
Stem Cell Models for Breast and Colon Cancer: Experimental Approach for Drug Discovery


Abstract
:1. Introduction
2. Experimental Models
2.1. Parental Breast and Colon Cancer Cell Lines
2.2. Conventional/Targeted Therapy
2.3. Pharmacological Inhibition
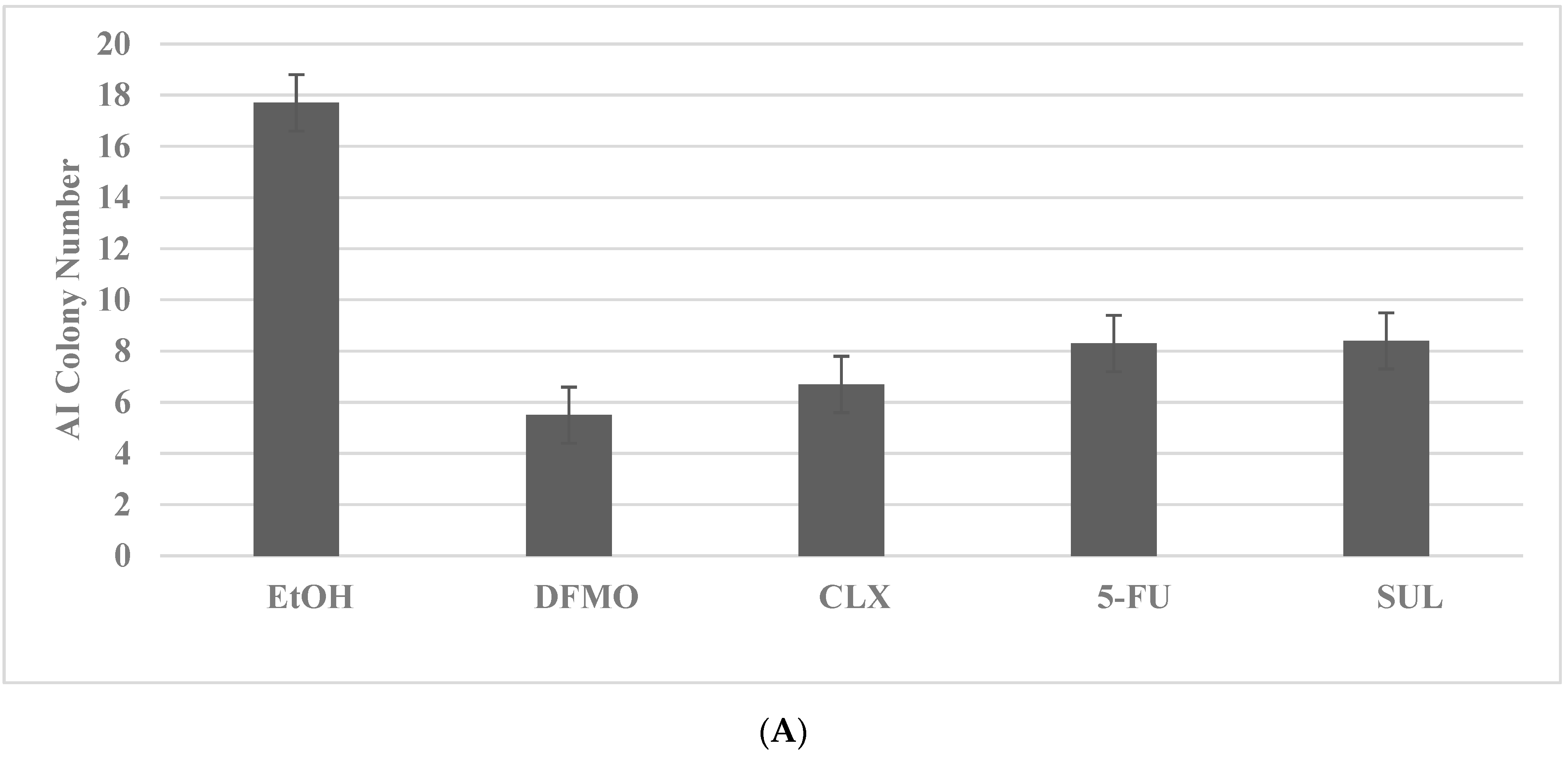
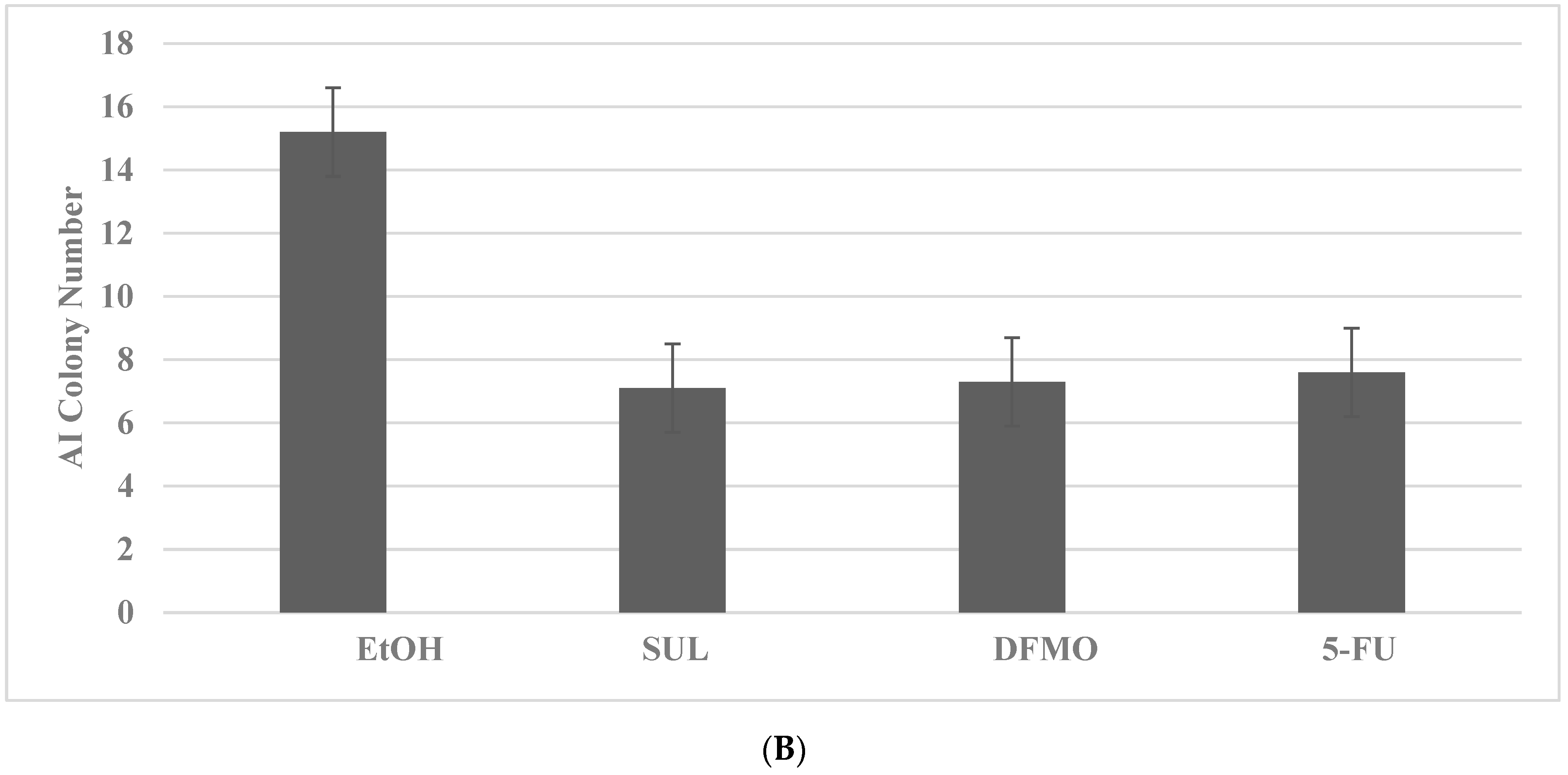
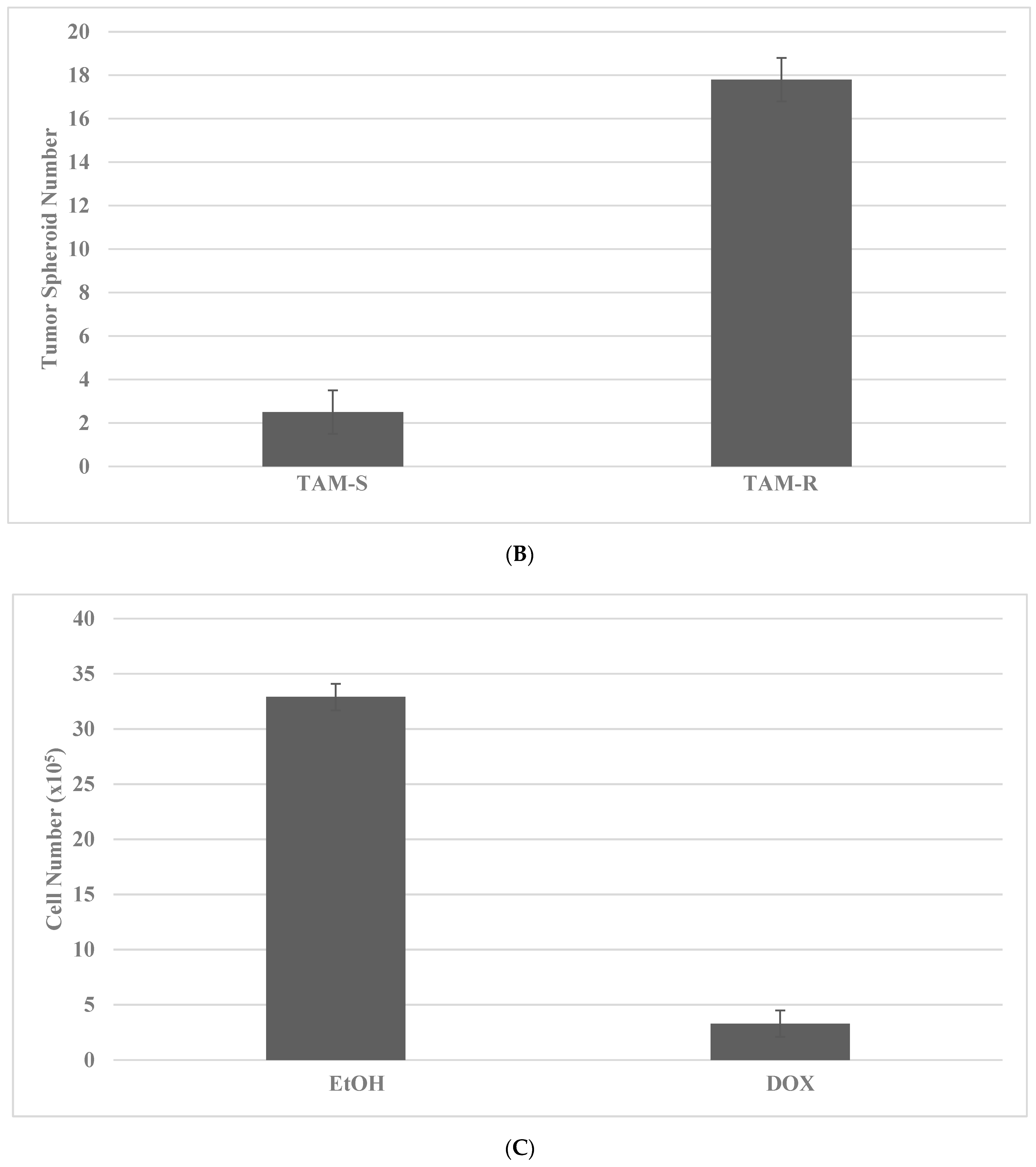
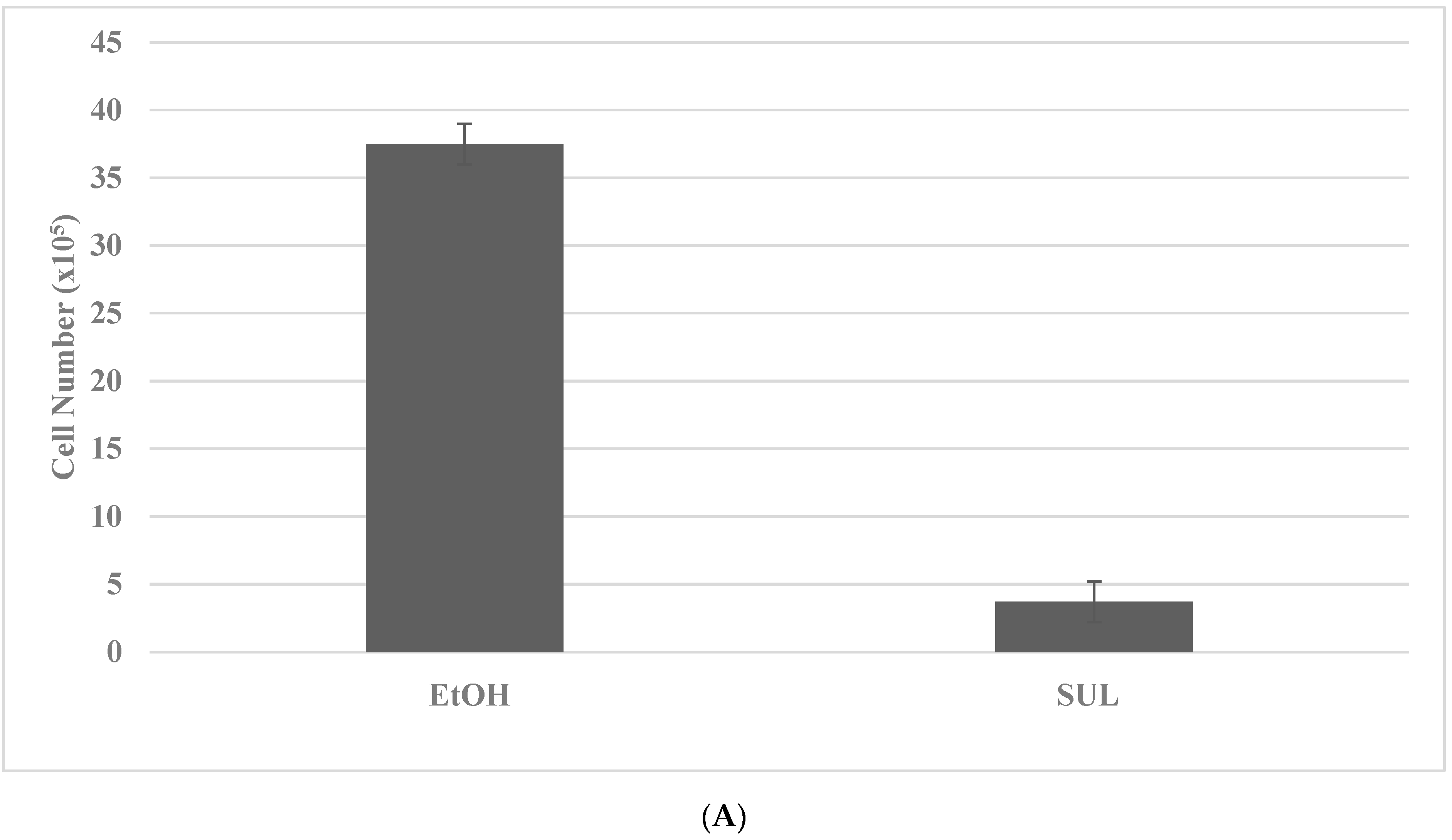
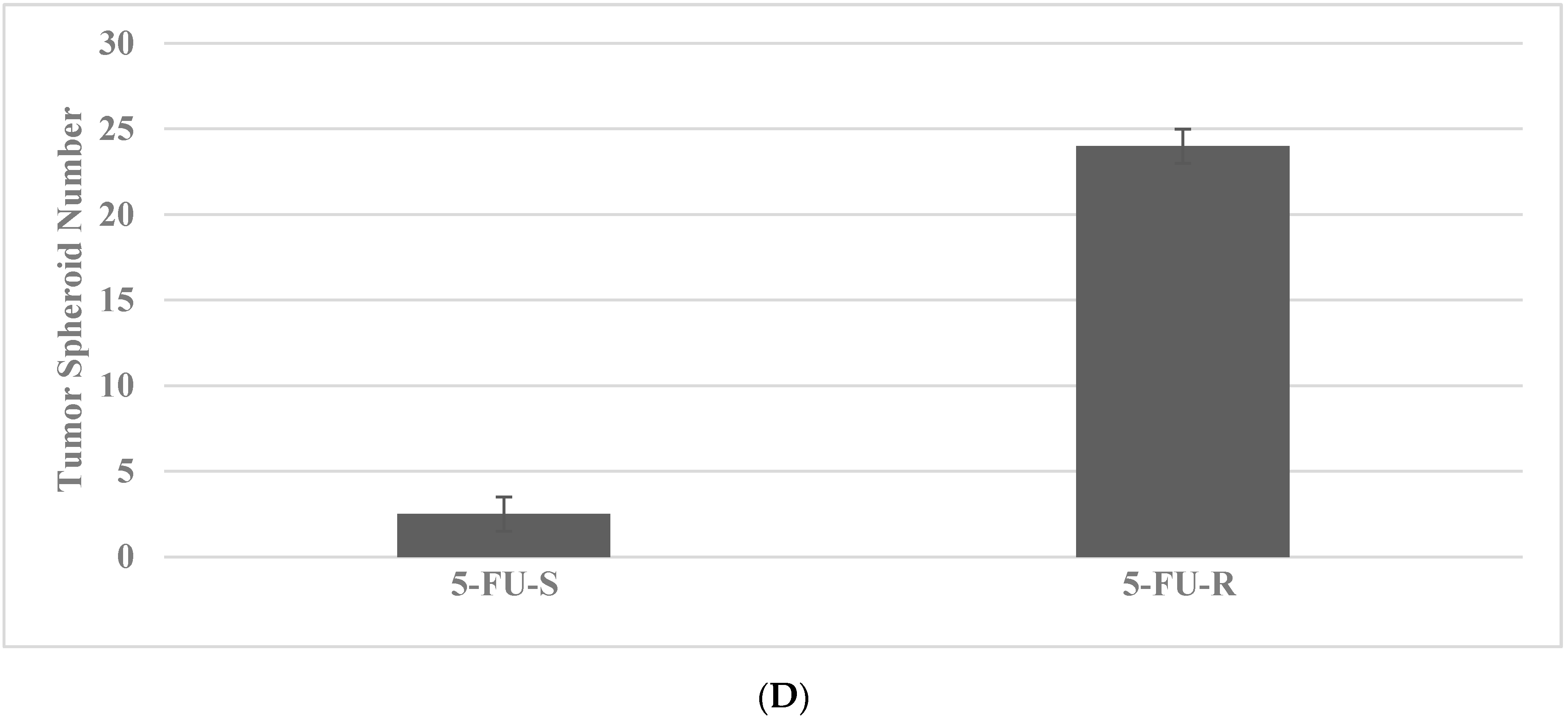
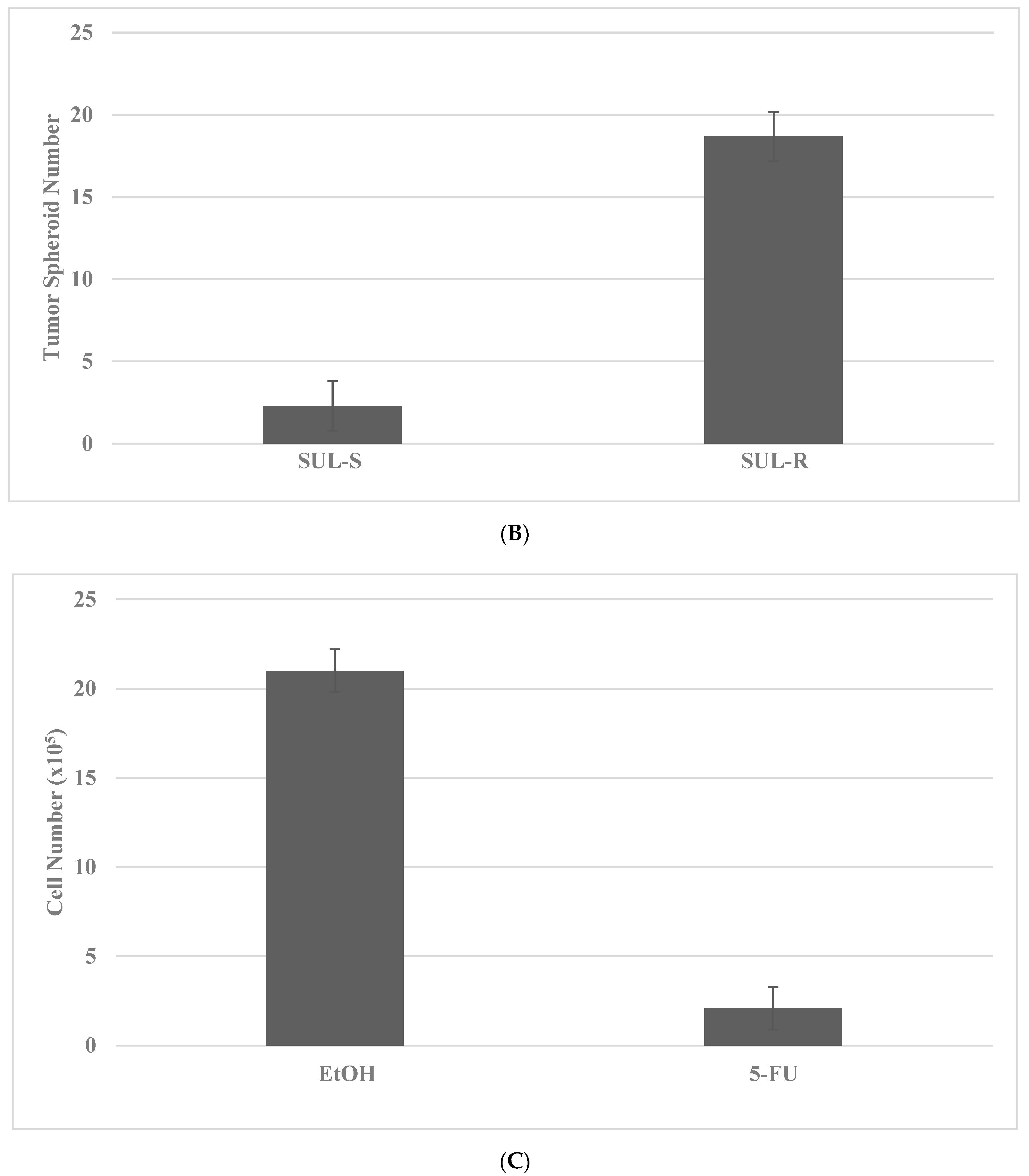
2.4. Drug-Resistant Stem Cell Models
3. Experimental Modulation
3.1. HER-2-Enriched Breast Cancer
3.2. FAP Model for Colon Cancer
4. Conclusions
5. Future Research
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Cancer Society Facts and Figures-2021; American Cancer Society: Atlanta, GA, USA, 2021.
- National Comprehensive Cancer Network-2022. Available online: http://www.nccn.org (accessed on 8 July 2022).
- Li, Y.; Wicha, M.S.; Schwartz, S.J.; Sun, D. Implication of cancer stem cell theory for cancer chemoprevention by natural dietary compounds. J. Nutr. Biochem. 2011, 22, 799–806. [Google Scholar] [CrossRef]
- Moy, B.; Goss, P.E. Estrogen receptor pathway: Resistance to endocrine therapy and new therapeutic approaches. Clin. Cancer Res. 2006, 12, 4790–4793. [Google Scholar] [CrossRef]
- Mathews, J.; Gustafson, J.-A. Estrogen signaling: A subtle balance between ER-α and ER-β. Mol. Interv. 2003, 3, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Cho, N.L.; Javid, S.H.; Carrothers, A.M.; Redston, M.; Bertagnolli, M.M. Estrogen receptor α and β are inhibitory modifiers of Apc-dependent tumorigenesis in the proximal colon of Min+/mice. Cancer Res. 2007, 67, 2366–2372. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.; Di Leo, A.; Niv, Y.; Gustafson, J.-A. Estrogen receptor-β as a target for colorectal cancer prevention. Cancer Lett. 2016, 372, 28–56. [Google Scholar] [CrossRef] [PubMed]
- Filho, R.P.S.; Junior, S.A.; Begnami, M.D.; Farreira, F.O.; Nakagava, W.T. Estrogen receptor-β as a prognostic marker of tumor progression in colorectal cancer with familial adenomatous polyps and sporadic polyps. Pathol. Oncol. Res. 2018, 24, 533–540. [Google Scholar] [CrossRef]
- Ye, J.; Jia, Y.; Ji, K.-E.; Saunders, A.J.; Xue, K.; Ji, J.; Mason, M.D.; Jiang, W.G. Traditional Chinese medicine in the prevention and treatment of cancer and cancer metastasis. Oncol. Lett. 2015, 10, 1240–1250. [Google Scholar] [CrossRef] [PubMed]
- Aiello, P.; Sharghi, M.; Mansourkhani, S.M.; Ardekan, A.P.; Jouibari, L.; Daraei, N.; Piero, K.; Mahamodian, S.; Rezaei, M.; Heidari, M.; et al. Medicinal plants in the treatment and prevention of colon cancer. Oxidative Med. Cell. Longev. 2019, 2019, 2075614. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, Q.; Yu, L.; Zhu, J.; Cao, Y.; Gao, X. The signaling pathways and targets of traditional Chinese medicine and natural medicine in triple-negative breast cancer. J. Ethnopharmacol. 2021, 264, 113249. [Google Scholar] [CrossRef] [PubMed]
- Shibue, T.; Weinberg, R.A. EMT, CSCs and drug resistance. Nat. Rev. Clin. Oncol. 2017, 14, 611–629. [Google Scholar] [CrossRef]
- Lytle, N.K.; Barber, A.G.; Reya, T. Stem cell faith in cancer growth, progression and therapy resistance. Nat. Rev. Cancer 2018, 18, 669–680. [Google Scholar] [CrossRef]
- Gooding, A.J.; Schiemann, W.P. Epithelial-mesenchymal transition programs and cancer stem cell phenotypes: Mediators of breast cancer therapy resistance. Mol. Cancer Res. 2020, 18, 1257–1270. [Google Scholar] [CrossRef]
- Su, L.K.; Kinzler, K.W.; Vogelstein, B.; Preisinger, A.C.; Moser, A.R.; Luongo, C.; Gould, K.A.; Dove, W.F. Multiple intestinal neoplasia caused by a mutation in the murine homolog of the APC gene. Science 1992, 256, 668–670. [Google Scholar] [CrossRef]
- Fodde, R.; Edelmann, W.; Yang, K.; Van Leuven, C.; Carlson, C.; Renault, B.; Brukel, C.; Alt, E.; Lipkin, M.; Khan, P.M.; et al. A targeted chain-termination mutation in the mouse Apc gene results in multiple intestinal tumors. Proc. Natl. Acad. Sci. USA 1994, 91, 8969–8973. [Google Scholar]
- Edelmann, W.; Yang, K.; Karaguchi, M.; Heyer, J.; Lia, M.; Kneitz, B.; Fan, K.; Brown, A.M.; Lipkin, M.; Kucherlapati, R. Tumorigenesis in Mlh1/1638 N mutant mice. Cancer Res. 1999, 59, 1301–1307. [Google Scholar] [PubMed]
- Karaguchi, M.; Edelmann, W.; Yang, K.; Lipkin, M.; Kucherlapati, R.; Brown, A.M. Tumor-associated Apc mutations in Mlh1−/− Apc 1638N mice reveal a mutational significance of Mlh1 deficiency. Oncogene 2000, 19, 5755–5763. [Google Scholar] [CrossRef]
- Telang, N.; Katdare, M. Novel cell culture model for prevention of carcinogenic risk in in familial adenomatous polyposis syndrome. Oncol. Rep. 2009, 21, 1017–1021. [Google Scholar] [CrossRef]
- Telang, N. Anti-inflammatory drug resistance selects putative cancer stem cells in a cellular model for genetically predisposed colon cancer. Oncol. Lett. 2018, 15, 642–648. [Google Scholar] [CrossRef]
- Telang, N. Isolation and characterization of chemo-resistant stem cells from a mouse model of hereditary non-polyposis colon cancer. Stem Cells Cloning Adv. Appl. 2021, 14, 19–25. [Google Scholar]
- Dean, M.; Fojo, T.; Bates, S. Tumor stem cells and drug resistance. Nat. Rev. Cancer 2005, 5, 275–284. [Google Scholar] [CrossRef]
- Patel, S.A.; Ndabahaliye, A.; Lim, P.K.; Milton, R.; Rameshwar, P. Challenges in the development of future treatments for breast cancer stem cells. Breast Cancer 2010, 2, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Barker, N. Adult intestinal stem cells: Critical drivers of epithelial homeostasis and regeneration. Nat. Rev. Mol. Cell. Biol. 2014, 15, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Soteriou, D.; Fuchs, Y. A matter of life and death: Stem cell survival in tissue regeneration and tumor formation. Nat. Rev. Cancer 2018, 18, 187–201. [Google Scholar] [CrossRef]
- Kouri, J.; Zhong, L.; Hao, J. Targeting signaling pathways in cancer stem cells for cancer treatment. Stem Cell Int. 2017, 2017, 2925869. [Google Scholar] [CrossRef] [PubMed]
- Kudaravalli, S.; Hollander, P.D.; Mani, S.A. Role of p38 MAP kinase in cancer stem cells and metastasis. Oncogene 2022, 41, 3177–3185. [Google Scholar] [CrossRef]
- Kumari, M.; Krishnamurthy, P.T.; Sola, P. Targeted drug therapy to overcome chemo-resistance in triple-negative breast cancer. Curr. Cancer Drug Targets 2020, 20, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.G.; Collier, A.L.; Lee, D.; Hoeffer, R.A.; Zelava, V.; van Reesema, L.L.S.; Tang-Tan, A.M.; Guye, M.L.; Chang, D.Z.; Winston, J.S.; et al. Perspectives on triple-negative breast cancer: Current treatment strategies, unmet needs and potential targets for future therapies. Cancers 2020, 12, 2392. [Google Scholar] [CrossRef] [PubMed]
- Gorican, L.; Gole, B.; Potocnic, U. Head and neck cancer stem cell-enriched spheroid model for anticancer compound screening. Cells 2020, 9, 1707. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Desai, R.; Conrad, D.N.; Leite, N.C.; Akshinthala, D.; Lim, C.M.; Gonzalez, R.; Muthuswamy, L.B.; Gartener, Z.; Muthuswamy, S.K. Commitment and oncogene-induced plasticity of human stem cell-derived pancreatic acinar and ductal organoids. Stem Cell Cell 2021, 28, 1090–1104. [Google Scholar] [CrossRef] [PubMed]
- Breunig, M.; Merkle, J.; Wagner, M.; Melzer, M.K.; Barth, T.F.E.; Engleitner, T.; Krumm, J.; Weidenmann, S.; Cohrs, C.M.; Perkhofer, H.; et al. Modelling plasticity and dysplasia of pancreatic ductal organoids derived from human pluripotent stem cells. Cell Stem Cell 2021, 28, 1105–1124. [Google Scholar] [CrossRef]
- Ponomarev, A.; Gilazieva, Z.; Solovyeva, V.; Allegrucci, C.; Rizanov, A. Intrinsic and extrinsic factors impacting cancer stemness and tumor progression. Cancers 2022, 14, 970. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.E.; Nambiar, R.; De Souza, C.; Nguyen, A.; Chien, J.; Lam, K.S. Targeting epigenetic modifiers of tumor plasticity and cancer stem cell behavior. Cells 2022, 11, 1403. [Google Scholar] [CrossRef] [PubMed]
- Telang, N. The divergent effects of ovarian steroid hormones in the MCF-7 model for Luminal A breast cancer: Mechanistic leads for therapy. Int. J. Mol. Sci. 2022, 23, 4800. [Google Scholar] [CrossRef] [PubMed]
- Telang, N. Drug-resistant stem cells: Novel approach for colon cancer therapy. Int. J. Mol. Sci. 2022, 23, 2519. [Google Scholar] [CrossRef]
- Telang, N. Stem cell models for cancer therapy. Int. J. Mol. Sci. 2022, 23, 7055. [Google Scholar] [CrossRef]
- Park, I.H.; Zhou, R.; West, J.A.; Yabuchi, A.; Huo, H.; Ince, T.A.; Larou, P.H.; Lensch, M.W.; Daley, G.Q. Reprograming of human somatic cells to pluripotency with defined factors. Nature 2008, 151, 141–146. [Google Scholar] [CrossRef]
- Yu, J.; Hu, K.; Smuga-Otto, K.; Tian, S.; Stuart, R.; Slukvin, I.I.; Thompson, J.A. Human induced pluripotent cells free of vector and transgene sequences. Science 2009, 324, 797–801. [Google Scholar] [CrossRef]
- Katdare, M.; Osborne, M.P.; Telang, N.T. Soy isoflavone genistein modulates cell cycle progression and induces apoptosis in HER-2/neu oncogene expressing human breast epithelial cells. Int. J. Oncol. 2002, 21, 809–816. [Google Scholar] [CrossRef]
- Naujokat, C.; Mc Kee, D.L. The ‘Big Five’ phytochemicals targeting the cancer stem cells: Curcumin, EGCG, sulforaphane, resveratrol, and genistein. Curr. Med. Chem. 2021, 28, 4321–4342. [Google Scholar] [CrossRef]
- Manogaran, P.; Umapathy, D.; Karthikeyan, M.; Venkatachalam, K.; Singarvelu, A. Dietary phytochemicals as a potential source for targeting cancer stem cells. Cancer Investig. 2021, 39, 349–368. [Google Scholar] [CrossRef]
- Meerson, A.; Khatib, S.; Mahjna, J. Natural products targeting cancer stem cells for augmenting cancer therapeutics. Int. J. Mol. Sci. 2021, 22, 13044. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Tan, H.Y.; Li, S.; Cheung, F.; Wang, N.; Nagamatsu, T.; Feng, Y. Cancer stem cells: The potential targets of Chinese medicines and their active compounds. Int. J. Mol. Sci. 2016, 17, 893. [Google Scholar] [CrossRef] [PubMed]
- Nunes, T.; Hamdan, D.; El Boutachtaoui, M.D.; Gapihan, G.; Ngyuen, T.T.; Meles, S.; Angeli, E.; Ratajczak, P.; Lu, H.; Di Benedetto, M.; et al. Targeting cancer stem cells to overcome chemo-resistance. Int. J. Mol. Sci. 2018, 19, 4036. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Khan, M.A.; Du, J.; Cheng, J.; Tania, M.; Leung, E.L.-H.; Fu, J. Cordycepin inhibits triple-negative breast cancer migration and invasion by regulating EMT-TFs SLUG, TWIST1, SNAIL1 and ZEB1. Front. Oncol. 2022, 12, 898583. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Desai, N.S. Cancer stem cells: Acquisition, characteristics therapeutic implications targeting strategies and future prospects. Stem Cell Rev. Rep. 2019, 15, 331–355. [Google Scholar] [CrossRef]
- Wang, X. Stem cells in tissues, organoids and cancer. Cell. Mol. Life Sci. 2019, 76, 4043–4070. [Google Scholar] [CrossRef]
- Huang, B.; Yan, X.; Li, Y. Cancer stem cells for tumor therapy. Cancers 2021, 13, 4841. [Google Scholar] [CrossRef]
- Sharma, H.W.; Sokolski, J.A.; Perez, J.R.; Maltese, J.Y.; Sartorelli, A.C.; Stein, C.A.; Nichols, G.; Khaled, Z.; Telang, N.T.; Narayanan, R. Differentiation of immortal cells inhibits telomerase activity. Proc. Natl. Acad. Sci. USA 1995, 92, 12343–12346. [Google Scholar] [CrossRef] [PubMed]
- Fragiadaki, P.; Ranieri, E.; Kalliantasi, K.; Kouvidi, E.; Apalaki, E.; Vakonaki, E.; Mamoulakis, C.; Spandidos, D.A.; Tsalsakis, A. Telomerase inhibition and activation: A systematic review. Mol. Med. Rep. 2022, 25, 158. [Google Scholar] [CrossRef]
- Shi, R.; Radulovich, N.; Ng, C.; Liu, N.; Notsuda, H.; Cabanero, M.; Martins-Filho, S.N.; Raghavan, V.; Li, Q.; Mer, A.S.; et al. Organoid cultures as preclinical models of non-small cell lung cancer. Clin. Cancer Res. 2019, 26, 1162–1174. [Google Scholar] [CrossRef]
- Bednar, F.; Simeone, D.M. Pancreatic cancer stem cell biology and its therapeutic implications. J. Gastroenterol. 2011, 46, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- Farshadi, E.A.; Chang, J.; Sampadi, B.; Dukas, M.; Van’t Land, F.; Sijde, F.V.D.; Vietsch, E.E.; Pothof, J.; Koerkamp, B.G.; Van Eijck, C.H.J. Organoids derived from neoadjuvant FOLFIRINOX patients recapitulate therapy resistance in pancreatic ductal adenocarcinoma. Clin. Cancer Res. 2021, 27, 6602–6612. [Google Scholar] [CrossRef] [PubMed]
- Crespo, M.; Villar, E.; Tsai, S.Y.; Chang, K.; Amin, S.; Srinivasan, T.; Zhang, T.; Pipalia, N.H.; Chen, H.J.; Witherspoon, M.; et al. Colonic organoids derived from human induced pluripotent stem cells for modeling colorectal cancer and drug testing. Nat. Med. 2017, 23, 878–884. [Google Scholar] [CrossRef] [PubMed]
- Sachs, N.; Di Ligt, J.; Kopper, Z.O.; Gogola, E.; Bouvnova, G.; Weeber, F.; Balgobind, A.V.; Wind, K.; Gracanin, A.; Begthel, H.; et al. A living biobank of breast cancer organoids captures disease heterogeneity. Cell 2018, 172, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Drost, J.; Clevers, H. Organoids in cancer research. Nat. Rev. Cancer 2018, 18, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Devall, M.A.M.; Drew, D.A.; Dampier, C.H.; Plummer, S.J.; Eaton, S.; Bryant, J.; Diez-obrero, V.; Mo, J.; Kedrin, D.; Zerjav, D.C.; et al. Transcriptome-wide in vitro effects of aspirin in patient-derived normal colon organoids. Cancer Prev. Res. 2021, 14, 1089–1100. [Google Scholar] [CrossRef]
- Cao, J.; Chan, W.C. Use of conditional reprograming cell, patient-derived xenograft and organoid for drug screening for individualized prostate cancer therapy: Current and future perspectives (Review). Int. J. Oncol. 2022, 60, 52. [Google Scholar] [CrossRef] [PubMed]
- Hsu, K.-S.; Adileh, M.; Martin, M.L.; Makarov, V.; Chen, J.; Wu, C.; Bodo, S.; Klinger, S.; Gabriel, C.-E.; Szeglin, B.C.; et al. Colorectal cancer develops inherent radio-sensitivity that can be predicted using patient-derived organoids. Cancer Res. 2022, 82, 2298–2312. [Google Scholar] [CrossRef]
- Bruna, A.; Rueda, O.M.; Greenwood, W.; Batra, A.S.; Callari, M.; Batra, R.N.; Pogrebniak, K.; Sandoval, J.; Cassidy, J.E.; Tufegdzic-Vidakovic, A.; et al. A biobank of breast cancer explants with preserved intra-tumor heterogeneity to screen anticancer compounds. Cell 2016, 167, 260–274. [Google Scholar] [CrossRef]
- Gavert, N.; Zwang, Y.; Weiser, R.; Greenberg, O.; Halperin, S.; Jacoby, O.; Mallel, G.; Sandler, O.; Berger, A.J.; Stossel, E.; et al. Ex vivo organotypic cultures for synergistic therapy prioritization identify patient specific responses to combined MEK and Src inhibition in colorectal cancer. Nat. Cancer 2022, 3, 219–231. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parental Cells | Clinical and Biological Markers | Clinical Subtype | |||||
|---|---|---|---|---|---|---|---|
| ER | PR | HER-2 | AICF | Tumor | |||
| Breast | |||||||
| 184-B5 | - | - | - | - | - | Normal breast | |
| MCF-7 | + | + | + | + | + | Luminal A | |
| MDA-MB-231 | - | - | - | + | + | Triple-negative | |
| Colon | Apc [+/+] | Mlh1 [+/+] | |||||
| C57 COL | + | + | - | - | Normal colon | ||
| 1638N COL | - | + | + | + | Apc mutation codon 1638. FAP | ||
| Mlh1/1638N COL | - | - | + | + | HNPCC | ||
| Organ Type | Therapy | Clinical Subtype | |
|---|---|---|---|
| Conventional | Targeted | ||
| Breast | Multi-drug combination DOX, PCT, CPT | SERM, CDKI, AI, HER-2 inhibitors | Luminal A, Luminal B, TNBC |
| Organ Type | Therapy | Clinical Subtype | |
|---|---|---|---|
| Conventional | Targeted | ||
| Colon | Multi-drug combination 5-FU, OPN, ITC | NSAID COX-2 inhibitors ODC inhibitors | FAP, HNPCC sporadic cancer |
| Agent | Concentration | Stem Cell Model |
|---|---|---|
| Breast | ||
| Tamoxifen (TAM) | 1.5 µM | TAM-R Luminal A |
| Doxorubicin (DOX) | 0.5 µM | DOX-R TNBC |
| Colon | ||
| Sulindac (SUL) | 10 µM | SUL-R, FAP |
| 5-fluoro-uracil (5-FU) | 0.2 µM | 5-FU-R, HNPCC |
| Organ Type | Molecular Marker | Clinical Subtype | ||
|---|---|---|---|---|
| CD44 | NANOG | OCT-4 | ||
| Breast | ||||
| TAM-R | + | + | + | Luminal A |
| DOX-R | + | + | + | TNBC |
| Colon | CD44 | CD133 | c-Myc | |
| SUL-R | + | + | + | FAP |
| 5-FU-R | + | + | + | HNPCC |
| Treatment | Concentration | Stem Cell Markers | |||
|---|---|---|---|---|---|
| TS | CD44 | NANOG | OCT-4 | ||
| DMSO | 0.1% | 14.8 ± 1.9 | 20.8 ± 4.4 | 11.8 ± 3.1 | 14.2 ± 3.8 |
| CSOL | 5 µM | 1.9 ± 0.2 | 2.9 ± 0.6 | 2.7 ± 0.7 | 2.9 ± 0.9 |
| X2 | 7.74 | ||||
| p-value | 0.010 | 0.010 | 0.020 | 0.020 | |
| Inhibition | 87.2% | 86.0% | 77.1% | 79.6% | |
| Treatment | Concentration | Stem Cell Markers | |||
|---|---|---|---|---|---|
| TS | CD44 | CD133 | c-Myc | ||
| EtOH | 0.01% | 20.1 ± 4.0 | 15.9 ± 3.4 | 16.7 ± 3.4 | 8.3 ± 1.8 |
| CUR | 10 µM | 4.0 ± 0.3 | 3.5 ± 0.3 | 5.3 ± 0.5 | 3.8 ± 0.4 |
| X2 | 7.74 | ||||
| p-value | <0.01 | 0.01 | 0.01 | 0.03 | |
| Inhibition | 80.1% | 77.9% | 68.3% | 54.2% | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Telang, N.T. Stem Cell Models for Breast and Colon Cancer: Experimental Approach for Drug Discovery. Int. J. Mol. Sci. 2022, 23, 9223. https://doi.org/10.3390/ijms23169223
Telang NT. Stem Cell Models for Breast and Colon Cancer: Experimental Approach for Drug Discovery. International Journal of Molecular Sciences. 2022; 23(16):9223. https://doi.org/10.3390/ijms23169223
Chicago/Turabian StyleTelang, Nitin T. 2022. "Stem Cell Models for Breast and Colon Cancer: Experimental Approach for Drug Discovery" International Journal of Molecular Sciences 23, no. 16: 9223. https://doi.org/10.3390/ijms23169223
APA StyleTelang, N. T. (2022). Stem Cell Models for Breast and Colon Cancer: Experimental Approach for Drug Discovery. International Journal of Molecular Sciences, 23(16), 9223. https://doi.org/10.3390/ijms23169223