
Role of MicroRNA-7 (MiR-7) in Cancer Physiopathology


Abstract

1. Introduction
1.1. Emergence of MiRNAs
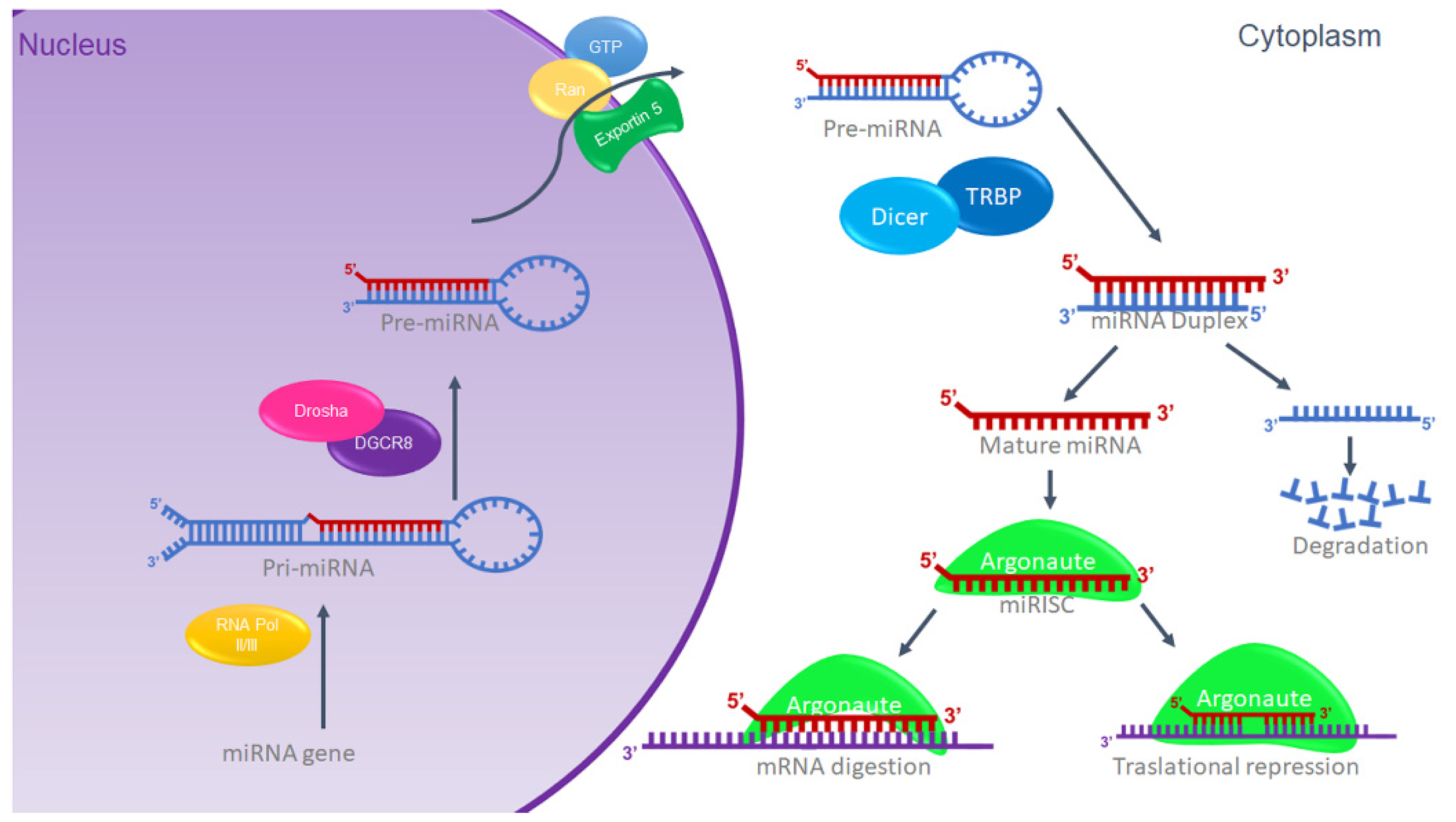
1.2. MiRNAs Biogenesis
1.3. MiRNAs in Cancer
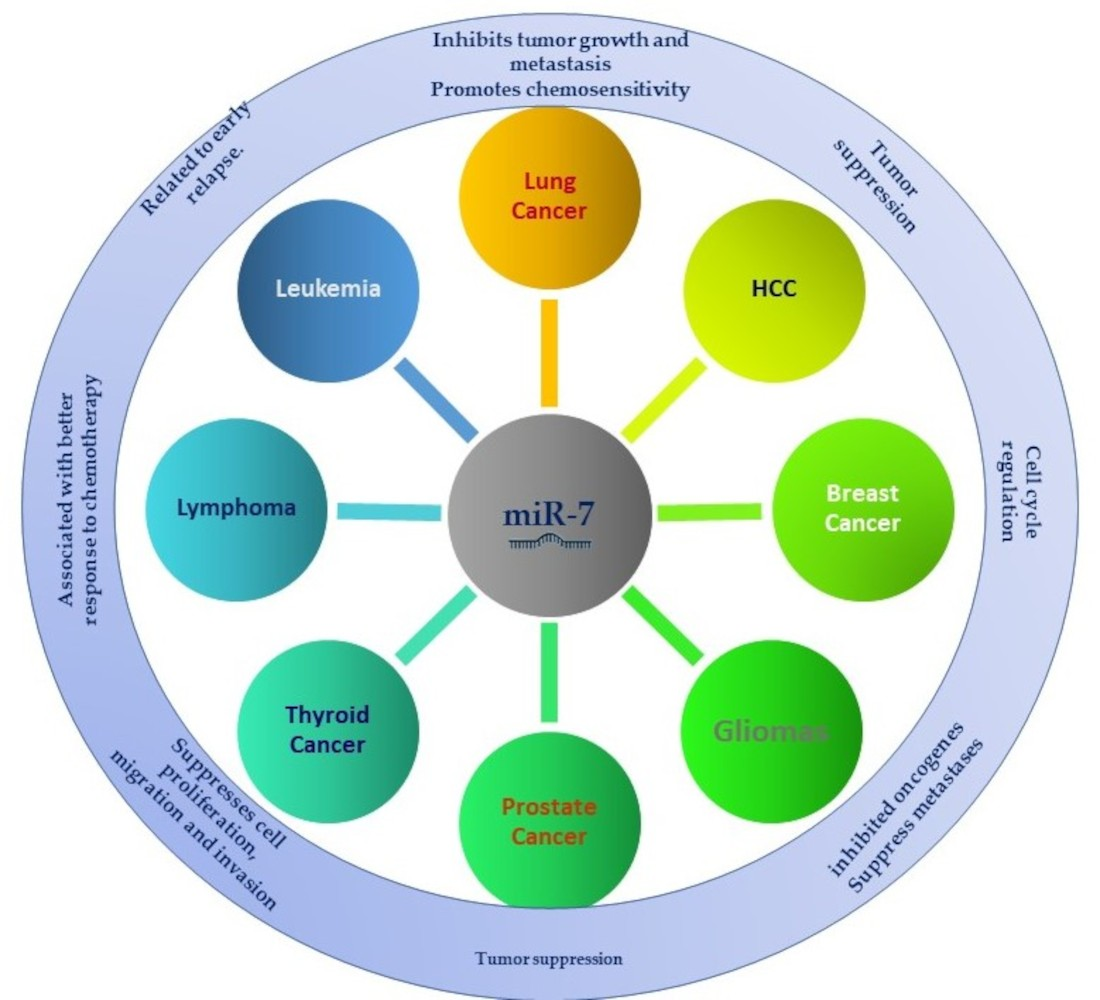
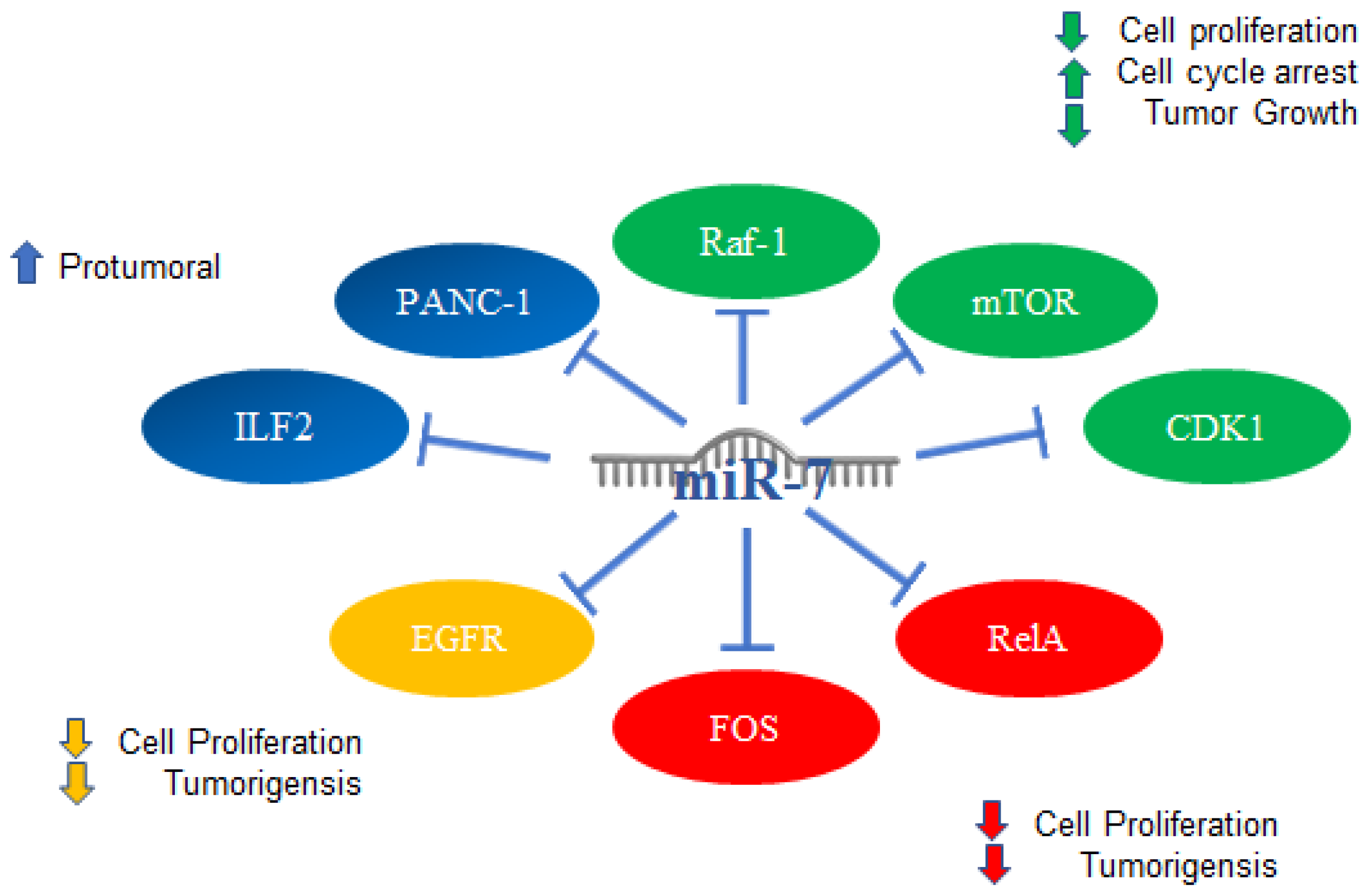
2. MiR-7 in Cancer
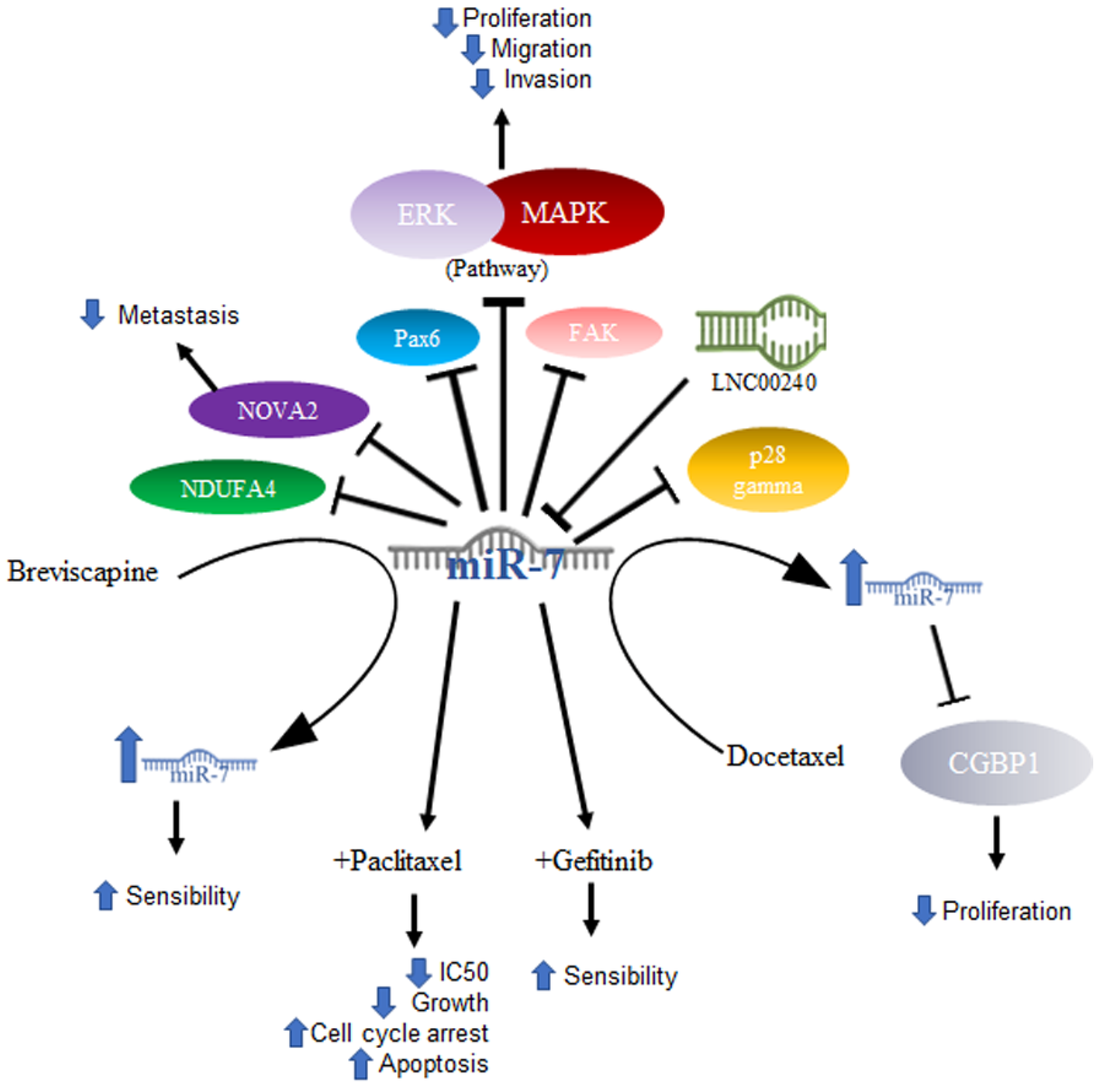
2.1. MiR-7 in Lung Cancer
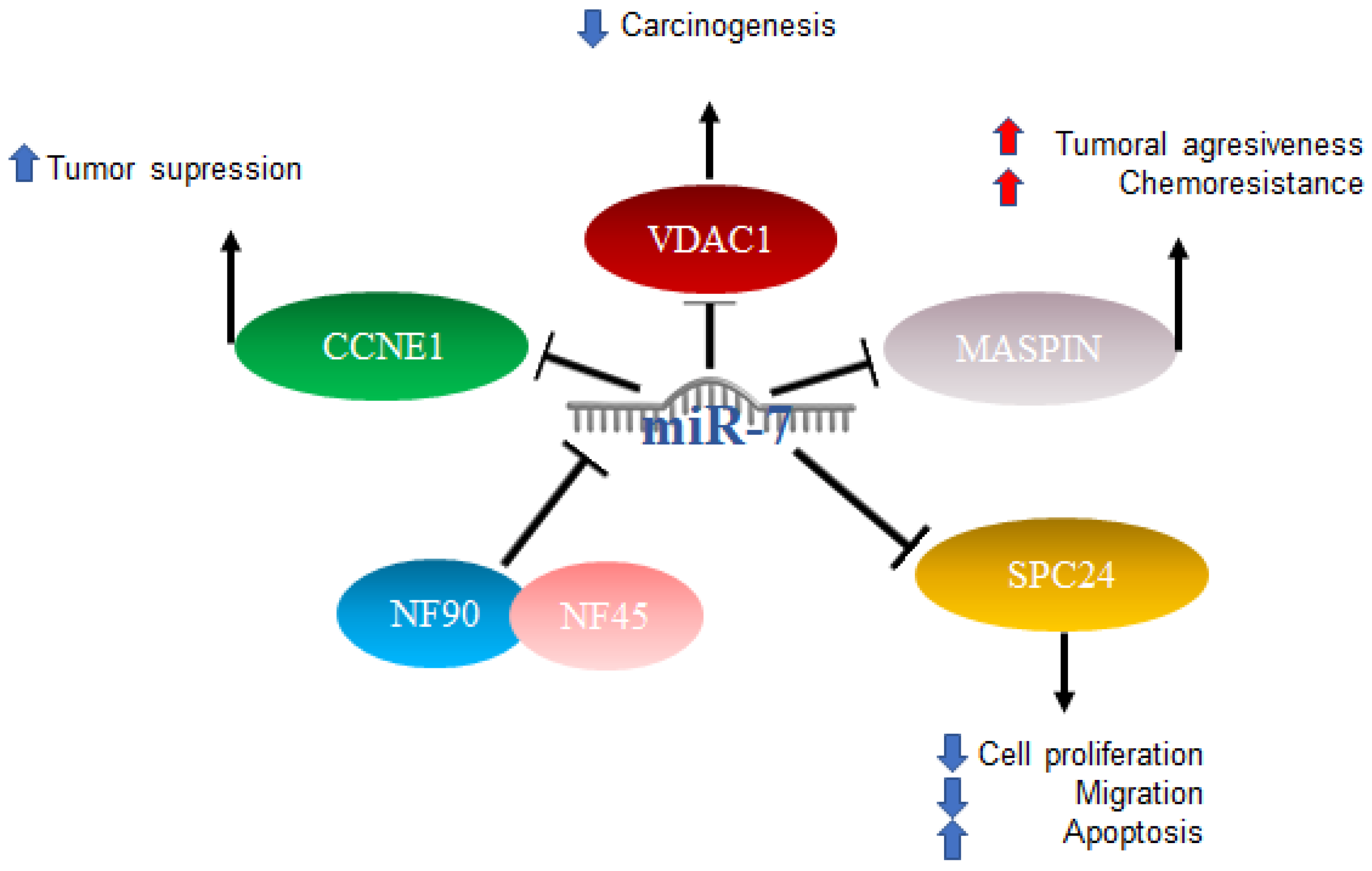
2.2. MiR-7 in Hepatocellular Cancer
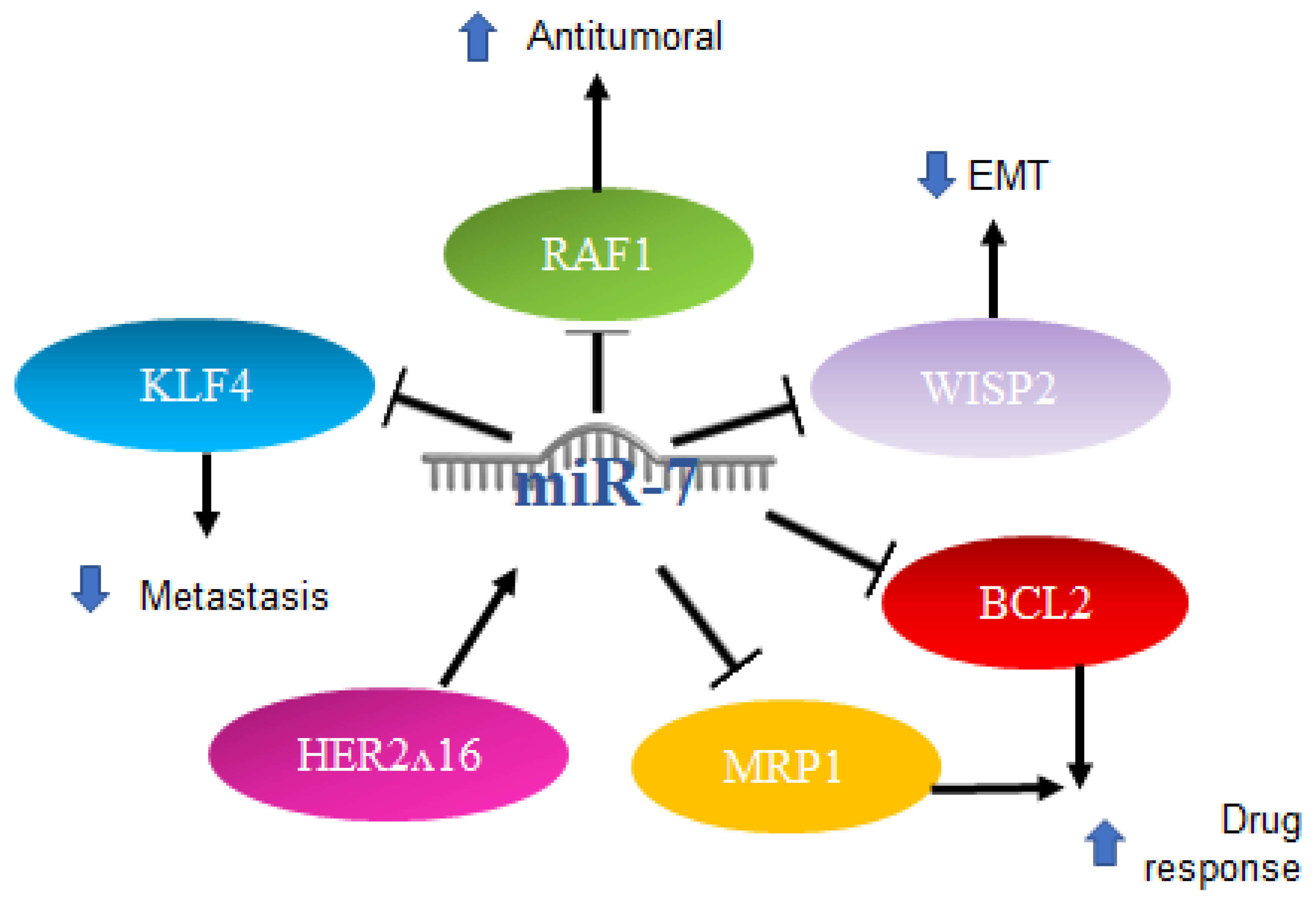
2.3. MiR-7 in Breast Cancer
2.4. Role of MiR-7 in Gliomas
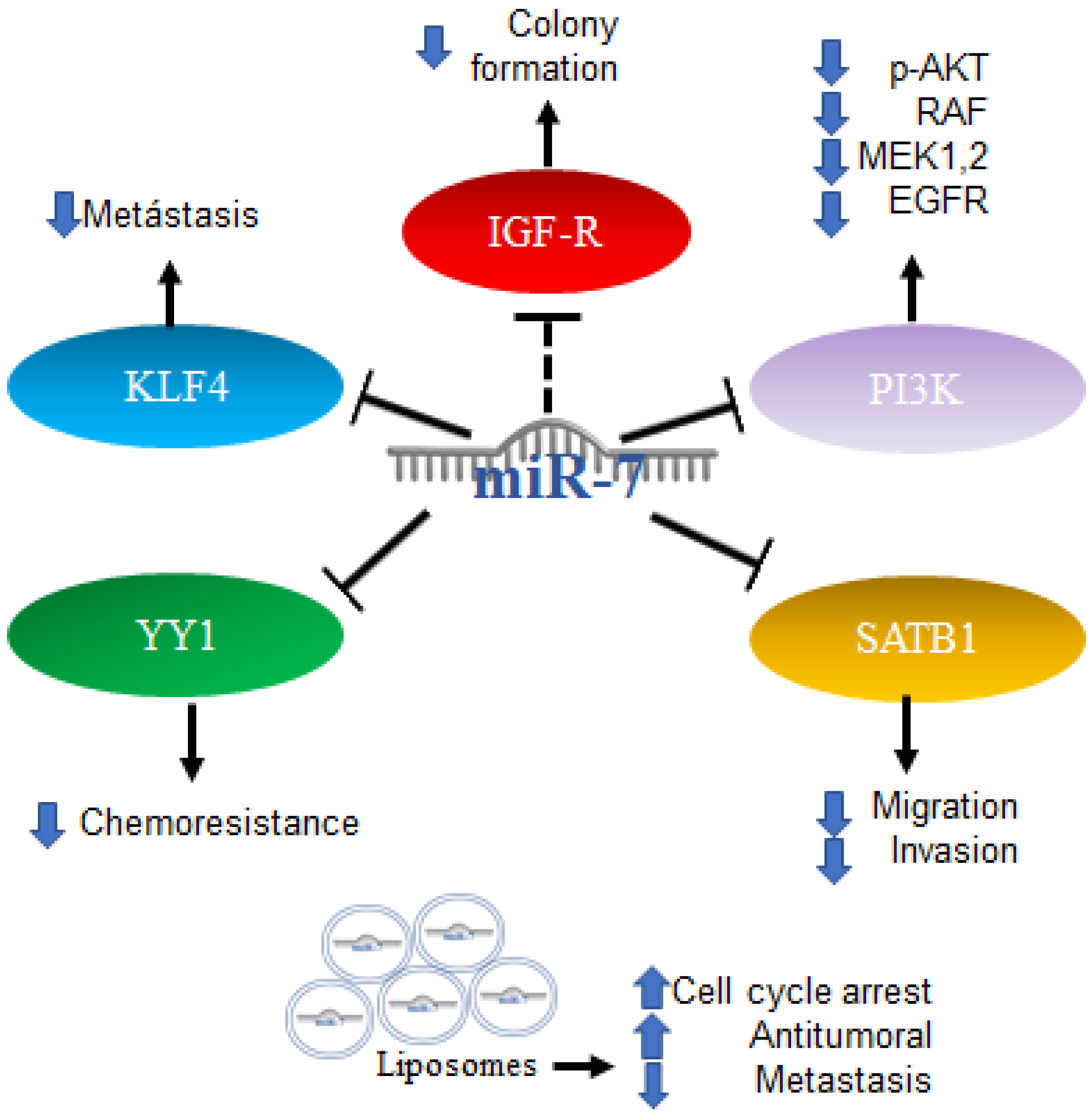
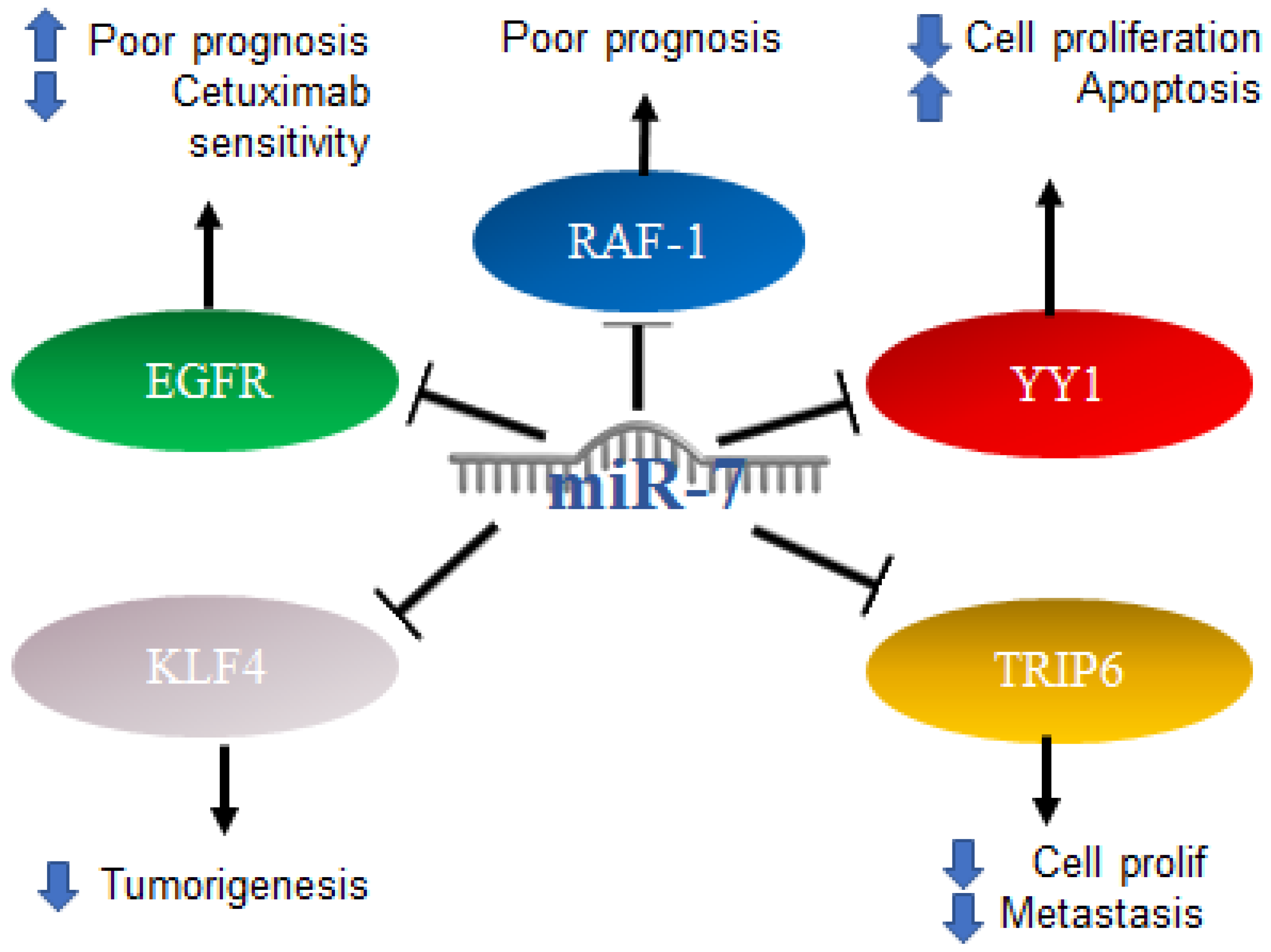
2.5. Role of MiR-7 in Colorectal Cancer
2.6. Role of MiR-7 in Prostate Cancer
2.7. MiR-7 in Oral Cancer
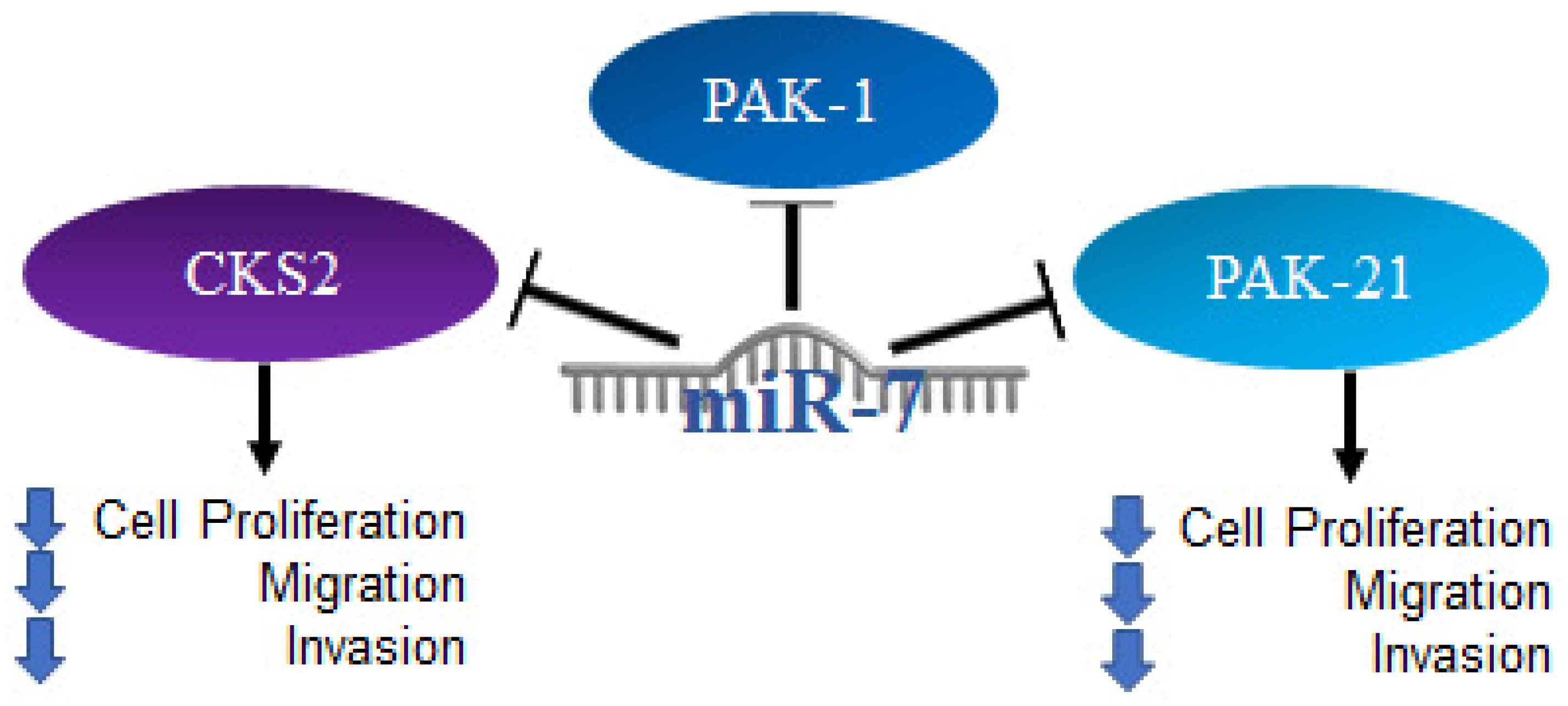
2.8. MiR-7 in Thyroid Cancer
2.9. Role of MiR-7 in Melanoma
2.10. MiR-7 in Cervical Cancer
2.11. Role of MiR-7 in Other Types of Cancer
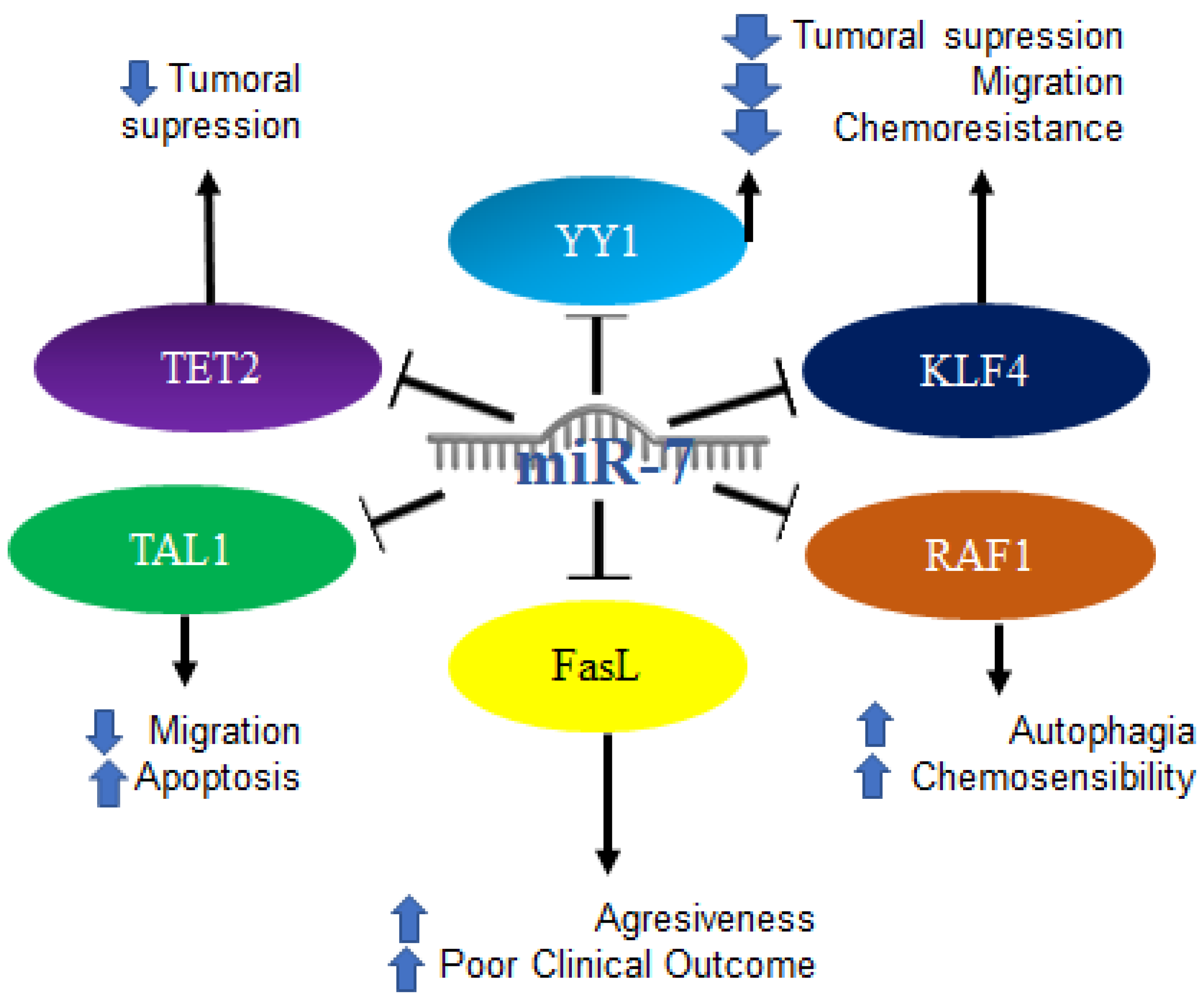
2.12. MiR-7 in Hematologic Malignancies
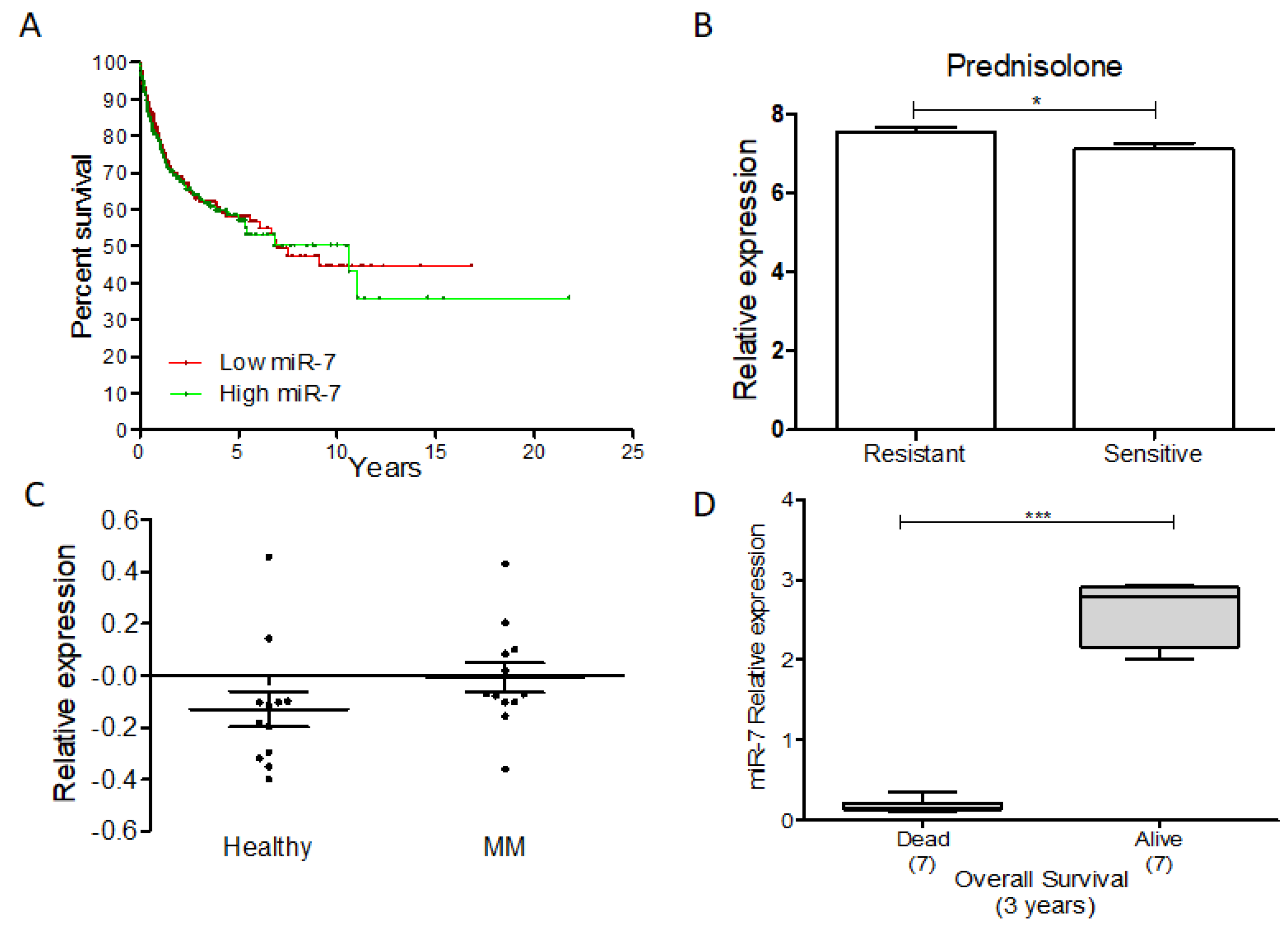
3. MiR-7 and Its Clinical and Therapeutic Potential in Cancer
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations:
| BVP | Breviscapine |
| CAGRs | Cancer Associates Genomics Regions |
| CLL | Chronic Lymphocytic Leukemia |
| CRC | Colorectal Cancer |
| CTL | Cytotoxic T Lymphocytes |
| DNA | Desoxyribonucleic Acid |
| GEO | Gene Expression Omnibus |
| miR, miRNA | microRNA |
| NHL | Non-Hodgkin Lymphoma |
| NSCLC | Non-small Cell Lung Cancer |
| PCR | Polymerase Chain Reaction |
| Pre-mRNA | Precursor microRNA |
| Pri-miRNA | Primary microRNA |
| PTX | Paclitaxel |
| RISC | RNA-interference Silence Complex |
| RNA | Ribonucleic Acid |
| UTR | Untranslated Region |
| WHO | World Health Organization |
References
- OMS. “Cáncer,” Fact Sheets, 2017. Available online: http://www.who.int/mediacentre/factsheets/fs297/es/ (accessed on 6 December 2017).
- Reckel, S.; Hantschel, O. Bcr-Abl: One kinase, two isoforms, two diseases. Oncotarget 2017, 8, 78257–78258. [Google Scholar] [CrossRef]
- Koeffler, H.P.; McCormick, F.; Denny, C. Molecular mechanisms of cancer. West. J. Med. 1991, 155, 505–514. [Google Scholar] [PubMed]
- Rivlin, N.; Brosh, R.; Oren, M.; Rotter, V. Mutations in the p53 Tumor Suppressor Gene: Important Milestones at the Various Steps of Tumorigenesis. Genes Cancer 2011, 2, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Garzon, R.; Fabbri, M.; Cimmino, A.; Calin, G.A.; Croce, C.M. MicroRNA expression and function in cancer. Trends Mol. Med. 2006, 12, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Ambros, V. microRNAs: Tiny Regulators with Great Potential. Cell 2001, 107, 823–826. [Google Scholar] [CrossRef]
- Lee, R.C.; Ambros, V. An extensive class of small RNAs in Caenorhabditis elegans. Science 2001, 294, 862–864. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of Novel Genes Coding for Small Expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef]
- MLagos-Quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of Tissue-Specific MicroRNAs from Mouse. Curr. Biol. 2002, 12, 735–739. [Google Scholar] [CrossRef]
- Mourelatos, Z.; Dostie, J.; Paushkin, S.; Sharma, A.; Charroux, B.; Abel, L.; Rappsilber, J.; Mann, M.; Dreyfuss, G. miRNPs: A novel class of ribonucleoproteins containing numerous microRNAs. Genes Dev. 2002, 16, 720–728. [Google Scholar] [CrossRef] [PubMed]
- Dostie, J.; Mourelatos, Z.; Yang, M.; Sharma, A.; Dreyfuss, G. Numerous microRNPs in neuronal cells containing novel microRNAs. RNA 2003, 9, 180–186. [Google Scholar] [CrossRef]
- Ambros, V. MicroRNA Pathways in Flies and Worms: Growth, Death, Fat, Stress, and Timing. Cell 2003, 113, 673–676. [Google Scholar] [CrossRef]
- Aravin, A.A.; Lagos-Quintana, M.; Yalcin, A.; Zavolan, M.; Marks, D.; Snyder, B.; Gaasterland, T.; Meyer, J.; Tuschl, T. The Small RNA Profile during Drosophila melanogaster Development. Dev. Cell 2003, 5, 337–350. [Google Scholar] [CrossRef]
- Lim, L.P.; Glasner, M.E.; Yekta, S.; Burge, C.B.; Bartel, D.P. Vertebrate MicroRNA Genes. Science 2003, 299, 1540. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Rådmark, O.; Kim, S.; et al. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Denli, A.M.; Tops, B.; Plasterk, R.H.A.; Ketting, R.F.; Hannon, G.J. Processing of primary microRNAs by the Microprocessor complex. Nature 2004, 432, 231–235. [Google Scholar] [CrossRef]
- Basyuk, E.; Suavet, F.; Doglio, A.; Bordonné, R.; Bertrand, E. Human let-7 stem-loop precursors harbor features of RNase III cleavage products. Nucleic Acids Res. 2003, 31, 6593–6597. [Google Scholar] [CrossRef]
- Chendrimada, T.P.; Gregory, R.I.; Kumaraswamy, E.; Norman, J.; Cooch, N.; Nishikura, K.; Shiekhattar, R. TRBP recruits the Dicer complex to Ago2 for microRNA processing and gene silencing. Nature 2005, 436, 740–744. [Google Scholar] [CrossRef]
- Bernstein, E.; Caudy, A.A.; Hammond, S.M.; Hannon, G.J. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 2001, 409, 363–366. [Google Scholar] [CrossRef]
- Hammond, S.M.; Bernstein, E.; Beach, D.; Hannon, G.J. An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells. Nature 2000, 404, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Hutvágner, G.; Mclachlan, J.; Pasquinelli, A.E.; Bálint, É.; Tuschl, T.; Zamore, P.D. A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNA. Science 2001, 293, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Huang, X.; Cui, H.; Luo, X.; Tang, Y.; Chen, S.; Wu, L.; Shen, N. miR-155 and its star-form partner miR-155* cooperatively regulate type I interferon production by human plasmacytoid dendritic cells. Blood 2010, 116, 5885–5894. [Google Scholar] [CrossRef] [PubMed]
- Elbashir, S.M.; Lendeckel, W.; Tuschl, T. RNA interference is mediated by 21- and 22-nucleotide RNAs. Genes Dev. 2001, 15, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Hutvagner, G.; Zamore, P.D. A microRNA in a Multiple-Turnover RNAi Enzyme Complex. Science 2002, 297, 2056–2060. [Google Scholar] [CrossRef]
- Tijsterman, M.; Plasterk, R.H. Dicers at RISC. Cell 2004, 117, 1–3. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Kim, B.; Kim, V.N. Re-evaluation of the roles of DROSHA, Exportin 5, and DICER in microRNA biogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, E1881–E1889. [Google Scholar] [CrossRef] [PubMed]
- Alles, J.; Fehlmann, T.; Fischer, U.; Backes, C.; Galata, V.; Minet, M.; Hart, M.; Abu-Halima, M.; Grässer, F.A.; Lenhof, H.-P.; et al. An estimate of the total number of true human miRNAs. Nucleic Acids Res. 2019, 47, 3353–3364. [Google Scholar] [CrossRef]
- Hao, Z.; Yang, J.; Wang, C.; Li, Y.; Zhang, Y.; Dong, X.; Zhou, L.; Liu, J.; Zhang, Y.; Qian, J. MicroRNA-7 inhibits metastasis and invasion through targeting focal adhesion kinase in cervical cancer. Int. J. Clin. Exp. Med. 2015, 8, 480–487. [Google Scholar]
- Wang, D.; Gu, J.; Wang, T.; Ding, Z. OncomiRDB: A database for the experimentally verified oncogenic and tumor-suppressive microRNAs. Bioinformatics 2014, 30, 2237–2238. [Google Scholar] [CrossRef]
- Weber, B.; Stresemann, C.; Brueckner, B.; Lyko, F. Methylation of Human MicroRNA Genes in Normal and Neoplastic Cells. Cell Cycle 2007, 6, 1001–1005. [Google Scholar] [CrossRef] [PubMed]
- Agirre, X.; A Martínez-Climent, J.; Odero, M.D.; Prósper, F. Epigenetic regulation of miRNA genes in acute leukemia. Leukemia 2011, 26, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Chim, C.S.; Wong, K.Y.; Qi, Y.; Loong, F.; Lam, W.L.; Wong, L.G.; Jin, D.Y.; Costello, J.F.; Liang, R. Epigenetic inactivation of the miR-34a in hematological malignancies. Carcinogenesis 2010, 31, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Stumpel, D.J.P.M.; Schotte, D.; Lange-Turenhout, E.A.M.; A Schneider, P.; Seslija, L.; De Menezes, R.X.; E Marquez, V.; Pieters, R.; Boer, M.D.; Stam, R.W. Hypermethylation of specific microRNA genes in MLL-rearranged infant acute lymphoblastic leukemia: Major matters at a micro scale. Leukemia 2010, 25, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef]
- Ozsolak, F.; Poling, L.L.; Wang, Z.; Liu, H.; Liu, X.S.; Roeder, R.G.; Zhang, X.; Song, J.S.; Fisher, D.E. Chromatin structure analyses identify miRNA promoters. Genes Dev. 2008, 22, 3172–3183. [Google Scholar] [CrossRef] [PubMed]
- Monteys, A.M.; Spengler, R.M.; Wan, J.; Tecedor, L.; Lennox, K.A.; Xing, Y.; Davidson, B.L. Structure and activity of putative intronic miRNA promoters. RNA 2010, 16, 495–505. [Google Scholar] [CrossRef]
- Galasso, M.; Sandhu, S.; Volinia, S. MicroRNA Expression Signatures in Solid Malignancies. Cancer J. 2012, 18, 238–243. [Google Scholar] [CrossRef]
- He, L.; Thomson, J.M.; Hemann, M.T.; Hernando-Monge, E.; Mu, D.; Goodson, S.; Powers, S.; Cordon-Cardo, C.; Lowe, S.W.; Hannon, G.J.; et al. A microRNA polycistron as a potential human oncogene. Nature 2005, 435, 828–833. [Google Scholar] [CrossRef]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef]
- Basso, K.; Sumazin, P.; Morozov, P.; Schneider, C.; Maute, R.L.; Kitagawa, Y.; Mandelbaum, J.; Haddad, J.; Chen, C.-Z.; Califano, A.; et al. Identification of the Human Mature B Cell miRNome. Immunity 2009, 30, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Hummel, R.; Hussey, D.J.; Haier, J. MicroRNAs: Predictors and modifiers of chemo- and radiotherapy in different tumour types. Eur. J. Cancer 2010, 46, 298–311. [Google Scholar] [CrossRef]
- Takeshita, F.; Patrawala, L.; Osaki, M.; Takahashi, R.-U.; Yamamoto, Y.; Kosaka, N.; Kawamata, M.; Kelnar, K.; Bader, A.G.; Brown, D.; et al. Systemic Delivery of Synthetic MicroRNA-16 Inhibits the Growth of Metastatic Prostate Tumors via Downregulation of Multiple Cell-cycle Genes. Mol. Ther. 2010, 18, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Wang, V.; Wu, W. MicroRNA-based therapeutics for cancer. BioDrugs 2009, 23, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Ferracin, M.; Liu, C.-G.; Veronese, A.; Spizzo, R.; Sabbioni, S.; Magri, E.; Pedriali, M.; Fabbri, M.; Campiglio, M.; et al. MicroRNA Gene Expression Deregulation in Human Breast Cancer. Cancer Res. 2005, 65, 7065–7070. [Google Scholar] [CrossRef] [PubMed]
- Glover, A.R.; Zhao, J.T.; Gill, A.J.; Weiss, J.; Mugridge, N.; Kim, E.; Feeney, A.L.; Ip, J.C.; Reid, G.; Clarke, S.; et al. microRNA-7 as a tumor suppressor and novel therapeutic for adrenocortical carcinoma. Oncotarget 2015, 6, 36675–36688. [Google Scholar] [CrossRef]
- Kalinowski, F.; Brown, R.A.; Ganda, C.; Giles, K.M.; Epis, M.R.; Horsham, J.; Leedman, P.J. microRNA-7: A tumor suppressor miRNA with therapeutic potential. Int. J. Biochem. Cell Biol. 2014, 54, 312–317. [Google Scholar] [CrossRef]
- Correa-Medina, M.; Bravo-Egana, V.; Rosero, S.; Ricordi, C.; Edlund, H.; Diez, J.; Pastori, R.L. MicroRNA miR-7 is preferentially expressed in endocrine cells of the developing and adult human pancreas. Gene Expr. Patterns 2009, 9, 193–199. [Google Scholar] [CrossRef]
- Reddy, S.D.N.; Ohshiro, K.; Rayala, S.K.; Kumar, R. MicroRNA-7, a Homeobox D10 Target, Inhibits p21-Activated Kinase 1 and Regulates Its Functions. Cancer Res. 2008, 68, 8195–8200. [Google Scholar] [CrossRef]
- McInnes, N.; Sadlon, T.J.; Brown, C.Y.; Pederson, S.; Beyer, M.; Schultze, J.L.; McColl, S.; Goodall, G.J.; Barry, S.C. FOXP3 and FOXP3-regulated microRNAs suppress SATB1 in breast cancer cells. Oncogene 2011, 31, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.-T.; Lin, H.-H.; Lien, Y.-C.; Wang, Y.-H.; Hong, C.-F.; Kao, Y.-R.; Lin, S.-C.; Chang, Y.-C.; Lin, S.-Y.; Chen, S.-J.; et al. EGFR Promotes Lung Tumorigenesis by Activating miR-7 through a Ras/ERK/Myc Pathway That Targets the Ets2 Transcriptional Repressor ERF. Cancer Res. 2010, 70, 8822–8831. [Google Scholar] [CrossRef]
- Wu, H.; Sun, S.; Tu, K.; Gao, Y.; Xie, B.; Krainer, A.; Zhu, J. A Splicing-Independent Function of SF2/ASF in MicroRNA Processing. Mol. Cell 2010, 38, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-J.; Wang, C.-H.; Zhou, Y.; Liao, Z.-Y.; Zhu, S.-F.; Hu, Y.; Chen, C.; Luo, J.-M.; Wen, Z.-K.; Xu, L. TLR9 signaling repressed tumor suppressor miR-7 expression through up-regulation of HuR in human lung cancer cells. Cancer Cell Int. 2013, 13, 90. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wen, Z.; Zhou, Y.; Liu, Z.; Li, Q.; Fei, G.; Luo, J.; Ren, T. MicroRNA-7–regulated TLR9 signaling–enhanced growth and metastatic potential of human lung cancer cells by altering the phosphoinositide-3-kinase, regulatory subunit 3/Akt pathway. Mol. Biol. Cell 2013, 24, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Bhere, D.; Arghiani, N.; Lechtich, E.R.; Yao, Y.; Alsaab, S.; Bei, F.; Matin, M.M.; Shah, K. Simultaneous downregulation of miR-21 and upregulation of miR-7 has anti-tumor efficacy. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- American-Cancer-Society, “American Cancer Society, Cancer facts and figures 2015”. 2015. Available online: www.cancer.org (accessed on 1 June 2022).
- Lei, L.; Chen, C.; Zhao, J.; Wang, H.; Guo, M.; Zhou, Y.; Luo, J.; Zhang, J.; Xu, L. Targeted Expression of miR-7 Operated by TTF-1 Promoter Inhibited the Growth of Human Lung Cancer through the NDUFA4 Pathway. Mol. Ther.-Nucleic Acids 2017, 6, 183–197. [Google Scholar] [CrossRef]
- Zeng, J.; Cai, S. Breviscapine suppresses the growth of non-small cell lung cancer by enhancing microRNA-7 expression. J. Biosci. 2017, 42, 121–129. [Google Scholar] [CrossRef]
- He, X.; Li, C.; Wu, X.; Yang, G. Docetaxel inhibits the proliferation of non-small-cell lung cancer cells via upregulation of microRNA-7 expression. Int. J. Clin. Exp. Pathol. 2015, 8, 9072–9080. [Google Scholar] [PubMed]
- Hu, Y.; Liao, Z.; Chen, C.; Qin, N.; Zheng, J.; Tian, D.; Li, Y.; Zhu, S.; Luo, J.; Xu, L. Over-expressed microRNA-7 inhibits the growth of human lung cancer cells via suppressing CGGBP1 expression. Chin. J. Cell. Mol. Immunol. 2014, 30, 125–130. [Google Scholar]
- Luo, J.; Li, H.; Zhang, C. MicroRNA-7 inhibits the malignant phenotypes of non-small cell lung cancer in vitro by targeting Pax6. Mol. Med. Rep. 2015, 12, 5443–5448. [Google Scholar] [CrossRef]
- Cao, Q.; Mao, Z.-D.; Shi, Y.-J.; Chen, Y.; Sun, Y.; Zhang, Q.; Song, L.; Peng, L.-P. MicroRNA-7 inhibits cell proliferation, migration and invasion in human non-small cell lung cancer cells by targeting FAK through ERK/MAPK signaling pathway. Oncotarget 2016, 7, 77468–77481. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; Zheng, Y.; Jiang, P.; Liu, R.; Liu, X.; Qian, J.; Gu, J.; Chang, L.; Ge, D.; Chu, Y. PA28gamma emerges as a novel functional target of tumour suppressor microRNA-7 in non-small-cell lung cancer. Br. J. Cancer 2013, 110, 353–362. [Google Scholar] [CrossRef]
- Xiao, H. MiR-7-5p suppresses tumor metastasis of non-small cell lung cancer by targeting NOVA2. Cell. Mol. Biol. Lett. 2019, 24, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, K.; Liao, Z.; Li, Y.; Yang, H.; Chen, C.; Zhou, Y.; Tao, Y.; Guo, M.; Ren, T.; et al. Promoter mutation of tumor suppressor microRNA-7 is associated with poor prognosis of lung cancer. Mol. Clin. Oncol. 2015, 3, 1329–1336. [Google Scholar] [CrossRef]
- Liu, R.; Liu, X.; Zheng, Y.; Gu, J.; Xiong, S.; Jiang, P.; Jiang, X.; Huang, E.; Yang, Y.; Ge, D.; et al. MicroRNA-7 sensitizes non-small cell lung cancer cells to paclitaxel. Oncol. Lett. 2014, 8, 2193–2200. [Google Scholar] [CrossRef]
- Zhao, J.-G.; Men, W.-F.; Tang, J. MicroRNA-7 enhances cytotoxicity induced by gefitinib in non-small cell lung cancer via inhibiting the EGFR and IGF1R signalling pathways. Współczesna Onkol. 2015, 3, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Ku, G.W.; Kang, Y.; Yu, S.-L.; Park, J.; Park, S.; Jeong, I.B.; Kang, M.W.; Son, J.W.; Kang, J. LncRNA LINC00240 suppresses invasion and migration in non-small cell lung cancer by sponging miR-7-5p. BMC Cancer 2021, 21, 44. [Google Scholar] [CrossRef] [PubMed]
- American-Cancer-Society. Acerca del Cáncer de Hígado, 2016. Available online: https://www.cancer.org/es/cancer/cancer-de-higado/acerca/que-es-cancer-de-higado.html (accessed on 1 June 2022).
- Fang, Y.; Xue, J.-L.; Shen, Q.; Chen, J.; Tian, L. MicroRNA-7 inhibits tumor growth and metastasis by targeting the phosphoinositide 3-kinase/Akt pathway in hepatocellular carcinoma. Hepatology 2012, 55, 1852–1862. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, T.; Todaka, H.; Sugiyama, Y.; Ono, M.; Tamaki, N.; Hatano, E.; Takezaki, Y.; Hanazaki, K.; Miwa, T.; Lai, S.; et al. Suppression of MicroRNA-7 (miR-7) Biogenesis by Nuclear Factor 90-Nuclear Factor 45 Complex (NF90-NF45) Controls Cell Proliferation in Hepatocellular Carcinoma. J. Biol. Chem. 2016, 291, 21074–21084. [Google Scholar] [CrossRef]
- Wang, F.; Qiang, Y.; Zhu, L.; Jiang, Y.; Wang, Y.; Shao, X.; Yin, L.; Chen, J.; Chen, Z. MicroRNA-7 downregulates the oncogene VDAC1 to influence hepatocellular carcinoma proliferation and metastasis. Tumor Biol. 2016, 37, 10235–10246. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-S.; Yen, C.-J.; Chen, Y.-J.; Chen, J.-Y.; Wang, L.-Y.; Chiu, S.-J.; Shih, W.-L.; Ho, C.-Y.; Wei, T.-T.; Pan, H.-L.; et al. miRNA-7/21/107 contribute to HBx-induced hepatocellular carcinoma progression through suppression of maspin. Oncotarget 2015, 6, 25962–25974. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hu, S.; Zhang, X.; Wang, L.; Zhang, X.; Yan, B.; Zhao, J.; Yang, A.; Zhang, R. MicroRNA-7 arrests cell cycle in G1 phase by directly targeting CCNE1 in human hepatocellular carcinoma cells. Biochem. Biophys. Res. Commun. 2013, 443, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, H.; Zhang, G.; Luo, C.; Zhang, S.; Luo, R.; Deng, B. hsa-miR-7-5p suppresses proliferation, migration and promotes apoptosis in hepatocellular carcinoma cell lines by inhibiting SPC24 expression. Biochem. Biophys. Res. Commun. 2021, 561, 80–87. [Google Scholar] [CrossRef]
- American-Cancer-Society. Que es el Cancer de Seno, 2016. Available online: https://www.cancer.org/es/cancer/cancer-de-seno/acerca/que-es-el-cancer-de-seno.html (accessed on 1 June 2022).
- Hsiao, Y.-C.; Yeh, M.-H.; Chen, Y.-J.; Liu, J.-F.; Tang, C.-H.; Huang, W.-C. Lapatinib increases motility of triple-negative breast cancer cells by decreasing miRNA-7 and inducing Raf-1/MAPK-dependent interleukin-6. Oncotarget 2015, 6, 37965–37978. [Google Scholar] [CrossRef] [PubMed]
- Huynh, F.C.; Jones, F.E. MicroRNA-7 Inhibits Multiple Oncogenic Pathways to Suppress HER2Δ16 Mediated Breast Tumorigenesis and Reverse Trastuzumab Resistance. PLoS ONE 2014, 9, e114419. [Google Scholar] [CrossRef]
- Akalay, I.; Tan, T.Z.; Kumar, P.; Janji, B.; Mami-Chouaib, F.; Charpy, C.; Vielh, P.; Larsen, A.K.; Thiery, J.P.; Sabbah, M.; et al. Targeting WNT1-inducible signaling pathway protein 2 alters human breast cancer cell susceptibility to specific lysis through regulation of KLF-4 and miR-7 expression. Oncogene 2014, 34, 2261–2271. [Google Scholar] [CrossRef] [PubMed]
- Okuda, H.; Xing, F.; Pandey, P.R.; Sharma, S.; Watabe, M.; Pai, S.K.; Mo, Y.-Y.; Iiizumi-Gairani, M.; Hirota, S.; Liu, Y.; et al. miR-7 Suppresses Brain Metastasis of Breast Cancer Stem-Like Cells By Modulating KLF4. Cancer Res. 2013, 73, 1434–1444. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.; Ding, J.; Li, W. miR-7 Reverses Breast Cancer Resistance to Chemotherapy by Targeting MRP1 and BCL2. OncoTargets Ther. 2019, 12, 11097–11105. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.; Mawrin, C.; Scherlach, C.; Skalej, M.; Firsching, R. Gliomas in Adults. Dtsch. Arztebl. Int. 2010, 107, 799–808. [Google Scholar] [CrossRef]
- Wang, B.; Sun, F.; Dong, N.; Sun, Z.; Diao, Y.; Zheng, C.; Sun, J.; Yang, Y.; Jiang, D. MicroRNA-7 directly targets insulin-like growth factor 1 receptor to inhibit cellular growth and glucose metabolism in gliomas. Diagn. Pathol. 2014, 9, 211. [Google Scholar] [CrossRef]
- Koshkin, F.A.; Chistyakov, D.A.; Nikitin, A.G.; Konovalov, A.N.; Potapov, A.; Usachyov, D.Y.; Pitskhelauri, D.I.; Kobyakov, G.L.; Shishkina, L.V.; Chekhonin, V.P. Profile of MicroRNA Expression in Brain Tumors of Different Malignancy. Bull. Exp. Biol. Med. 2014, 157, 794–797. [Google Scholar] [CrossRef]
- Liu, Z.; Jiang, Z.; Huang, J.; Huang, S.; Li, Y.; Yu, S.; Yu, S.; Liu, X. miR-7 inhibits glioblastoma growth by simultaneously interfering with the PI3K/ATK and Raf/MEK/ERK pathways. Int. J. Oncol. 2014, 44, 1571–1580. [Google Scholar] [CrossRef]
- Yin, C.; Kong, W.; Jiang, J.; Xu, H.; Zhao, W. miR-7-5p inhibits cell migration and invasion in glioblastoma through targeting SATB1. Oncol. Lett. 2018, 17, 1819–1825. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Liu, W.; Gu, J.; Wang, J.; Lv, W.; Zhang, W.; Hao, Q.; Pang, Z.; Mu, N.; Zhang, W.; et al. MiR-7-5p suppresses stemness and enhances temozolomide sensitivity of drug-resistant glioblastoma cells by targeting Yin Yang 1. Exp. Cell Res. 2018, 375, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Dai, L.X.; Zhang, S.; Yang, Y.; Yan, N.; Fan, P.; Dai, L.; Tian, H.W.; Cheng, L.; Zhang, X.M.; et al. Regulation of Epidermal Growth Factor Receptor Signaling by plasmid-based MicroRNA-7 inhibits human malignant gliomas growth and metastasis in vivo. Neoplasma 2013, 60, 274–283. [Google Scholar] [CrossRef] [PubMed]
- American-Cancer-Society. Acerca del Cáncer de Colon y Recto, 2016. Available online: https://www.cancer.org/es/cancer/cancer-de-colon-o-recto/acerca/que-es-cancer-de-colon-o-recto.html (accessed on 1 June 2022).
- Nagano, Y.; Toiyama, Y.; Okugawa, Y.; Imaoka, H.; Fujikawa, H.; Yasuda, H.; Yoshiyama, S.; Hiro, J.; Kobayashi, M.; Ohi, M.; et al. MicroRNA-7 Is Associated with Malignant Potential and Poor Prognosis in Human Colorectal Cancer. Anticancer Res. 2016, 36, 6521–6526. [Google Scholar] [CrossRef] [PubMed]
- Suto, T.; Yokobori, T.; Yajima, R.; Morita, H.; Fujii, T.; Yamaguchi, S.; Altan, B.; Tsutsumi, S.; Asao, T.; Kuwano, H. MicroRNA-7 expression in colorectal cancer is associated with poor prognosis and regulates cetuximab sensitivity via EGFR regulation. Carcinogenesis 2014, 36, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Li, X.; Wu, C.W.; Dong, Y.; Cai, M.; Mok, M.T.S.; Wang, H.; Chen, J.; Ng, S.S.M.; Chen, M.; et al. microRNA-7 is a novel inhibitor of YY1 contributing to colorectal tumorigenesis. Oncogene 2012, 32, 5078–5088. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Xie, Y.; Xu, Y. miR-7-5p regulates the proliferation and migration of colorectal cancer cells by negatively regulating the expression of Krüppel-like factor 4. Oncol. Lett. 2019, 17, 3241–3246. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.; Cao, C.; Li, S.; Qiu, M.; Shen, G.; Chen, Z.; Yao, F.; Chen, W. TRIP6, as a target of miR-7, regulates the proliferation and metastasis of colorectal cancer cells. Biochem. Biophys. Res. Commun. 2019, 514, 231–238. [Google Scholar] [CrossRef]
- Romero-Lorca, A.; Novillo, A.; Gaibar, M.; Gilsanz, M.F.; Galán, M.; Beltrán, L.; Antón, B.; Malón, D.; Moreno, A.; Fernández-Santander, A. miR-7, miR-10a and miR-143 Expression May Predict Response to Bevacizumab Plus Chemotherapy in Patients with Metastatic Colorectal Cancer. Pharm. Pers. Med. 2021, 14, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- American-Cancer-Society. Acerca del Cáncer de Próstata, 2016. Available online: https://www.cancer.org/es/cancer/cancer-de-prostata/acerca/que-es-cancer-de-prostata.html (accessed on 1 June 2022).
- Chang, Y.-L.; Zhou, P.-J.; Wei, L.; Li, W.; Ji, Z.; Fang, Y.-X.; Gao, W.-Q. MicroRNA-7 inhibits the stemness of prostate cancer stem-like cells and tumorigenesis by repressing KLF4/PI3K/Akt/p21 pathway. Oncotarget 2015, 6, 24017–24031. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.I.; Teixeira, A.L.; Dias, F.; Maurício, J.; Lobo, F.; Morais, A.; Medeiros, R. Influence of peripheral whole-blood microRNA-7 and microRNA-221 high expression levels on the acquisition of castration-resistant prostate cancer: Evidences from in vitro and in vivo studies. Tumor Biol. 2014, 35, 7105–7113. [Google Scholar] [CrossRef]
- Kirita, T.; Omura, K. Oral Cancer: Diagnosis and Therapy; Springer: Tokyo, Japan, 2015; Volume 27. [Google Scholar] [CrossRef]
- Hara, K.; Miyazaki, T.; Yokobori, T.; Yoshida, T.; Kumakura, Y.; Honjyo, H.; Sakai, M.; Sohda, M.; Fukuchi, M.; Kuwano, H. Significance and Function of MicroRNA-7 in Oesophageal Squamous Cell Carcinoma. Anticancer Res. 2017, 37, 1043–1048. [Google Scholar] [CrossRef]
- Dong, W.; Li, B.; Wang, J.; Song, Y.; Zhang, Z.; Fu, C.; Zhang, P. Diagnostic and predictive significance of serum microRNA-7 in esophageal squamous cell carcinoma. Oncol. Rep. 2015, 35, 1449–1456. [Google Scholar] [CrossRef] [PubMed]
- American-Cancer-Society. Acerca del Cáncer de Tiroides, 2016. Available online: https://www.cancer.org/es/cancer/cancer-de-tiroides/acerca/que-es-cancer-de-tiroides.html (accessed on 1 June 2022).
- Hua, K.; Jin, J.; Zhang, H.; Zhao, B.; Wu, C.; Xu, H.; Fang, L. MicroRNA-7 inhibits proliferation, migration and invasion of thyroid papillary cancer cells via targeting CKS2. Int. J. Oncol. 2016, 49, 1531–1540. [Google Scholar] [CrossRef]
- Yue, K.; Wang, X.; Wu, Y.; Zhou, X.; He, Q.; Duan, Y. microRNA-7 regulates cell growth, migration and invasion via direct targeting of PAK1 in thyroid cancer. Mol. Med. Rep. 2016, 14, 2127–2134. [Google Scholar] [CrossRef][Green Version]
- American-Cancer-Society. Acerca del Cáncer de Piel Tipo Melanoma, 2016. Available online: https://www.cancer.org/es/cancer/cancer-de-piel-tipo-melanoma/acerca/que-es-melanoma.html (accessed on 1 June 2022).
- Sun, X.; Li, J.; Sun, Y.; Zhang, Y.; Dong, L.; Shen, C.; Yang, L.; Yang, M.; Li, Y.; Shen, G.; et al. miR-7 reverses the resistance to BRAFi in melanoma by targeting EGFR/IGF-1R/CRAF and inhibiting the MAPK and PI3K/AKT signaling pathways. Oncotarget 2016, 7, 53558–53570. [Google Scholar] [CrossRef]
- Giles, K.M.; Brown, R.A.; Ganda, C.; Podgorny, M.J.; Candy, P.A.; Wintle, L.C.; Richardson, K.L.; Kalinowski, F.C.; Stuart, L.M.; Epis, M.R.; et al. microRNA-7-5p inhibits melanoma cell proliferation and metastasis by suppressing RelA/NF-κB. Oncotarget 2016, 7, 31663–31680. [Google Scholar] [CrossRef]
- Alejandro López Saavedra, M.L.S. Cáncer cérvicouterino y el virus del papiloma humano: La historia que no termina. In Unidad de Investigación Biomédica en Cáncer; UNAM-INCan: Tlalpan, Mexico, 2006; pp. 31–55. [Google Scholar]
- Liu, S.; Zhang, P.; Chen, Z.; Liu, M.; Li, X.; Tang, H. MicroRNA-7 downregulates XIAP expression to suppress cell growth and promote apoptosis in cervical cancer cells. FEBS Lett. 2013, 587, 2247–2253. [Google Scholar] [CrossRef]
- Bi, Y.; Shen, W.; Min, M.; Liu, Y. MicroRNA-7 functions as a tumor-suppressor gene by regulating ILF2 in pancreatic carcinoma. Int. J. Mol. Med. 2017, 39, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Lu, Y.-Y.; Guo, H.; Xie, H.-H.; He, L.-J.; Shen, G.-F.; Zhou, J.-F.; Li, T.; Hu, S.-J.; Zhou, L.; et al. MicroRNA-7/NF-κB signaling regulatory feedback circuit regulates gastric carcinogenesis. J. Cell Biol. 2015, 210, 613–627. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Hu, Y.; Dai, L.; Wang, Y.; Zhou, J.; Wang, W.; Di, W.; Qiu, L. MicroRNA-7 Inhibits Tumor Metastasis and Reverses Epithelial-Mesenchymal Transition through AKT/ERK1/2 Inactivation by Targeting EGFR in Epithelial Ovarian Cancer. PLoS ONE 2014, 9, e96718. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Ni, L.; Chen, D.; Zhang, Q.; Su, Z.; Wang, Y.; Yu, W.; Wu, X.; Ye, J.; Yang, S.; et al. Identification of miR-7 as an oncogene in renal cell carcinoma. Histochem. J. 2013, 44, 669–677. [Google Scholar] [CrossRef]
- Cheng, J.; Guo, S.; Chen, S.; Mastriano, S.J.; Liu, C.; D’Alessio, A.C.; Hysolli, E.; Guo, Y.; Yao, H.; Megyola, C.M.; et al. An Extensive Network of TET2-Targeting MicroRNAs Regulates Malignant Hematopoiesis. Cell Rep. 2013, 5, 471–481. [Google Scholar] [CrossRef]
- Xu, L.; Liang, Y.; Luo, X.; Liu, X.; Guo, H. Association of miRNAs expression profiles with prognosis and relapse in childhood acute lymphoblastic leukemia. Zhonghua Xue Ye Xue Za Zhi 2011, 32, 178–181. [Google Scholar] [CrossRef]
- Chen, X.; Xiong, W.; Li, H. Comparison of microRNA expression profiles in K562-cells-derived microvesicles and parental cells, and analysis of their roles in leukemia. Oncol. Lett. 2016, 12, 4937–4948. [Google Scholar] [CrossRef]
- Blume, C.J.; Hotz-Wagenblatt, A.; Hüllein, J.; Sellner, L.; Jethwa, A.; Stolz, T.; Slabicki, M.; Lee, K.; Sharathchandra, A.; Benner, A.; et al. p53-dependent non-coding RNA networks in chronic lymphocytic leukemia. Leukemia 2015, 29, 2015–2023. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, Z.; Luo, W.; Liu, J.; Lou, Y.; Xia, S. MiR-7 Functions as a Tumor Suppressor by Targeting the Oncogenes TAL1 in T-Cell Acute Lymphoblastic Leukemia. Technol. Cancer Res. Treat. 2020, 19-1. [Google Scholar] [CrossRef]
- Sorrentino, D.; Frentzel, J.; Mitou, G.; Blasco, R.B.; Torossian, A.; Hoareau-Aveilla, C.; Pighi, C.; Farcé, M.; Meggetto, F.; Manenti, S.; et al. High Levels of miR-7-5p Potentiate Crizotinib-Induced Cytokilling and Autophagic Flux by Targeting RAF1 in NPM-ALK Positive Lymphoma Cells. Cancers 2020, 12, 2951. [Google Scholar] [CrossRef]
- Blosse, A.; Levy, M.; Robe, C.; Staedel, C.; Copie-Bergman, C.; Lehours, P. Deregulation of miRNA in Helicobacter pylori-Induced Gastric MALT Lymphoma: From Mice to Human. J. Clin. Med. 2019, 8, 845. [Google Scholar] [CrossRef] [PubMed]
- Lou, X.; Fu, J.; Zhao, X.; Zhuansun, X.; Rong, C.; Sun, M.; Niu, H.; Wu, L.; Zhang, Y.; An, L.; et al. MiR-7e-5p downregulation promotes transformation of low-grade follicular lymphoma to aggressive lymphoma by modulating an immunosuppressive stroma through the upregulation of FasL in M1 macrophages. J. Exp. Clin. Cancer Res. 2020, 39, 237. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Corrigan-Cummins, M.; Hudson, J.; Maric, I.; Simakova, O.; Neelapu, S.S.; Kwak, L.; Janik, J.E.; Gause, B.; Jaffe, E.S.; et al. MicroRNA profiling of follicular lymphoma identifies microRNAs related to cell proliferation and tumor response. Haematologica 2011, 97, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Morales-Martinez, M.; Vega, G.G.; Neri, N.; Nambo, M.J.; Alvarado, I.; Cuadra, I.; Duran-Padilla, M.A.; Huerta-Yepez, S.; Vega, M.I. MicroRNA-7 Regulates Migration and Chemoresistance in Non-Hodgkin Lymphoma Cells Through Regulation of KLF4 and YY1. Front. Oncol. 2020, 10, 588893. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.-N.; Huang, Q.; Tian, L. The molecular mechanisms and therapeutic potential of microRNA-7 in cancer. Expert Opin. Ther. Targets 2014, 19, 415–426. [Google Scholar] [CrossRef]
- Horsham, J.L.; Kalinowski, F.C.; Epis, M.R.; Ganda, C.; Brown, R.A.M.; Leedman, P.J. Clinical Potential of microRNA-7 in Cancer. J. Clin. Med. 2015, 4, 1668–1687. [Google Scholar] [CrossRef]
- Alizadeh, A.A.; Gentles, A.J.; Alencar, A.J.; Liu, C.L.; Kohrt, H.E.; Houot, R.; Goldstein, M.J.; Zhao, S.; Natkunam, Y.; Advani, R.; et al. Prediction of survival in diffuse large B-cell lymphoma based on the expression of 2 genes reflecting tumor and microenvironment. Blood 2011, 118, 1350–1358. [Google Scholar] [CrossRef]
- Schotte, D.; De Menezes, R.X.; Moqadam, F.A.; Khankahdani, L.M.; Lange-Turenhout, E.; Chen, C.; Pieters, R.; Boer, M.L.D. MicroRNA characterize genetic diversity and drug resistance in pediatric acute lymphoblastic leukemia. Haematologica 2011, 96, 703–711. [Google Scholar] [CrossRef]
- GEO Accession Viewer. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE124489 (accessed on 29 March 2022).
- GEO Accession Viewer. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=gse31312 (accessed on 29 March 2022).












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Function | Type of Cancer | References |
|---|---|---|---|
| EGFR | Promotes cell survival, proliferation, tumorigenesis, resistance to therapeutic targets and radiotherapy | Head and neck, Glioblastoma multiforme, cervical, lung, breast, and prostate | [2,3,4,5,6] |
| RAF1 | Promotes cell survival | Lung, breast, and HNC | [3,5,7] |
| PAK1 | Promotes cell survival, proliferation, cell motility, invasion, growth, and tumorigenesis | Breast, squamous cell carcinoma of the tongue, Schwannoma | [6,8,9] |
| IRS-1 | Promotes proliferation | Glioblastoma multiforme, breast and cervical | [8,10] |
| IRS-2 | Promotes cell survival, proliferation, motility, and invasion | Glioblastoma multiforme, lung, breast, prostate, squamous cell carcinoma of the tongue, and Schwannoma melanoma | [3,8,9,10,11] |
| ACK1 | Promotes cell proliferation, and tumorigenesis | Schwannoma | [6] |
| PI3KCD | Promotes cell survival, proliferation, tumorigenesis, and metastasis | Hepatocellular carcinoma | [12] |
| mTOR | Promotes cell survival, proliferation, tumorigenesis, and metastasis | Hepatocellular carcinoma | [12] |
| P70S6K | Promotes cell survival, proliferation, tumorigenesis, and metastasis | Hepatocellular carcinoma | [12] |
| BCL-2 | Promotes resistance to apoptosis, proliferation, and tumorigenesis | Lung | [13] |
| XIAP | Promotes cell survival and proliferation | Cervical | [14] |
| YY1 | Promotes cell survival, proliferation, and tumorigenesis | Colorectal | [15] |
| CCNE1 | Promotes cell survival | Hepatocellular carcinoma | [15,16] |
| PA28γ gamma | Promotes cell survival, proliferation, and tumorigenesis | Lung | [17] |
| FAK | Promotes cell proliferation, cell survival, tumorigenesis, and cell mobility and regulates EMT | Glioblastoma multiforme and breast | [18,19] |
| KLF4 | Promotes metastasis and self-renewal of stem-type cancer cells | Breast | [20] |
| IGF1R | Promotes cell survival, proliferation, migration, invasion, and metastasis | Squamous cell carcinoma of the tongue, gastric | [9,21] |
| MRP1 | Promotes resistance to chemotherapy | Breast | [22] |
| ERF | Represses checkpoints in the cell cycle | Lung | [23] |
| Type of Cancer | MiR-7 Expression Levels | Role | Reference |
|---|---|---|---|
| Lung | Low | Inhibits tumor growth and metastasis Promotes chemosensitivity | [24,25] |
| Hepatocellular | Low | Tumor suppression by CCNE1 inhibition | [26] |
| Breast | High | Cell cycle promotion | [27] |
| Gliomas | Low | Inhibits oncogenes Suppresses metastases | [20,28] |
| Colorectal | High | Tumor suppressor through YY1 inhibition | [92] |
| Prostate | High | Tumor suppression by inhibition of KLF4 | [30] |
| Oral | Low | Proliferation suppression | [31] |
| Thyroid | ND | Suppresses cell proliferation, migration and invasion | [32] |
| Melanoma | Low | Reverses resistance, decreases tumor growth | [33,34] |
| Cervix | Low | Promotes apoptosis, decreases cell viability. | [14,35] |
| Pancreatic | ND | Suppression of the epithelial–mesenchymal transition | [36] |
| Adrenocortical | Low | Reduction in cell proliferation | [37] |
| Gastric | ND | Prevents cell proliferation and tumorigenesis | [38] |
| Ovary | Low | Inhibits tumor metastasis and reverses epithelial–mesenchymal transition | [39] |
| Follicular lymphoma | High | Associated with better response to chemotherapy | [40] |
| Lymphoblastic leukemia | High | Related to early relapse. | [41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales-Martínez, M.; Vega, M.I. Role of MicroRNA-7 (MiR-7) in Cancer Physiopathology. Int. J. Mol. Sci. 2022, 23, 9091. https://doi.org/10.3390/ijms23169091
Morales-Martínez M, Vega MI. Role of MicroRNA-7 (MiR-7) in Cancer Physiopathology. International Journal of Molecular Sciences. 2022; 23(16):9091. https://doi.org/10.3390/ijms23169091
Chicago/Turabian StyleMorales-Martínez, Mario, and Mario I. Vega. 2022. "Role of MicroRNA-7 (MiR-7) in Cancer Physiopathology" International Journal of Molecular Sciences 23, no. 16: 9091. https://doi.org/10.3390/ijms23169091
APA StyleMorales-Martínez, M., & Vega, M. I. (2022). Role of MicroRNA-7 (MiR-7) in Cancer Physiopathology. International Journal of Molecular Sciences, 23(16), 9091. https://doi.org/10.3390/ijms23169091