 322, 308, 291 | Palmatine | This study, [25] |
| 13 | [M + H]+ | 13.5 | C20H17NO4 | 336.1230 | 336.1256 | −7.66 | 13 | 321, 306, 292, 275 | Berberine | This study, [25] |
| 14 | [M + H]+ | 17.0 | C21H19NO4 | 350.1387 | 350.1415 | −8.06 | 13 | 335, 320, 306, 292 | 13-Methylberberine | This study, [25] |
| 15 | [M + H]+ | 10.6 | C20H15NO4 | 334.1074 | 334.1075 | −0.35 | 14 | 319, 304, 290, 276 | Methylcoptisine | This study, [29] |
| 16 | [M + H]+ | 8.6 | C20H23NO4 | 342.1700 | 342.1668 | 9.33 | 10 | 297, 285, 265, 188 | Phellodendrine | This study, [30] |
| 17 | [M + H]+ | 9.8 | C20H21NO4 | 340.1543 | 340.1576 | −9.63 | 11 | 325, 308, 192 | Tetrahydroberberine (canadine) | This study, [30] |
| 18 | [M + H]+ | 16.5 | C20H15NO4 | 334.1074 | 334.1077 | −0.95 | 14 | 321, 304, 292, 278 | Worenine | This study, [19] |
| Compound | IC50 Value ± S.E.M. | |
|---|---|---|
| ACC-201 Cells | NCI-N87 Cells | |
| Berberine (13) | 0.999 µg/mL (2.97 µM) ± 0.013 | 2.023 µg/mL (6.01 µM) ± 0.016 |
| Coptisine (11) | 1.260 µg/mL (3.93 µM) ± 0.250 | 2.110 µg/mL (6.58 µM) ± 0.750 |
| Jatrorrhizine (10) | 12.15 µg/mL (35.90 µM) ± 0.016 | 17.85 µg/mL (52.75 µM) ± 0.018 |
| Palmatine (12) | 4.909 µg/mL (13.93 µM) ± 0.017 | 20.08 µg/mL (56.98 µM) ± 0.017 |
| IC50exp (μg/mL) | nexp | IC50add (μg/mL) | nadd | Interaction |
|---|---|---|---|---|
| 0.38 ± 0.09 | 96 | 1.13 ± 0.23 ** | 236 | Synergy |
| IC50exp (μg/mL) | nexp | L-IC50add (μg/mL) | nadd | U-IC50add (μg/mL) | Interaction |
|---|---|---|---|---|---|
| 2.476 ± 0.550 | 96 | 0.824 ± 0.367 | 236 | 1.291 ± 0.944 | Additivity |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakonieczna, S.; Grabarska, A.; Gawel, K.; Wróblewska-Łuczka, P.; Czerwonka, A.; Stepulak, A.; Kukula-Koch, W. Isoquinoline Alkaloids from Coptis chinensis Franch: Focus on Coptisine as a Potential Therapeutic Candidate against Gastric Cancer Cells. Int. J. Mol. Sci. 2022, 23, 10330. https://doi.org/10.3390/ijms231810330
Nakonieczna S, Grabarska A, Gawel K, Wróblewska-Łuczka P, Czerwonka A, Stepulak A, Kukula-Koch W. Isoquinoline Alkaloids from Coptis chinensis Franch: Focus on Coptisine as a Potential Therapeutic Candidate against Gastric Cancer Cells. International Journal of Molecular Sciences. 2022; 23(18):10330. https://doi.org/10.3390/ijms231810330
Chicago/Turabian StyleNakonieczna, Sylwia, Aneta Grabarska, Kinga Gawel, Paula Wróblewska-Łuczka, Arkadiusz Czerwonka, Andrzej Stepulak, and Wirginia Kukula-Koch. 2022. "Isoquinoline Alkaloids from Coptis chinensis Franch: Focus on Coptisine as a Potential Therapeutic Candidate against Gastric Cancer Cells" International Journal of Molecular Sciences 23, no. 18: 10330. https://doi.org/10.3390/ijms231810330
APA StyleNakonieczna, S., Grabarska, A., Gawel, K., Wróblewska-Łuczka, P., Czerwonka, A., Stepulak, A., & Kukula-Koch, W. (2022). Isoquinoline Alkaloids from Coptis chinensis Franch: Focus on Coptisine as a Potential Therapeutic Candidate against Gastric Cancer Cells. International Journal of Molecular Sciences, 23(18), 10330. https://doi.org/10.3390/ijms231810330