
Genome-Wide Analysis of the SPL Gene Family and Expression Analysis during Flowering Induction in Prunus × yedoensis ‘Somei-yoshino’
Abstract
:1. Introduction
2. Results
2.1. Screening, Identification and Sequence Analysis of PySPLs
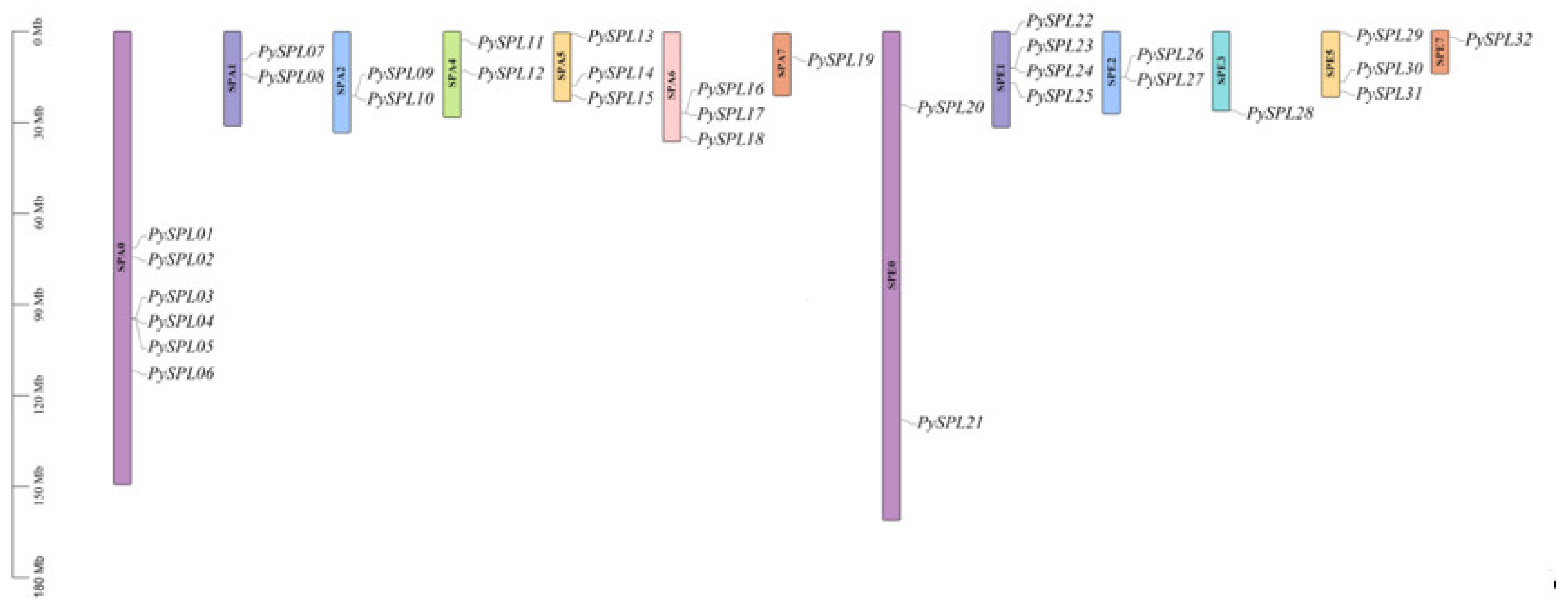
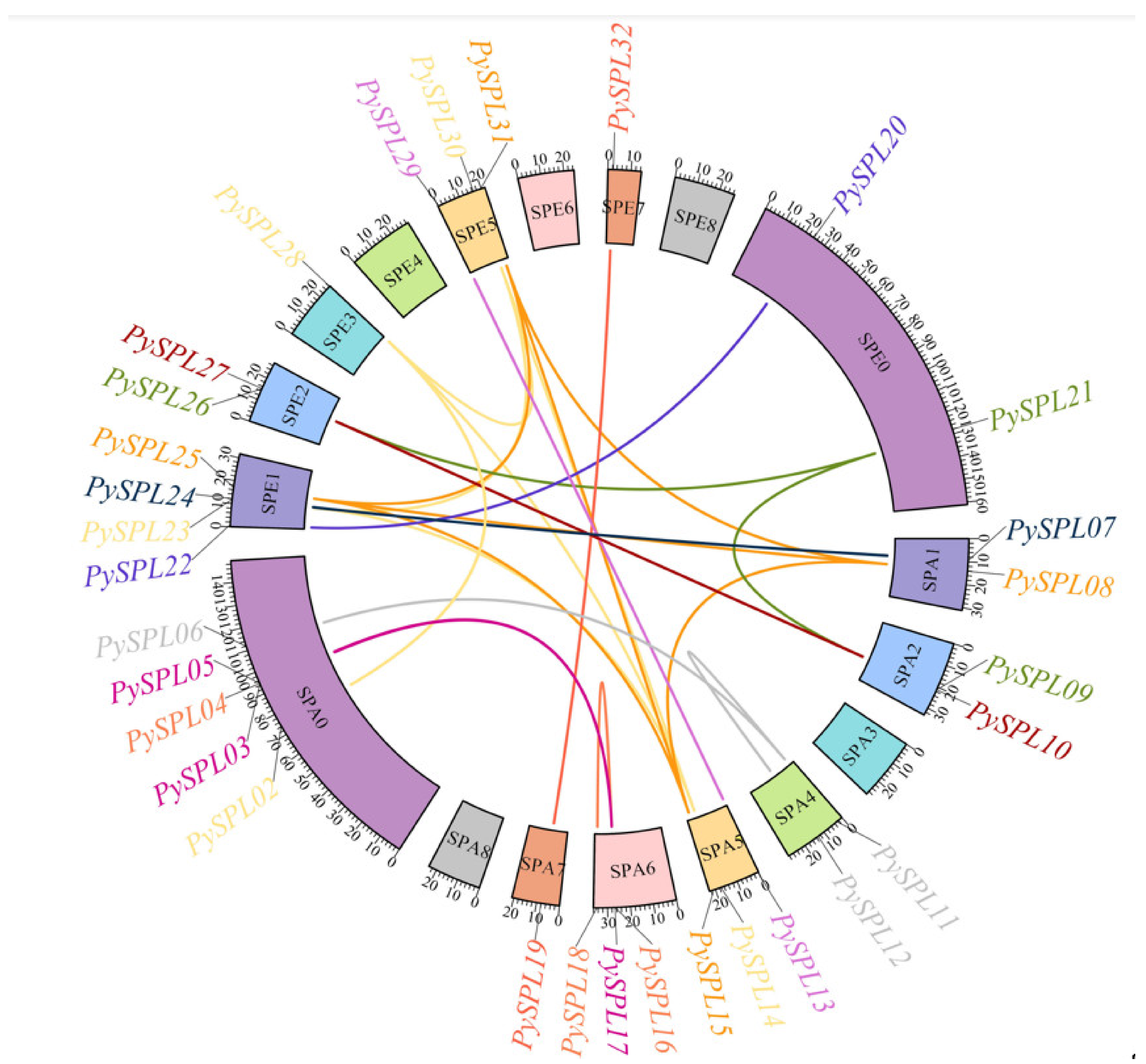
2.2. Chromosomal Localization of PySPLs
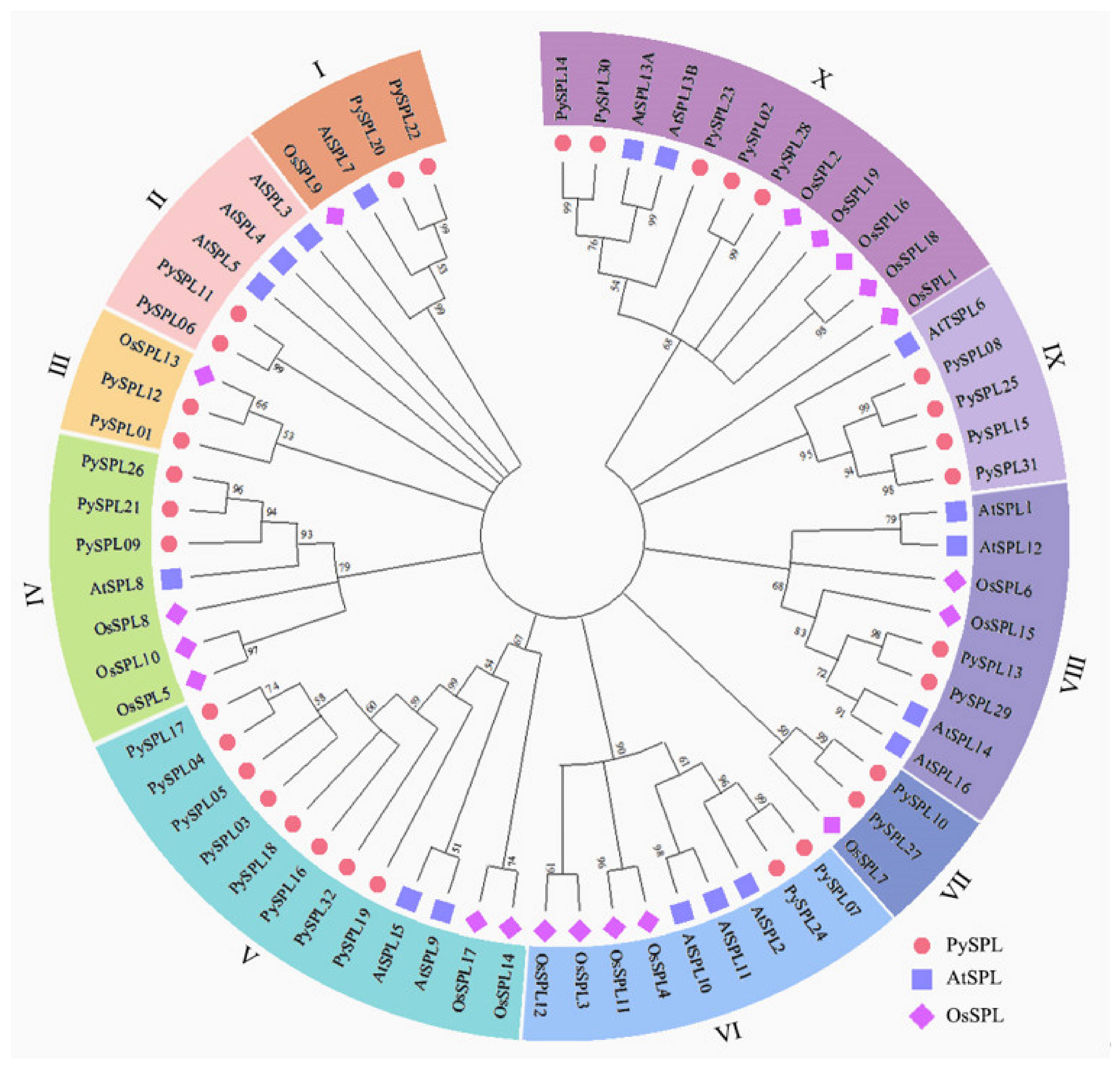
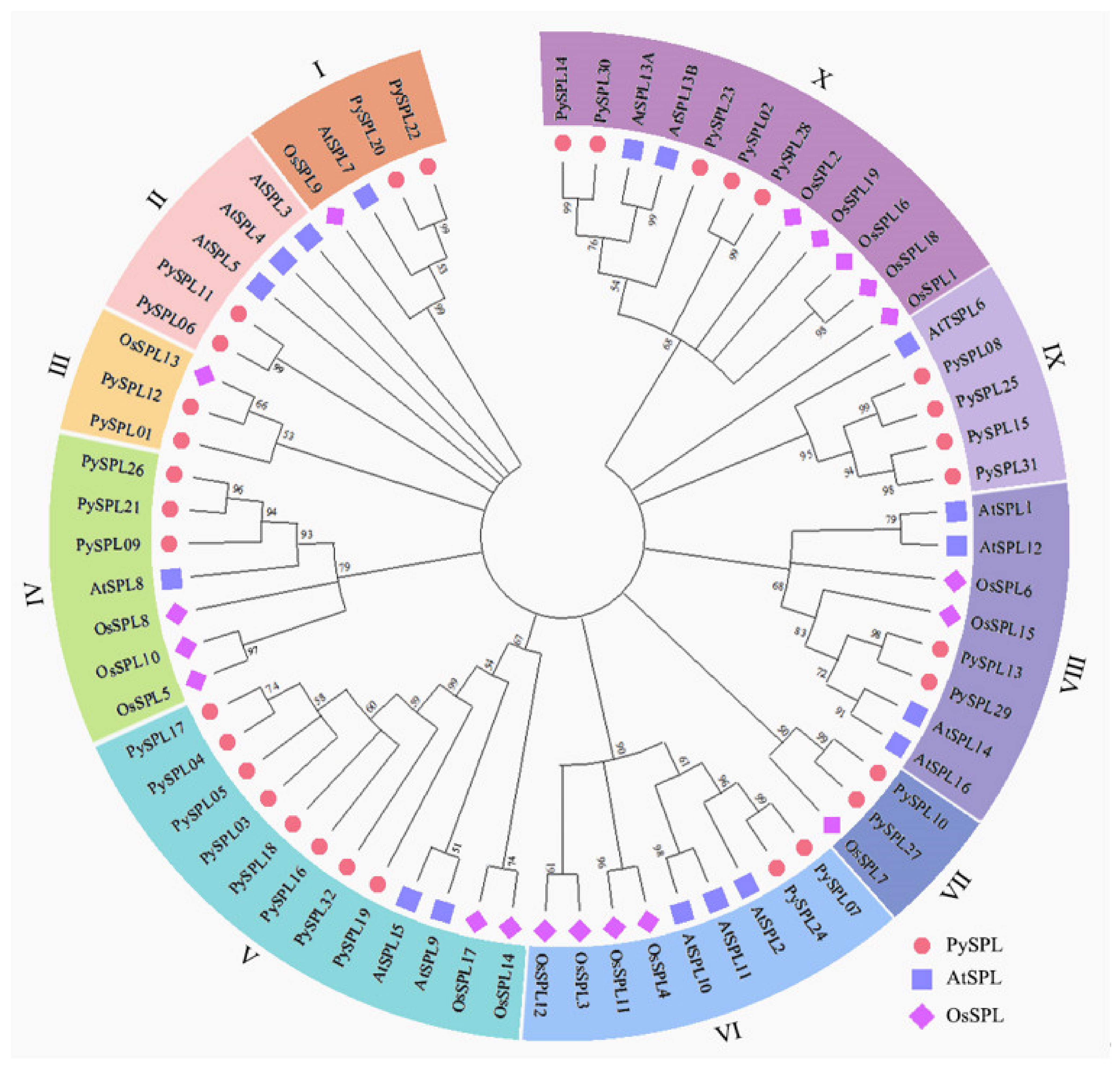
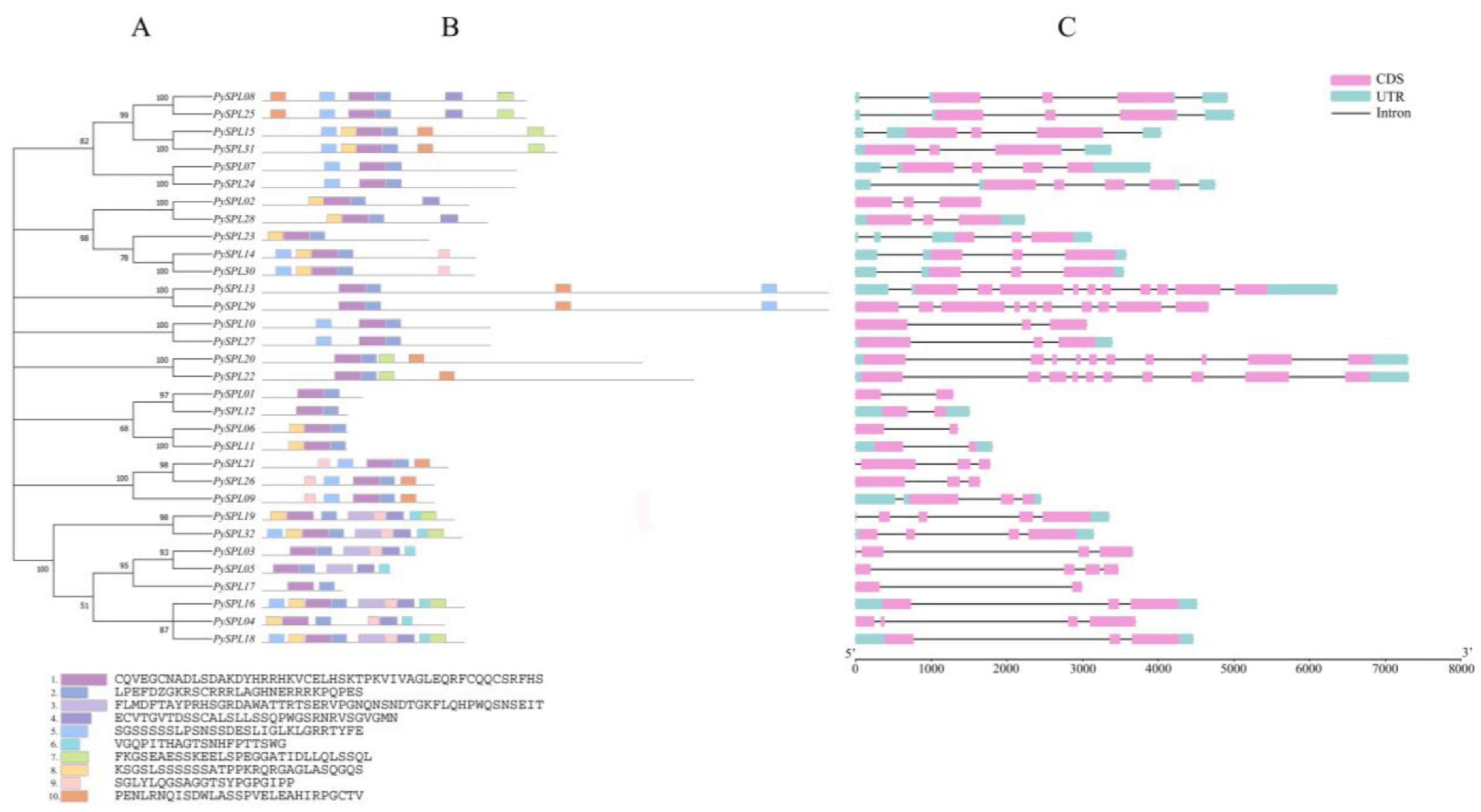
2.3. Phylogeny Analysis of PySPLs
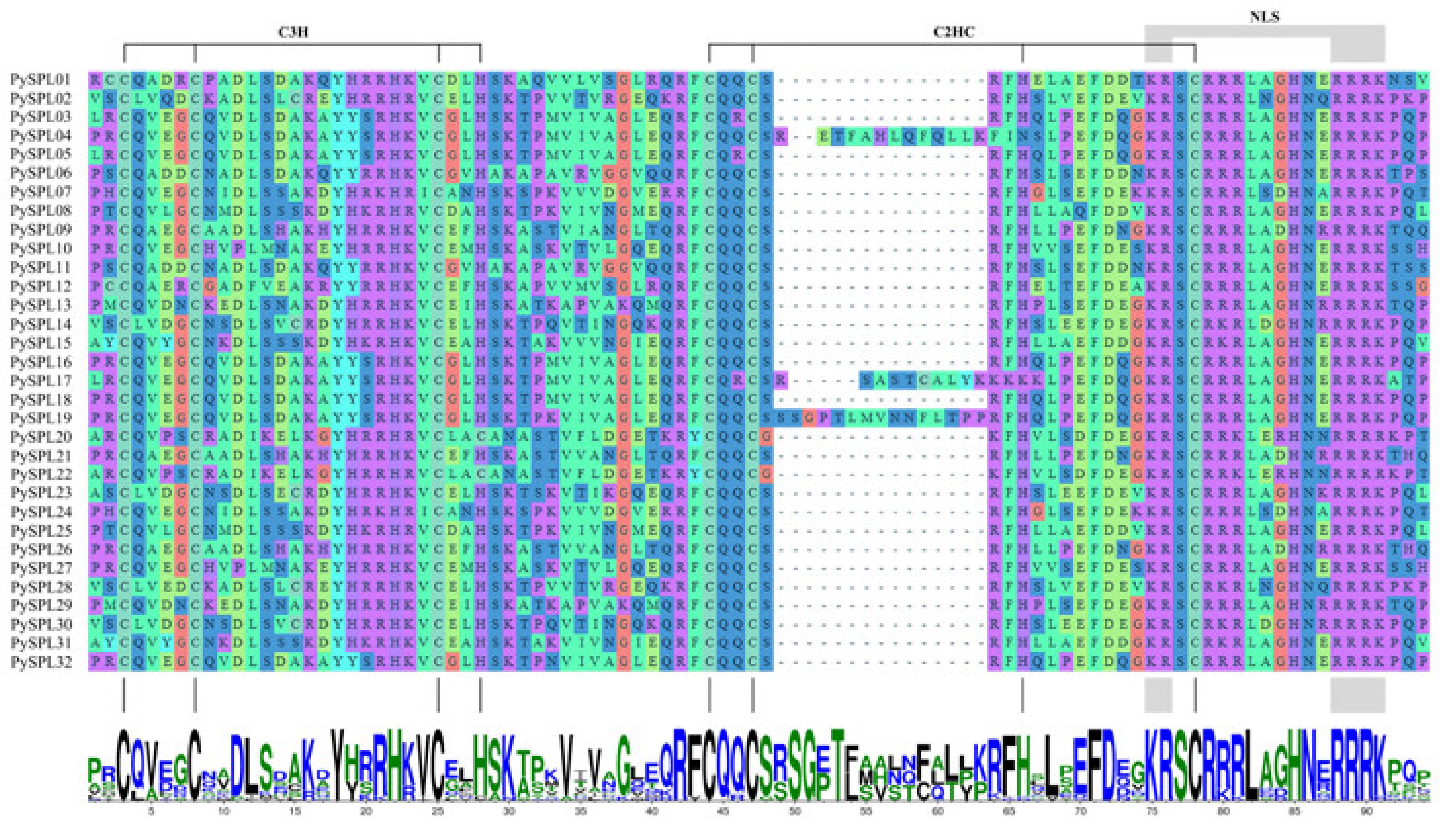
2.4. Multiple Sequence Alignment and Conserved Domain Visualization of PySPLs
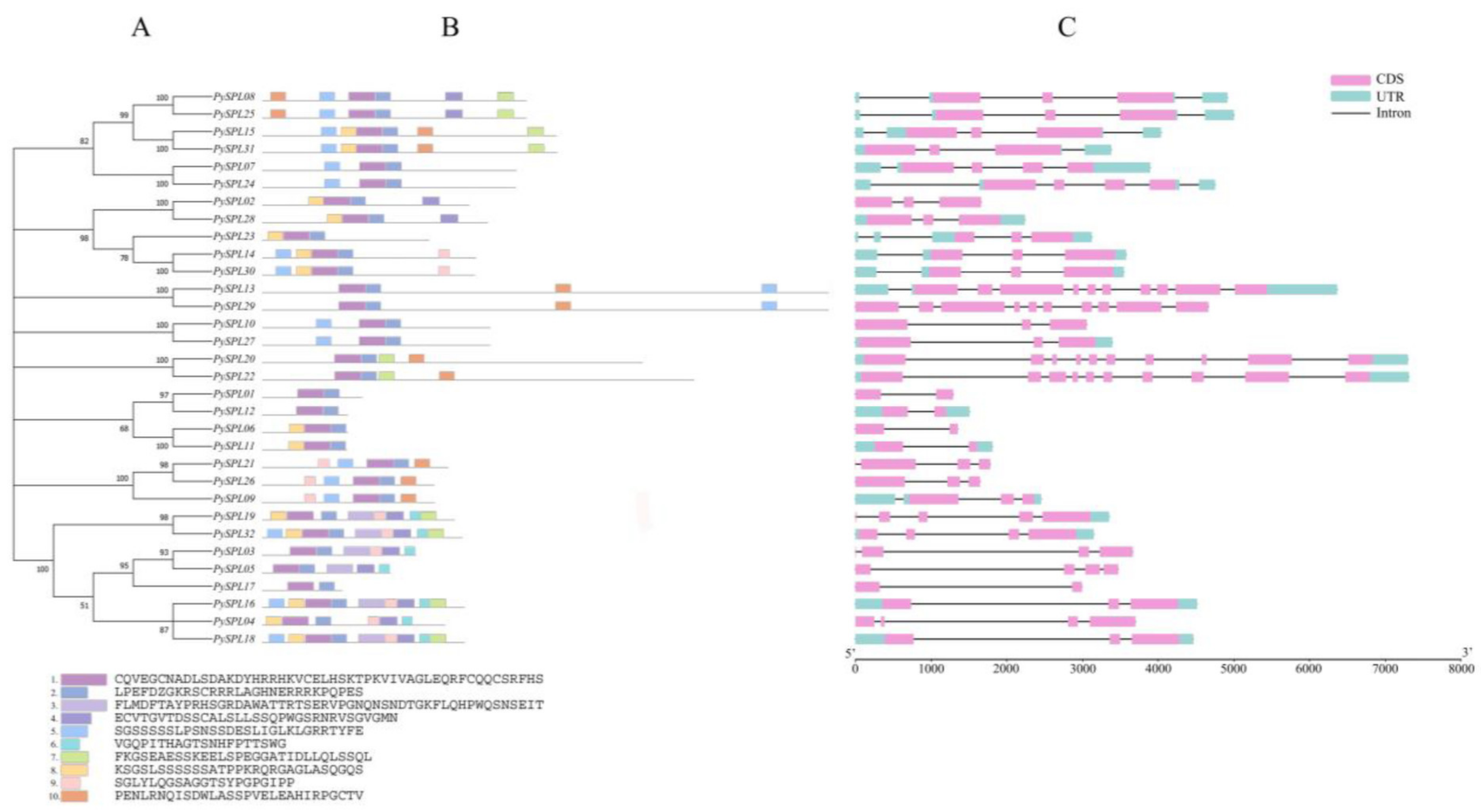
2.5. Conserved Motifs and Gene Structure Analysis of PySPLs
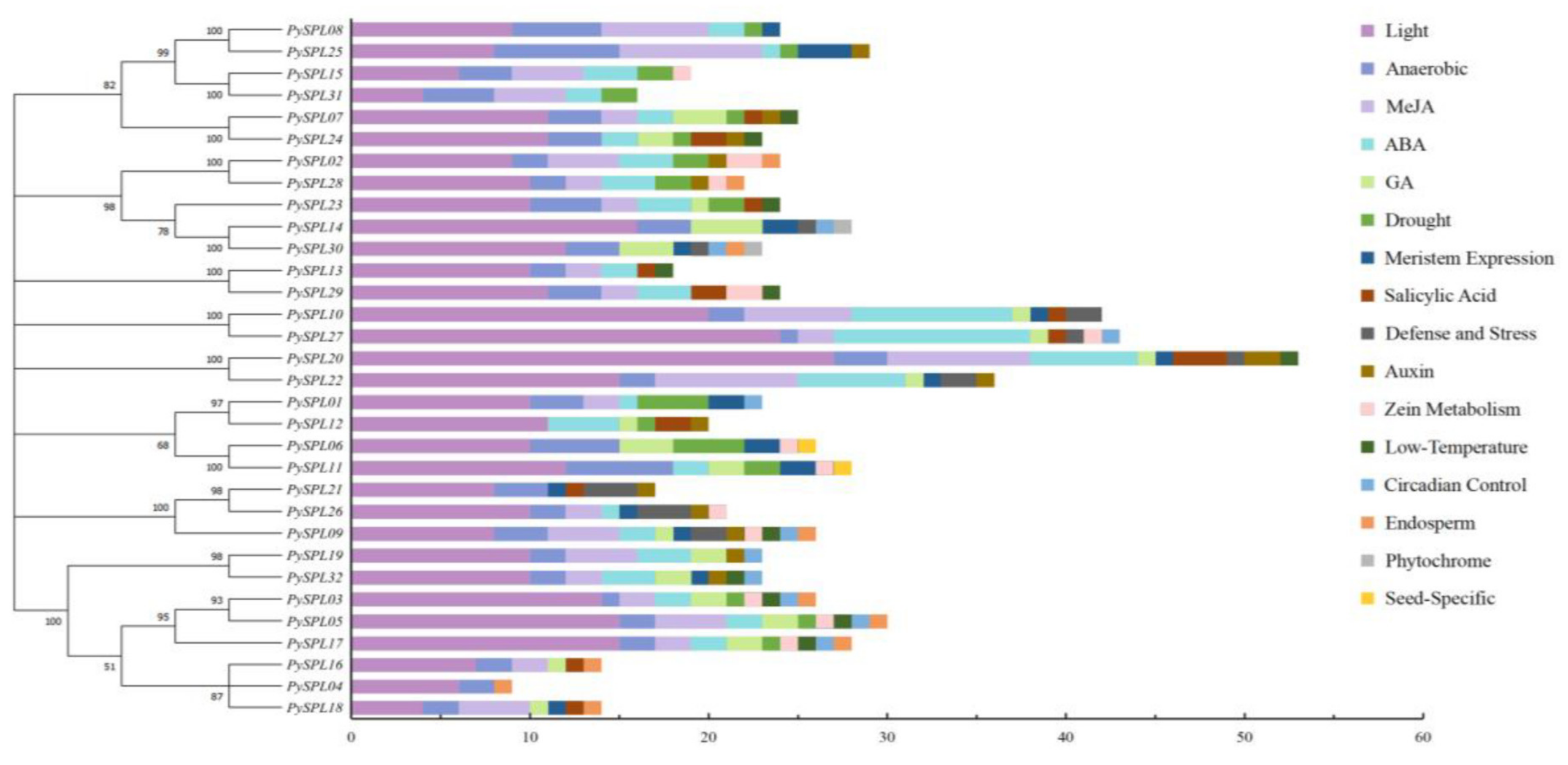
2.6. Cis-Acting Elements Analysis of PySPLs Promoter
2.7. Collinearity Analysis of PySPLs
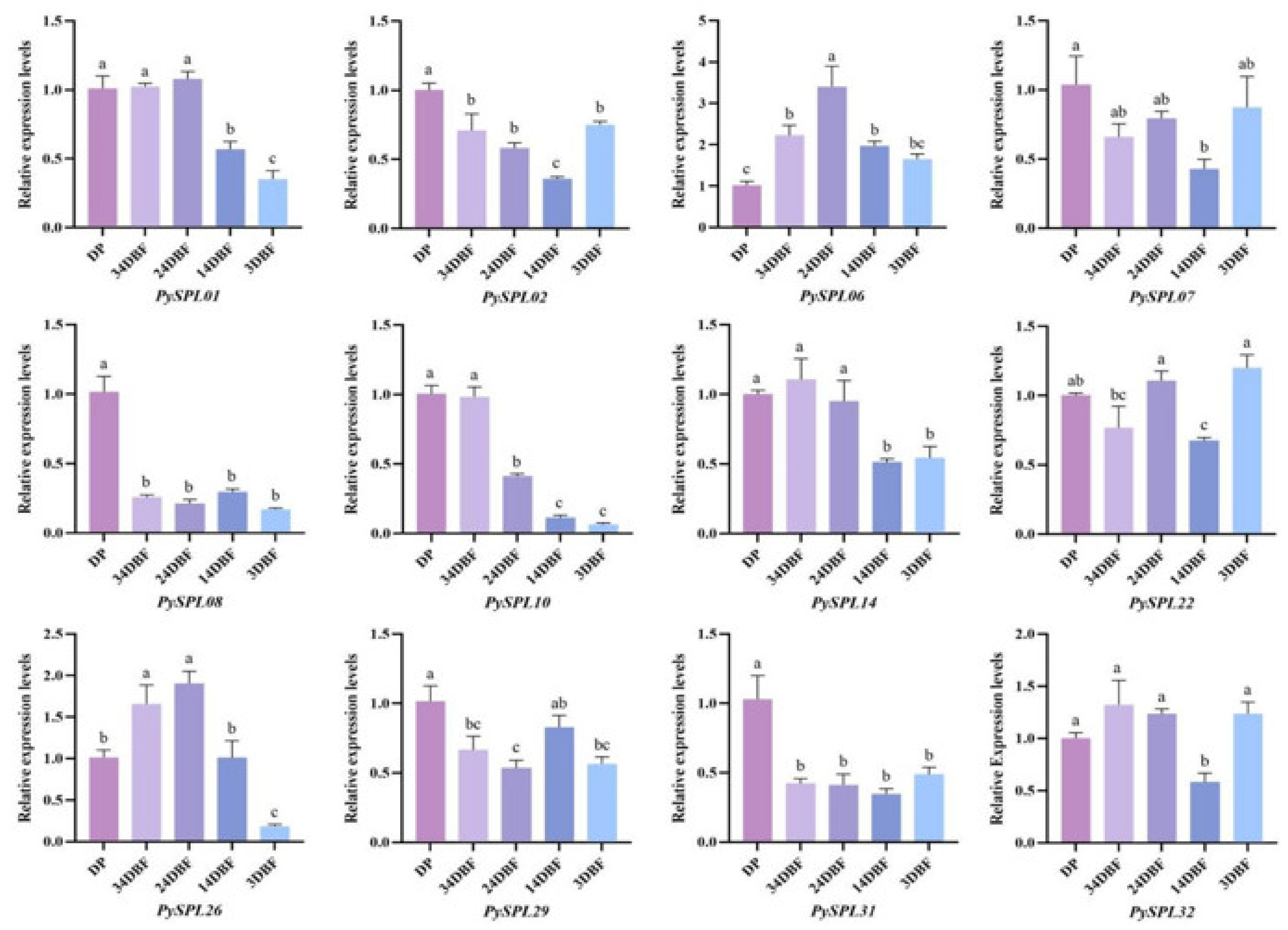
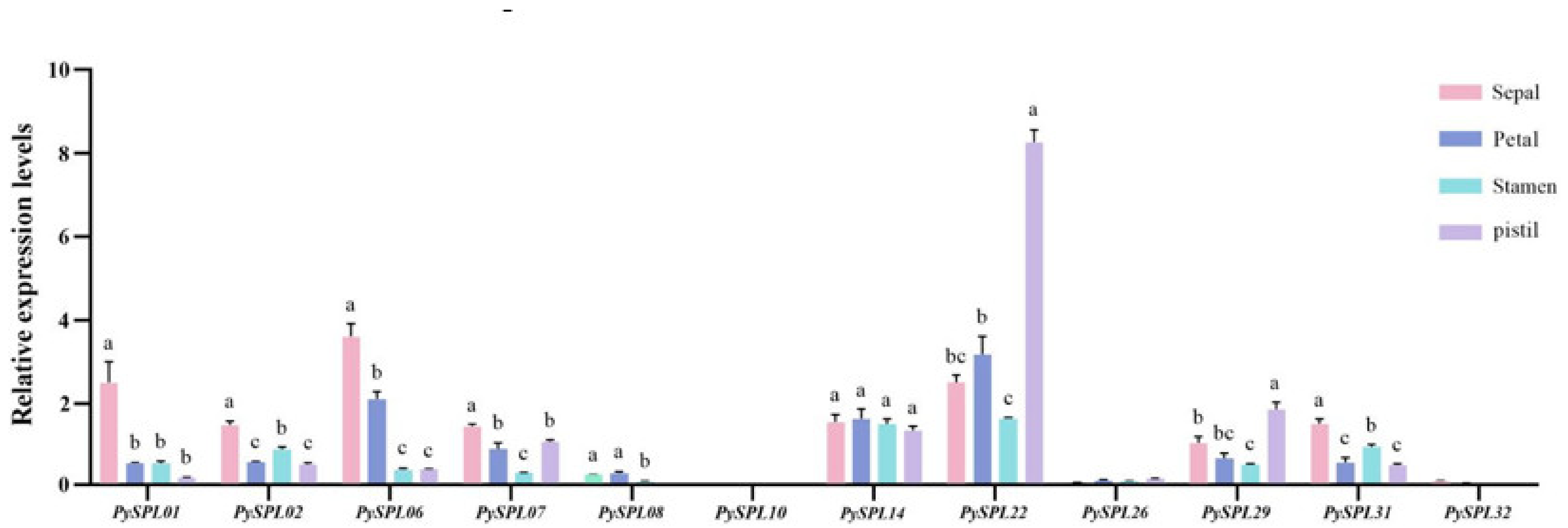
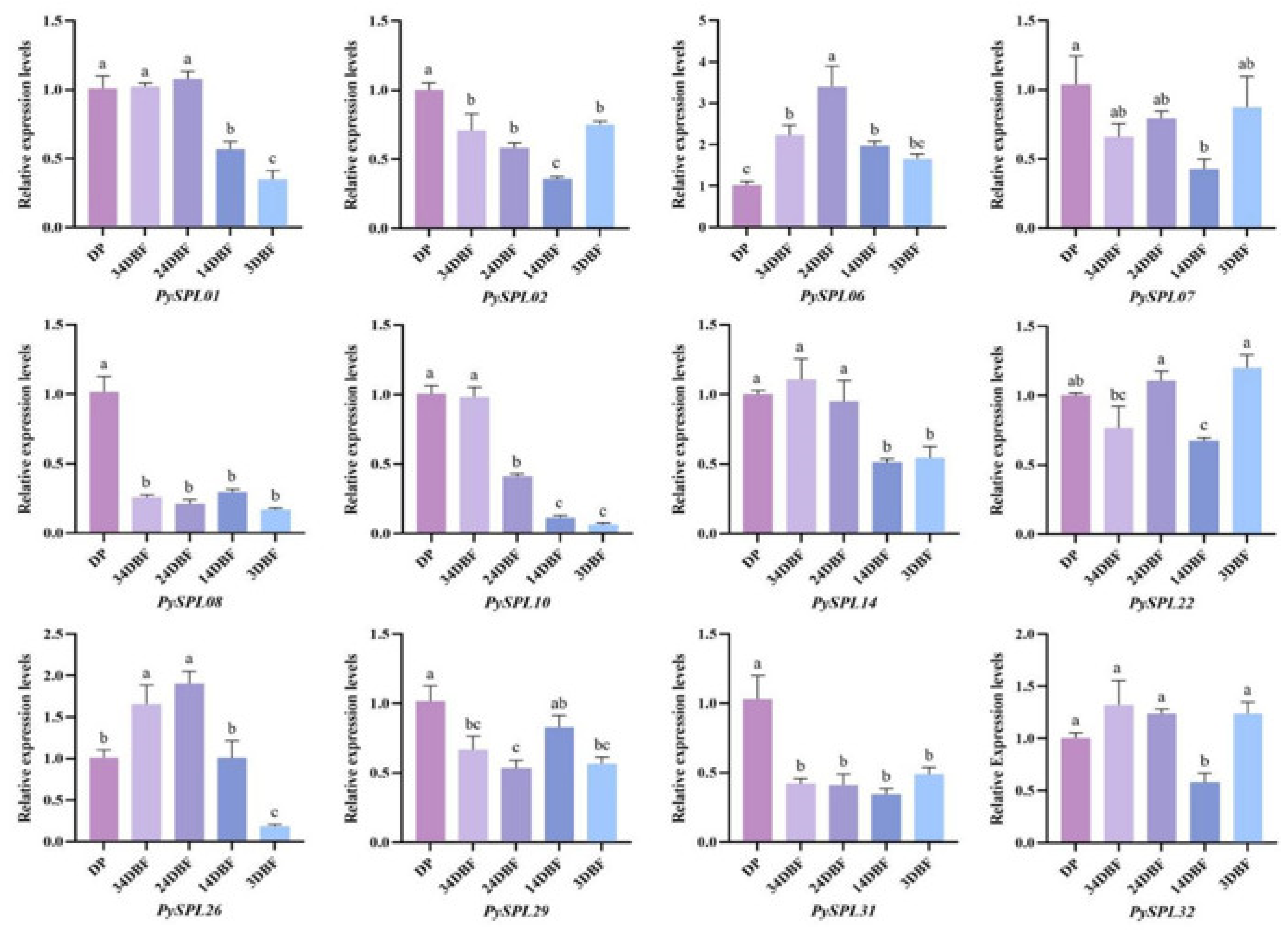
2.8. Expression Analysis of PySPLs in Flowering Induction
3. Discussion
4. Materials and Methods
4.1. Experimental Materials
4.2. Identification and Chromosomal Location of PySPLs
4.3. Phylogenetic Analysis of PySPLs
4.4. Conserved Domains, Conserved Motifs and Gene Structure Analysis of PySPLs
4.5. Cis-acting Elements Analysis of PySPLs Promoter
4.6. Syntenic Analysis of PySPLs
4.7. qRT-PCR
ΔΔCt = ΔCt (test sample) − ΔCt (standard sample)
ΔCt = Ct (Target gene) − Ct (Internal reference gene)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Luscombe, N.M.; Austin, S.E.; Berman, H.M.; Thornton, J.M. An overview of the structures of protein-DNA complexes. Genome Biol. 2000, 1, reviews001.1. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Kigawa, T.; Inou, M.; Tateno, M.; Yamasaki, T.; Yabuki, T.; Aoki, M.; Seki, E.; Matsuda, T.; Nunokawa, E.; et al. A novel zinc-binding motif revealed by solution structures of DNA-binding do-mains of Arabidopsis SBP-family transcription factors. J. Mol. Biol. 2004, 337, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Birkenbihl, R.P.; Jach, G.; Saedler, H.; Huijser, P. Functional dissection of the plant-specific sbp-domain: Overlap of the DNA-binding and nuclear localization domains. J. Mol. Biol. 2005, 352, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.; Saedler, H.; Huijser, P. A new family of DNA binding proteins includes putative transcriptional regulators of the Antirrhinum majus floral meristem identity gene squamosa. Mol. Gen. Genet. MGG 1996, 250, 7–16. [Google Scholar] [CrossRef]
- Wu, G.; Poethig, R.S. Temporal regulation of shoot development in Arabidopsis thaliana by miR156 and its target SPL3. Development 2006, 133, 3539–3547. [Google Scholar] [CrossRef]
- Hou, H.M.; Yan, Q.; Wang, X.P.; Xu, H. A sbp-box gene vpsbp5 from Chinese wild vitis species responds to erysiphe necator and defense signaling molecules. Plant Mol. Biol. Report. 2013, 31, 1261–1270. [Google Scholar] [CrossRef]
- Chao, L.; Liu, Y.; Chen, D.; Xue, X.; Mao, Y.; Chen, X. Arabidopsis transcription factors SPL1 and SPL12 confer plant thermo tolerance at reproductive stage. Mol. Plant 2017, 10, 735–748. [Google Scholar] [CrossRef]
- Nodine, M.D.; Bartel, D.P. MicroRNAs prevent precocious gene expression and enable pattern formation during plant embryogenesis. Genes Dev. 2010, 24, 2678–2692. [Google Scholar] [CrossRef]
- Martin, R.C.; Asahina, M.; Liu, P.P.; Kristof, J.R.; Coppersmith, J.L.; Pluskota, W.E.; Bassel, G.W.; Goloviznina, N.A.; Nguyen, T.T.; Martinez-Andújar, C.; et al. The microRNA156 and micro-RNA172 gene regulation cascades at post-germinative stages in Arabidopsis. Seed Sci. Res. 2010, 20, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Xing, S.; Salinas, M.; Höhmann, S.; Berndtgen, R.; Huijser, P. miR156-targeted and nontargeted SBP-box transcription factors act in concert to secure male fertility in Arabidopsis. Plant Cell 2010, 22, 3935–3950. [Google Scholar] [CrossRef]
- Cai, C.; Guo, W.; Zhang, B. Genome-wide identification and characterization of SPL transcription factor family and their evolution and expression profiling analysis in cotton. Sci. Rep. 2018, 8, 762. [Google Scholar] [CrossRef]
- Li, C.; Lu, S. Molecular characterization of the SPL gene family in Populus trichocarpa. BMC Plant Biol. 2014, 14, 131. [Google Scholar] [CrossRef]
- Li, X.; Lin, E.; Huang, H.; Niu, M.; Tong, Z.; Zhang, J. Molecular characterization of SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) gene family in Betula luminifera. Front. Plant Sci. 2018, 9, 608. [Google Scholar] [CrossRef]
- Shao, F.; Lu, Q.; Wilson, I.W.; Qiu, D. Genome-wide identification and characterization of the SPL gene family in Ziziphus jujuba. Gene 2017, 627, 315–321. [Google Scholar] [CrossRef]
- Shruti, L.; Leo Bryan, P.; Harley, S. Regulation of the SQUAMOSA PROMOTER-BINDING PROTEIN-LIKE genes/microRNA156 module by the homeodomain proteins PENNYWISE and POUND-FOOLISH in Arabidopsis. Mol. Plant 2011, 4, 1123–1132. [Google Scholar] [CrossRef]
- Cardon, G.H.; Höhmann, S.; Nettesheim, K.; Saedler, H.; Huijser, P. Functional analysis of the Arabidopsis thaliana SBP-box gene SPL3: A novel gene involved in the floral transition. Plant J. 1997, 12, 367–377. [Google Scholar] [CrossRef]
- Hou, H.; Yan, X.; Sha, T.; Yan, Q.; Wang, X. The sbp-box gene vpsbp11 from Chinese wild vitis is involved in floral transition and affects leaf development. Int. J. Mol. Sci. 2017, 18, 1493. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, Z.; Yang, Y.; Chen, X.; Chen, G. Function annotation of an sbp-box gene in Arabidopsis based on analysis of co-expression networks and promoters. Int. J. Mol. Sci. 2009, 10, 116–132. [Google Scholar] [CrossRef]
- Jung, J.; Ju, Y.; Seo, P.J.; Lee, J.; Prak, C. The SOC1-SPL module integrates photoperiod and gibberellic acid signals to control flowering time in Arabidopsis. Plant J. 2011, 69, 577–588. [Google Scholar] [CrossRef]
- Kim, J.J.; Lee, J.H.; Kim, W.; Jung, H.S.; Huijser, P.; Ahn, J.H. The microRNA156-SQUAMOSA PROMOTER BINDING PROTEIN-LIKE3 module regulates ambient temperature-responsive flowering via LOWERING LOCUS T in Arabidopsis. Plant Physiol. 2012, 159, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Czech, B.; Weigel, D. miR156-regulated SPL transcription factors define an endogenous flowering pathway in Arabidopsis thaliana. Cell 2009, 138, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.W.; Weigel, D.; Poethig, R.S. The sequential action of mir156 and mir172 regulates developmental timing in Arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Pan, H.; Wang, J.; Yang, W.; Zhang, Q. Identification and profiling of novel and conserved micrornas during the flower opening process in Prunus mume via deep sequencing. Mol. Genet. Genom. 2014, 289, 169–183. [Google Scholar] [CrossRef]
- Preston, J.C.; Jorgensen, S.A.; Orozco, R.; Hileman, L.C. Paralogous squamosa promoter binding protein-like (SPL) genes differentially regulate leaf initiation and reproductive phase change in petunia. Planta 2016, 243, 429–440. [Google Scholar] [CrossRef]
- Xie, K.; Wu, C.; Xiong, L. Genomic organization, differential expression, and interaction of squamosa promoter-binding-like transcription factors and microrna156 in rice. Plant Physiol. 2006, 142, 280–293. [Google Scholar] [CrossRef]
- Masahito, S.; Tomotsugu, K.; Nobutaka, M.; Masaru, O. Arabidopsis sbp-box genes SPL10, SPL11 and SPL2 control morphological change in association with shoot maturation in the reproductive phase. Plant Cell Physiol. 2009, 50, 2133. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Kohalmi, S.E.; Amyot, L.; Hannoufa, A. Squamosa promoter binding protein-like 2 controls floral organ development and plant fertility by activating asymmetric leaves 2 in Arabidopsis thaliana. Plant Mol. Biol. 2016, 92, 661–674. [Google Scholar] [CrossRef]
- Shirasawa, K.; Esumi, T.; Hirakawa, H.; Tanaka, H.; Itai, A.; Ghelfi, A.; Nagasaki, H.; Isobe, S. Phased genome sequence of an interspecific hybrid flowering cherry, ‘somei-yoshino’ (Cerasus × Yedoensis). DNA Res. 2019, 26, 1–29. [Google Scholar] [CrossRef]
- Song, J.; Ma, D.; Yin, J.; Yang, L.; Gao, C. Genome-wide characterization and expression profiling of squamosa promoter binding protein-like (SBP) transcription factors in wheat (Triticum aestivum L.). Agronomy 2019, 9, 527. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Wang, X.; Gu, S.; Hu, Z.; Hua, X.; Xu, C. Comparative study of SBP-box gene family in Arabidopsis and rice. Gene 2008, 407, 1–11. [Google Scholar] [CrossRef]
- Preston, J.C.; Hileman, L.C. Functional evolution in the plant squamosa-promoter binding protein-like (SPL) gene family. Front. Plant Sci. 2013, 4, 80. [Google Scholar] [CrossRef]
- Hideki, I.; Fyodor, K. The evolution of gene duplications: Classifying and distinguishing between models. Nat. Rev. Genet. 2010, 11, 97–108. [Google Scholar] [CrossRef]
- Michael, F. Bias in Plant Gene Content Following Different Sorts of Duplication: Tandem, Whole-Genome, Segmental, or by Transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef]
- Conant, G.C.; Wolfe, K.H. Turning a hobby into a job: How duplicated genes find new functions. Nat. Rev. Genet. 2008, 9, 938–950. [Google Scholar] [CrossRef]
- Ling, L.; Zhang, S. Unraveling the Distribution and Evolution of miR156-targeted SPLs in Plants by Phylogenetic Analysis. Plant Divers. Resour. 2012, 34, 33–46. [Google Scholar] [CrossRef]
- Jores, T.; Tonnies, J.; Wrightsman, T.; Buckler, E.S.; Cuperus, J.T.; Fields, S.; Queitsch, C. Synthetic promoter designs enabled by a comprehensive analysis of plant core promoters. Nat. Plants 2021, 7, 842–855. [Google Scholar] [CrossRef]
- Lee, J.; Oh, M.; Park, H.; Lee, I. SOC1 translocated to the nucleus by interaction with AGL24 directly regulates LEAFY. Plant J. 2008, 55, 832–843. [Google Scholar] [CrossRef]
- Schmid, M. Dissection of floral induction pathways using global expression analysis. Development 2003, 130, 6001–6012. [Google Scholar] [CrossRef]
- Preston, J.C.; Hileman, L.C. SQUAMOSA-PROMOTER BINDING PROTEIN 1 initiates flowering in Antirrhinum majus through the activation of meristem identity genes. Plant J. 2010, 62, 704–712. [Google Scholar] [CrossRef]
- Matsoukas, I.G. Interplay between sugar and hormone signaling pathways modulate floral signal transduction. Front. Genet. 2014, 5, 218. [Google Scholar] [CrossRef] [PubMed]
- Blümel, M.; Dally, N.; Jung, C. Flowering time regulation in crops-what did we learn from Arabidopsis? Curr. Opin. Biotechnol. 2015, 32, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Bouché, F.; Lobet, G.; Tocquin, P.; Périlleux, C. Flor-id: An interactive database of flowering-time gene networks in Arabidopsis thaliana. Nucleic Acids Res. 2015, 44, D1167–D1171. [Google Scholar] [CrossRef] [PubMed]
- Beemster, G.T.S.; Fiorani, F.; Inzé, D. Cell cycle: The key to plant growth control? Trends Plant Sci. 2003, 8, 154–158. [Google Scholar] [CrossRef]
- Mizukami, Y. A matter of size: Developmental control of organ size in plants. Curr. Opin. Plant Biol. 2001, 4, 533–539. [Google Scholar] [CrossRef]
- Usami, T.; Horiguchi, G.; Yano, S.; Tsukaya, H. The more and smaller cells mutants of Arabidopsis thaliana identify novel roles for SQUAMOSA PROMOTER BINDING PROTEIN-LIKE genes in the control of heteroblasty. Development 2009, 136, 955–964. [Google Scholar] [CrossRef]
- Jung, S.; Lee, T.; Cheng, C.-H.; Buble, K.; Zheng, P.; Yu, J.; Humann, J.; Ficklin, S.P.; Gasic, K.; Scott, K.; et al. 15 years of GDR: New data and functionality in the Genome Database for Rosaceae. Nucleic Acids Res. 2019, 47, D1137–D1145. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Farideh, C.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Marc, G.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, 265–268. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. Smart: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2020, 49, D458–D460. [Google Scholar] [CrossRef]
- Chou, K.; Shen, H. Cell-PLoc 2.0: An improved package of web-servers for predicting subcellular localization of proteins in various organisms. Nat. Sci. 2010, 2, 1090–1103. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Xia, R. Tbtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (itol) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; De Peer, Y.V.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene ID | Chromosome | Gene Length (bp) | CDS Length (bp) | Number of Aminocacids | Molecular Wieght (kD) | Theoretical pI | Instability Index | Aliphatic Index | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|---|---|
| PySPL01 | CYE_r3.1SPA0_g098740.1 | SPA0 | 1307 | 570 | 189 | 21,562.17 | 9.26 | 57.8 | 53.07 | −1.138 | Nucleus. |
| PySPL02 | CYE_r3.1SPA0_g102580.1 | SPA0 | 1665 | 1176 | 391 | 43,168.56 | 7.98 | 59.51 | 65.14 | −0.587 | Nucleus. |
| PySPL03 | CYE_r3.1SPA0_g124820.1 | SPA0 | 3667 | 876 | 291 | 32,288.48 | 9.65 | 48.7 | 64.60 | −0.537 | Nucleus. |
| PySPL04 | CYE_r3.1SPA0_g125190.1 | SPA0 | 3704 | 1038 | 346 | 37,451.89 | 9.94 | 49.04 | 78.9 | −0.256 | Cytoplasm. Nucleus. |
| PySPL05 | CYE_r3.1SPA0_g125250.1 | SPA0 | 3475 | 732 | 243 | 27,329.90 | 9.64 | 50.37 | 63.33 | −0.603 | Nucleus. |
| PySPL06 | CYE_r3.1SPA0_g142750.1 | SPA0 | 1354 | 489 | 162 | 18,589.14 | 5.75 | 98.68 | 36.17 | −1.457 | Nucleus. |
| PySPL07 | CYE_r3.1SPA1_g010660.1 | SPA1 | 3898 | 1443 | 480 | 52,717.28 | 8.75 | 42.71 | 54.48 | −0.774 | Nucleus. |
| PySPL08 | CYE_r3.1SPA1_g017590.1 | SPA1 | 4919 | 1500 | 499 | 55,318.71 | 7.59 | 50.89 | 61.5 | −0.654 | Nucleus. |
| PySPL09 | CYE_r3.1SPA2_g023420.1 | SPA2 | 2457 | 984 | 327 | 36,237.85 | 8.95 | 64.88 | 54.74 | −0.788 | Nucleus. |
| PySPL10 | CYE_r3.1SPA2_g023430.1 | SPA2 | 3059 | 1296 | 431 | 47,188.83 | 8.07 | 60.66 | 56.91 | −0.694 | Nucleus. |
| PySPL11 | CYE_r3.1SPA4_g004730.1 | SPA4 | 1811 | 486 | 161 | 18,419.89 | 5.92 | 96.44 | 36.4 | 1.456 | Nucleus. |
| PySPL12 | CYE_r3.1SPA4_g018900.1 | SPA4 | 1510 | 489 | 162 | 18,418.56 | 9.14 | 81.42 | 42.1 | −1.156 | Nucleus. |
| PySPL13 | CYE_r3.1SPA5_g000450.1 | SPA5 | 6368 | 3213 | 1070 | 118,298.50 | 8.29 | 55.62 | 76.48 | −0.471 | Nucleus. |
| PySPL14 | CYE_r3.1SPA5_g020750.1 | SPA5 | 3580 | 1215 | 404 | 44,268.17 | 8.26 | 51.4 | 62.95 | −0.687 | Nucleus. |
| PySPL15 | CYE_r3.1SPA5_g026190.1 | SPA5 | 4040 | 1671 | 556 | 60,834.75 | 7.05 | 45.3 | 67.48 | −0.524 | Nucleus. |
| PySPL16 | CYE_r3.1SPA6_g034400.1 | SPA6 | 4517 | 1152 | 383 | 41,145.63 | 9.5 | 55.23 | 53.73 | −0.679 | Nucleus. |
| PySPL17 | CYE_r3.1SPA6_g034460.1 | SPA6 | 2997 | 456 | 152 | 17,505.46 | 10.12 | 60.19 | 75.72 | −0.391 | Nucleus. |
| PySPL18 | CYE_r3.1SPA6_g047460.1 | SPA6 | 4467 | 1152 | 383 | 41,130.66 | 9.5 | 53.79 | 54.49 | −0.662 | Nucleus. |
| PySPL19 | CYE_r3.1SPA7_g008830.1 | SPA7 | 3356 | 1098 | 365 | 39,954.44 | 9.68 | 42.75 | 50.52 | −0.77 | Cytoplasm. Nucleus. |
| PySPL20 | CYE_r3.1SPE0_g030560.1 | SPE0 | 7304 | 2160 | 719 | 81,272.87 | 7.2 | 53.55 | 78.9 | −0.403 | Cytoplasm. Nucleus. |
| PySPL21 | CYE_r3.1SPE0_g159120.1 | SPE0 | 1785 | 1056 | 352 | 39,548.91 | 10.07 | 69 | 51.16 | −0.871 | Nucleus. |
| PySPL22 | CYE_r3.1SPE1_g001190.1 | SPE1 | 7316 | 2451 | 816 | 91,614.14 | 6.5 | 51.94 | 76.81 | −0.42 | Cytoplasm. Nucleus. |
| PySPL23 | CYE_r3.1SPE1_g014110.1 | SPE1 | 3126 | 951 | 316 | 34,821.23 | 9.54 | 60.85 | 65.19 | −0.615 | Cytoplasm. Nucleus. |
| PySPL24 | CYE_r3.1SPE1_g014420.1 | SPE1 | 4755 | 1440 | 479 | 52,541.15 | 8.85 | 45.11 | 55.41 | −0.765 | Nucleus. |
| PySPL25 | CYE_r3.1SPE1_g021460.1 | SPE1 | 5005 | 1500 | 499 | 55,328.80 | 7.85 | 49.39 | 63.47 | −0.621 | Nucleus. |
| PySPL26 | CYE_r3.1SPE2_g016170.1 | SPE2 | 1650 | 978 | 326 | 36,160.76 | 8.94 | 65.78 | 54.33 | −0.776 | Nucleus. |
| PySPL27 | CYE_r3.1SPE2_g016180.1 | SPE2 | 3397 | 1296 | 431 | 47,228.89 | 8.07 | 60.96 | 57.38 | −0.688 | Nucleus. |
| PySPL28 | CYE_r3.1SPE3_g036000.1 | SPE3 | 2243 | 1281 | 426 | 46,967.87 | 7.13 | 60.05 | 66.41 | −0.543 | Nucleus. |
| PySPL29 | CYE_r3.1SPE5_g000520.1 | SPE5 | 4666 | 3213 | 1070 | 118,346.45 | 8.21 | 56.96 | 75.83 | −0.479 | Nucleus. |
| PySPL30 | CYE_r3.1SPE5_g021220.1 | SPE5 | 3555 | 1212 | 403 | 44,157.07 | 8.27 | 51.85 | 63.35 | −0.681 | Nucleus. |
| PySPL31 | CYE_r3.1SPE5_g026260.1 | SPE5 | 3384 | 1674 | 557 | 61,020.94 | 6.72 | 46.49 | 68.58 | −0.511 | Cytoplasm. Nucleus. |
| PySPL32 | CYE_r3.1SPE7_g002940.1 | SPE7 | 3152 | 1140 | 379 | 41,296.61 | 9.6 | 51.09 | 47.89 | −0.811 | Nucleus. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, L.; Lyu, T.; Lyu, Y. Genome-Wide Analysis of the SPL Gene Family and Expression Analysis during Flowering Induction in Prunus × yedoensis ‘Somei-yoshino’. Int. J. Mol. Sci. 2022, 23, 10052. https://doi.org/10.3390/ijms231710052
Gao L, Lyu T, Lyu Y. Genome-Wide Analysis of the SPL Gene Family and Expression Analysis during Flowering Induction in Prunus × yedoensis ‘Somei-yoshino’. International Journal of Molecular Sciences. 2022; 23(17):10052. https://doi.org/10.3390/ijms231710052
Chicago/Turabian StyleGao, Lan, Tong Lyu, and Yingmin Lyu. 2022. "Genome-Wide Analysis of the SPL Gene Family and Expression Analysis during Flowering Induction in Prunus × yedoensis ‘Somei-yoshino’" International Journal of Molecular Sciences 23, no. 17: 10052. https://doi.org/10.3390/ijms231710052
APA StyleGao, L., Lyu, T., & Lyu, Y. (2022). Genome-Wide Analysis of the SPL Gene Family and Expression Analysis during Flowering Induction in Prunus × yedoensis ‘Somei-yoshino’. International Journal of Molecular Sciences, 23(17), 10052. https://doi.org/10.3390/ijms231710052