
Rescue of Mitochondrial SIRT3 Ameliorates Ischemia-like Injury in Human Endothelial Cells

Abstract
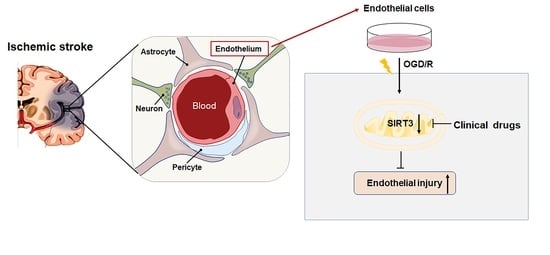
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
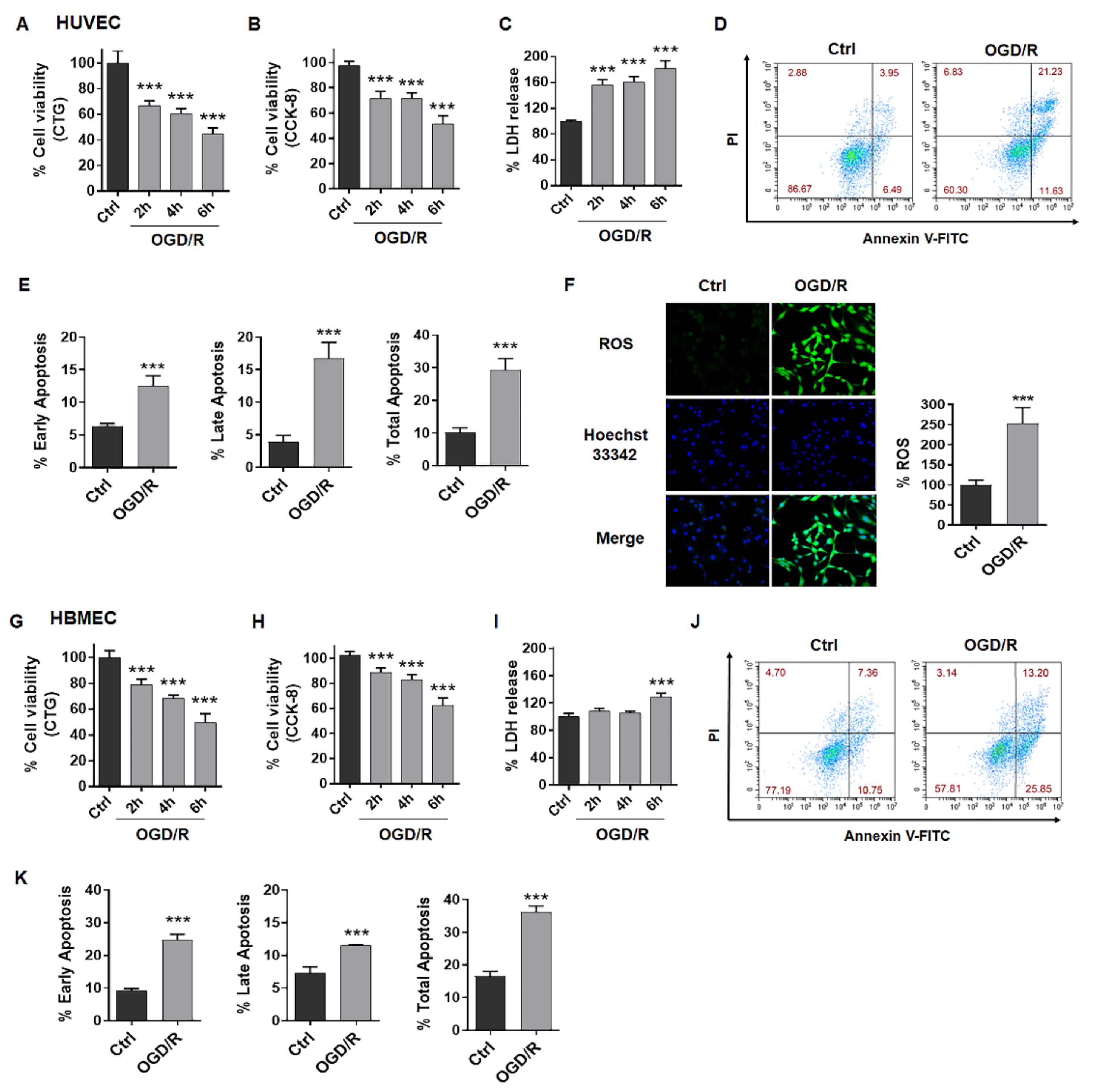
2.1. OGD/R Induces Injury in ECs
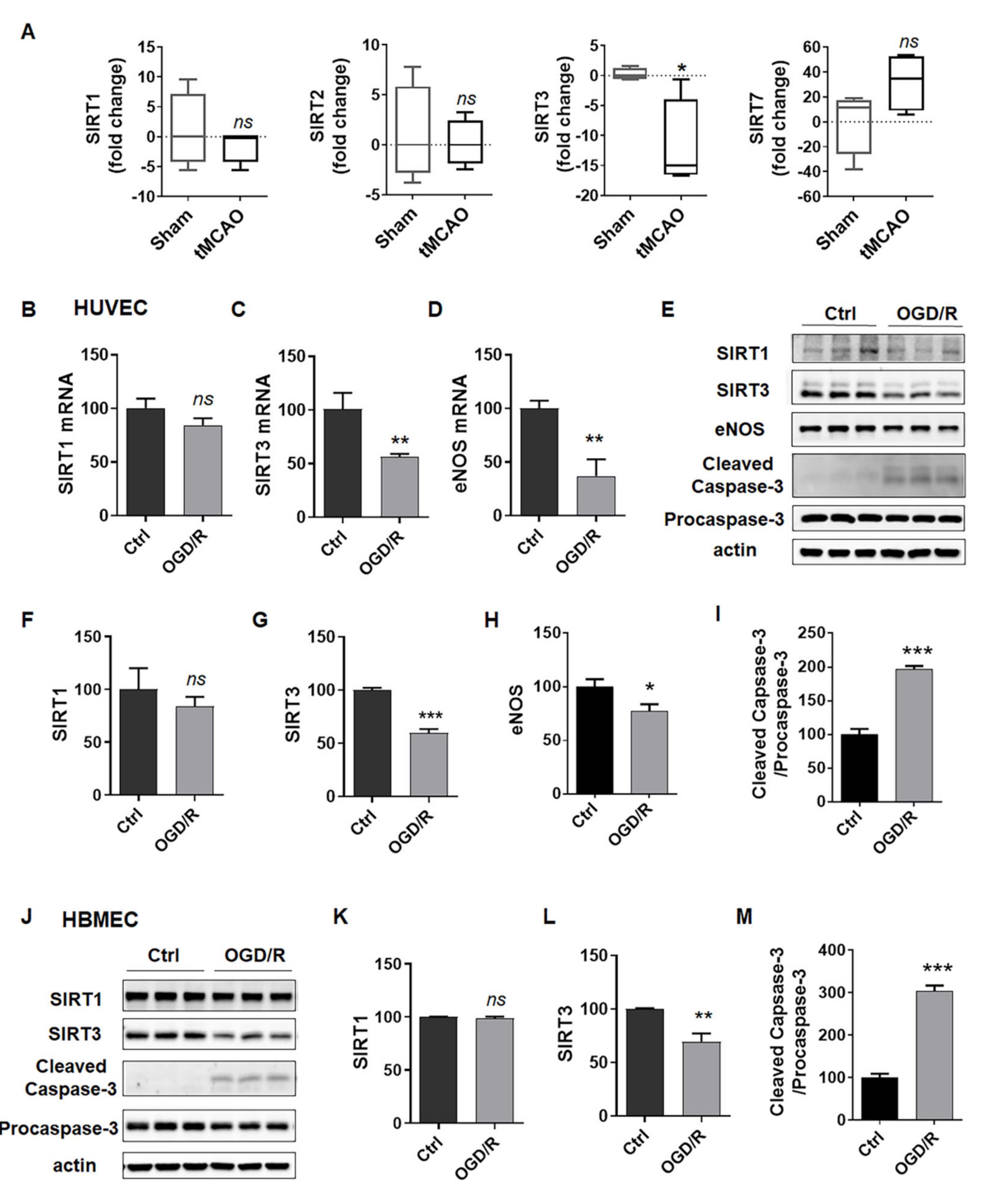
2.2. SIRT3 Expression Is Reduced in ECs after Exposure to OGD/R
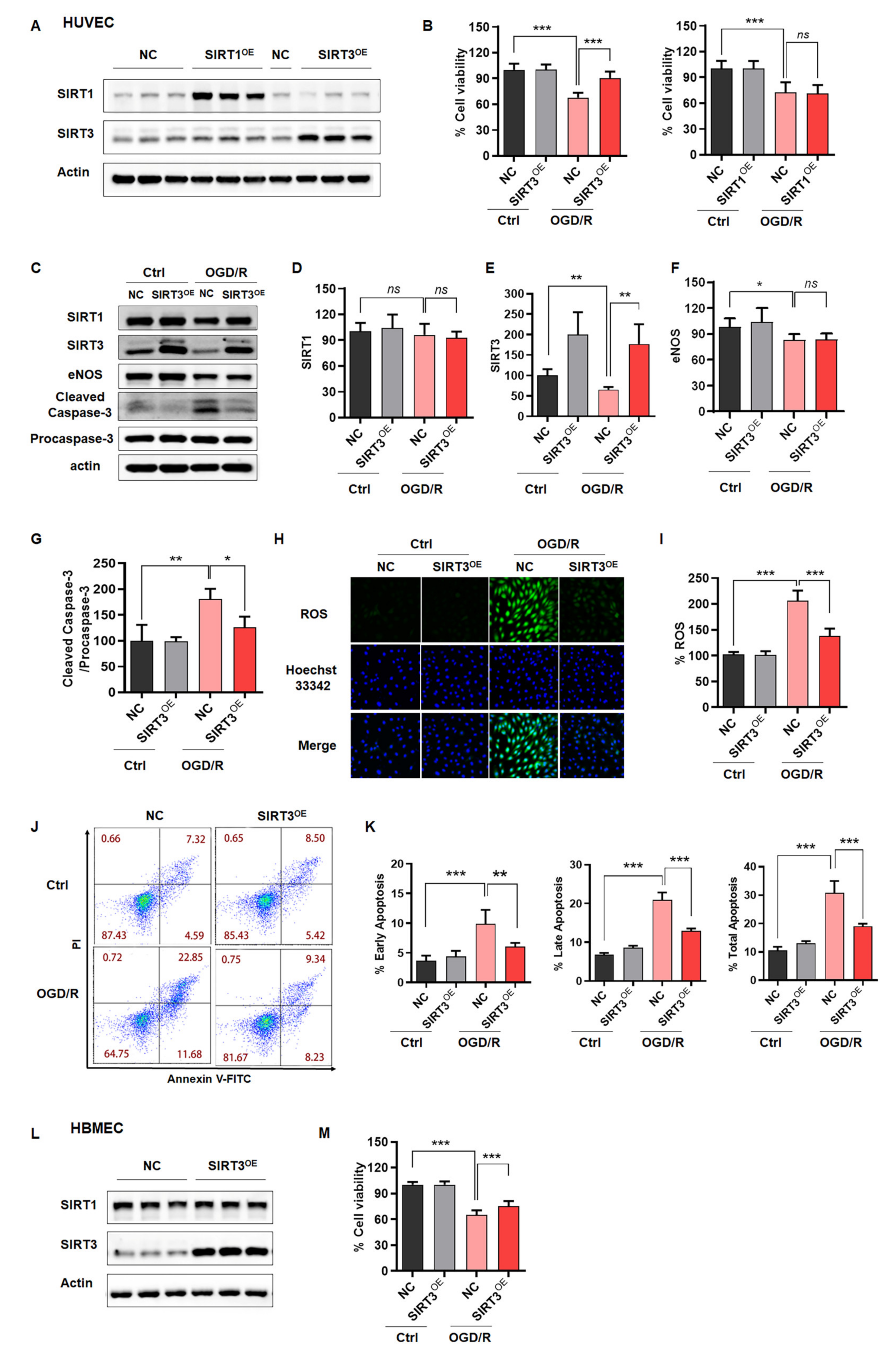
2.3. Rescue of SIRT3 Expression Reverses ODG/R-Induced Cell Damage in ECs
2.4. Clinical Drugs Rescue SIRT3 to Reduce OGD/R-Induced Injury
3. Discussion
4. Materials and Methods
4.1. Drugs and Reagents
4.2. Animals
4.3. Cell Culture
4.4. OGD/R-Induced Cell Injury Model
4.5. H2O2-Induced Cell Injury Model
4.6. Cell Viability
4.7. Lactate Dehydrogenase (LDH) Release Assay
4.8. Cell Apoptosis Assay with Flow Cytometry
4.9. ROS Detection
4.10. Plasmids and Lentiviral Constructs and Infection
4.11. Reverse Transcription and Quantitative Real-Time PCR
4.12. Western Blotting
4.13. Liquid Chromatography/Mass Spectrometry (LC/MS)
4.14. Platelet Aggregation Assay
4.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Acronyms
| ECs | Endothelial cells |
| OGD/R | Oxygen and glucose deprived/reperfusion |
| SIRT1/2/3/7 | Sirtuin 1/2/3/7 |
| NBP | Dl-3-n-butylphthalide |
| OHNBP | 10-hydroxy-NBP |
| rtPA | Recombinant tissue plasminogen activator |
| Clo | Clopidogrel |
| Asp | Aspirin |
| ED | Edaravone |
References
- Rabinstein, A.A. Update on treatment of acute ischemic stroke. Continuum 2020, 26, 268–286. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ma, F.; Huang, L.; Zhang, Y.; Peng, Y.; Xing, C.; Feng, Y.; Wang, X.; Peng, Y. Dl-3-n-butylphthalide (nbp): A promising therapeutic agent for ischemic stroke. CNS Neurol. Disord. Drug Targets 2018, 17, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Pan, Y.; Zhao, X.; Li, H.; Wang, D.; Johnston, S.C.; Liu, L.; Meng, X.; Wang, A.; Wang, C.; et al. Clopidogrel with aspirin in acute minor stroke or transient ischemic attack (chance) trial: One-year outcomes. Circulation 2015, 132, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Paciaroni, M.; Ince, B.; Hu, B.; Jeng, J.S.; Kutluk, K.; Liu, L.; Lou, M.; Parfenov, V.; Wong, K.S.L.; Zamani, B.; et al. Benefits and risks of clopidogrel vs. Aspirin monotherapy after recent ischemic stroke: A systematic review and meta-analysis. Cardiovasc. Ther. 2019, 2019, 1607181. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, Y.; Wang, A.; Gao, Z.; Gao, X.; Chen, H.; Zhou, J.; Zhao, X.; Wang, Y. Safety and efficacy of edaravone dexborneol versus edaravone for patients with acute ischaemic stroke: A phase ii, multicentre, randomised, double-blind, multiple-dose, active-controlled clinical trial. Stroke Vasc. Neurol. 2019, 4, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.Z.; Fan, C.X.; Zhang, Z.L.; Zhao, X.; Sun, Y.; Liu, H.H.; Nie, Z.X.; Pu, X.P. Effects of dl-3-n-butylphthalide on cerebral ischemia infarction in rat model by mass spectrometry imaging. Int. J. Mol. Sci. 2017, 18, 2451. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, R.; Fu, D.; Wu, H.; Zhao, X.; Sun, Y.; Wang, M.; Pu, X. Dl-3-n-butylphthalide inhibits neuroinflammation by stimulating foxp3 and ki-67 in an ischemic stroke model. Aging 2021, 13, 3763–3778. [Google Scholar] [CrossRef] [PubMed]
- Yousufuddin, M.; Young, N. Aging and ischemic stroke. Aging 2019, 11, 2542–2544. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; De Silva, T.M.; Chen, J.; Faraci, F.M. Cerebral vascular disease and neurovascular injury in ischemic stroke. Circ. Res. 2017, 120, 449–471. [Google Scholar] [CrossRef]
- Bonnin, P.; Mazighi, M.; Charriaut-Marlangue, C.; Kubis, N. Early collateral recruitment after stroke in infants and adults. Stroke 2019, 50, 2604–2611. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, Q.; Anrather, J.; Shi, F.D. Immune interventions in stroke. Nat. Rev. Neurol. 2015, 11, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Zhang, K.; Li, P.; Zhu, L.; Xu, J.; Yang, B.; Hu, X.; Lu, Z.; Chen, J. Dysfunction of the neurovascular unit in ischemic stroke and neurodegenerative diseases: An aging effect. Ageing Res. Rev. 2017, 34, 77–87. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, E.; Peterson, B.S.; Bomze, H.M.; Hirschey, M.D. Sirt3 regulates progression and development of diseases of aging. Trends Endocrinol. Metab. TEM 2015, 26, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Satoh, A.; Imai, S.I.; Guarente, L. The brain, sirtuins, and ageing. Nat. Rev. Neurosci. 2017, 18, 362–374. [Google Scholar] [CrossRef]
- Parodi-Rullán, R.M.; Chapa-Dubocq, X.; Rullán, P.J.; Jang, S.; Javadov, S. High sensitivity of sirt3 deficient hearts to ischemia-reperfusion is associated with mitochondrial abnormalities. Front. Pharmacol. 2017, 8, 275. [Google Scholar] [CrossRef] [PubMed]
- Porter, G.A.; Urciuoli, W.R.; Brookes, P.S.; Nadtochiy, S.M. Sirt3 deficiency exacerbates ischemia-reperfusion injury: Implication for aged hearts. Am. J. Physiol.. Heart Circ. Physiol. 2014, 306, H1602–H1609. [Google Scholar] [CrossRef] [PubMed]
- Koentges, C.; Pfeil, K.; Meyer-Steenbuck, M.; Lother, A.; Hoffmann, M.M.; Odening, K.E.; Hein, L.; Bode, C.; Bugger, H. Preserved recovery of cardiac function following ischemia-reperfusion in mice lacking sirt3. Can. J. Physiol. Pharmacol. 2016, 94, 72–80. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zeng, H.; Chen, J.X. Ablation of sirt3 causes coronary microvascular dysfunction and impairs cardiac recovery post myocardial ischemia. Int. J. Cardiol. 2016, 215, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Klishadi, M.S.; Zarei, F.; Hejazian, S.H.; Moradi, A.; Hemati, M.; Safari, F. Losartan protects the heart against ischemia reperfusion injury: Sirtuin3 involvement. J. Pharm. Pharm. Sci. A Publ. Can. Soc. Pharm. Sci. Soc. Can. Des Sci. Pharm. 2015, 18, 112–123. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Y.; Geng, K.; Yang, K.; Shao, J.; Xia, W. Sirt3 protects against ischemic stroke injury by regulating hif-1α/vegf signaling and blood-brain barrier integrity. Cell. Mol. Neurobiol. 2021, 41, 1203–1215. [Google Scholar] [CrossRef]
- Li, Y.; Hu, K.; Liang, M.; Yan, Q.; Huang, M.; Jin, L.; Chen, Y.; Yang, X.; Li, X. Stilbene glycoside upregulates sirt3/ampk to promotes neuronal mitochondrial autophagy and inhibit apoptosis in ischemic stroke. Adv. Clin. Exp. Med. Off. Organ Wroc. Med. Univ. 2021, 30, 139–146. [Google Scholar] [CrossRef]
- Zhao, L.; Cao, J.; Hu, K.; He, X.; Yun, D.; Tong, T.; Han, L. Sirtuins and their biological relevance in aging and age-related diseases. Aging Dis. 2020, 11, 927–945. [Google Scholar] [CrossRef]
- Wang, I.M.; Zhang, B.; Yang, X.; Zhu, J.; Stepaniants, S.; Zhang, C.; Meng, Q.; Peters, M.; He, Y.; Ni, C.; et al. Systems analysis of eleven rodent disease models reveals an inflammatome signature and key drivers. Mol. Syst. Biol. 2012, 8, 594. [Google Scholar] [CrossRef] [PubMed]
- Diao, X.; Deng, P.; Xie, C.; Li, X.; Zhong, D.; Zhang, Y.; Chen, X. Metabolism and pharmacokinetics of 3-n-butylphthalide (nbp) in humans: The role of cytochrome p450s and alcohol dehydrogenase in biotransformation. Drug Metab. Dispos. Biol. Fate Chem. 2013, 41, 430–444. [Google Scholar] [CrossRef] [PubMed]
- Diao, X.; Ma, Z.; Wang, H.; Zhong, D.; Zhang, Y.; Jin, J.; Fan, Y.; Chen, X. Simultaneous quantitation of 3-n-butylphthalide (nbp) and its four major metabolites in human plasma by lc-ms/ms using deuterated internal standards. J. Pharm. Biomed. Anal. 2013, 78–79, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Diao, X.X.; Zhong, K.; Li, X.L.; Zhong, D.F.; Chen, X.Y. Isomer-selective distribution of 3-n-butylphthalide (nbp) hydroxylated metabolites, 3-hydroxy-nbp and 10-hydroxy-nbp, across the rat blood-brain barrier. Acta Pharmacol. Sin. 2015, 36, 1520–1527. [Google Scholar] [CrossRef]
- Chouchani, E.T.; Pell, V.R.; James, A.M.; Work, L.M.; Saeb-Parsy, K.; Frezza, C.; Krieg, T.; Murphy, M.P. A unifying mechanism for mitochondrial superoxide production during ischemia-reperfusion injury. Cell Metab. 2016, 23, 254–263. [Google Scholar]
- Zhang, J.; Xiang, H.; Liu, J.; Chen, Y.; He, R.R.; Liu, B. Mitochondrial sirtuin 3: New emerging biological function and therapeutic target. Theranostics 2020, 10, 8315–8342. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Li, Y.; Zhang, Z.; Lu, J.; Pei, G.; Huang, S. Rescue of Mitochondrial SIRT3 Ameliorates Ischemia-like Injury in Human Endothelial Cells. Int. J. Mol. Sci. 2022, 23, 9118. https://doi.org/10.3390/ijms23169118
Liu X, Li Y, Zhang Z, Lu J, Pei G, Huang S. Rescue of Mitochondrial SIRT3 Ameliorates Ischemia-like Injury in Human Endothelial Cells. International Journal of Molecular Sciences. 2022; 23(16):9118. https://doi.org/10.3390/ijms23169118
Chicago/Turabian StyleLiu, Xi, Yi Li, Zhen Zhang, Juan Lu, Gang Pei, and Shichao Huang. 2022. "Rescue of Mitochondrial SIRT3 Ameliorates Ischemia-like Injury in Human Endothelial Cells" International Journal of Molecular Sciences 23, no. 16: 9118. https://doi.org/10.3390/ijms23169118
APA StyleLiu, X., Li, Y., Zhang, Z., Lu, J., Pei, G., & Huang, S. (2022). Rescue of Mitochondrial SIRT3 Ameliorates Ischemia-like Injury in Human Endothelial Cells. International Journal of Molecular Sciences, 23(16), 9118. https://doi.org/10.3390/ijms23169118