
Fertility Impairment after Trekking at High Altitude: A Proof of Mechanisms on Redox and Metabolic Seminal Changes

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  , and
, and 
Abstract
1. Introduction
1.1. High-Altitude Physiology
1.2. Male Reproductive Physiology at High Altitude
1.3. Redox Role
1.4. The Role of Cholesterol and Its Metabolites
1.5. Purpose
2. Results
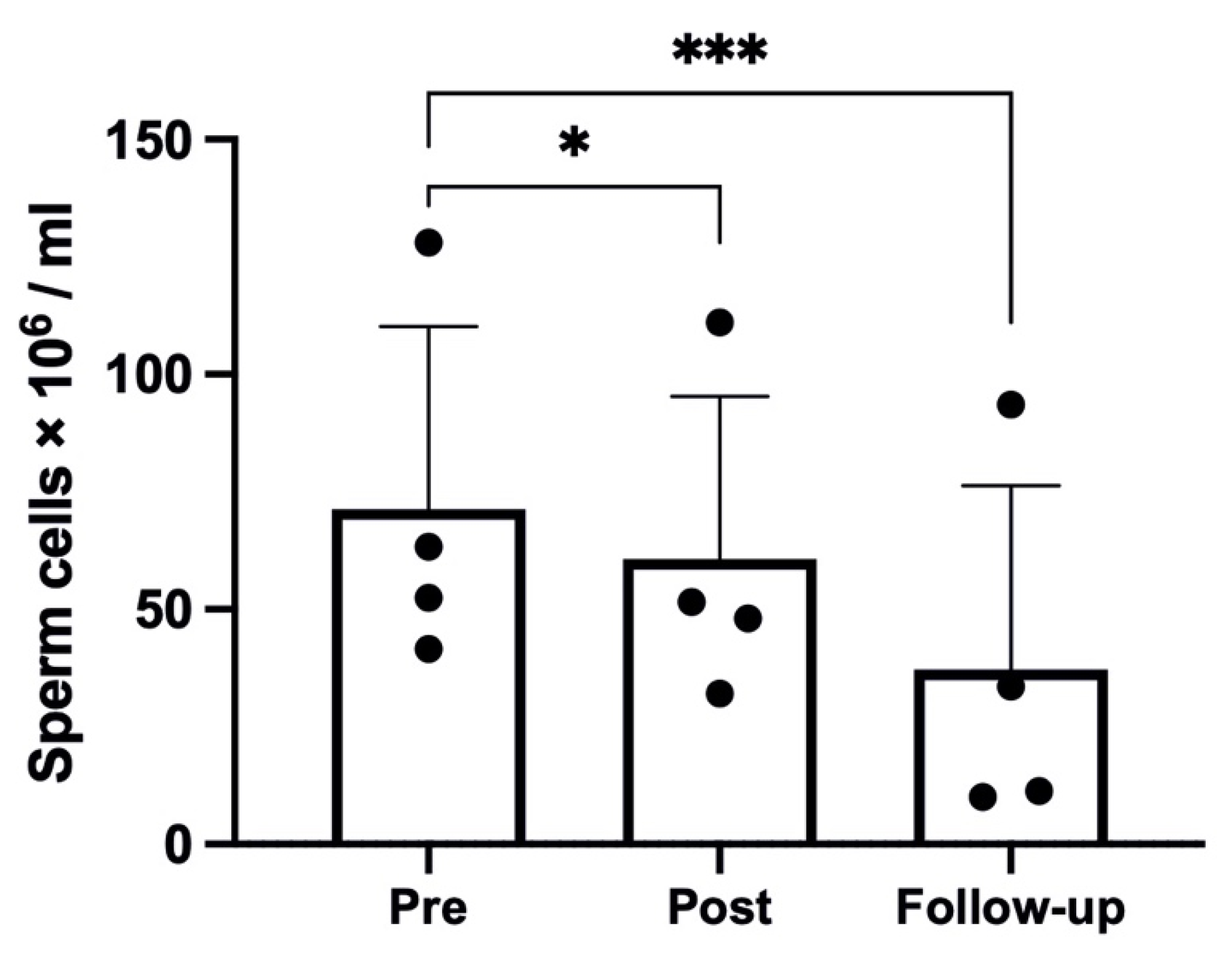
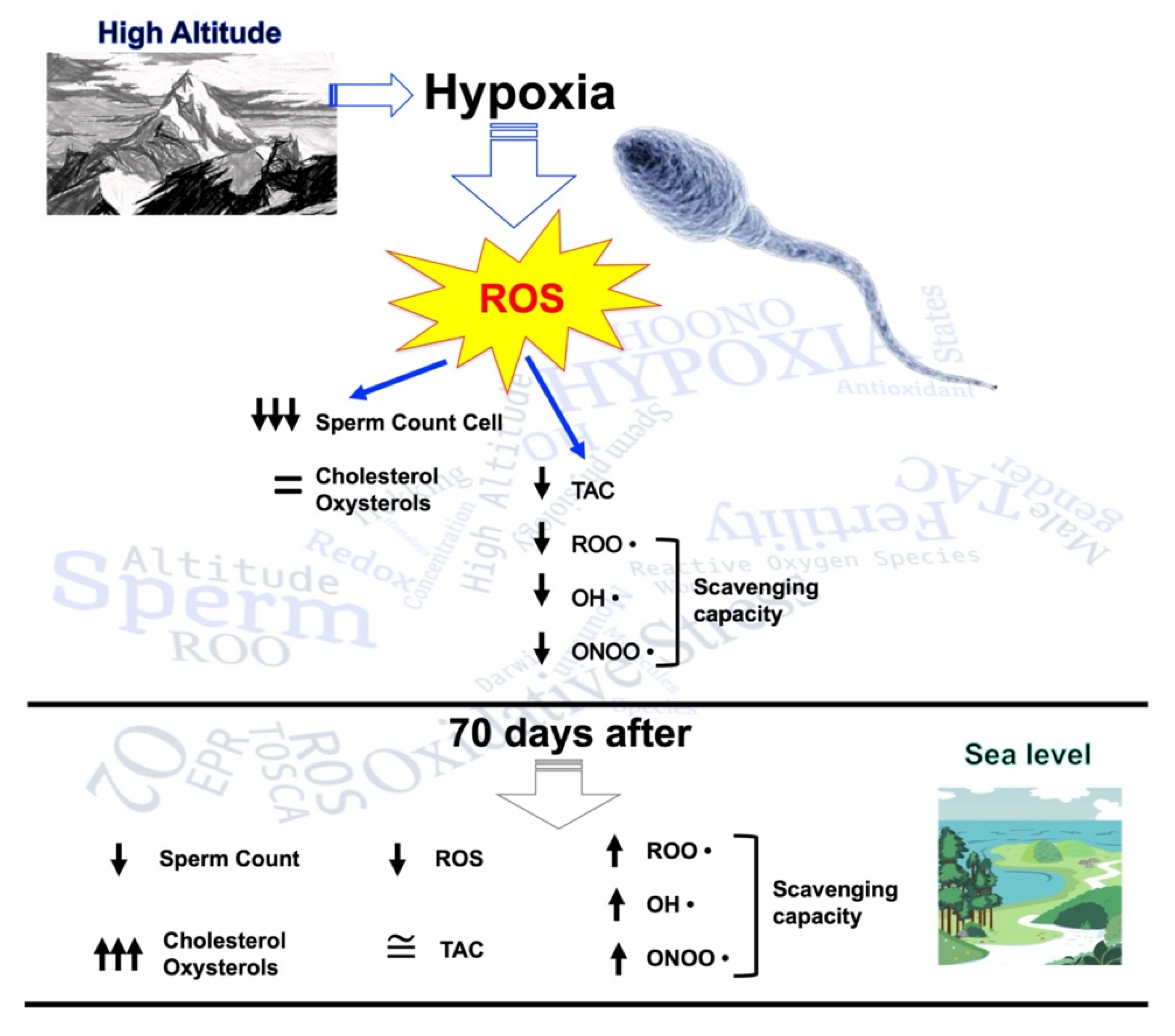
2.1. Sperm Parameters
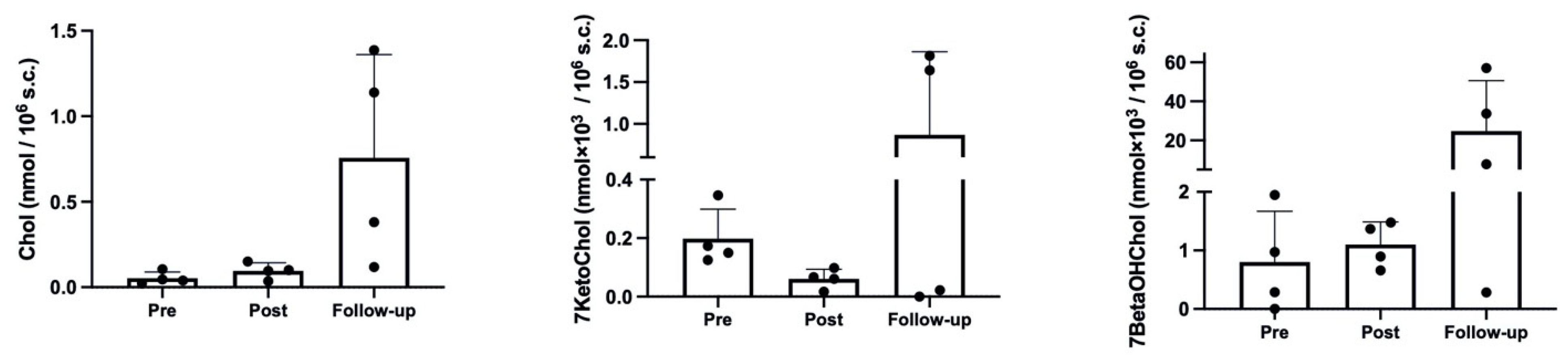
2.2. Cholesterol and Oxysterol Membrane Content
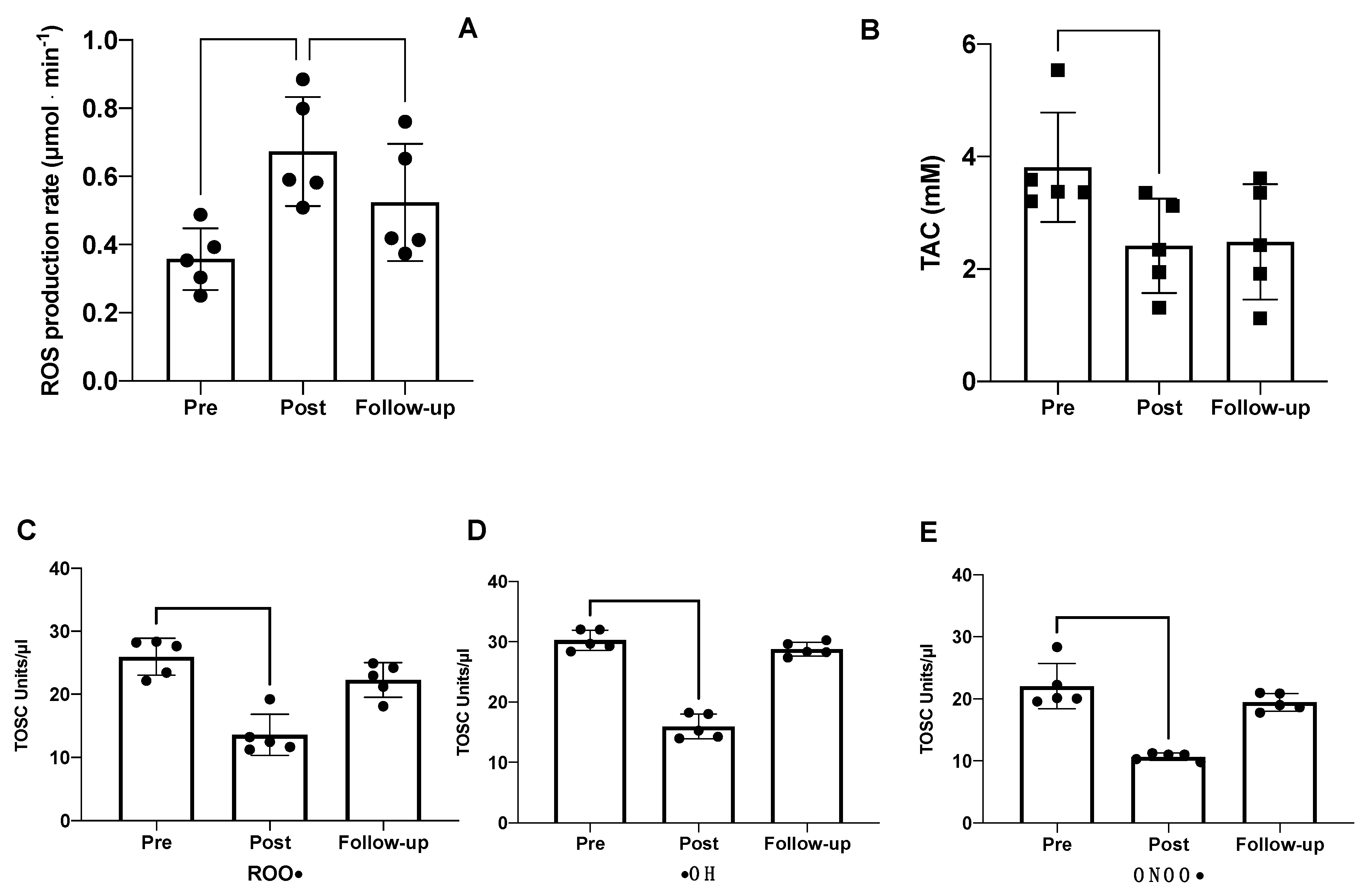
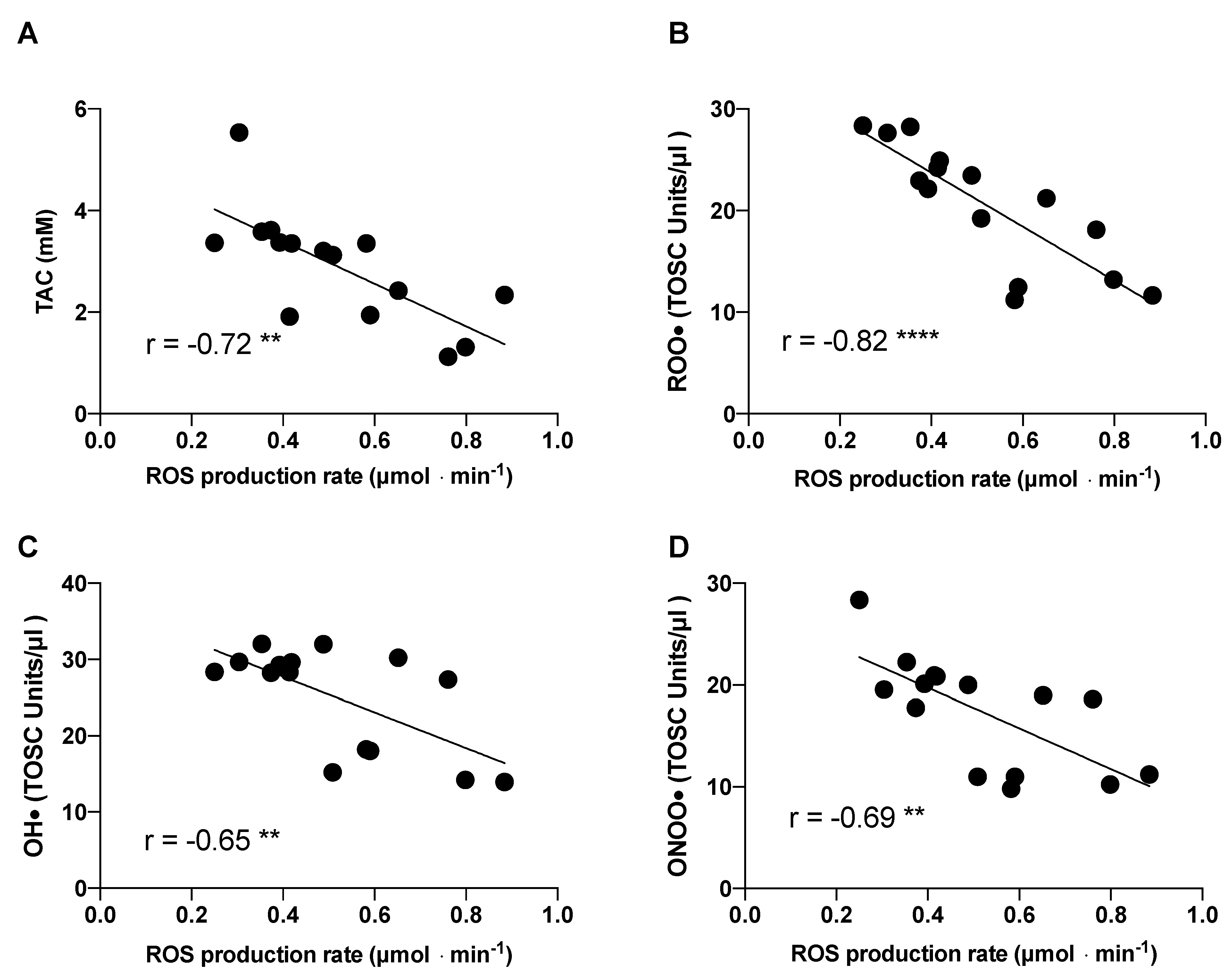
2.3. Redox Analysis
2.4. Other Redox and Hormonal Results
3. Discussion
“What checks the natural tendency of each species to increase in number is most obscure [...] Each species, even where it most abounds, is constantly suffering enormous destruction at some period of its life, from enemies or from competitors for the same place and food [...] When we travel southward and see a species decreasing in numbers, we may feel sure that the cause lies quite as much in other species being favoured, as in this one being hurt. So it is when we travel northward, but in a somewhat lesser degree, for the number of species of all kinds, and therefore of competitors, decreases northwards; hence in going northward, or in ascending a mountain, we far oftener meet with stunted forms, due to the directly injurious action of climate, than we do in proceeding southwards or in descending a mountain. When we reach the Arctic regions, or snow-capped summits, or absolute deserts, the struggle for life is almost exclusively with the elements”[55]
4. Materials and Methods
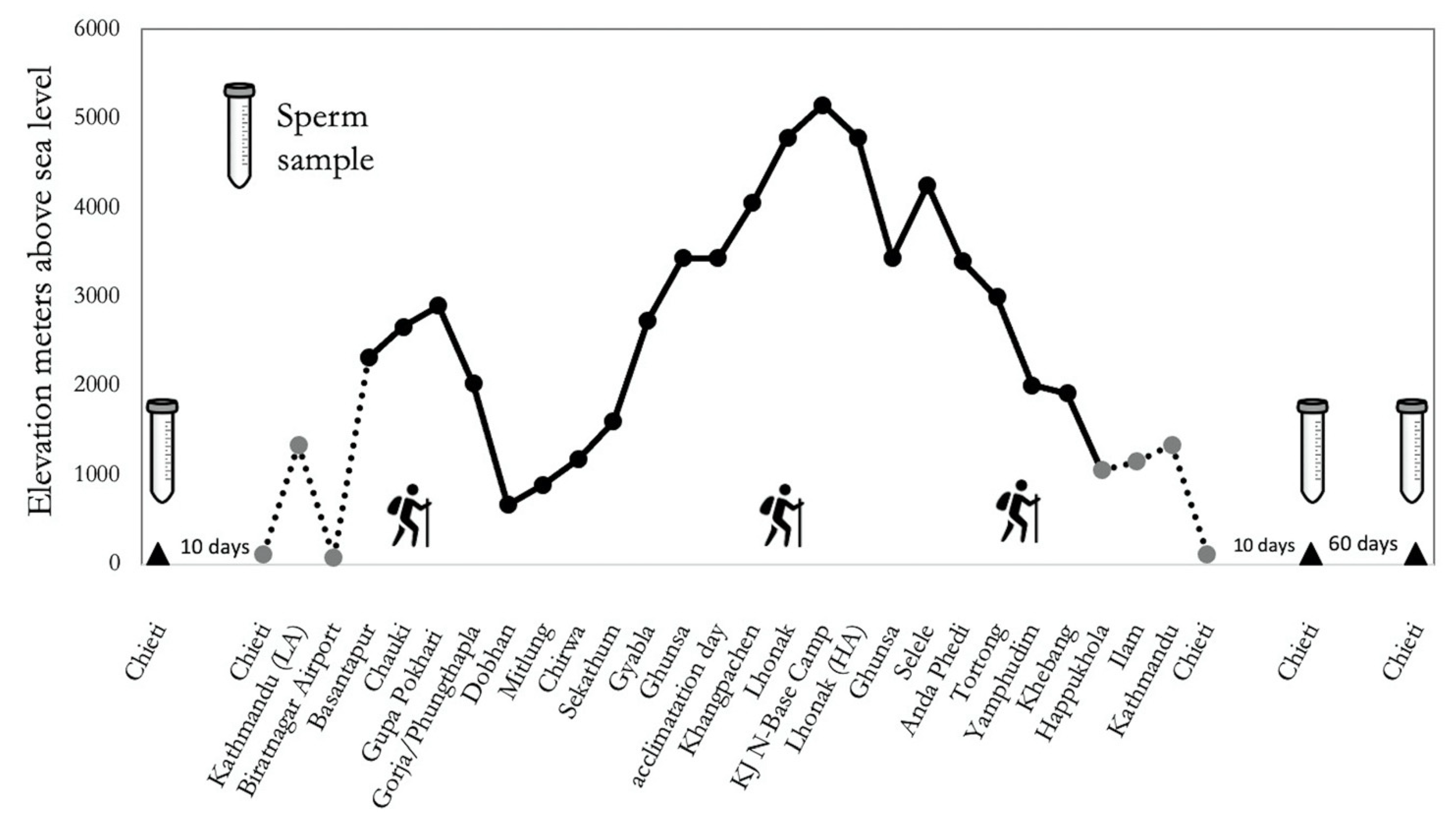
4.1. Participants and Study Protocol
4.2. Semen Collection and Analysis
4.3. Extraction and Cholesterol/Oxysterols Measurements Using LC-MS
4.4. ROS and TAC Determination via Electron Paramagnetic Resonance
4.5. Total Oxyradical Scavenging Capacity Assay (TOSCA)
4.6. Other Markers from the Same Project
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Verratti, V.; Ferrante, C.; Soranna, D.; Zambon, A.; Bhandari, S.; Orlando, G.; Brunetti, L.; Parati, G. Effect of High-Altitude Trekking on Blood Pressure and on Asymmetric Dimethylarginine and Isoprostane Production: Results from a Mount Ararat Expedition. J. Clin. Hypertens. 2020, 22, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Luks, A.M.; Levett, D.; Martin, D.S.; Goss, C.H.; Mitchell, K.; Fernandez, B.O.; Feelisch, M.; Grocott, M.P.; Swenson, E.R. Caudwell Xtreme Everest Investigators Changes in Acute Pulmonary Vascular Responsiveness to Hypoxia during a Progressive Ascent to High Altitude (5300 m). Exp. Physiol. 2017, 102, 711–724. [Google Scholar] [CrossRef] [PubMed]
- Pelliccione, F.; Verratti, V.; D’Angeli, A.; Micillo, A.; Doria, C.; Pezzella, A.; Iacutone, G.; Francavilla, F.; Di Giulio, C.; Francavilla, S. Physical Exercise at High Altitude Is Associated with a Testicular Dysfunction Leading to Reduced Sperm Concentration but Healthy Sperm Quality. Fertil. Steril. 2011, 96, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Doria, C.; Verratti, V.; Pietrangelo, T.; Fanò-Illic, G.; Bisconti, A.V.; Shokohyar, S.; Rampichini, S.; Limonta, E.; Coratella, G.; Longo, S.; et al. Changes in Energy System Contributions to the Wingate Anaerobic Test in Climbers after a High Altitude Expedition. Eur. J. Appl. Physiol. 2020, 120, 1629–1636. [Google Scholar] [CrossRef] [PubMed]
- Magliulo, L.; Bondi, D.; Pietrangelo, T.; Fulle, S.; Piccinelli, R.; Jandova, T.; Blasio, G.D.; Taraborrelli, M.; Verratti, V. Serum Ferritin and Vitamin D Evaluation in Response to High Altitude Comparing Italians Trekkers vs Nepalese Porters. Eur. J. Sport Sci. 2020, 21, 994–1002. [Google Scholar] [CrossRef]
- Tam, E.; Bruseghini, P.; Calabria, E.; Dal Sacco, L.; Doria, C.; Grassi, B.; Pietrangelo, T.; Pogliaghi, S.; Reggiani, C.; Salvadego, D.; et al. Gokyo Khumbu/Ama Dablam Trek 2012: Effects of Physical Training and High-Altitude Exposure on Oxidative Metabolism, Muscle Composition, and Metabolic Cost of Walking in Women. Eur. J. Appl. Physiol. 2016, 116, 129–144. [Google Scholar] [CrossRef]
- Verratti, V.; Bondi, D.; Mulliri, G.; Ghiani, G.; Crisafulli, A.; Pietrangelo, T.; Marinozzi, M.E.; Cerretelli, P. Muscle Oxygen Delivery in the Forearm and in the Vastus Lateralis Muscles in Response to Resistance Exercise: A Comparison Between Nepalese Porters and Italian Trekkers. Front. Physiol. 2020, 11, 607616. [Google Scholar] [CrossRef]
- Julian, C.G.; Moore, L.G. Human Genetic Adaptation to High Altitude: Evidence from the Andes. Genes 2019, 10, 150. [Google Scholar] [CrossRef]
- Martin, D.S.; Gilbert-Kawai, E.; Levett, D.Z.; Mitchell, K.; Kumar BC, R.; Mythen, M.G.; Grocott, M.P. Xtreme Everest 2: Unlocking the Secrets of the Sherpa Phenotype? Extreme Physiol. Med. 2013, 2, 30. [Google Scholar] [CrossRef]
- Mrakic-Sposta, S.; Gussoni, M.; Dellanoce, C.; Marzorati, M.; Montorsi, M.; Rasica, L.; Pratali, L.; D’Angelo, G.; Martinelli, M.; Bastiani, L.; et al. Effects of Acute and Sub-Acute Hypobaric Hypoxia on Oxidative Stress: A Field Study in the Alps. Eur. J. Appl. Physiol. 2021, 121, 297–306. [Google Scholar] [CrossRef]
- Strapazzon, G.; Malacrida, S.; Vezzoli, A.; Dal Cappello, T.; Falla, M.; Lochner, P.; Moretti, S.; Procter, E.; Brugger, H.; Mrakic-Sposta, S. Oxidative Stress Response to Acute Hypobaric Hypoxia and Its Association with Indirect Measurement of Increased Intracranial Pressure: A Field Study. Sci. Rep. 2016, 6, 32426. [Google Scholar] [CrossRef] [PubMed]
- Cassinelli, L.M. Biology of fertility; acclimatization of the ovine animals to various altitudes. An. Univ. Nac. Mayor San Marcos Fac. Med. 1953, 36, 419–446. [Google Scholar]
- Calancha, A. Crónicas Agustinianas del Perú; Consejo Superior de Investigaciones Científicas: Madrid, Spain, 1972. [Google Scholar]
- Monge, C. Life in the Andes and Chronic Mountain Sickness. Science 1942, 95, 79–84. [Google Scholar] [CrossRef]
- Zou, Z.; Hu, H.; Song, M.; Shen, Y.; Guo, X.; McElreavey, K.; Bittles, A.H.; Wang, W. Semen Quality Analysis of Military Personnel from Six Geographical Areas of the People’s Republic of China. Fertil. Steril. 2011, 95, 2018–2023.e3. [Google Scholar] [CrossRef] [PubMed][Green Version]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; WHO Press: Geneva, Switzerland, 2010. [Google Scholar]
- Wan, L.; Wang, Y.-M. Impact of high altitude and hypoxia on sperm concentration. Zhonghua Nan Ke Xue Natl. J. Androl. 2012, 18, 835–839. [Google Scholar]
- He, J.; Cui, J.; Wang, R.; Gao, L.; Gao, X.; Yang, L.; Zhang, Q.; Cao, J.; Yu, W. Exposure to Hypoxia at High Altitude (5380 m) for 1 Year Induces Reversible Effects on Semen Quality and Serum Reproductive Hormone Levels in Young Male Adults. High Alt. Med. Biol. 2015, 16, 216–222. [Google Scholar] [CrossRef]
- Verratti, V.; Berardinelli, F.; Di Giulio, C.; Bosco, G.; Cacchio, M.; Pellicciotta, M.; Nicolai, M.; Martinotti, S.; Tenaglia, R. Evidence That Chronic Hypoxia Causes Reversible Impairment on Male Fertility. Asian J. Androl. 2008, 10, 602–606. [Google Scholar] [CrossRef]
- Zheng, S.; Liu, Y.; Li, P.; Tian, H. Short-Term High-Altitude Exposure (3600 m) Alters the Type Distribution of Sperm Deformity. High Alt. Med. Biol. 2019, 20, 198–202. [Google Scholar] [CrossRef]
- Gu, B.-H.; Liu, F.; Li, J.; Zhu, Z.-J.; Ni, Y.-T.; Li, P.; Chen, M.; Duoji, Z.; Xue, Y.-J.; Chen, H.-X.; et al. Impact of plateau environment on seminal characteristics of native Tibetans and immigrated Tibetan Hans. Zhonghua Nan Ke Xue Natl. J. Androl. 2018, 24, 27–32. [Google Scholar]
- Cofré, E.; Peralta, O.A.; Raggi, A.; De los Reyes, M.; Sales, F.; González-Bulnes, A.; Parraguez, V.H. Ram Semen Deterioration by Short-Term Exposure to High Altitude Is Prevented by Improvement of Antioxidant Status. Animal 2018, 12, 1007–1014. [Google Scholar] [CrossRef]
- Farias, J.G.; Bustos-Obregón, E.; Orellana, R.; Bucarey, J.L.; Quiroz, E.; Reyes, J.G. Effects of Chronic Hypobaric Hypoxia on Testis Histology and Round Spermatid Oxidative Metabolism. Andrologia 2005, 37, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Verratti, V.; Tartaro, A.; Falone, S.; Pellegrini, M.; Pelliccione, F.; Di Giulio, C. Long Trekking Experience at High Altitude Causes Testicular Volumetric Reduction in Humans: Evidence Based on Magnetic Resonance Imaging. High Alt. Med. Biol. 2017, 18, 191–192. [Google Scholar] [CrossRef] [PubMed]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, Oxidative Damage and Oxygen Deprivation Stress: A Review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Zepeda, A.B.; Figueroa, C.A.; Calaf, G.M.; Farías, J.G. Male Reproductive System and Antioxidants in Oxidative Stress Induced by Hypobaric Hypoxia. Andrologia 2014, 46, 1–8. [Google Scholar] [CrossRef]
- Sharma, R.K.; Pasqualotto, F.F.; Nelson, D.R.; Thomas, A.J.; Agarwal, A. The Reactive Oxygen Species-Total Antioxidant Capacity Score Is a New Measure of Oxidative Stress to Predict Male Infertility. Hum. Reprod. Oxf. Engl. 1999, 14, 2801–2807. [Google Scholar] [CrossRef]
- Zerbinati, C.; Caponecchia, L.; Puca, R.; Ciacciarelli, M.; Salacone, P.; Sebastianelli, A.; Pastore, A.; Palleschi, G.; Petrozza, V.; Porta, N.; et al. Mass Spectrometry Profiling of Oxysterols in Human Sperm Identifies 25-Hydroxycholesterol as a Marker of Sperm Function. Redox Biol. 2017, 11, 111–117. [Google Scholar] [CrossRef]
- Garolla, A.; Šabović, I.; Tescari, S.; Toni, L.D.; Menegazzo, M.; Cosci, I.; Filippis, V.D.; Giarola, M.; Foresta, C. Impaired Sperm Function in Infertile Men Relies on the Membrane Sterol Pattern. Andrology 2018, 6, 325–334. [Google Scholar] [CrossRef]
- Mrakic-Sposta, S.; Biagini, D.; Bondi, D.; Pietrangelo, T.; Vezzoli, A.; Lomonaco, T.; Di Francesco, F.; Verratti, V. OxInflammation at High Altitudes: A Proof of Concept from the Himalayas. Antioxidants 2022, 11, 368. [Google Scholar] [CrossRef]
- Tafuri, A.; Bondi, D.; Princiotta, A.; Pietrangelo, T.; Yadav, P.; Altieri, V.M.; Cerruto, M.A.; Pelliccione, F.; Antonelli, A.; Verratti, V. Mid-Term Effects of Physical Activity at High-Altitude on Hormonal Profiles in Italian Trekkers and Nepalese Porters. In Best Practice in Health Care; Springer: Berlin/Heidelberg, Germany, 2020; Volume 1335, pp. 111–119. [Google Scholar] [CrossRef]
- Mrakic-Sposta, S.; Montorsi, M.; Porcelli, S.; Marzorati, M.; Healey, B.; Dellanoce, C.; Vezzoli, A. Effects of Prolonged Exposure to Hypobaric Hypoxia on Oxidative Stress: Overwintering in Antarctic Concordia Station. Oxidative Med. Cell. Longev. 2022, 2022, e4430032. [Google Scholar] [CrossRef]
- Agarwal, A.; Saleh, R.A.; Bedaiwy, M.A. Role of Reactive Oxygen Species in the Pathophysiology of Human Reproduction. Fertil. Steril. 2003, 79, 829–843. [Google Scholar] [CrossRef]
- Debevec, T.; Millet, G.P.; Pialoux, V. Hypoxia-Induced Oxidative Stress Modulation with Physical Activity. Front. Physiol. 2017, 8, 84. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive Oxygen Species (ROS) as Pleiotropic Physiological Signalling Agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Smith, T.B.; Jobling, M.S.; Baker, M.A.; De Iuliis, G.N. Oxidative Stress and Male Reproductive Health. Asian J. Androl. 2014, 16, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Mannucci, A.; Argento, F.R.; Fini, E.; Coccia, M.E.; Taddei, N.; Becatti, M.; Fiorillo, C. The Impact of Oxidative Stress in Male Infertility. Front. Mol. Biosci. 2022, 8, 799294. [Google Scholar] [CrossRef] [PubMed]
- Dada, R.; Bisht, S. Oxidative Stress and Male Infertility. In Male Infertility: Understanding, Causes and Treatment; SINGH, R., Singh, K., Eds.; Springer: Singapore, 2017; pp. 151–165. ISBN 978-981-10-4017-7. [Google Scholar]
- Cao, L.; Leers-Sucheta, S.; Azhar, S. Aging Alters the Functional Expression of Enzymatic and Non-Enzymatic Anti-Oxidant Defense Systems in Testicular Rat Leydig Cells. J. Steroid Biochem. Mol. Biol. 2004, 88, 61–67. [Google Scholar] [CrossRef]
- Selvaraju, V.; Baskaran, S.; Agarwal, A.; Henkel, R. Environmental Contaminants and Male Infertility: Effects and Mechanisms. Andrologia 2021, 53, e13646. [Google Scholar] [CrossRef]
- Sikka, S.C. Oxidative Stress and Role of Antioxidants in Normal and Abnormal Sperm Function. Front. Biosci. J. Virtual Libr. 1996, 1, e78–e86. [Google Scholar] [CrossRef]
- Opuwari, C.S.; Henkel, R.R. An Update on Oxidative Damage to Spermatozoa and Oocytes. BioMed Res. Int. 2016, 2016, 9540142. [Google Scholar] [CrossRef]
- Agarwal, A.; Bui, A.D. Oxidation-Reduction Potential as a New Marker for Oxidative Stress: Correlation to Male Infertility. Investig. Clin. Urol. 2017, 58, 385–399. [Google Scholar] [CrossRef]
- Desai, N.; Sharma, R.; Makker, K.; Sabanegh, E.; Agarwal, A. Physiologic and Pathologic Levels of Reactive Oxygen Species in Neat Semen of Infertile Men. Fertil. Steril. 2009, 92, 1626–1631. [Google Scholar] [CrossRef]
- Agarwal, A.; Said, T.M. Oxidative Stress, DNA Damage and Apoptosis in Male Infertility: A Clinical Approach. BJU Int. 2005, 95, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Athayde, K.S.; Cocuzza, M.; Agarwal, A.; Krajcir, N.; Lucon, A.M.; Srougi, M.; Hallak, J. Development of Normal Reference Values for Seminal Reactive Oxygen Species and Their Correlation with Leukocytes and Semen Parameters in a Fertile Population. J. Androl. 2007, 28, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Khandwala, Y.S.; Zhang, C.A.; Li, S.; Behr, B.; Guo, D.; Eisenberg, M.L. Racial Variation in Semen Quality at Fertility Evaluation. Urology 2017, 106, 96–102. [Google Scholar] [CrossRef] [PubMed]
- De Rooij, D.G.; Russell, L.D. All You Wanted to Know about Spermatogonia but Were Afraid to Ask. J. Androl. 2000, 21, 776–798. [Google Scholar]
- Muciaccia, B.; Boitani, C.; Berloco, B.P.; Nudo, F.; Spadetta, G.; Stefanini, M.; de Rooij, D.G.; Vicini, E. Novel Stage Classification of Human Spermatogenesis Based on Acrosome Development. Biol. Reprod. 2013, 89, 60. [Google Scholar] [CrossRef]
- Heller, C.G.; Clermont, Y. Spermatogenesis in Man: An Estimate of Its Duration. Science 1963, 140, 184–186. [Google Scholar] [CrossRef]
- Oates, R. Evaluation of the Azoospermic Male. Asian J. Androl. 2012, 14, 82–87. [Google Scholar] [CrossRef]
- Haditsch, B.; Roessler, A.; Hinghofer-Szalkay, H.G. Renal Adrenomedullin and High Altitude Diuresis. Physiol. Res. 2007, 56, 779–787. [Google Scholar] [CrossRef]
- Bondi, D.; Aloisi, A.M.; Pietrangelo, T.; Piccinelli, R.; Le Donne, C.; Jandova, T.; Pieretti, S.; Taraborrelli, M.; Santangelo, C.; Lattanzi, B.; et al. Feeding Your Himalayan Expedition: Nutritional Signatures and Body Composition Adaptations of Trekkers and Porters. Nutrients 2021, 13, 460. [Google Scholar] [CrossRef]
- Agarwal, A.; Finelli, R.; Selvam, M.K.P.; Leisegang, K.; Majzoub, A.; Tadros, N.; Ko, E.; Parekh, N.; Henkel, R.; Durairajanayagam, D.; et al. A Global Survey of Reproductive Specialists to Determine the Clinical Utility of Oxidative Stress Testing and Antioxidant Use in Male Infertility. World J. Mens Health 2021, 39, 470. [Google Scholar] [CrossRef]
- Darwin, C.R. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life, 1st ed.; John Murray: London, UK, 1859. [Google Scholar]
- Vitzthum, V.J. Fifty Fertile Years: Anthropologists’ Studies of Reproduction in High Altitude Natives. Am. J. Hum. Biol. 2013, 25, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Brodmann Maeder, M.; Brugger, H.; Pun, M.; Strapazzon, G.; Dal Cappello, T.; Maggiorini, M.; Hackett, P.; Bärtsch, P.; Swenson, E.R.; Zafren, K. The STAR Data Reporting Guidelines for Clinical High Altitude Research. High Alt. Med. Biol. 2018, 19, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, J.F.; Boerke, A.; Silva, P.F.N.; Garcia-Gil, N.; van Gestel, R.A.; Helms, J.B.; van de Lest, C.H.A.; Gadella, B.M. Mass Spectrometric Detection of Cholesterol Oxidation in Bovine Sperm. Biol. Reprod. 2011, 85, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Mrakic-Sposta, S.; Gussoni, M.; Montorsi, M.; Porcelli, S.; Vezzoli, A. A Quantitative Method to Monitor Reactive Oxygen Species Production by Electron Paramagnetic Resonance in Physiological and Pathological Conditions. Oxidative Med. Cell. Longev. 2014, 2014, 306179. [Google Scholar] [CrossRef]
- Mrakic-Sposta, S.; Vezzoli, A.; Malacrida, S.; Falla, M.; Strapazzon, G. “Direct” and “Indirect” Methods to Detect Oxidative Stress During Acute or Chronic High-Altitude Exposure. High Alt. Med. Biol. 2017, 18, 303–304. [Google Scholar] [CrossRef] [PubMed]
- Mrakic-Sposta, S.; Vezzoli, A.; Rizzato, A.; Della Noce, C.; Malacrida, S.; Montorsi, M.; Paganini, M.; Cancellara, P.; Bosco, G. Oxidative Stress Assessment in Breath-Hold Diving. Eur. J. Appl. Physiol. 2019, 119, 2449–2456. [Google Scholar] [CrossRef]
- Zang, S.; Tian, S.; Jiang, J.; Han, D.; Yu, X.; Wang, K.; Li, D.; Lu, D.; Yu, A.; Zhang, Z. Determination of Antioxidant Capacity of Diverse Fruits by Electron Spin Resonance (ESR) and UV-Vis Spectrometries. Food Chem. 2017, 221, 1221–1225. [Google Scholar] [CrossRef]
- Daniele, S.; Pietrobono, D.; Fusi, J.; Iofrida, C.; Chico, L.; Petrozzi, L.; Gerfo, A.L.; Baldacci, F.; Galetta, F.; Siciliano, G.; et al. α-Synuclein Aggregates with β-Amyloid or Tau in Human Red Blood Cells: Correlation with Antioxidant Capability and Physical Exercise in Human Healthy Subjects. Mol. Neurobiol. 2018, 55, 2653–2675. [Google Scholar] [CrossRef]
- Fusi, J.; Bianchi, S.; Daniele, S.; Pellegrini, S.; Martini, C.; Galetta, F.; Giovannini, L.; Franzoni, F. An in Vitro Comparative Study of the Antioxidant Activity and SIRT1 Modulation of Natural Compounds. Biomed. Pharmacother. 2018, 101, 805–819. [Google Scholar] [CrossRef]
- Piccarducci, R.; Daniele, S.; Polini, B.; Carpi, S.; Chico, L.; Fusi, J.; Baldacci, F.; Siciliano, G.; Bonuccelli, U.; Nieri, P.; et al. Apolipoprotein E Polymorphism and Oxidative Stress in Human Peripheral Blood Cells: Can Physical Activity Reactivate the Proteasome System through Epigenetic Mechanisms? Oxid. Med. Cell. Longev. 2021, 2021, e8869849. [Google Scholar] [CrossRef]
- Cooper, T.G.; Noonan, E.; von Eckardstein, S.; Auger, J.; Baker, H.W.G.; Behre, H.M.; Haugen, T.B.; Kruger, T.; Wang, C.; Mbizvo, M.T.; et al. World Health Organization Reference Values for Human Semen Characteristics. Hum. Reprod. Update 2010, 16, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Skakkebaek, N.E. Normal Reference Ranges for Semen Quality and Their Relations to Fecundity. Asian J. Androl. 2010, 12, 95–98. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Years) | BMI Pre (kg/m2) | BMI Post (kg/m2) | |
|---|---|---|---|
| KJ2 | 63 | 28.91 | 27.34 |
| KJ4 | 59 | 21.91 | 21.35 |
| KJ5 | 25 | 24.31 | 23.13 |
| KJ6 | 32 | 24.14 | 23.14 |
| KJ7 | 48 | 30.54 | 27.98 |
| 44 ± 15 | 25.81 ± 3.25 | 24.60 ± 2.60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verratti, V.; Mrakic-Sposta, S.; Fusi, J.; Sabovic, I.; Franzoni, F.; Pietrangelo, T.; Bondi, D.; Dall’Acqua, S.; Daniele, S.; Scarfò, G.; et al. Fertility Impairment after Trekking at High Altitude: A Proof of Mechanisms on Redox and Metabolic Seminal Changes. Int. J. Mol. Sci. 2022, 23, 9066. https://doi.org/10.3390/ijms23169066
Verratti V, Mrakic-Sposta S, Fusi J, Sabovic I, Franzoni F, Pietrangelo T, Bondi D, Dall’Acqua S, Daniele S, Scarfò G, et al. Fertility Impairment after Trekking at High Altitude: A Proof of Mechanisms on Redox and Metabolic Seminal Changes. International Journal of Molecular Sciences. 2022; 23(16):9066. https://doi.org/10.3390/ijms23169066
Chicago/Turabian StyleVerratti, Vittore, Simona Mrakic-Sposta, Jonathan Fusi, Iva Sabovic, Ferdinando Franzoni, Tiziana Pietrangelo, Danilo Bondi, Stefano Dall’Acqua, Simona Daniele, Giorgia Scarfò, and et al. 2022. "Fertility Impairment after Trekking at High Altitude: A Proof of Mechanisms on Redox and Metabolic Seminal Changes" International Journal of Molecular Sciences 23, no. 16: 9066. https://doi.org/10.3390/ijms23169066
APA StyleVerratti, V., Mrakic-Sposta, S., Fusi, J., Sabovic, I., Franzoni, F., Pietrangelo, T., Bondi, D., Dall’Acqua, S., Daniele, S., Scarfò, G., Di Giulio, C., & Garolla, A. (2022). Fertility Impairment after Trekking at High Altitude: A Proof of Mechanisms on Redox and Metabolic Seminal Changes. International Journal of Molecular Sciences, 23(16), 9066. https://doi.org/10.3390/ijms23169066