
Comparative Proteomic Analysis of Liver Tissues and Serum in db/db Mice


 and
and 
Abstract
:1. Introduction
2. Results
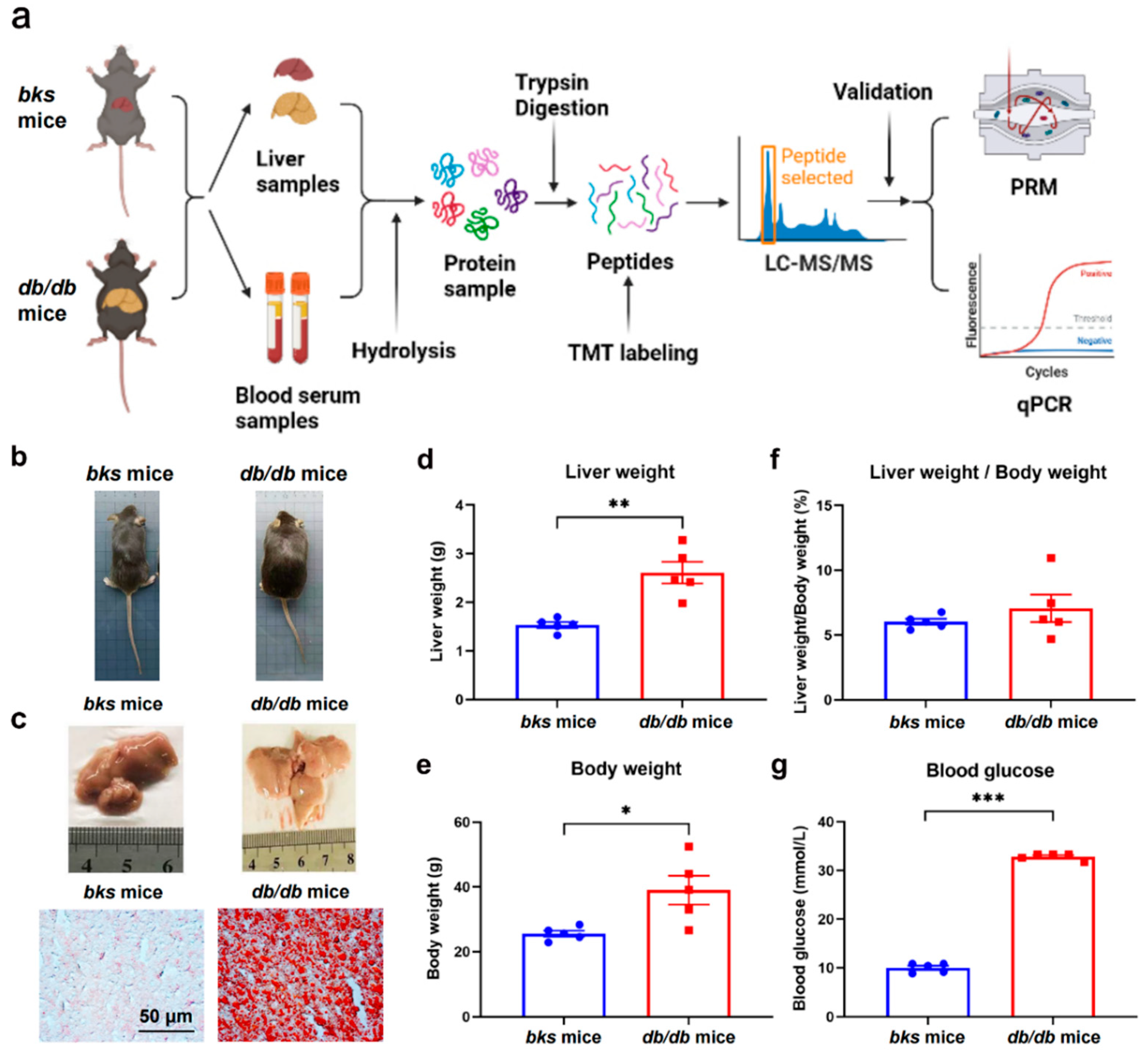
2.1. Validation of NAFLD Mouse Model Based on db/db Mice
2.2. Validations of Data Filtering and Quality Control in the Serum Samples
2.3. Validations of Data Filtering and Quality Control in the Liver Samples
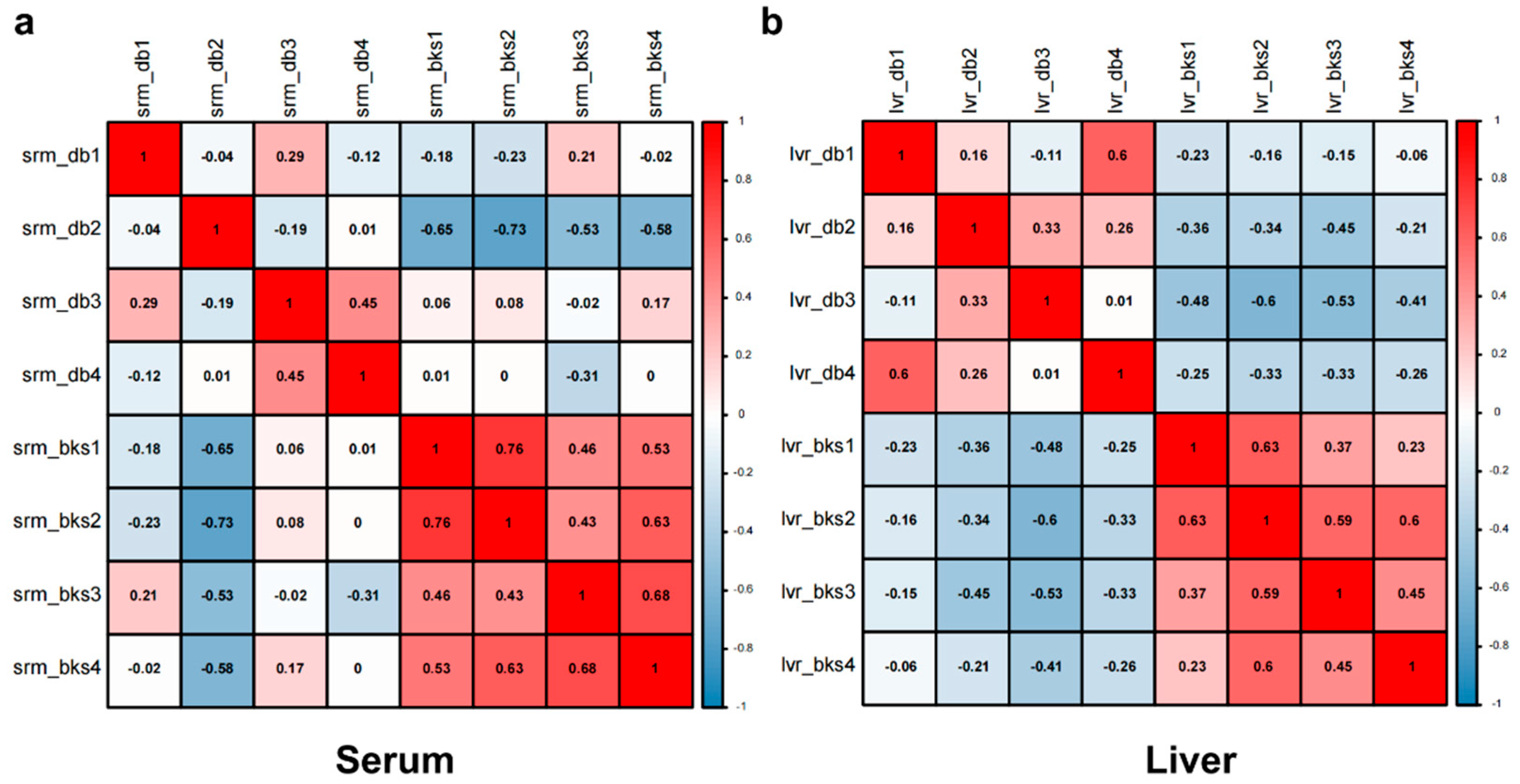
2.4. Biological Repeatability of the Proteome
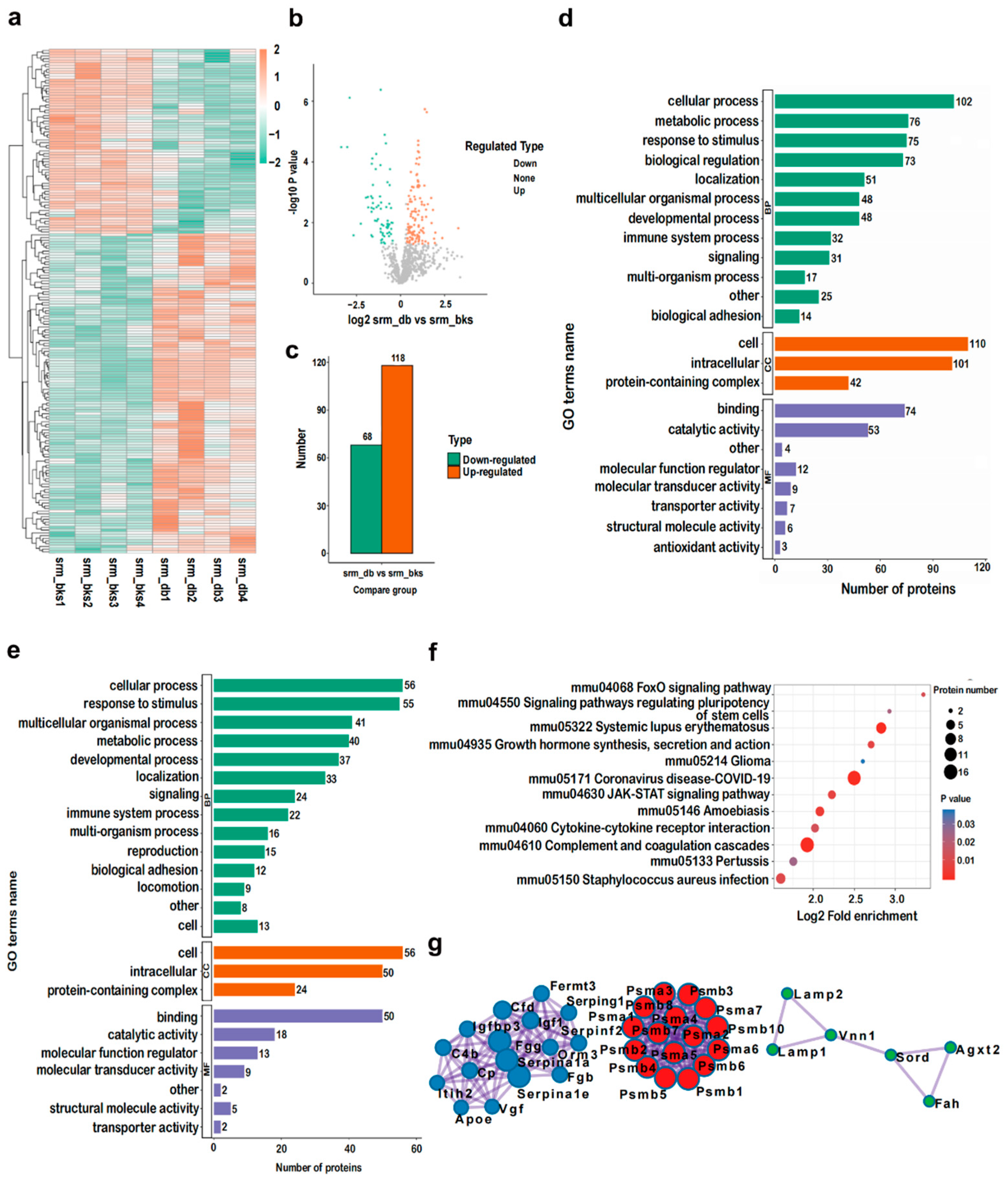
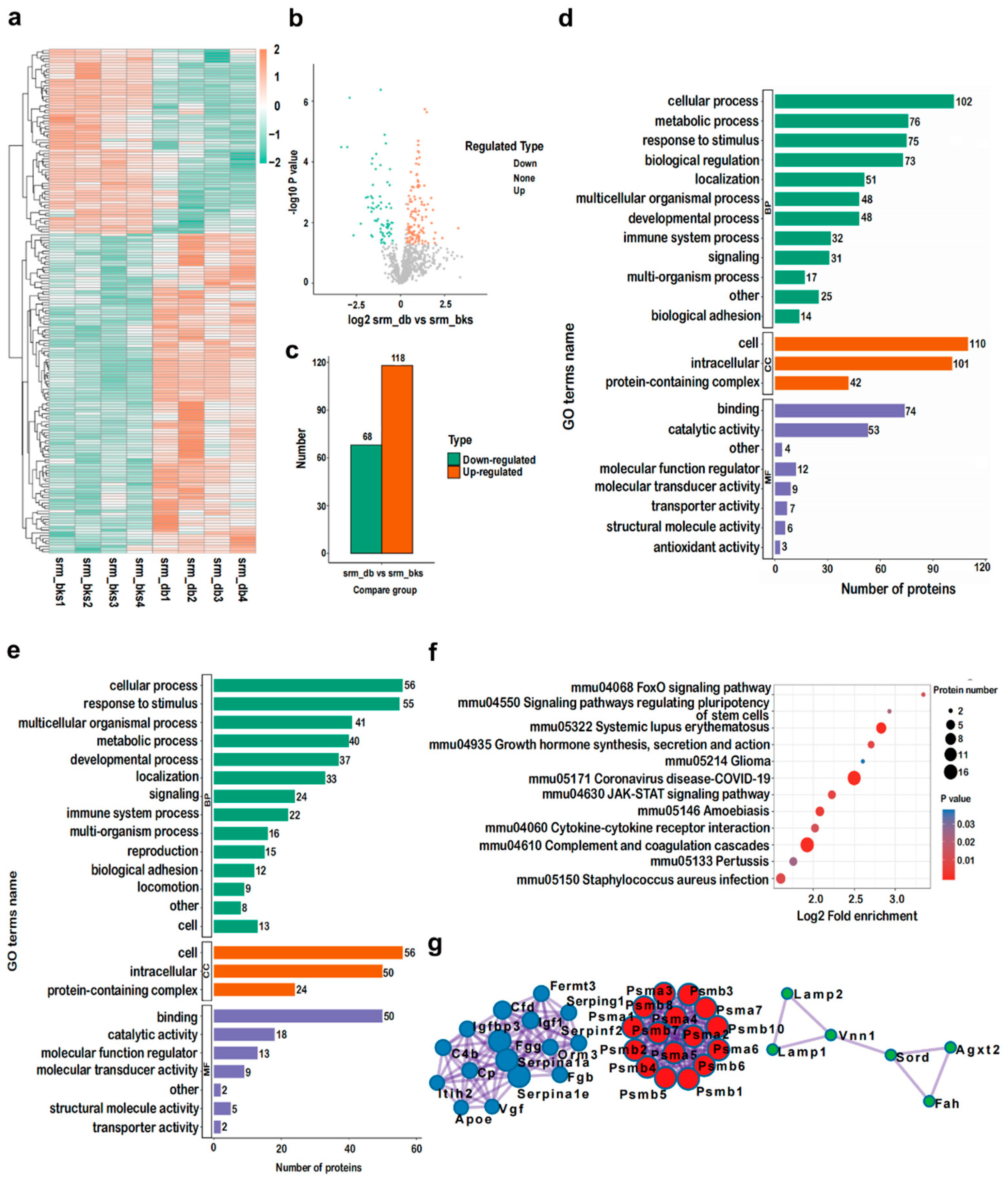
2.5. Identification and Bioinformatic Analysis of Serum DEPs of db/db Mice
2.6. PRM Validations of Serum DEPs
2.7. Identification and Bioinformatic Analysis of Hepatic DEPs in db/db Mice
2.8. PRM Validations of Hepatic DEPs
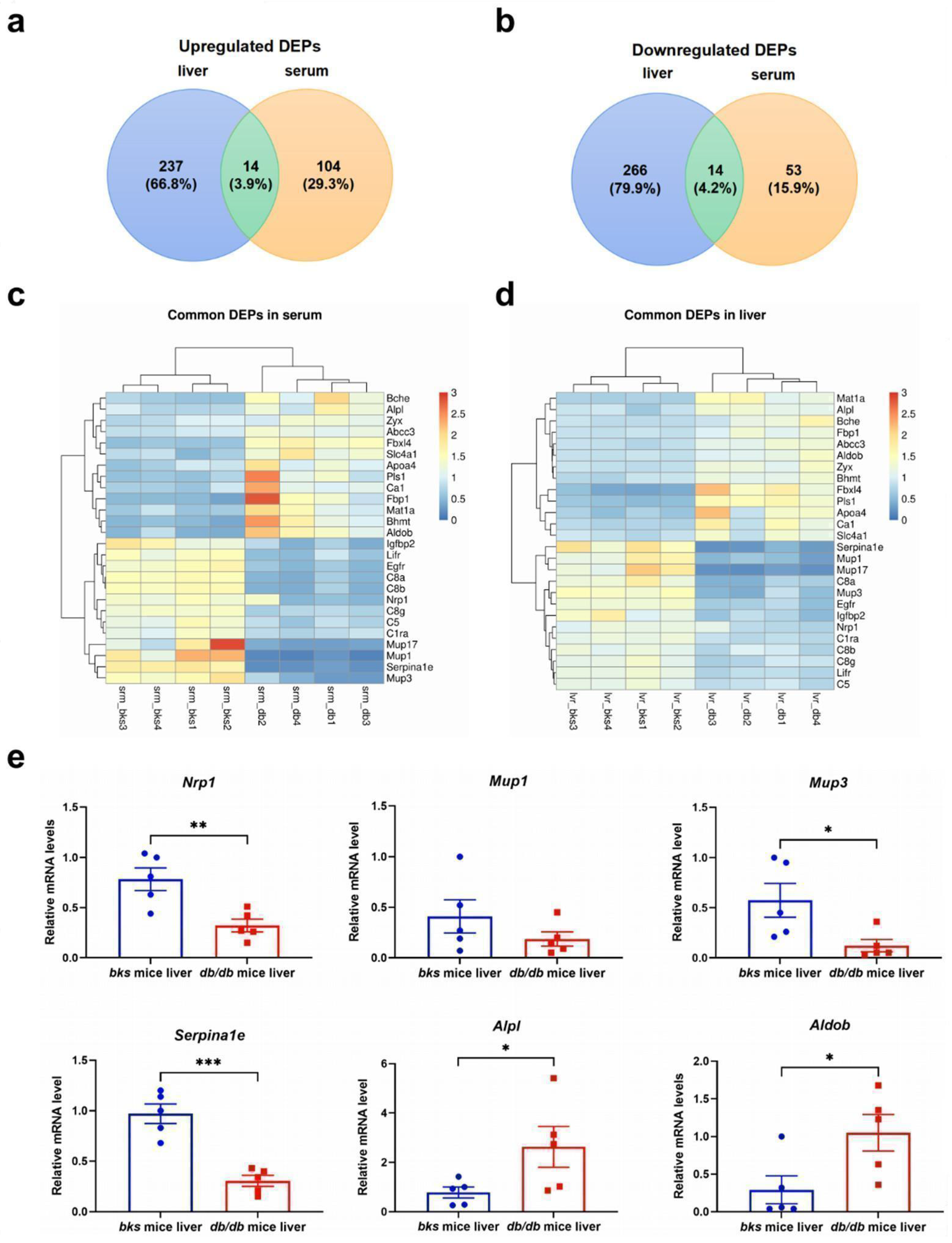
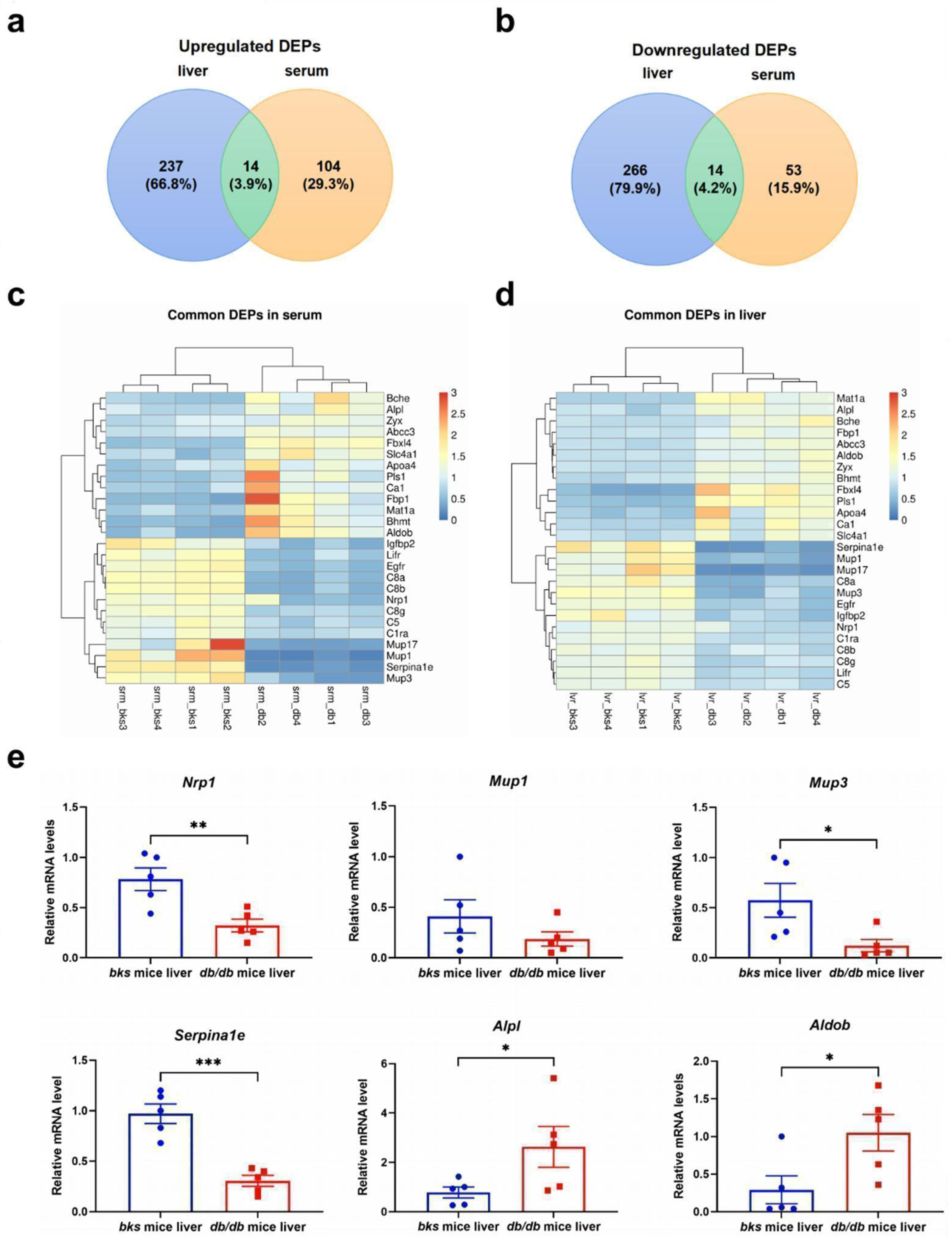
2.9. The Confirmation of Common DEPs in the Serum and Liver of db/db Mice
3. Discussion
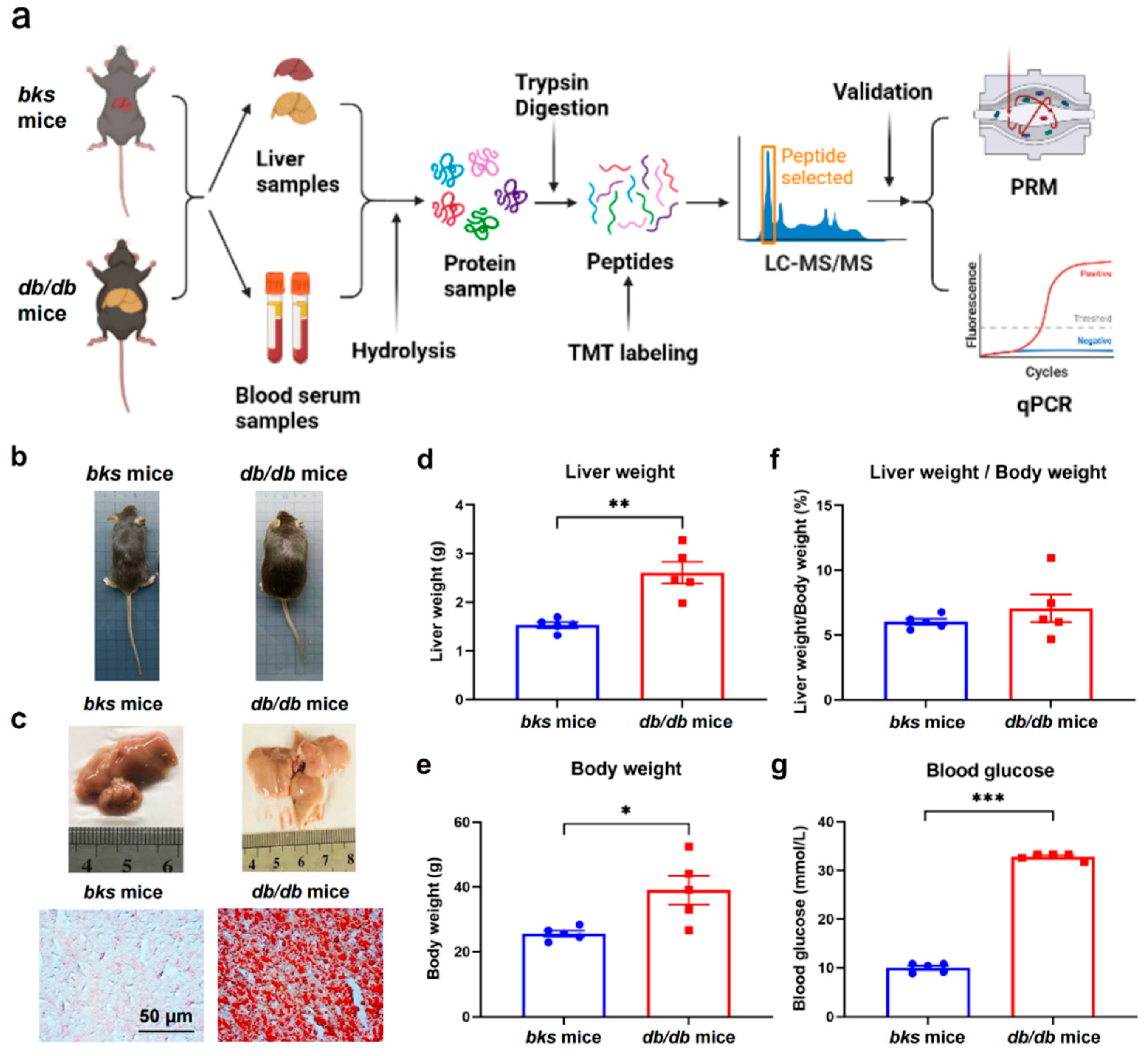
4. Methods
4.1. Animals
4.2. Sample Collections
4.3. Oil Red O Staining
4.4. The Extraction and Digestion of Proteins
4.5. TMT Labeling, HPLC Separation, and LC-MS/MS Analysis
4.6. Database Searches
4.7. Bioinformatics Analysis
4.8. Parallel Reaction Monitoring (PRM) Validation
4.9. RNA Extraction and qRT-PCR
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, C.; Zhou, B.; Meng, M.; Zhao, W.; Wang, D.; Yuan, Y.; Zheng, Y.; Qiu, J.; Li, Y.; Li, G.; et al. FOXA3 induction under endoplasmic reticulum stress contributes to non-alcoholic fatty liver disease. J. Hepatol. 2021, 75, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.P. The definition and classification of metabolic liver disease. Zhonghua Yi Xue Za Zhi 2020, 100, 401–406. [Google Scholar] [PubMed]
- Eslam, M.; Sanyal, A.J.; George, J.; on behalf of theInternational Consensus Panel. MAFLD: A Consensus-Driven Proposed Nomenclature for Metabolic Associated Fatty Liver Disease. Gastroenterology 2020, 158, 1999–2014.e1991. [Google Scholar] [CrossRef] [PubMed]
- Eslam, M.; Newsome, P.N.; Sarin, S.K.; Anstee, Q.M.; Targher, G.; Romero-Gomez, M.; Zelber-Sagi, S.; Wong, V.W.-S.; Dufour, J.-F.; Schattenberg, J.M.; et al. A new definition for metabolic dysfunction-associated fatty liver disease: An international expert consensus statement. J. Hepatol. 2020, 73, 202–209. [Google Scholar] [CrossRef]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef]
- Wong, V.W.-S.; Adams, L.A.; De Lédinghen, V.; Wong, G.L.-H.; Sookoian, S. Noninvasive biomarkers in NAFLD and NASH—Current progress and future promise. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 461–478. [Google Scholar] [CrossRef]
- Piazzolla, V.A.; Mangia, A. Noninvasive Diagnosis of Nafld and Nash. Cells 2020, 9, 1005. [Google Scholar] [CrossRef]
- Di Mauro, S.; Scamporrino, A.; Filippello, A.; Di Pino, A.; Scicali, R.; Malaguarnera, R.; Purrello, F.; Piro, S. Clinical and Molecular Biomarkers for Diagnosis and Staging of Nafld. Int. J. Mol. Sci. 2021, 22, 11905. [Google Scholar] [CrossRef]
- Niu, L.; E Geyer, P.; Albrechtsen, N.J.W.; Gluud, L.L.; Santos, A.; Doll, S.; Treit, P.V.; Holst, J.J.; Knop, F.K.; Vilsbøll, T.; et al. Plasma proteome profiling discovers novel proteins associated with non-alcoholic fatty liver disease. Mol. Syst. Biol. 2019, 15, e8793. [Google Scholar] [CrossRef]
- Zhu, W.; Zhang, Y.; Ren, C.-H.; Cheng, X.; Chen, J.-H.; Ge, Z.-Y.; Sun, Z.-P.; Zhuo, X.; Sun, F.-F.; Chen, Y.-L.; et al. Identification of proteomic markers for ram spermatozoa motility using a tandem mass tag (TMT) approach. J. Proteom. 2020, 210, 103438. [Google Scholar] [CrossRef]
- Zhu, Y.; Aebersold, R.; Mann, M.; Guo, T. SnapShot: Clinical proteomics. Cell 2021, 184, 4840.e1. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.; Schäfer, J.; Kuhn, K.; Kienle, S.; Schwarz, J.; Schmidt, G.; Neumann, T.; Hamon, C. Tandem mass tags: A novel quantification strategy for comparative analysis of complex protein mixtures by MS/MS. Anal. Chem. 2003, 75, 1895–1904. [Google Scholar] [CrossRef] [PubMed]
- Canessa, E.H.; Goswami, M.V.; Alayi, T.D.; Hoffman, E.P.; Hathout, Y. Absolute quantification of dystrophin protein in human muscle biopsies using parallel reaction monitoring (PRM). Biol. Mass Spectrom. 2020, 55, e4437. [Google Scholar] [CrossRef]
- Hu, Y.; Ye, S.; Li, Q.; Yin, T.; Wu, J.; He, J. Quantitative Proteomics Analysis Indicates that Upregulation of lncRNA HULC Promotes Pathogenesis of Glioblastoma Cells. OncoTargets Ther. 2020, 13, 5927–5938. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Pan, Y.; Li, K.; Li, D.; Li, P.; Gao, Z. Proteomics Reveals the Mechanism Underlying the Inhibition of Phytophthora sojae by Propyl Gallate. J. Agric. Food Chem. 2020, 68, 8151–8162. [Google Scholar] [CrossRef]
- Yuan, T.; Cai, M.; Sheng, Y.; Ding, X.; Shen, T.; Li, W.; Huang, H.; Liang, B.; Zhang, X.; Zhu, Q. Differentially expressed proteins identified by TMT proteomics analysis in children with verrucous epidermal naevi. J. Eur. Acad. Dermatol. Venereol. 2021, 35, 1393–1406. [Google Scholar] [CrossRef]
- Sahai, A.; Malladi, P.; Pan, X.; Paul, R.; Melin-Aldana, H.; Green, R.M.; Whitington, P.F. Obese and diabetic db/db mice develop marked liver fibrosis in a model of nonalcoholic steatohepatitis: Role of short-form leptin receptors and osteopontin. Am. J. Physiol. Liver Physiol. 2004, 287, G1035–G1043. [Google Scholar] [CrossRef] [PubMed]
- Moriles, E.K.; Azer, S.A. Alanine Amino Transferase. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2022. [Google Scholar]
- Liu, H.-W.; Kao, H.-H.; Wu, C.-H. Exercise training upregulates SIRT1 to attenuate inflammation and metabolic dysfunction in kidney and liver of diabetic db/db mice. Nutr. Metab. 2019, 16, 22. [Google Scholar] [CrossRef]
- Neuman, M.G.; Cohen, L.B.; Nanau, R.M. Biomarkers in nonalcoholic fatty liver disease. Can. J. Gastroenterol. Hepatol. 2014, 28, 607–618. [Google Scholar] [CrossRef]
- Cao, Y.; Szabolcs, A.; Dutta, S.K.; Yaqoob, U.; Jagavelu, K.; Wang, L.; Leof, E.B.; Urrutia, R.A.; Shah, V.H.; Mukhopadhyay, D. Neuropilin-1 Mediates Divergent R-Smad Signaling and the Myofibroblast Phenotype. J. Biol. Chem. 2010, 285, 31840–31848. [Google Scholar] [CrossRef] [Green Version]
- Arab, J.P.; Cabrera, D.; Sehrawat, T.; Jalan-Sakrikar, N.; Verma, V.K.; Simonetto, D.; Cao, S.; Yaqoob, U.; Leon, J.; Freire, M.; et al. Hepatic stellate cell activation promotes alcohol-induced steatohepatitis through Igfbp3 and SerpinA12. J. Hepatol. 2020, 73, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Elpek, G. Neuropilins and liver. World J. Gastroenterol. 2015, 21, 7065–7073. [Google Scholar] [CrossRef] [PubMed]
- Bondeva, T.; Rüster, C.; Franke, S.; Hammerschmid, E.; Klagsbrun, M.; Cohen, C.D.; Wolf, G. Advanced glycation end-products suppress neuropilin-1 expression in podocytes. Kidney Int. 2009, 75, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Bondeva, T.; Wolf, G. Role of Neuropilin-1 in Diabetic Nephropathy. J. Clin. Med. 2015, 4, 1293–1311. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, I.; Rüster, C.; Franke, S.; Liebisch, M.; Wolf, G. Erythropoietin ameliorates podocyte injury in advanced diabetic nephropathy in the db/db mouse. Am. J. Physiol. Ren. Physiol. 2013, 305, F911–F918. [Google Scholar] [CrossRef] [PubMed]
- Schiekofer, S.; Galasso, G.; Sato, K.; Kraus, B.J.; Walsh, K. Impaired Revascularization in a Mouse Model of Type 2 Diabetes Is Associated with Dysregulation of a Complex Angiogenic-Regulatory Network. Arter. Thromb. Vasc. Biol. 2005, 25, 1603–1609. [Google Scholar] [CrossRef]
- Zhou, Y.; Rui, L. Major Urinary Protein Regulation of Chemical Communication and Nutrient Metabolism. Vitam. Horm. 2010, 83, 151–163. [Google Scholar]
- Hayakawa, J.I.; Nikaido, H.; Koizumi, T. Components of major urinary proteins (MUP’s) in the mouse: Sex, strain, and subspecies differences. J. Hered. 1983, 74, 453–456. [Google Scholar] [CrossRef]
- Shahan, K.; Denaro, M.; Gilmartin, M.; Shi, Y.; Derman, E. Expression of six mouse major urinary protein genes in the mammary, parotid, sublingual, submaxillary, and lachrymal glands and in the liver. Mol. Cell. Biol. 1987, 7, 1947–1954. [Google Scholar]
- Shaw, P.H.; Held, W.A.; Hastie, N.D. The gene family for major urinary proteins: Expression in several secretory tissues of the mouse. Cell 1983, 32, 755–761. [Google Scholar] [CrossRef]
- Stopková, R.; Stopka, P.; Janotová, K.; Jedelský, P.L. Species-specific Expression of Major Urinary Proteins in the House Mice (Mus musculus musculus and Mus musculus domesticus). J. Chem. Ecol. 2007, 33, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Hui, X.; Zhu, W.; Wang, Y.; Lam, K.S.L.; Zhang, J.; Wu, D.; Kraegen, E.W.; Li, Y.; Xu, A. Major Urinary Protein-1 Increases Energy Expenditure and Improves Glucose Intolerance through Enhancing Mitochondrial Function in Skeletal Muscle of Diabetic Mice. J. Biol. Chem. 2009, 284, 14050–14057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Jiang, L.; Rui, L. Identification of MUP1 as a Regulator for Glucose and Lipid Metabolism in Mice. J. Biol. Chem. 2009, 284, 11152–11159. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-C.; Lee, T.-Y.; Kwok, C.-F.; Hsu, Y.-P.; Shih, K.-C.; Lin, Y.-J.; Ho, L.-T. Major urinary protein 1 interacts with cannabinoid receptor type 1 in fatty acid-induced hepatic insulin resistance in a mouse hepatocyte model. Biochem. Biophys. Res. Commun. 2015, 460, 1063–1068. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-H.; Kim, D.; Choi, I.; Bae, K. Identification of transcriptional regulatory elements required for the Mup2 expression in circadian clock mutant mice. Biochem. Biophys. Res. Commun. 2011, 410, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Schoeller, E.L.; Tonsfeldt, K.J.; Sinkovich, M.; Shi, R.; Mellon, P.L. Growth Hormone Pulses and Liver Gene Expression Are Differentially Regulated by the Circadian Clock Gene Bmal1. Endocrinology 2021, 162, bqab023. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhang, W.; Lu, S.; Lu, G.; Zhang, H.; Zhuang, Y.; Wang, Y.; Dong, M.; Zhang, Y.; Zhou, X.; et al. Mup-knockout mice generated through CRISPR/Cas9-mediated deletion for use in urinary protein analysis. Acta Biochim. Et Biophys. Sin. 2016, 48, 468–473. [Google Scholar] [CrossRef]
- Janciauskiene, S.; Wrenger, S.; Immenschuh, S.; Olejnicka, B.; Greulich, T.; Welte, T.; Chorostowska-Wynimko, J. The Multifaceted Effects of Alpha1-Antitrypsin on Neutrophil Functions. Front. Pharmacol. 2018, 9, 341. [Google Scholar] [CrossRef]
- Janciauskiene, S.M.; Bals, R.; Koczulla, R.; Vogelmeier, C.; Köhnlein, T.; Welte, T. The discovery of α1-antitrypsin and its role in health and disease. Respir. Med. 2011, 105, 1129–1139. [Google Scholar] [CrossRef]
- Fromme, M.; Schneider, C.V.; Trautwein, C.; Brunetti-Pierri, N.; Strnad, P. Alpha-1 antitrypsin deficiency: A re-surfacing adult liver disorder. J. Hepatol. 2021, 76, 946–958. [Google Scholar] [CrossRef]
- Martinez-Huenchullan, S.F.; Shipsey, I.; Hatchwell, L.; Min, D.; Twigg, S.M.; Larance, M. Blockade of High-Fat Diet Proteomic Phenotypes Using Exercise as Prevention or Treatment. Mol. Cell. Proteom. 2021, 20, 100027. [Google Scholar] [CrossRef] [PubMed]
- Grander, C.; Schaefer, B.; Schwärzler, J.; Grabherr, F.; de Graaf, D.M.; Enrich, B.; Oberhuber, G.; Mayr, L.; Sangineto, M.; Jaschke, N.; et al. Alpha-1 antitrypsin governs alcohol-related liver disease in mice and humans. Gut 2020, 70, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Kalas, M.A.; Chavez, L.; Leon, M.; Taweesedt, P.T.; Surani, S. Abnormal liver enzymes: A review for clinicians. World J. Hepatol. 2021, 13, 1688–1698. [Google Scholar] [CrossRef] [PubMed]
- Arakaki, T.L.; Pezza, J.A.; Cronin, M.A.; Hopkins, C.E.; Zimmer, D.B.; Tolan, D.R.; Allen, K.N. Structure of human brain fructose 1,6-(bis)phosphate aldolase: Linking isozyme structure with function. Protein Sci. 2004, 13, 3077–3084. [Google Scholar] [CrossRef] [PubMed]
- Nesteruk, M.; Hennig, E.; Mikula, M.; Karczmarski, J.; Dzwonek, A.; Goryca, K.; Rubel, T.; Paziewska, A.; Woszczyński, M.; Ledwon, J.; et al. Mitochondrial-related proteomic changes during obesity and fasting in mice are greater in the liver than skeletal muscles. Funct. Integr. Genom. 2014, 14, 245–259. [Google Scholar] [CrossRef]
- James, C.L.; Rellos, P.; Ali, M.; Heeley, A.F.; Cox, T.M. Neonatal screening for hereditary fructose intolerance: Frequency of the most common mutant aldolase B allele (A149P) in the British population. J. Med Genet. 1996, 33, 837–841. [Google Scholar] [CrossRef]
- Gruchota, J.; Pronicka, E.; Korniszewski, L.; Stolarski, B.; Pollak, A.; Rogaszewska, M.; Płoski, R. Aldolase B mutations and prevalence of hereditary fructose intolerance in a Polish population. Mol. Genet. Metab. 2006, 87, 376–378. [Google Scholar] [CrossRef]
- Coffee, E.M.; Yerkes, L.; Ewen, E.P.; Zee, T.; Tolan, D.R. Increased prevalence of mutant null alleles that cause hereditary fructose intolerance in the American population. J. Inherit. Metab. Dis. 2010, 33, 33–42. [Google Scholar] [CrossRef]
- Oppelt, S.A.; Sennott, E.M.; Tolan, D.R. Aldolase-B knockout in mice phenocopies hereditary fructose intolerance in humans. Mol. Genet. Metab. 2015, 114, 445–450. [Google Scholar] [CrossRef]
- Lanaspa, M.A.; Andres-Hernando, A.; Orlicky, D.J.; Cicerchi, C.; Jang, C.; Li, N.; Milagres, T.; Kuwabara, M.; Wempe, M.F.; Rabinowitz, J.D.; et al. Ketohexokinase C blockade ameliorates fructose-induced metabolic dysfunction in fructose-sensitive mice. J. Clin. Investig. 2018, 128, 2226–2238. [Google Scholar] [CrossRef]
- Murphy, S.; Yang, H.; Moylan, C.A.; Pang, H.; Dellinger, A.; Abdelmalek, M.; Garrett, M.E.; Ashley-Koch, A.; Suzuki, A.; Tillmann, H.L.; et al. Relationship Between Methylome and Transcriptome in Patients with Nonalcoholic Fatty Liver Disease. Gastroenterology 2013, 145, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.; Fernández-Ramos, D.; Varela-Rey, M.; Martínez-Arranz, I.; Navasa, N.; Van Liempd, S.M.; Lavín Trueba, J.L.; Mayo, R.; Ilisso, C.P.; de Juan, V.G.; et al. Metabolomic Identification of Subtypes of Nonalcoholic Steatohepatitis. Gastroenterology 2017, 152, 1449–1461.e7. [Google Scholar] [CrossRef] [Green Version]
- Moylan, C.A.; Pang, H.; Dellinger, A.; Suzuki, A.; Garrett, M.E.; Guy, C.D.; Murphy, S.K.; Ashley-Koch, A.E.; Choi, S.S.; Michelotti, G.; et al. Hepatic gene expression profiles differentiate presymptomatic patients with mild versus severe nonalcoholic fatty liver disease. Hepatology 2014, 59, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Turecky, L.; Kupcova, V.; Durfinova, M.; Uhlikova, E. Serum butyrylcholinesterase activities in patients with non-alcoholic fatty liver disease. Comparison with liver proteosynthetic function and liver fibrosis. Bratisl. Med J. 2021, 122, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Hardwick, R.N.; Fisher, C.D.; Canet, M.J.; Scheffer, G.L.; Cherrington, N.J. Variations in ATP-Binding Cassette Transporter Regulation during the Progression of Human Nonalcoholic Fatty Liver Disease. Drug Metab. Dispos. 2011, 39, 2395–2402. [Google Scholar] [CrossRef]
- Chen, Y.-M.; Liu, Y.; Zhou, R.-F.; Chen, X.-L.; Wang, C.; Tan, X.-Y.; Wang, L.-J.; Zheng, R.-D.; Zhang, H.-W.; Ling, W.-H.; et al. Associations of gut-flora-dependent metabolite trimethylamine-N-oxide, betaine and choline with non-alcoholic fatty liver disease in adults. Sci. Rep. 2016, 6, 19076. [Google Scholar] [CrossRef]
- Huang, T.; Yu, L.; Pan, H.; Ma, Z.; Wu, T.; Zhang, L.; Liu, K.; Qi, Q.; Miao, W.; Song, Z.; et al. Integrated Transcriptomic and Translatomic Inquiry of the Role of Betaine on Lipid Metabolic Dysregulation Induced by a High-Fat Diet. Front. Nutr. 2021, 8, 763. [Google Scholar] [CrossRef]
- Fan, C.; Hu, H.; Huang, X.; Su, D.; Huang, F.; Zhuo, Z.; Tan, L.; Xu, Y.; Wang, Q.; Hou, K.; et al. Betaine Supplementation Causes an Increase in Fatty Acid Oxidation and Carbohydrate Metabolism in Livers of Mice Fed a High-Fat Diet: A Proteomic Analysis. Foods 2022, 11, 881. [Google Scholar] [CrossRef]
- Sy, S.M.-H.; Lai, P.B.-S.; Pang, E.; Wong, N.L.-Y.; To, K.-F.; Johnson, P.J.; Wong, N. Novel identification of zyxin upregulations in the motile phenotype of hepatocellular carcinoma. Mod. Pathol. 2006, 19, 1108–1116. [Google Scholar] [CrossRef]
- Bergen, H.R., 3rd; Vasmatzis, G.; Cliby, W.A.; Johnson, K.L.; Oberg, A.L.; Muddiman, D.C. Discovery of ovarian cancer biomarkers in serum using NanoLC electrospray ionization TOF and FT-ICR mass spectrometry. Dis. Markers 2003, 19, 239–249. [Google Scholar] [CrossRef]
- El-Hattab, A.W.; Dai, H.; Almannai, M.; Wang, J.; Faqeih, E.A.; Al Asmari, A.; Saleh, M.A.M.; Elamin, M.A.O.; Alfadhel, M.; Alkuraya, F.S.; et al. Molecular and clinical spectra of FBXL4 deficiency. Hum. Mutat. 2017, 38, 1649–1659. [Google Scholar] [PubMed]
- Shinomiya, H. Plastin Family of Actin-Bundling Proteins: Its Functions in Leukocytes, Neurons, Intestines, and Cancer. Int. J. Cell Biol. 2012, 2012, 213492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Wang, Z.; Liu, Y.; Huo, Y.; Liu, H.; Xu, C.; Mao, R.; Zhu, Y.; Liu, L.; Wei, D.; et al. Plastin 1 drives metastasis of colorectal cancer through the IQGAP1/Rac1/ERK pathway. Cancer Sci. 2020, 111, 2861–2871. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Kohan, A.B.; Lo, C.-M.; Liu, M.; Howles, P.; Tso, P. Apolipoprotein A-IV: A protein intimately involved in metabolism. J. Lipid Res. 2015, 56, 1403–1418. [Google Scholar] [CrossRef]
- Kang, M.; Kim, J.; An, H.; Ko, J. Human leucine zipper protein promotes hepatic steatosis via induction of apolipoprotein A-IV. FASEB J. 2017, 31, 2548–2561. [Google Scholar] [CrossRef]
- Wang, P.-W.; Hung, Y.-C.; Wu, T.-H.; Chen, M.-H.; Yeh, C.-T.; Pan, T.-L. Proteome-based identification of apolipoprotein A-IV as an early diagnostic biomarker in liver fibrosis. Oncotarget 2017, 8, 88951–88964. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic anhydrase inhibitors and activators for novel therapeutic applications. Futur. Med. Chem. 2011, 3, 1165–1180. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic anhydrases—An overview. Curr. Pharm. Des. 2008, 14, 603–614. [Google Scholar] [CrossRef]
- Yuan, L.; Wang, M.; Liu, T.; Lei, Y.; Miao, Q.; Li, Q.; Wang, H.; Zhang, G.; Hou, Y.; Chang, X. Carbonic Anhydrase 1-Mediated Calcification Is Associated With Atherosclerosis, and Methazolamide Alleviates Its Pathogenesis. Front. Pharmacol. 2019, 10, 766. [Google Scholar] [CrossRef]
- Arakawa, T.; Kobayashi-Yurugi, T.; Alguel, Y.; Iwanari, H.; Hatae, H.; Iwata, M.; Abe, Y.; Hino, T.; Ikeda-Suno, C.; Kuma, H.; et al. Crystal structure of the anion exchanger domain of human erythrocyte band 3. Science 2015, 350, 680–684. [Google Scholar] [CrossRef]
- Romero, M.F.; Chen, A.P.; Parker, M.D.; Boron, W.F. The SLC4 family of bicarbonate (HCO3−) transporters. Mol. Asp. Med. 2013, 34, 159–182. [Google Scholar] [CrossRef] [PubMed]
- Remigante, A.; Spinelli, S.; Pusch, M.; Sarikas, A.; Morabito, R.; Marino, A.; Dossena, S. Role of SLC4 and SLC26 solute carriers during oxidative stress. Acta Physiol. 2022, 235, e13796. [Google Scholar] [CrossRef]
- Bhushan, B.; Michalopoulos, G.K. Role of epidermal growth factor receptor in liver injury and lipid metabolism: Emerging new roles for an old receptor. Chem. Interact. 2020, 324, 109090. [Google Scholar] [CrossRef] [PubMed]
- Collin de l’Hortet, A.; Zerrad-Saadi, A.; Prip-Buus, C.; Fauveau, V.; Helmy, N.; Ziol, M.; Vons, C.; Billot, K.; Baud, V.; Gilgenkrantz, H.; et al. GH administration rescues fatty liver regeneration impairment by restoring GH/EGFR pathway deficiency. Endocrinology 2014, 155, 2545–2554. [Google Scholar] [CrossRef] [PubMed]
- Giraudi, P.J.; Giuricin, M.; Bonazza, D.; de Manzini, N.; Tiribelli, C.; Palmisano, S.; Rosso, N. Modifications of IGF2 and EGFR plasma protein concentrations in NAFLD patients after bariatric surgery. Int. J. Obes. 2021, 45, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Giraldez, M.D.; Carneros, D.; Garbers, C.; Rose-John, S.; Bustos, M. New insights into IL-6 family cytokines in metabolism, hepatology and gastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 787–803. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, J.; Wang, J.; Hu, W.; Feng, Z. The emerging role of leukemia inhibitory factor in cancer and therapy. Pharmacol. Ther. 2021, 221, 107754. [Google Scholar] [CrossRef]
- Yuan, Y.; Li, K.; Teng, F.; Wang, W.; Zhou, B.; Zhou, X.; Lin, J.; Ye, X.; Deng, Y.; Liu, W.; et al. Leukemia inhibitory factor protects against liver steatosis in nonalcoholic fatty liver disease patients and obese mice. J. Biol. Chem. 2022, 298, 101946. [Google Scholar] [CrossRef]
- Meng, Q.; Li, X.; Xiong, X. Identification of Hub Genes Associated with Non-alcoholic Steatohepatitis Using Integrated Bioinformatics Analysis. Front. Genet. 2022, 13, 872518. [Google Scholar] [CrossRef]
- Stanley, T.L.; Fourman, L.T.; Zheng, I.; McClure, C.M.; Feldpausch, M.N.; Torriani, M.; Corey, K.E.; Chung, R.T.; Lee, H.; Kleiner, D.E.; et al. Relationship of IGF-1 and IGF-Binding Proteins to Disease Severity and Glycemia in Nonalcoholic Fatty Liver Disease. J. Clin. Endocrinol. Metab. 2021, 106, e520–e533. [Google Scholar] [CrossRef]
- Fahlbusch, P.; Knebel, B.; Hörbelt, T.; Barbosa, D.M.; Nikolic, A.; Jacob, S.; Al-Hasani, H.; Van De Velde, F.; Van Nieuwenhove, Y.; Müller-Wieland, D.; et al. Physiological Disturbance in Fatty Liver Energy Metabolism Converges on IGFBP2 Abundance and Regulation in Mice and Men. Int. J. Mol. Sci. 2020, 21, 4144. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhou, W.; Wu, Y.; Xu, L.; Wang, Y.; Xu, Z.; Yang, Y. Circulating IGFBP-2 levels are inversely associated with the incidence of nonalcoholic fatty liver disease: A cohort study. J. Int. Med. Res. 2020, 48, 0300060520935219. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Janech, M.G.; Sobolesky, P.M.; Bland, A.M.; Samsuddin, S.; Alazawi, W.; Syn, W.-K. Proteomic screening of plasma identifies potential noninvasive biomarkers associated with significant/advanced fibrosis in patients with nonalcoholic fatty liver disease. Biosci. Rep. 2020, 40, BSR20190395. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Wang, M.; Yin, C.; Li, S.; Liu, Y.; Xiao, Y. Serum complement factor 5a levels are associated with nonalcoholic fatty liver disease in obese children. Acta Paediatr. 2018, 107, 322–327. [Google Scholar] [CrossRef]
- Hillebrandt, S.; Wasmuth, H.E.; Weiskirchen, R.; Hellerbrand, C.; Keppeler, H.; Werth, A.; Schirin-Sokhan, R.; Wilkens, G.; Geier, A.; Lorenzen, J.; et al. Complement factor 5 is a quantitative trait gene that modifies liver fibrogenesis in mice and humans. Nat. Genet. 2005, 37, 835–843. [Google Scholar] [CrossRef]
- Viiklepp, K.; Nissinen, L.; Ojalill, M.; Riihilä, P.; Kallajoki, M.; Meri, S.; Heino, J.; Kähäri, V.-M. C1r Upregulates Production of Matrix Metalloproteinase-13 and Promotes Invasion of Cutaneous Squamous Cell Carcinoma. J. Investig. Dermatol. 2022, 142, 1478–1488.e9. [Google Scholar] [CrossRef]
- Fukuda, S.; Sumii, M.; Masuda, Y.; Takahashi, M.; Koike, N.; Teishima, J.; Yasumoto, H.; Itamoto, T.; Asahara, T.; Dohi, K.; et al. Murine and human SDF2L1 is an endoplasmic reticulum stress-inducible gene and encodes a new member of the Pmt/rt protein family. Biochem. Biophys. Res. Commun. 2001, 280, 407–414. [Google Scholar] [CrossRef]
- Nonogaki, K.; Kaji, T. Whey protein isolate inhibits hepatic FGF21 production, which precedes weight gain, hyperinsulinemia and hyperglycemia in mice fed a high-fat diet. Sci. Rep. 2020, 10, 15784. [Google Scholar] [CrossRef]
- Meunier, L.; Usherwood, Y.-K.; Chung, K.T.; Hendershot, L.M. A Subset of Chaperones and Folding Enzymes Form Multiprotein Complexes in Endoplasmic Reticulum to Bind Nascent Proteins. Mol. Biol. Cell 2002, 13, 4456–4469. [Google Scholar] [CrossRef]
- Bies, C.; Blum, R.; Dudek, J.; Nastainczyk, W.; Oberhauser, S.; Jung, M.; Zimmermann, R. Characterization of pancreatic ERj3p, a homolog of yeast DnaJ-like protein Scj1p. Biol. Chem. 2004, 385, 389–395. [Google Scholar] [CrossRef]
- Tongaonkar, P.; Selsted, M.E. SDF2L1, a Component of the Endoplasmic Reticulum Chaperone Complex, Differentially Interacts with α-, β-, and θ-Defensin Propeptides. J. Biol. Chem. 2009, 284, 5602–5609. [Google Scholar] [CrossRef] [PubMed]
- Walter, P.; Ron, D. The Unfolded Protein Response: From Stress Pathway to Homeostatic Regulation. Science 2011, 334, 1081–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasako, T.; Ohsugi, M.; Kubota, N.; Itoh, S.; Okazaki, Y.; Terai, A.; Kubota, T.; Yamashita, S.; Nakatsukasa, K.; Kamura, T.; et al. Hepatic Sdf2l1 controls feeding-induced ER stress and regulates metabolism. Nat. Commun. 2019, 10, 947. [Google Scholar] [CrossRef] [PubMed]
- Schott, A.; Ravaud, S.; Keller, S.; Radzimanowski, J.; Viotti, C.; Hillmer, S.; Sinning, I.; Strahl, S. Arabidopsis Stromal-derived Factor2 (SDF2) Is a Crucial Target of the Unfolded Protein Response in the Endoplasmic Reticulum. J. Biol. Chem. 2010, 285, 18113–18121. [Google Scholar] [CrossRef]
- Loveland, J.L.; Lank, D.B.; Küpper, C. Gene Expression Modification by an Autosomal Inversion Associated with Three Male Mating Morphs. Front. Genet. 2021, 12, 794. [Google Scholar] [CrossRef]
- Huang, X.F.; Luu-The, V. Molecular characterization of a first human 3(α→β)-hydroxysteroid epimerase. J. Biol. Chem. 2000, 275, 29452–29457. [Google Scholar] [CrossRef]
- Huang, X.F.; Luu-The, V. Gene structure, chromosomal localization and analysis of 3-ketosteroid reductase activity of the human 3(α→β)-hydroxysteroid epimerase. Biochim. Biophys. Acta 2001, 1520, 124–130. [Google Scholar] [CrossRef]
- Chan, Y.X.; Yeap, B.B. Dihydrotestosterone and cancer risk. Curr. Opin. Endocrinol. Diabetes Obes. 2018, 25, 209–217. [Google Scholar] [CrossRef]
- Kohl, M.; Wiese, S.; Warscheid, B. Cytoscape: Software for Visualization and Analysis of Biological Networks. Comput. Aided Tissue Eng. 2011, 696, 291–303. [Google Scholar]
- Bader, G.D.; Hogue, C.W.V. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinform. 2003, 4, 2–27. [Google Scholar] [CrossRef]
- Zhou, Z.; Li, Y.; Hao, H.; Wang, Y.; Zhou, Z.; Wang, Z.; Chu, X. Screening Hub Genes as Prognostic Biomarkers of Hepatocellular Carcinoma by Bioinformatics Analysis. Cell Transpl. 2019, 28 (Suppl. S1), 76s–86s. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Zhang, K.; Long, R.; Sun, Y.; Kang, J.; Zhang, T.; Cao, S. iTRAQ-based comparative proteomic analysis reveals tissue-specific and novel early-stage molecular mechanisms of salt stress response in Carex rigescens. Environ. Exp. Bot. 2017, 143, 99–114. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MCODE Cluster-ID | Gene Symbol | MCODE Score | Biological Functions of These Genes |
|---|---|---|---|
| Cluster 1 | Psmb3 | 7.50 | 20S proteasome; Proteasome; Cross-presentation of soluble exogenous antigens (endosomes) |
| Psmb2 | |||
| Psma7 | |||
| Psma6 | |||
| Psma5 | |||
| Psma4 | |||
| Psma1 | |||
| Psmb7 | |||
| Psmb6 | |||
| Psmb5 | |||
| Psmb4 | |||
| Psmb10 | |||
| Psmb1 | |||
| Psma3 | |||
| Psma2 | |||
| Psmb8 | |||
| Cluster 2 | Vgf | 5.06 | Complement and coagulation cascades; Regulation of insulin-like growth factor (IGF) transport and uptake by insulin-like growth factor binding proteins (IGFBPs); Post-translational protein phosphorylation |
| Fgb | |||
| Fermt3 | |||
| Fgg | |||
| Serpina1e | |||
| Serpina1a | |||
| Serpinf2 | |||
| Orm3 | |||
| Itih2 | |||
| Igfbp3 | |||
| Igf1 | |||
| Cp | |||
| C4b | |||
| Serping1 | |||
| Apoe | |||
| Cfd | |||
| Cluster 3 | Agxt2 | 1.17 | Small molecule catabolic process; Neutrophil degranulation; Monocarboxylic acid metabolic process |
| Vnn1 | |||
| Sord | |||
| Lamp2 | |||
| Lamp1 | |||
| Fah | |||
| Cluster 4 | Rap1b | 2.00 | Integrin-mediated cell adhesion; Focal adhesion; Rap1 signaling pathway; Cell adhesion mediated by integrin |
| Itgb3 | |||
| Itgb2 | |||
| Itgam | |||
| Itga2b | |||
| Cluster 5 | C8a | 2.00 | Terminal pathway of complement; Complement activation, alternative pathway cytolysis |
| C8b | |||
| C8g | |||
| Hc | |||
| C9 | |||
| Cluster 6 | Apom | 1.50 | Plasma lipoprotein particle remodeling; Protein–lipid complex remodeling; Protein-containing complex remodeling |
| Pltp | |||
| Apoh | |||
| Apoa4 | |||
| Cluster 7 | Krt12 | 1.50 | Epidermis development; Formation of the cornified envelope; Keratinization |
| Krt76 | |||
| Krt2 | |||
| Krt10 | |||
| Cluster 8 | Aldob | 1.50 | Carbon metabolism; Hexose metabolic process; Monosaccharide metabolic process |
| H6pd | |||
| Pgam2 | |||
| Eno1 | |||
| Cluster 9 | Dstn | 1.00 | Actin cytoskeleton organization; Actin-filament-based process |
| Pfn1 | |||
| Cap1 |
| Protein Accession | Protein Description | Gene Name | Proteomics (Fold Change) | PRM Validation (Fold Change) | ||
|---|---|---|---|---|---|---|
| srm_db/srm_bks Ratio | srm_db/srm_bks p Value | srm_db/srm_bks Ratio | srm_db/srm bks p Value | |||
| Q9R1P0 | Proteasome subunit alpha type-4 OS = Mus musculus OX = 10,090 GN = Psma4 PE = 1 SV = 1 | Psma4 | 1.78 | *** | 2.82 | * |
| O09061 | Proteasome subunit beta type-1 OS = Mus musculus OX = 10,090 GN = Psmb1 PE = 1 SV = 1 | Psmb1 | 1.86 | *** | 2.83 | ** |
| Q9R1P4 | Proteasome subunit alpha type-1 OS = Mus musculus OX = 10,090 GN = Psma1 PE = 1 SV = 1 | Psma1 | 1.95 | *** | 2.38 | * |
| Q9Z2U0 | Proteasome subunit alpha type-7 OS = Mus musculus OX = 10,090 GN = Psma7 PE = 1 SV = 1 | Psma7 | 2.04 | *** | 2.29 | * |
| Q91Y97 | Fructose-bisphosphate aldolase B OS = Mus musculus OX = 10,090 GN = Aldob PE = 1 SV = 3 | Aldob | 2.93 | *** | 8.08 | * |
| Q00898 | Alpha-1-antitrypsin 1–5 OS = Mus musculus OX = 10,090 GN = Serpina1e PE = 1 SV = 1 | Serpina1e | 0.14 | *** | 0.01 | ** |
| Q60692 | Proteasome subunit beta type-6 OS = Mus musculus OX = 10,090 GN = Psmb6 PE = 1 SV = 3 | Psmb6 | 2.01 | *** | 2.99 | *** |
| Q9QUM9 | Proteasome subunit alpha type-6 OS = Mus musculus OX = 10,090 GN = Psma6 PE = 1 SV = 1 | Psma6 | 2.03 | *** | 3.04 | ** |
| Q9R1P1 | Proteasome subunit beta type-3 OS = Mus musculus OX = 10,090 GN = Psmb3 PE = 1 SV = 1 | Psmb3 | 2.12 | *** | 3.43 | * |
| P07758 | Alpha-1-antitrypsin 1–1 OS = Mus musculus OX = 10,090 GN = Serpina1a PE = 1 SV = 4 | Serpina1a | 0.47 | * | 0.46 | * |
| MCODE Cluster-ID | Gene Symbol | MCODE Score | Biological Functions of These Genes |
|---|---|---|---|
| Cluster 1 | Hspa13 | 4.33 | Protein folding; ER-localized multiprotein complex, in absence of Ig heavy chains ER-localized multiprotein complex, Ig heavy chain-associated; |
| Eif4b | |||
| Dnajc25 | |||
| Rps27l | |||
| Nudt7 | |||
| Nop56 | |||
| Rpl11 | |||
| Rsl1d1 | |||
| Sdf2l1 | |||
| Dnaja4 | |||
| Rps28 | |||
| Sec61a1 | |||
| Pnrc2 | |||
| Rps7 | |||
| Rps3a1 | |||
| Rbm3 | |||
| Ppib | |||
| Sqstm1 | |||
| Hsp90aa1 | |||
| Hspa1b | |||
| Hsph1 | |||
| Hspa5 | |||
| Pdia3 | |||
| Fbl | |||
| Pdia4 | |||
| Hyou1 | |||
| Cluster 2 | Cyp2c23 | 6.62 | Retinol metabolism; Chemical carcinogenesis—DNA adducts; Xenobiotic metabolic process |
| Cyp2c70 | |||
| Gstt3 | |||
| Gstk1 | |||
| Ugt2a3 | |||
| Cyp2u1 | |||
| Mgst3 | |||
| Erg28 | |||
| Cyp3a25 | |||
| Aldh1a7 | |||
| Hsd3b5 | |||
| Gstp1 | |||
| Gsta2 | |||
| Gsta1 | |||
| Cyp4a14 | |||
| Cyp4a10 | |||
| Cyp2b9 | |||
| Cyp2b10 | |||
| Cyp26a1 | |||
| Cyp1a2 | |||
| Cluster 3 | Dnajc3 | 4.50 | Post-translational protein phosphorylation; Regulation of insulin-like growth factor (IGF) transport and uptake by insulin-like growth factor binding proteins (IGFBPs); Plasma lipoprotein remodeling |
| Pdia6 | |||
| Apoa5 | |||
| Hsp90b1 | |||
| Serpina1e | |||
| Serpina1b | |||
| Calu | |||
| Apoa2 | |||
| Alb | |||
| Cluster 4 | Aldob | 1.90 | Glycolysis/gluconeogenesis; Glucose metabolism; Gluconeogenesis |
| H1f5 | |||
| Maoa | |||
| H2ax | |||
| H1f3 | |||
| Fbp1 | |||
| Eno3 | |||
| Bpgm | |||
| Aldoc | |||
| Cluster 5 | Ttll3 | 1.25 | GnRH secretion; Bacterial invasion of epithelial cells; RHO GTPase effectors |
| Arpc1a | |||
| Trpc4 | |||
| Prkca | |||
| Itpr2 | |||
| Cdh1 | |||
| Casp3 | |||
| Arpc1b | |||
| Cluster 6 | Abcb8 | 1.43 | Supramolecular fiber organization; Actin cytoskeleton organization; Actin filament-based process |
| Crip2 | |||
| Lima1 | |||
| Abcb10 | |||
| Ppl | |||
| Gas2 | |||
| Cluster 7 | Acss3 | 2.50 | Valine, leucine and isoleucine degradation; Propanoate metabolism; Carboxylic acid catabolic process |
| Abat | |||
| Aldh1b1 | |||
| Hadh | |||
| Acadm | |||
| Cluster 8 | Txndc5 | 2.00 | Neutrophil degranulation |
| Dpp7 | |||
| Orm2 | |||
| Fabp5 | |||
| Cluster 9 | Spcs2 | 1.80 | Signal peptide processing; Synthesis, secretion, and deacylation of ghrelin; Protein export |
| Sec11c | |||
| Sec11a | |||
| Alpl | |||
| Cluster 10 | Rpn1 | 1.50 | Protein N-linked glycosylation; Macromolecule glycosylation; Protein glycosylation |
| Ostc | |||
| Stt3a | |||
| Cluster 11 | Acot4 | 1.50 | Peroxisomal protein import; Protein localization; Fatty acid metabolism |
| Acot3 | |||
| Acot1 | |||
| Cluster 12 | Akr1d1 | 1.50 | Steroid hormone biosynthesis; Androgen metabolic process; Steroid catabolic process |
| Hsd17b6 | |||
| Hsd17b2 | |||
| Cluster 13 | Necap1 | 1.50 | Clathrin-mediated endocytosis; Cargo recognition for clathrin-mediated endocytosis; Membrane Trafficking |
| Egfr | |||
| Cttn | |||
| Cluster 14 | C8b | 1.50 | Terminal pathway of complement Complement activation, alternative pathway; Cytolysis |
| C8g | |||
| Hc |
| Protein Accession | Protein Description | Gene Name | Proteomics (Fold Change) | PRM Validation (Fold Change) | ||
|---|---|---|---|---|---|---|
| lvr_db/lvr_bks Ratio | lvr_db/lvr_bks p Value | lvr_db/lvr_bks Ratio | lvr_db/lvr_bks p Value | |||
| Q00898 | Alpha-1-antitrypsin 1–5 OS = Mus musculus OX = 10,090 GN = Serpina1e PE = 1 SV = 1 | Serpina1e | 0.19 | *** | 0.10 | *** |
| Q9ESP1 | Stromal cell-derived factor 2-like protein 1 OS = Mus musculus OX = 10,090 GN = Sdf2l1 PE = 1 SV = 2 | Sdf2l1 | 0.47 | *** | 0.34 | ** |
| P58044 | Isopentenyl-diphosphate Delta-isomerase 1 OS = Mus musculus OX = 10,090 GN = Idi1 PE = 1 SV = 1 | Idi1 | 0.43 | *** | 0.25 | ** |
| Q61694 | NADPH-dependent 3-keto-steroid reductase Hsd3b5 OS = Mus musculus OX = 10,090 GN = Hsd3b5 PE = 1 SV = 4 | Hsd3b5 | 0.17 | *** | 0.01 | * |
| Q9R092 | 17-beta-hydroxysteroid dehydrogenase type 6 OS = Mus musculus OX = 10,090 GN = Hsd17b6 PE = 1 SV = 1 | Hsd17b6 | 0.41 | *** | 0.19 | *** |
| P51658 | Estradiol 17-beta-dehydrogenase 2 OS = Mus musculus OX = 10,090 GN = Hsd17b2 PE = 1 SV = 2 | Hsd17b2 | 0.41 | *** | 0.18 | *** |
| Q99L20 | Glutathione S-transferase theta-3 OS = Mus musculus OX = 10,090 GN = Gstt3 PE = 1 SV = 1 | Gstt3 | 2.26 | *** | 6.01 | *** |
| Q920E5 | Farnesyl pyrophosphate synthase OS = Mus musculus OX = 10,090 GN = Fdps PE = 1 SV = 1 | Fdps | 0.48 | * | 0.27 | *** |
| Q91W64 | Cytochrome P450 2C70 OS = Mus musculus OX = 10,090 GN = Cyp2c70 PE = 1 SV = 2 | Cyp2c70 | 0.34 | *** | 0.10 | *** |
| P12790 | Cytochrome P450 2B9 OS = Mus musculus OX = 10,090 GN = Cyp2b9 PE = 1 SV = 2 | Cyp2b9 | 3.53 | *** | 33.56 | *** |
| Q60598 | Src substrate cortactin OS = Mus musculus OX = 10,090 GN = Cttn PE = 1 SV = 2 | Cttn | 1.54 | ** | 2.04 | *** |
| Q14DH7 | Acyl-CoA synthetase short-chain family member 3, mitochondrial OS = Mus musculus OX = 10,090 GN = Acss3 PE = 1 SV = 2 | Acss3 | 2.79 | *** | 6.49 | *** |
| Q8BWN8 | Peroxisomal succinyl-coenzyme A thioesterase OS = Mus musculus OX = 10,090 GN = Acot4 PE = 1 SV = 1 | Acot4 | 1.79 | *** | 2.81 | *** |
| Q9QYR7 | Acyl-coenzyme A thioesterase 3 OS = Mus musculus OX = 10,090 GN = Acot3 PE = 1 SV = 1 | Acot3 | 2.70 | *** | 6.99 | *** |
| O55137 | Acyl-coenzyme A thioesterase 1 OS = Mus musculus OX = 10,090 GN = Acot1 PE = 1 SV = 1 | Acot1 | 2.42 | *** | 4.19 | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Wu, X.; Xu, M.; Yue, T.; Ling, P.; Fang, T.; Luo, S.; Xu, S.; Weng, J. Comparative Proteomic Analysis of Liver Tissues and Serum in db/db Mice. Int. J. Mol. Sci. 2022, 23, 9687. https://doi.org/10.3390/ijms23179687
Zhang Y, Wu X, Xu M, Yue T, Ling P, Fang T, Luo S, Xu S, Weng J. Comparative Proteomic Analysis of Liver Tissues and Serum in db/db Mice. International Journal of Molecular Sciences. 2022; 23(17):9687. https://doi.org/10.3390/ijms23179687
Chicago/Turabian StyleZhang, Yu, Xiumei Wu, Mengyun Xu, Tong Yue, Ping Ling, Tingyu Fang, Sihui Luo, Suowen Xu, and Jianping Weng. 2022. "Comparative Proteomic Analysis of Liver Tissues and Serum in db/db Mice" International Journal of Molecular Sciences 23, no. 17: 9687. https://doi.org/10.3390/ijms23179687
APA StyleZhang, Y., Wu, X., Xu, M., Yue, T., Ling, P., Fang, T., Luo, S., Xu, S., & Weng, J. (2022). Comparative Proteomic Analysis of Liver Tissues and Serum in db/db Mice. International Journal of Molecular Sciences, 23(17), 9687. https://doi.org/10.3390/ijms23179687