
The Key Role of Fatty Acid Synthase in Lipid Metabolism and Metamorphic Development in a Destructive Insect Pest, Spodoptera litura (Lepidoptera: Noctuidae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
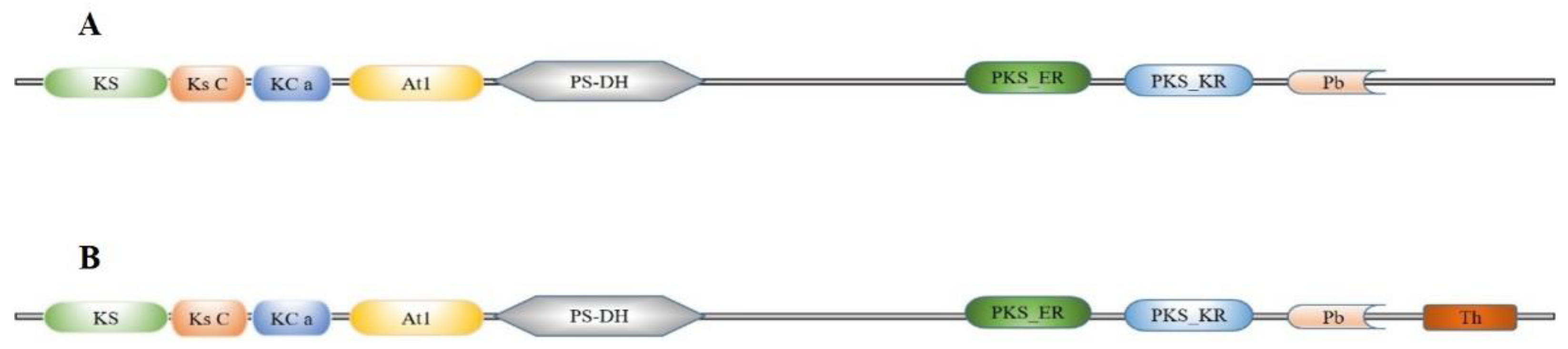
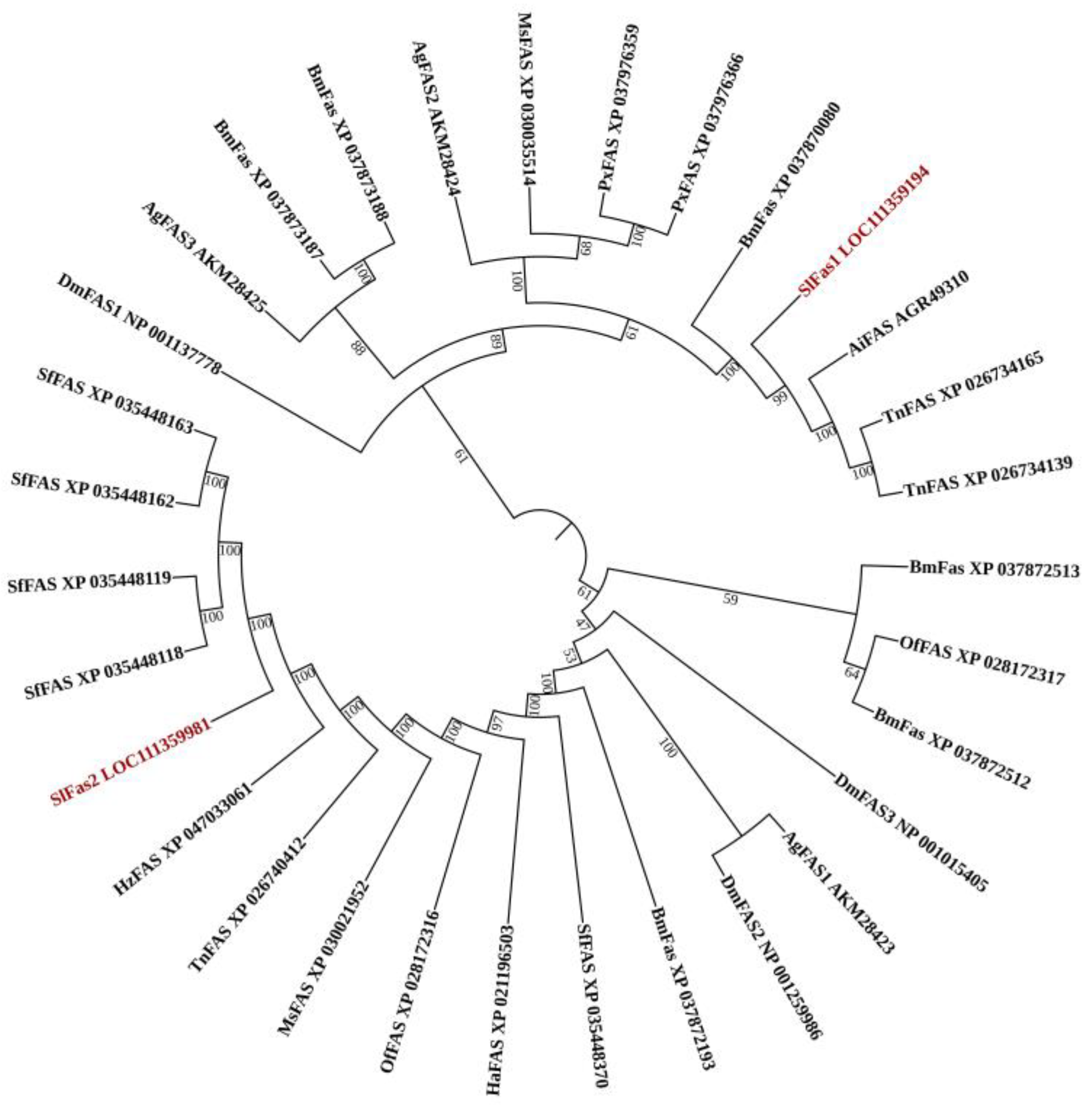
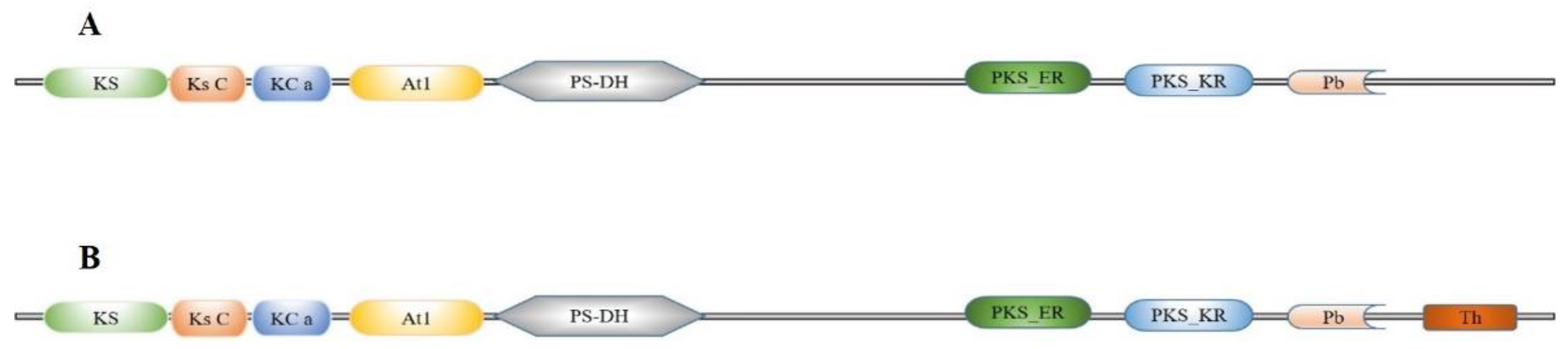
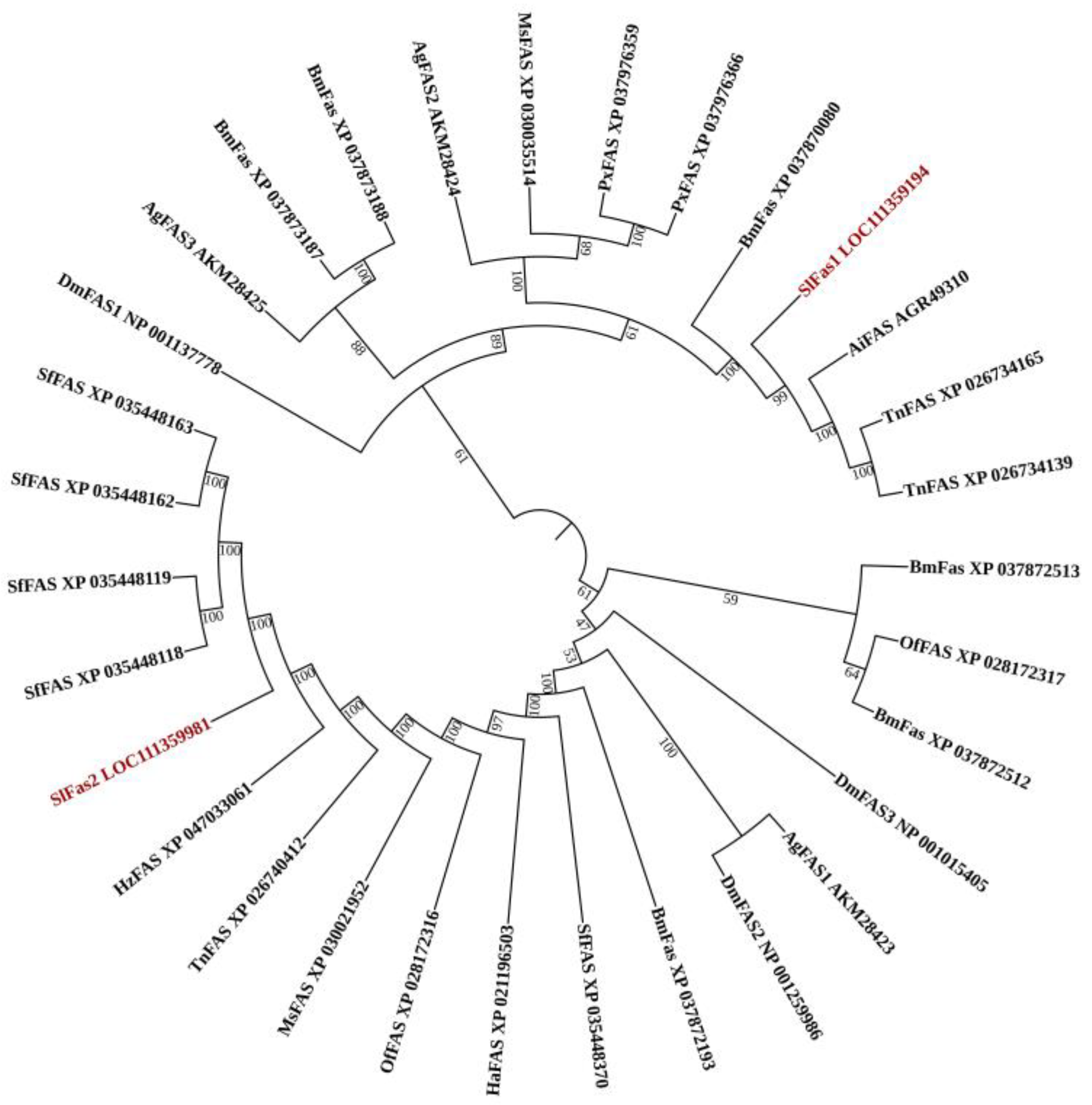
2.1. Gene Identification and Sequence Analysis of FAS Genes in S. litura
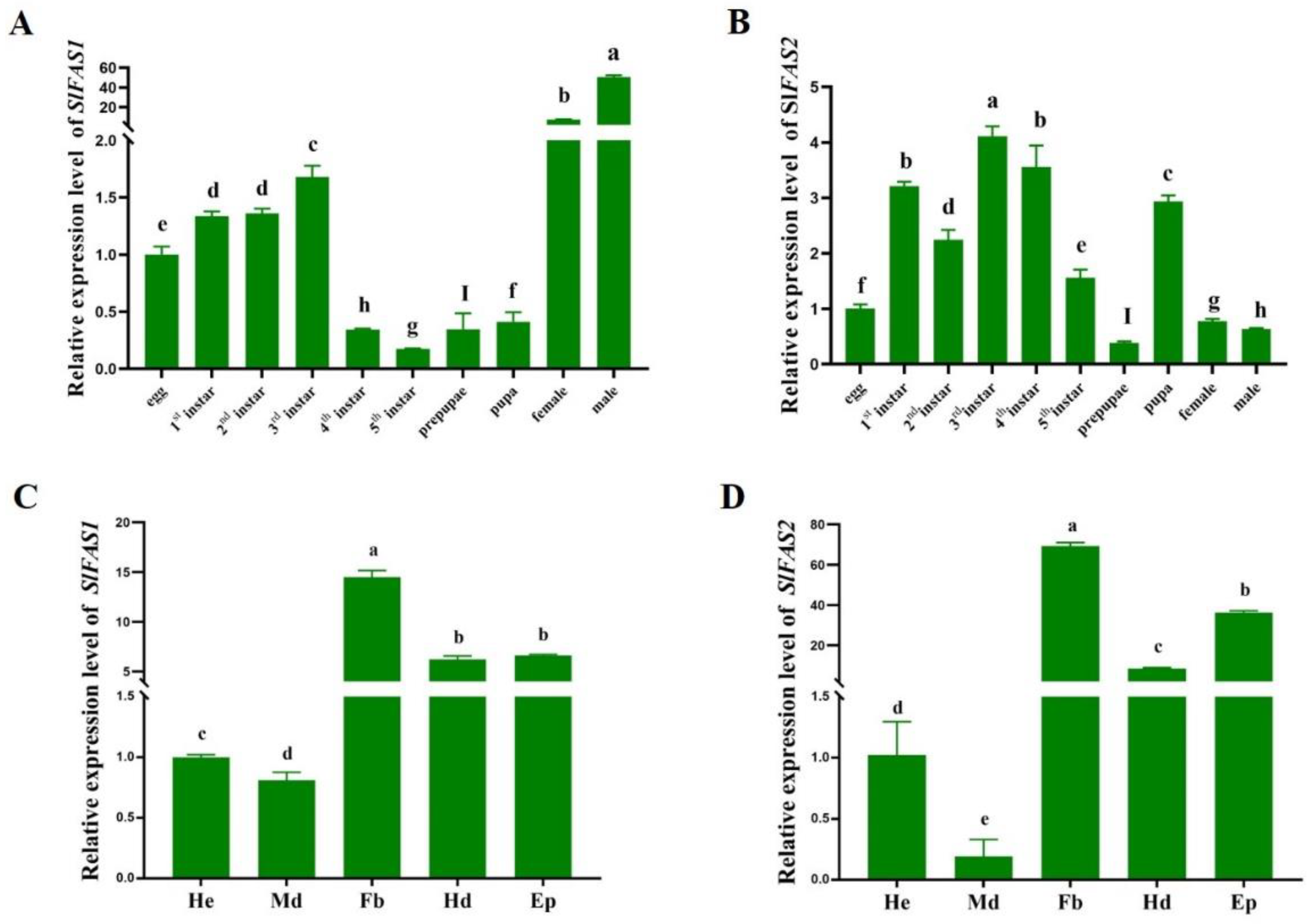
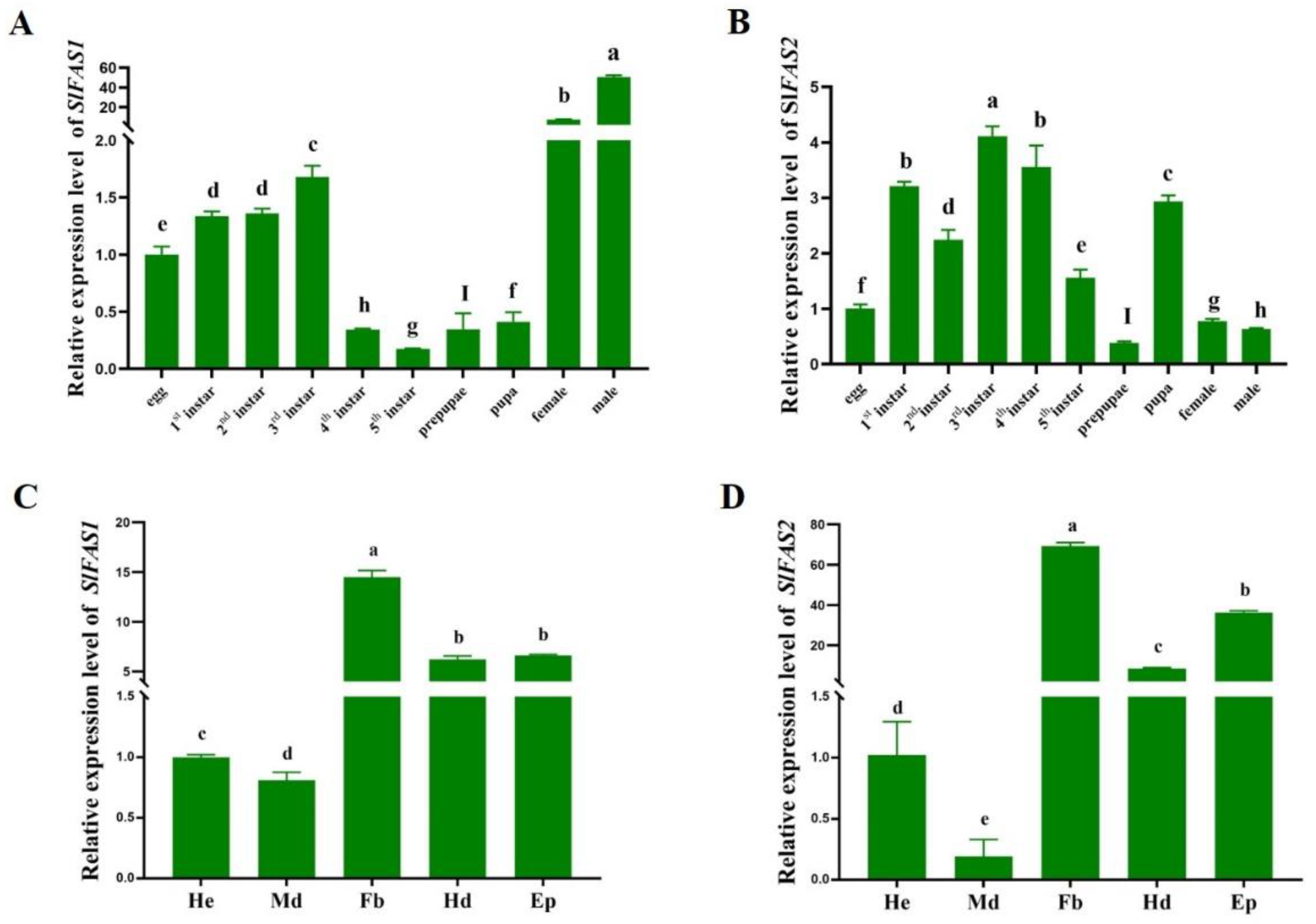
2.2. Expression Patterns of SlFAS1 and SlFAS2
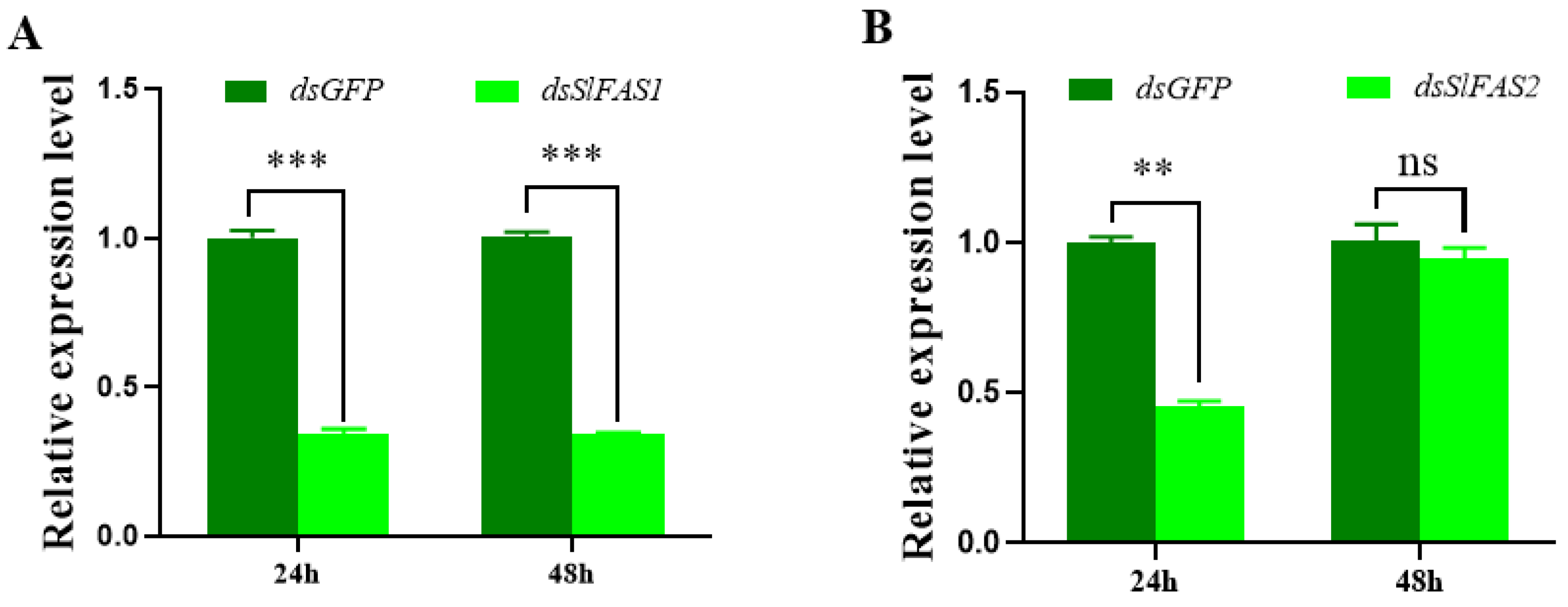
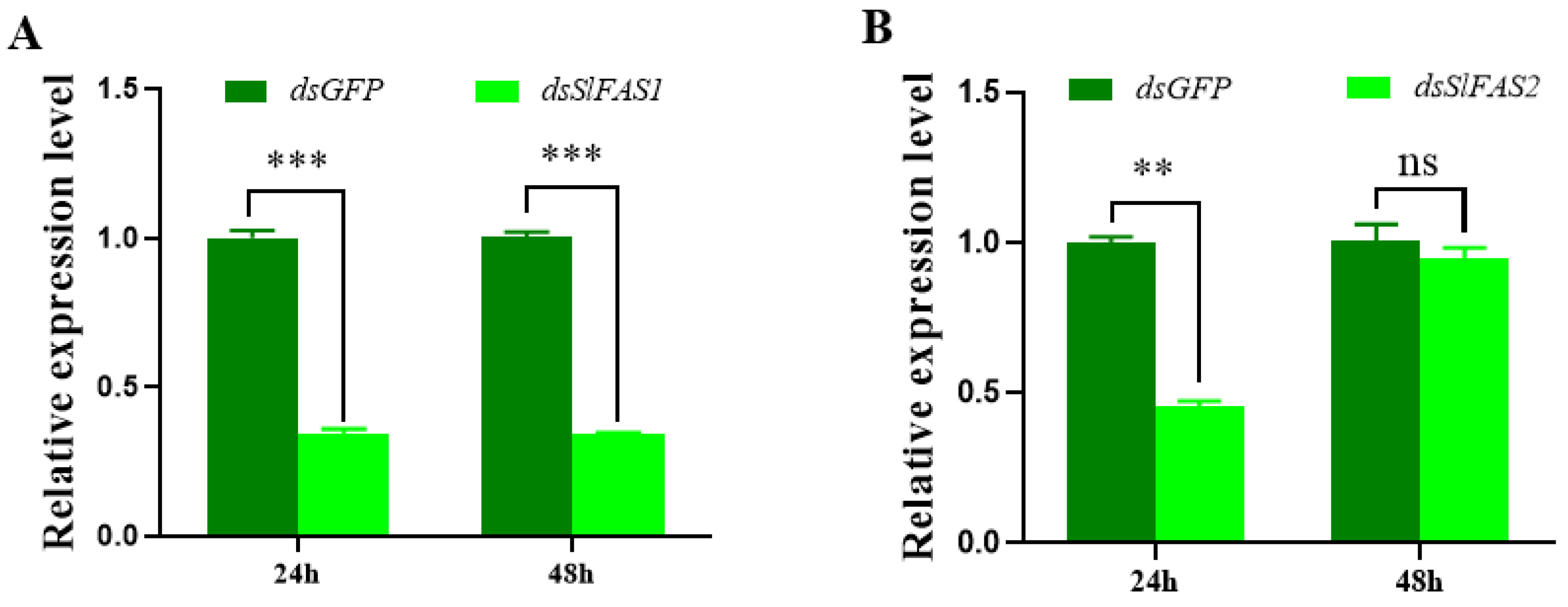
2.3. Analysis of the Function of SlFASs Using RNAi
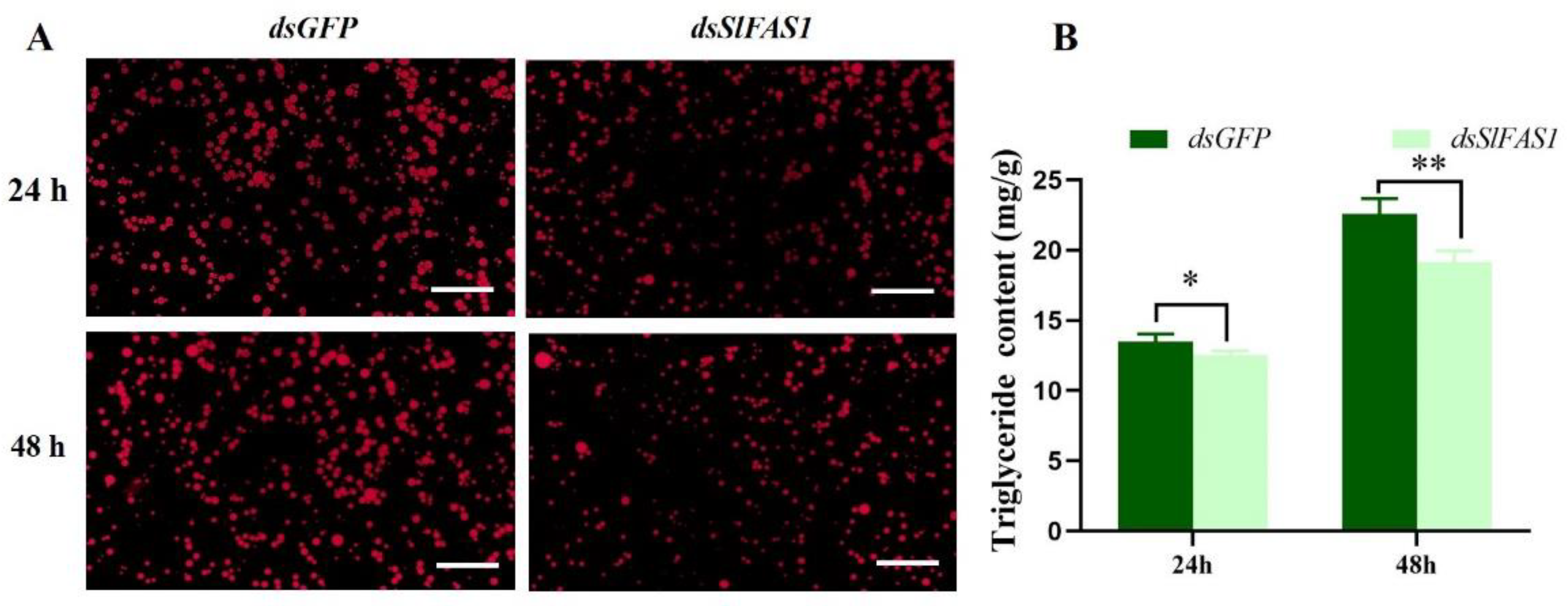
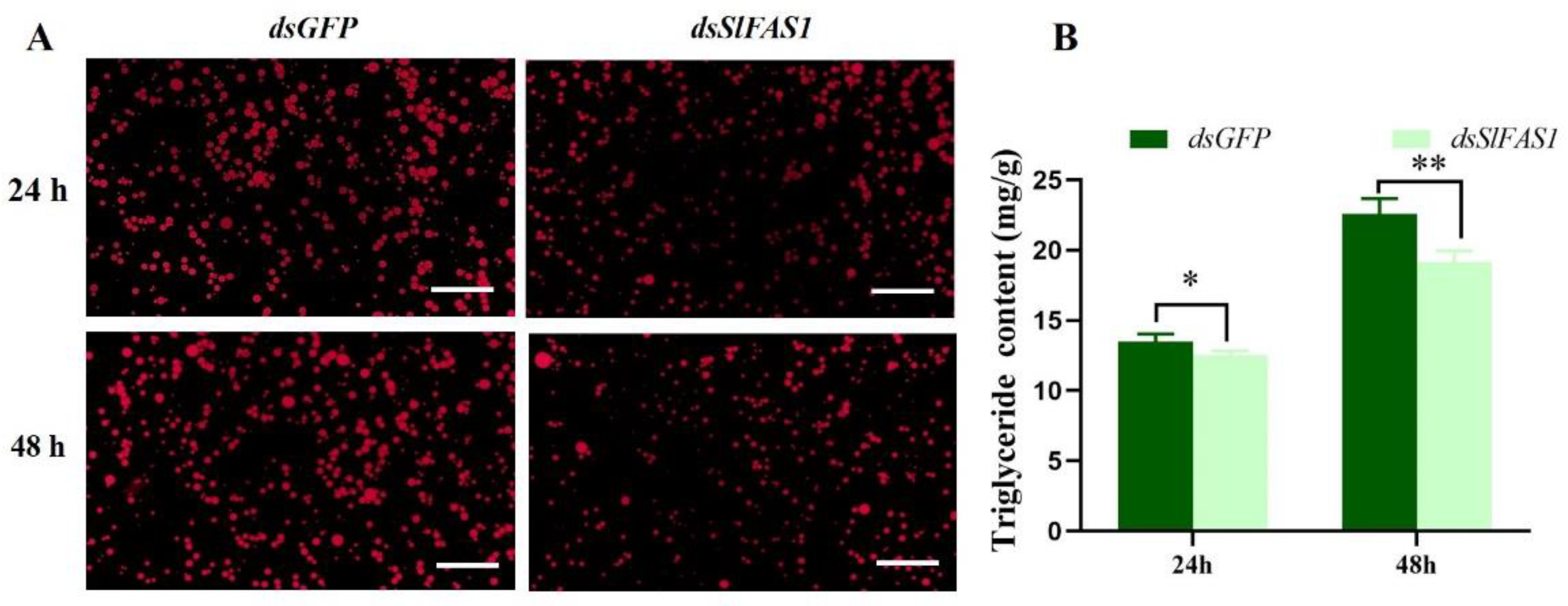
2.4. Effect of Silencing SlFAS1 on Lipid Accumulation in S. litura Larvae
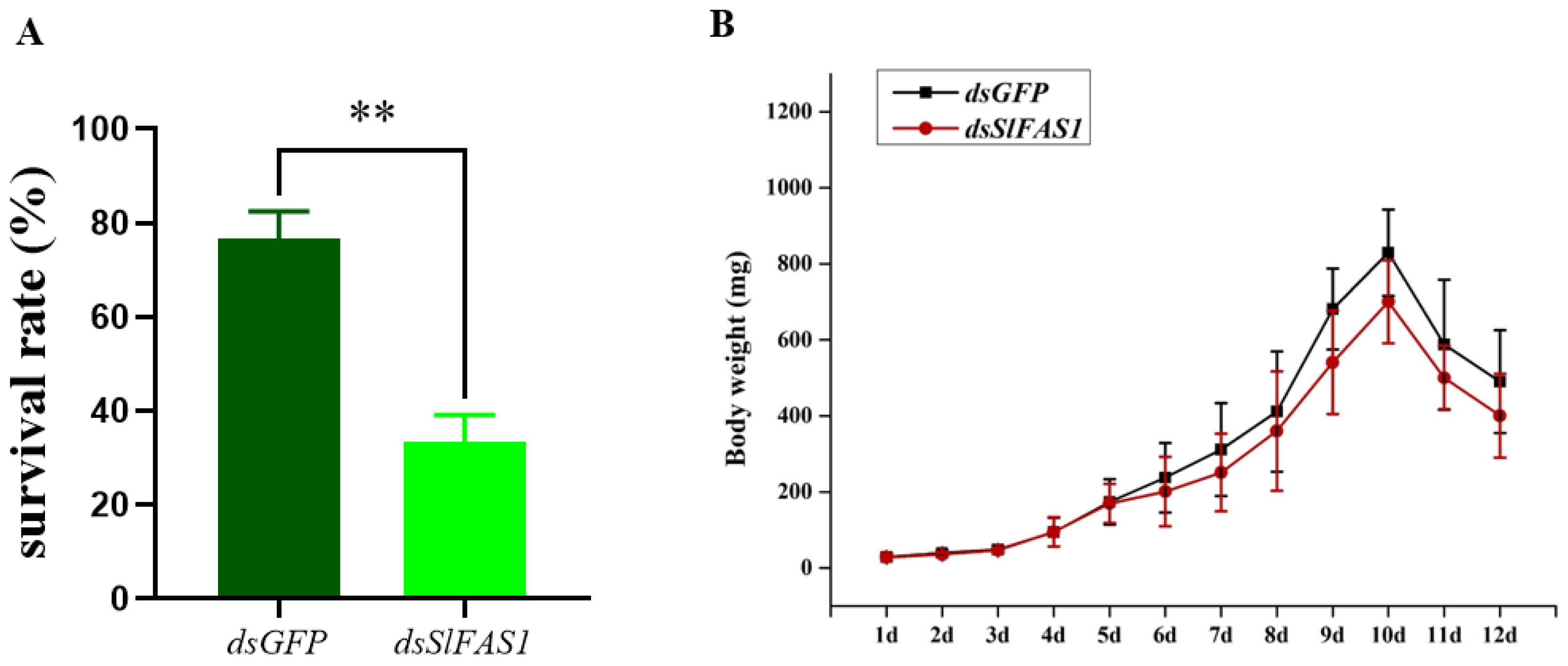
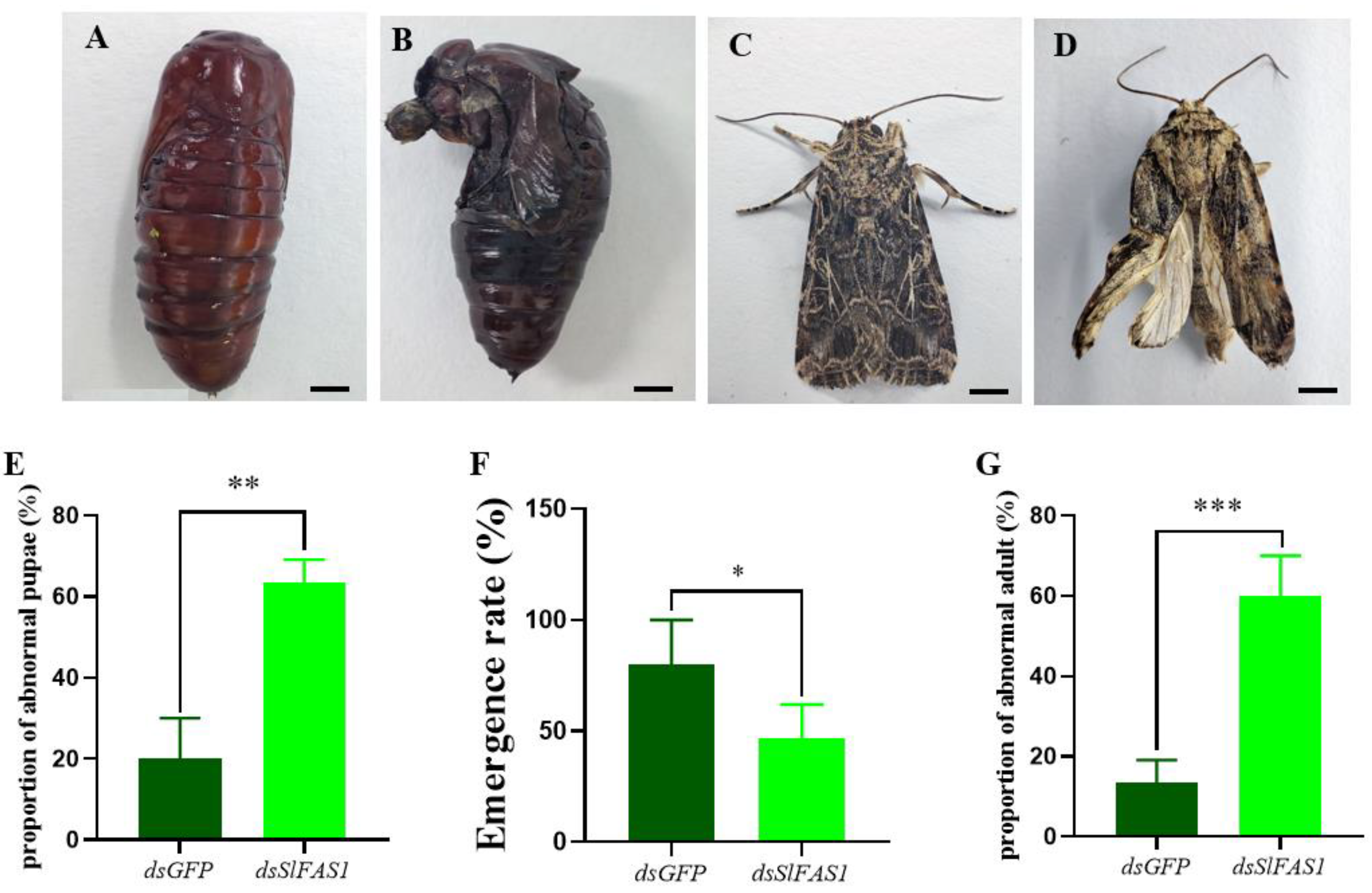
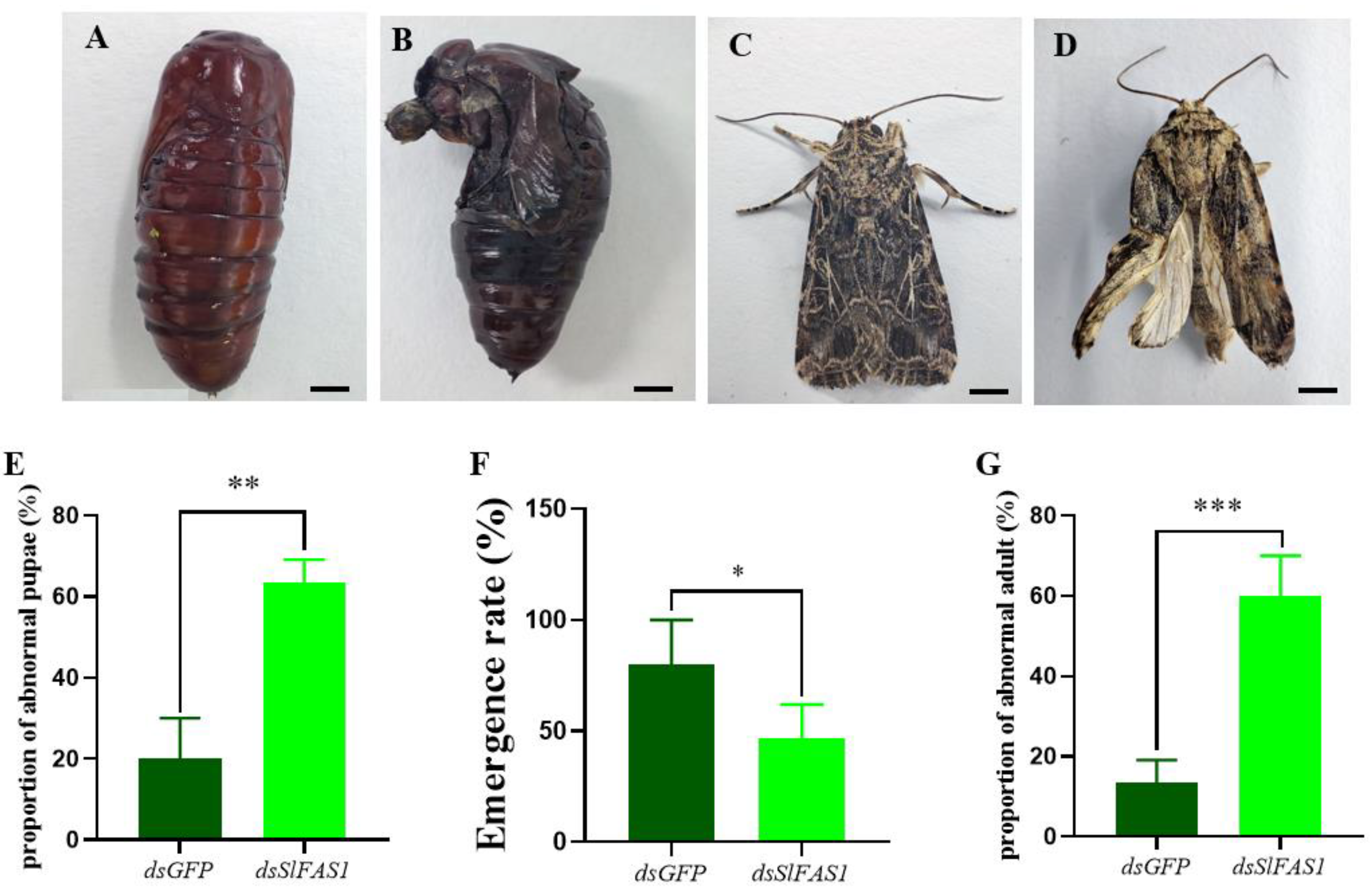
2.5. Effect of Silencing SlFAS1 on the Development of S. litura Larvae
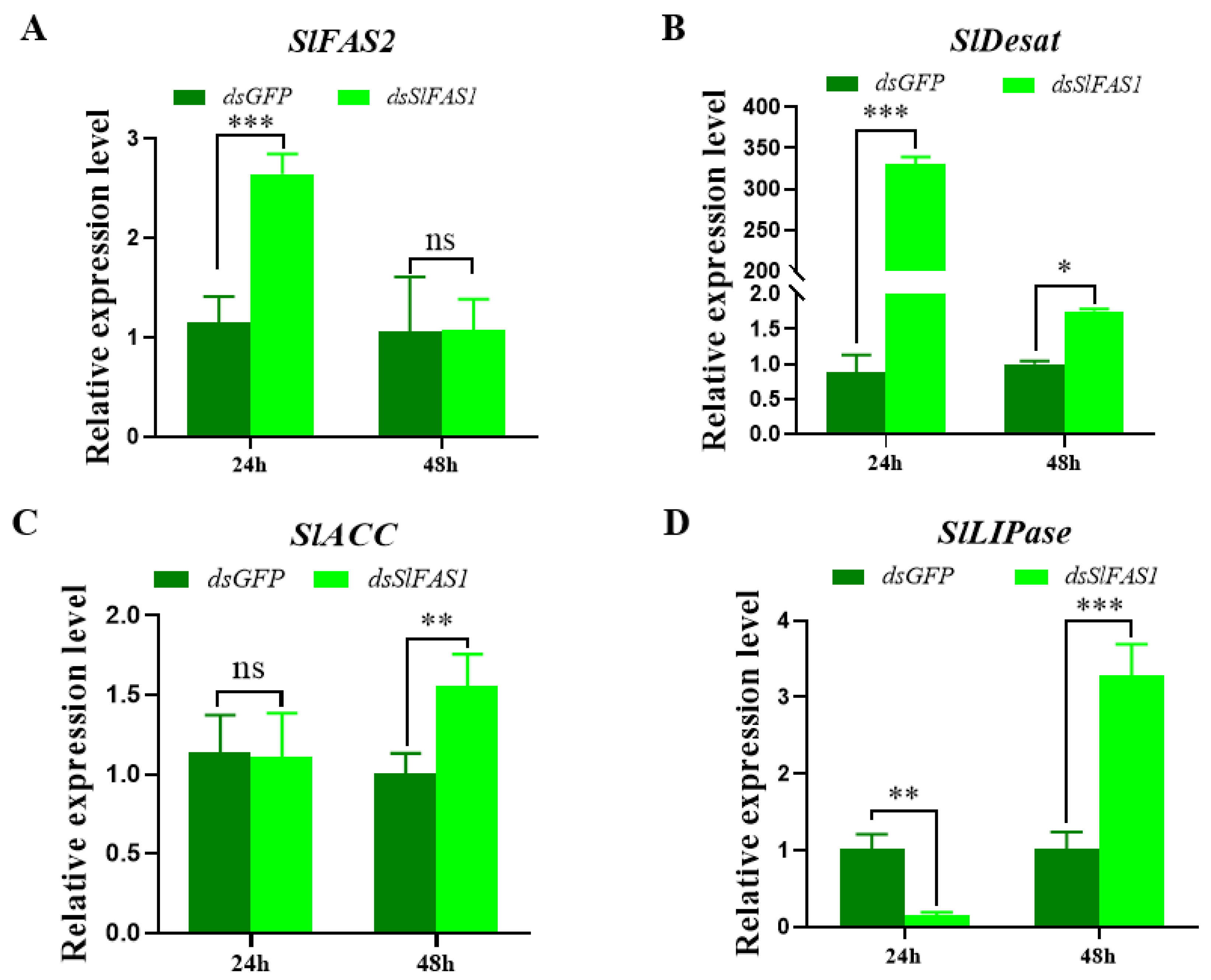
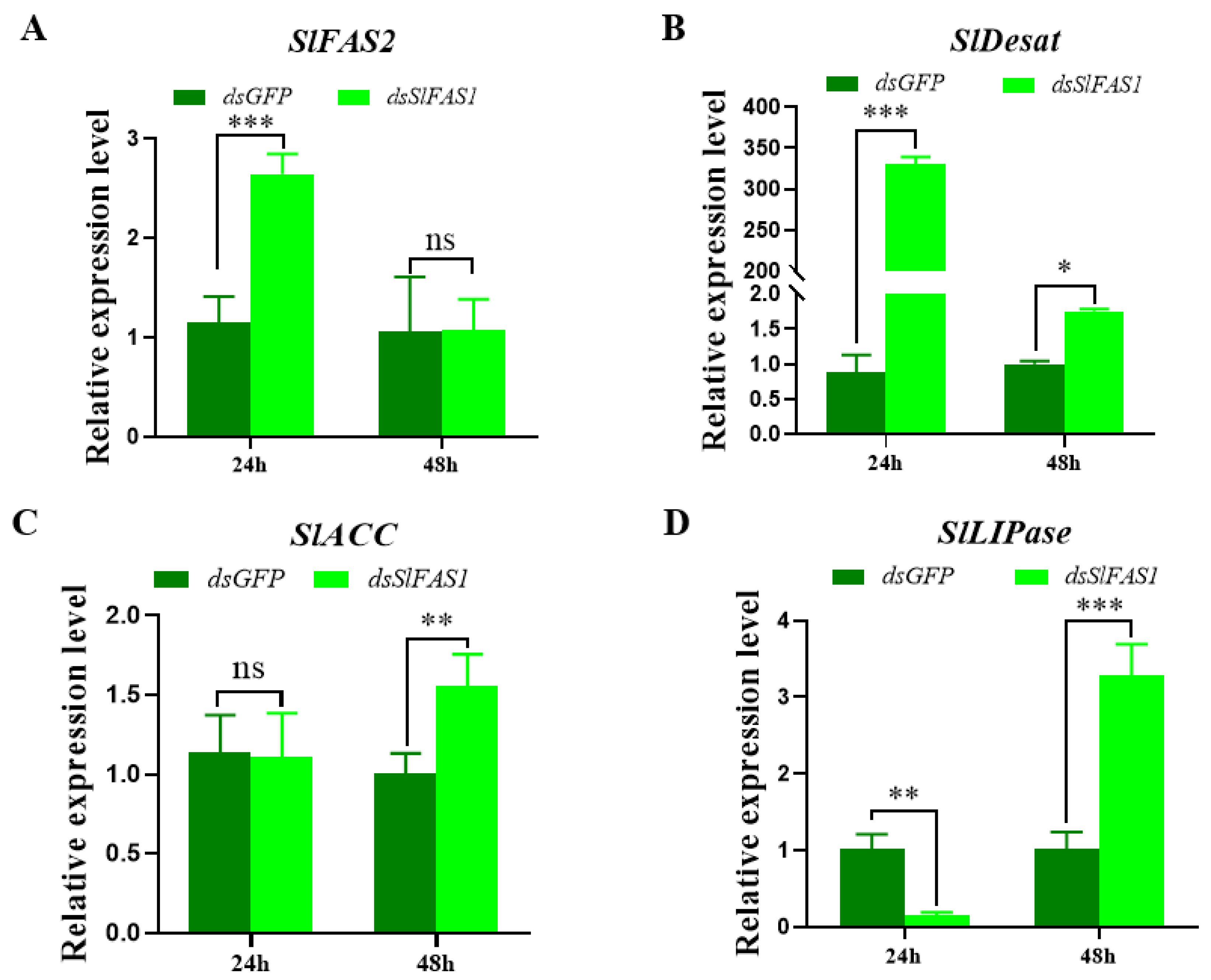
2.6. Effect of Silencing SlFAS1 on the Expression of other Key Genes in the Lipogenesis Pathway
3. Discussion
4. Materials and Methods
4.1. Insects
4.2. Bioinformatics Analysis of FAS Genes in S. litura
4.3. RNA Isolation and Quantitative RT-PCR Validation
4.4. RNA Interference in S. litura Larvae
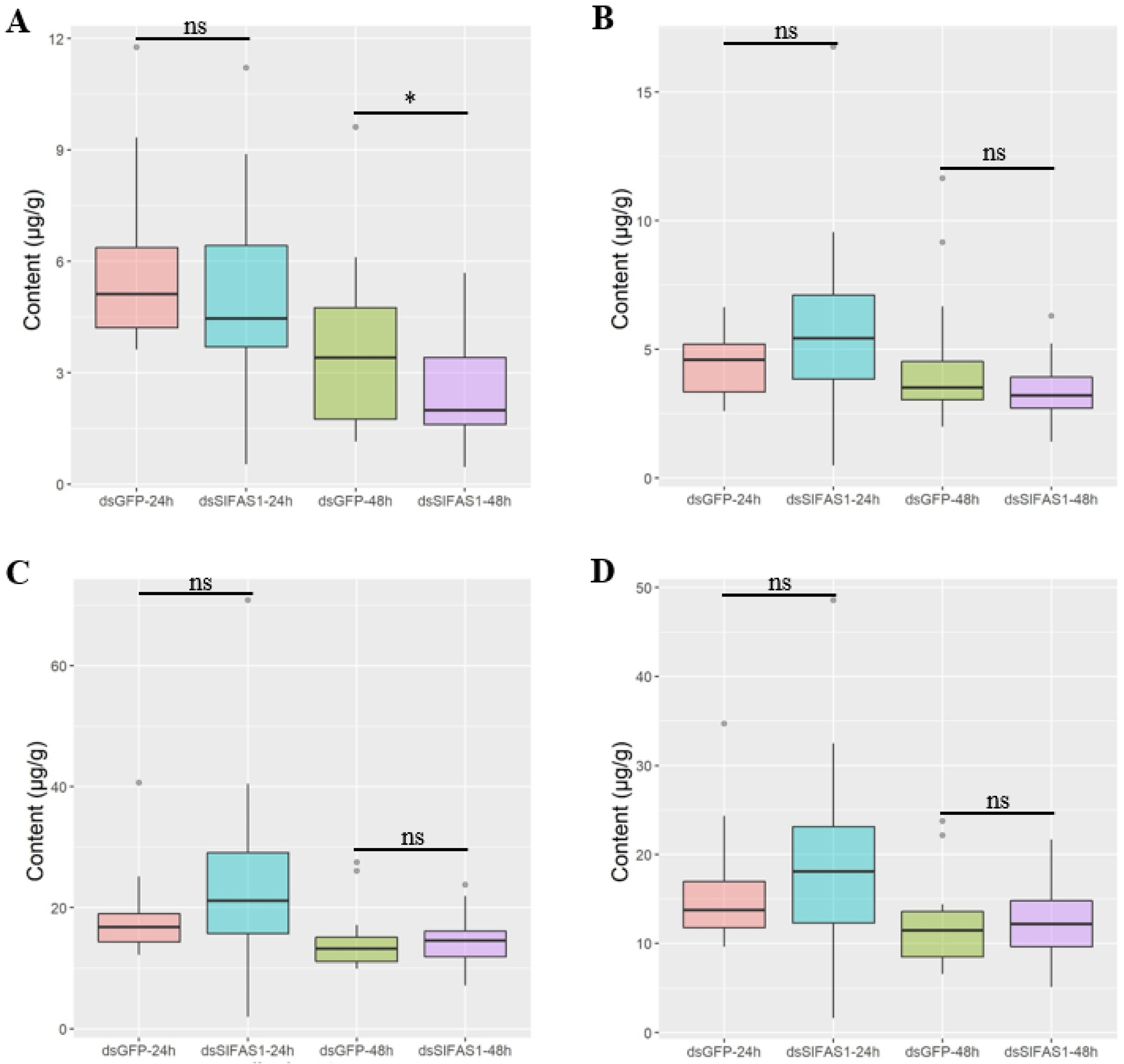
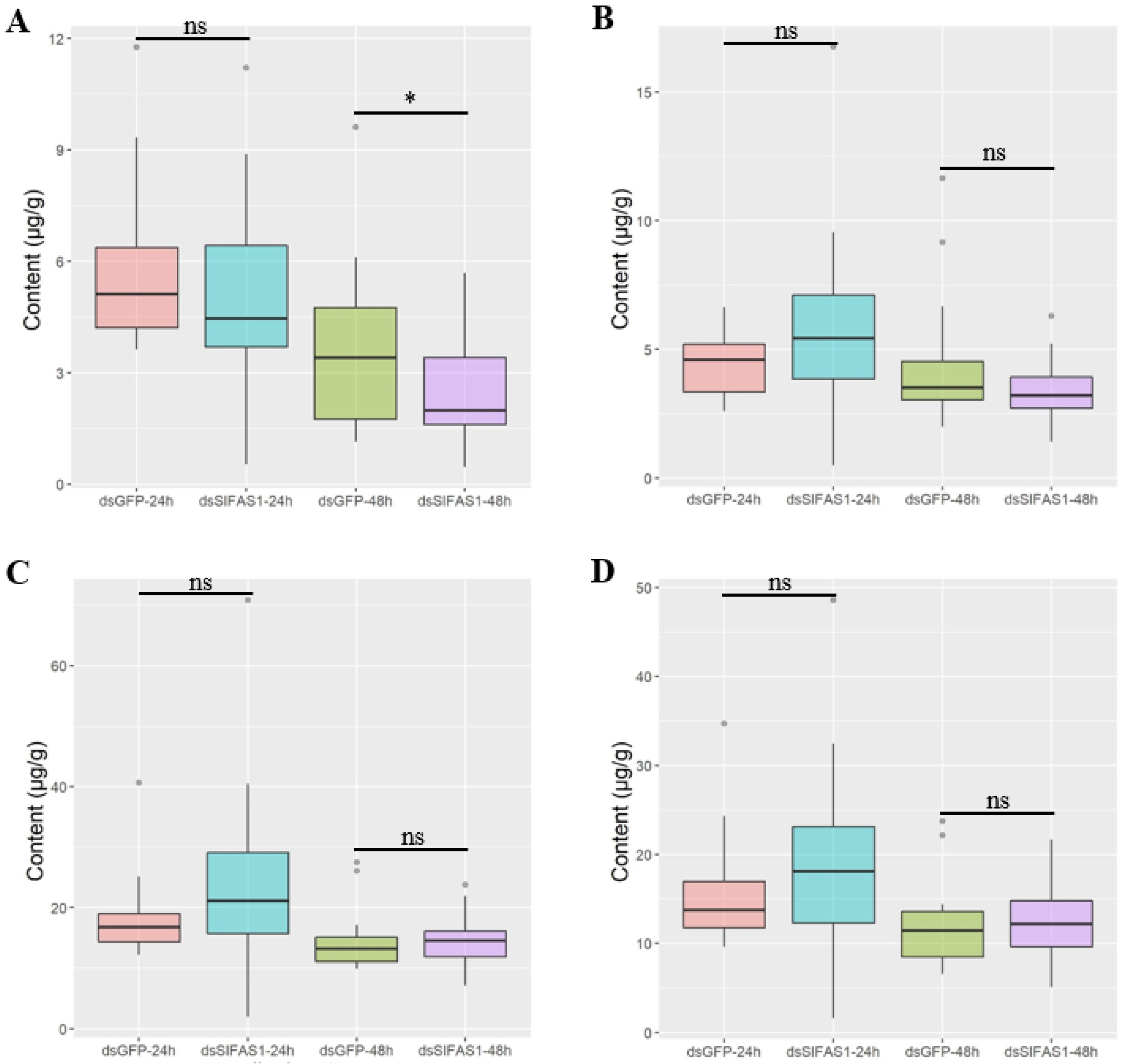
4.5. Determination of Fatty Acid Content
4.6. Measurements of Triglyceride Content
4.7. Nile Red Staining
4.8. Development of S. litura
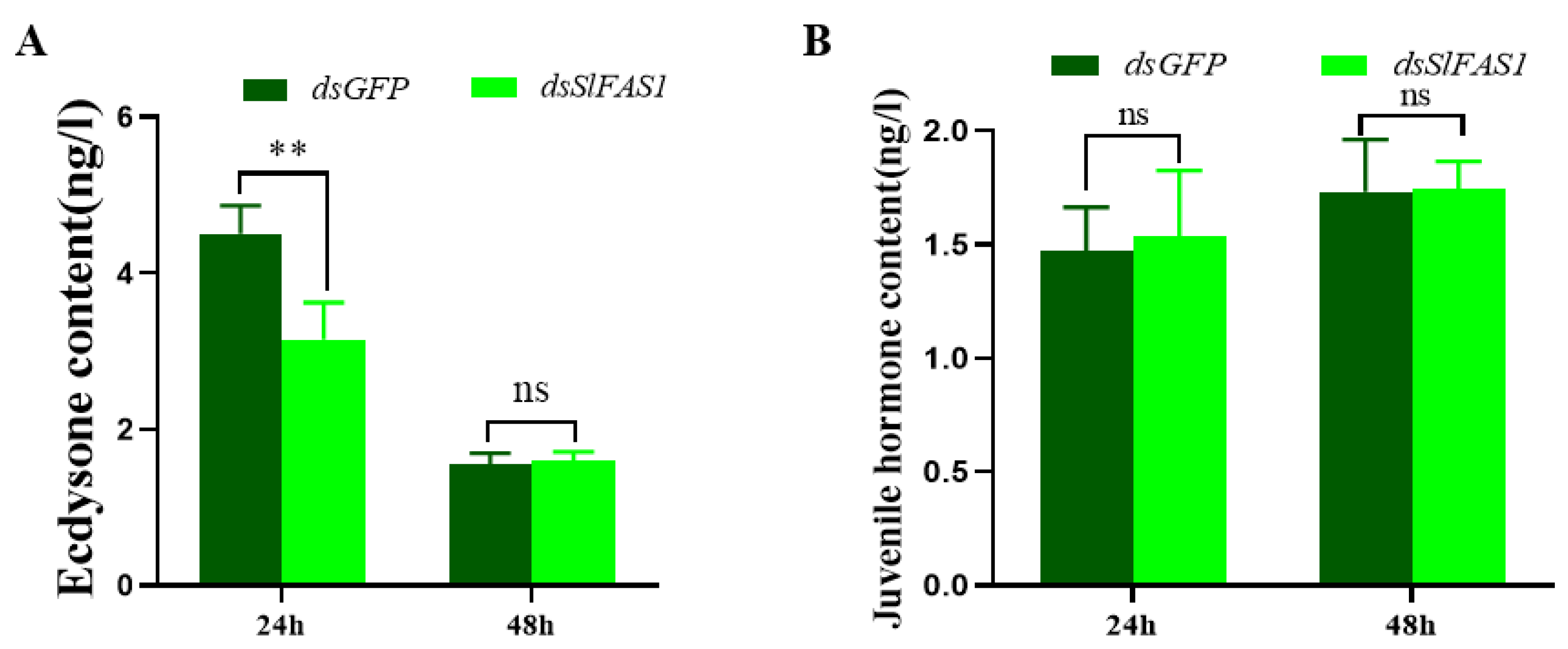
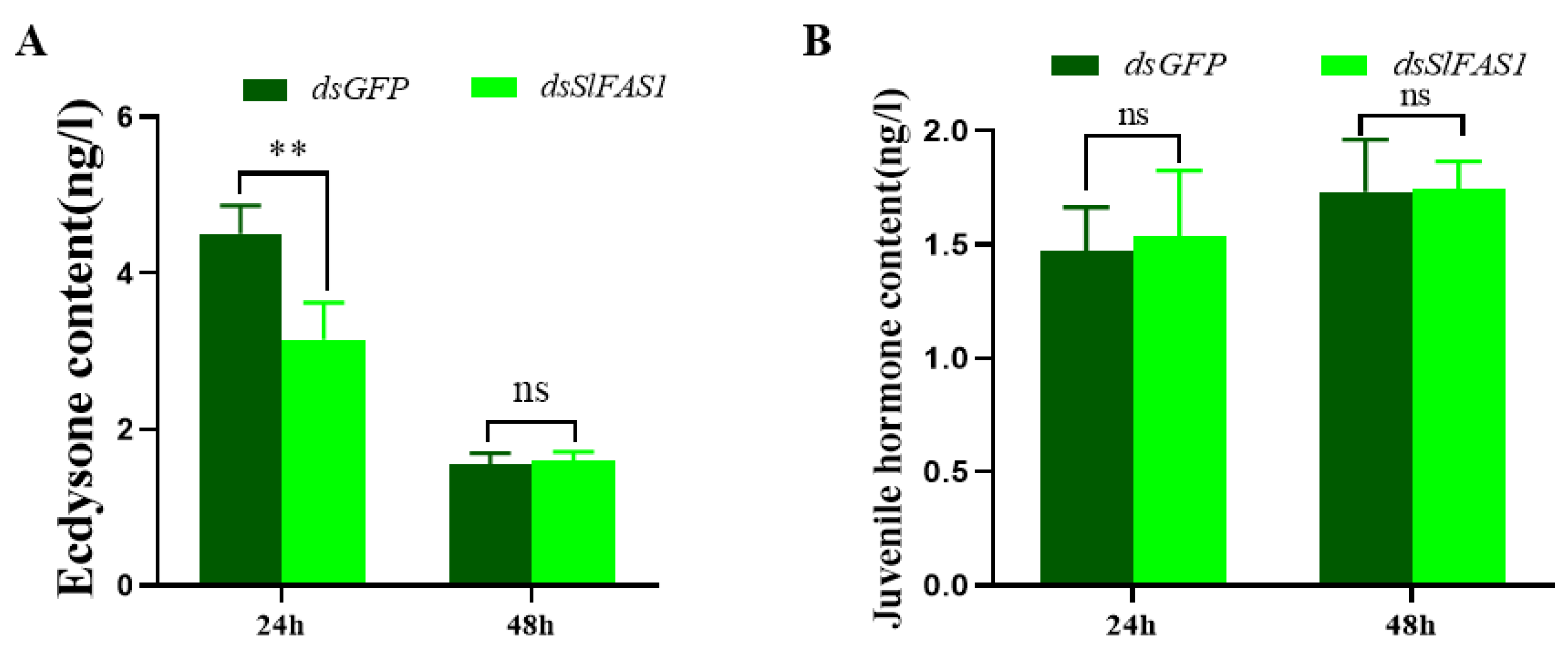
4.9. Content of Juvenile Hormone and Ecdysone in Haemolymph of Fifth Instar S. litura after Silencing SlFAS
4.10. Effect of Silencing SlFAS1 on the Expression of Other Key Genes in the Lipid Synthesis Pathway
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jenni, S.; Leibundgut, M.; Boehringer, D.; Frick, C.; Mikolásek, B.; Ban, N. Structure of fungal fatty acid synthase and impli-cations for iterative substrate shuttling. Science 2007, 316, 254–261. [Google Scholar] [CrossRef]
- Maier, T.; Leibundgut, M.; Ban, N. The Crystal Structure of a Mammalian Fatty Acid Synthase. Science 2008, 321, 1315–1322. [Google Scholar] [CrossRef]
- Finzel, K.; Lee, D.J.; Burkart, M.D. Using Modern Tools To Probe the Structure-Function Relationship of Fatty Acid Synthases. ChemBioChem 2015, 16, 528–547. [Google Scholar] [CrossRef] [PubMed]
- Finck, J.; Berdan, E.L.; Mayer, F.; Ronacher, B.; Geiselhardt, S. Divergence of cuticular hydrocarbons in two sympatric grass-hopper species and the evolution of fatty acid synthases and elongases across insects. Sci. Rep. 2016, 6, 33695. [Google Scholar] [CrossRef] [PubMed]
- Parvy, J.-P.; Napal, L.; Rubin, T.; Poidevin, M.; Perrin, L.; Wicker-Thomas, C.; Montagne, J. Drosophila melanogaster Ace-tyl-CoA-carboxylase sustains a fatty acid-dependent remote signal to waterproof the respiratory system. PLoS Genet. 2012, 8, e1002925. [Google Scholar] [CrossRef]
- Wicker-Thomas, C.; Garrido, D.; Bontonou, G.; Napal, L.; Mazuras, N.; Denis, B.; Rubin, T.; Parvy, J.-P.; Montagne, J. Flexible origin of hydrocarbon/pheromone precursors in Drosophila melanogaster. J. Lipid Res. 2015, 56, 2094–2101. [Google Scholar] [CrossRef]
- Renobales, M.; Woodin, T.S.; Blomquist, G.J. Drosophila melanogaster fatty acid synthetase: Characteristics and effect of protease inhibitors. Insect Biochem. 1986, 16, 887–894. [Google Scholar] [CrossRef]
- Garrido, D.; Rubin, T.; Poidevin, M.; Maroni, B.; Le Rouzic, A.; Parvy, J.-P.; Montagne, J. Fatty Acid Synthase Cooperates with Glyoxalase 1 to Protect against Sugar Toxicity. PLoS Genet. 2015, 11, e1004995. [Google Scholar] [CrossRef]
- Chung, H.; Loehlin, D.W.; Dufour, H.D.; Vaccarro, K.; Millar, J.G.; Carroll, S.B. A single gene affects both ecological di-vergence and mate choice in Drosophila. Science 2014, 343, 1148–1151. [Google Scholar] [CrossRef] [PubMed]
- Arrese, E.L.; Canavoso, L.E.; Jouni, Z.E.; Pennington, J.E.; Tsuchida, K.; Wells, M.A. Lipid storage and mobilization in insects: Current status and future directions. Insect Biochem. Mol. Biol. 2001, 31, 7–17. [Google Scholar] [CrossRef]
- Alabaster, A.; Isoe, J.; Zhou, G.; Lee, A.; Murphy, A.; Day, W.A.; Miesfeld, R.L. Deficiencies in acetyl-CoA carboxylase and fatty acid synthase 1 differentially affect eggshell formation and blood meal digestion in Aedes aegypti. Insect Biochem. Mol. Biol. 2011, 41, 946–955. [Google Scholar] [CrossRef]
- Menendez, J.A.; Lupu, R. Fatty acid synthase and the lipogenic phenotype in cancer pathogenesis. Nat. Rev. Cancer 2007, 7, 763–777. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Song, Y.; Hwarari, D.T.; Liang, X.-H.; Ding, J.-H.; Yan, M.W.; Wu, F.A.; Wang, J.; Sheng, S. Fatty acid synthases and desaturases are essential for the biosynthesis of α-linolenic acid and metamorphosis in a major mulberry pest, Glyphodes pyloalis walker (Lepidoptera: Pyralidae). Pest Manag. Sci. 2022, 78, 2629–2642. [Google Scholar] [CrossRef] [PubMed]
- Gondim, K.C.; Atella, G.C.; Pontes, E.G.; Majerowicz, D. Lipid metabolism in insect disease vectors. Insect Biochem. Mol. Biol. 2018, 101, 108–123. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Hussain, D.; Ghouse, G.; Abbas, M.; Fisher, S.W. Monitoring of insecticide resistance in Spodoptera litura (Lepi-doptera: Noctuidae) from four districts of Punjab, Pakistan to conventional and new chemistry insecticides. Crop Prot. 2016, 79, 177–184. [Google Scholar] [CrossRef]
- Wen, L.; Gao, G.P.; Huang, Z.Q.; Zheng, S.C.; Feng, Q.L.; Liu, L. Expression, regulation and binding affinity of fatty ac-id-binding protein 2 in Spodoptera litura. J. Integr. Agr. 2020, 19, 2095–3119. [Google Scholar] [CrossRef]
- Zhou, J.-M.; Yan, J.; You, K.-K.; Chen, X.; Yuan, Z.-N.; Zhou, Q.; Lu, K. Characterization of a Nilaparvata lugens (Stal) brummer gene and analysis of its role in lipid metabolism. Arch. Insect Biochem. Physiol. 2018, 97, e21442. [Google Scholar] [CrossRef]
- Lambert, C.; Poyart, C.; Gruss, A.; Fouet, A. FabT, a Bacterial Transcriptional Repressor That Limits Futile Fatty Acid Biosynthesis. Microbiol. Mol. Biol. Rev. 2022, 21, e0002922. [Google Scholar] [CrossRef]
- Zhang, X.-Y.; Li, B.; Huang, B.-C.; Wang, F.-B.; Zhang, Y.-Q.; Zhao, S.-G.; Li, M.; Wang, H.-Y.; Yu, X.-J.; Liu, X.-Y.; et al. Production, Biosynthesis, and Commercial Applications of Fatty Acids From Oleaginous Fungi. Front. Nutr. 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.-Q.; Liu, W.; Zhu, F.; Lei, C.-L.; Wang, X.-P. Fatty acid synthase 2 contributes to diapause preparation in a beetle by regulating lipid accumulation and stress tolerance genes expression. Sci. Rep. 2017, 7, 40509. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shen, L.-W.; Xing, X.-R.; Xie, Y.-Q.; Li, Y.-J.; Liu, Z.-X.; Wang, J.; Wu, F.-A.; Sheng, S. Lipid Dynamics, Identification, and Expression Patterns of Fatty Acid Synthase Genes in an Endoparasitoid, Meteorus pulchricornis (Hymenoptera: Braconidae). Int. J. Mol. Sci. 2020, 21, 6228. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect Fat Body: Energy, Metabolism, and Regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- Moriconi, D.E.; Dulbecco, A.B.; Juárez, M.P.; Calderón-Fernández, G.M. A fatty acid synthase gene (FASN3) from the in-tegument tissue of Rhodnius prolixus contributes to cuticle water loss regulation. Insect Mol. Biol. 2019, 28, 850–861. [Google Scholar] [CrossRef]
- Stanley-Samuelson, D.W.; Jurenka, R.A.; Cripps, C.; Blomquist, G.J.; de Renobales, M. Fatty acids in insects: Composition, metabolism, and biological significance. Arch. Insect Biochem. Physiol. 1988, 9, 1–33. [Google Scholar] [CrossRef]
- Dembeck, L.M.; Böröczky, K.; Huang, W.; Schal, C.; Anholt, R.R.H.; Mackay, T.F.C. Genetic architecture of natural variation in cuticular hydrocarbon composition in Drosophila melanogaster. eLife 2015, 4, e09861. [Google Scholar] [CrossRef]
- Pei, X.-J.; Chen, N.; Bai, Y.; Qiao, J.-W.; Li, S.; Fan, Y.-L.; Liu, T.-X. BgFas1: A fatty acid synthase gene required for both hydro-carbon and cuticular fatty acid biosynthesis in the German cockroach, Blattella germanica (L.). Insect Biochem. Mol. Biol. 2019, 112, 103203. [Google Scholar] [CrossRef]
- Paszkiewicz, M.; Sikora, A.; Boguś, M.I.; Włóka, E.; Stepnowski, P.; Gołębiowski, M. Effect of exposure to chlorpyrifos on the cuticular and internal lipid composition of Blattella germanica males. Insect Sci. 2016, 23, 94–104. [Google Scholar] [CrossRef]
- Law, J.H.; Wells, M.A. Insects as biochemical models. J. Biol. Chem. 1989, 264, 16335–16338. [Google Scholar] [CrossRef]
- Zhou, X.; Ling, X.; Guo, H.; Zhu-Salzman, K.; Ge, F.; Sun, Y. Serratia symbiotica Enhances Fatty Acid Metabolism of Pea Aphid to Promote Host Development. Int. J. Mol. Sci. 2021, 22, 5951. [Google Scholar] [CrossRef]
- Cheng, Y.; Lu, T.; Guo, J.; Lin, Z.; Jin, Q.; Zhang, X.; Zou, Z. Helicoverpa armigera miR-2055 regulates lipid metabolism via fatty acid synthase expression. Open Biol. 2022, 12. [Google Scholar] [CrossRef]
- Li, Y.; Paik, Y.-K. A potential role for fatty acid biosynthesis genes during molting and cuticle formation in Caenorhabditis elegans. BMB Rep. 2011, 44, 285–290. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, X.; Niu, N.; Zhao, Y.; Liu, W.; Moussian, B.; Zhang, J. Two fatty acid synthase genes from the integument contribute to cuticular hydrocarbon biosynthesis and cuticle permeability in Locusta migratoria. Insect Mol. Biol. 2020, 29, 555–568. [Google Scholar] [CrossRef]
- Fang, H.; Wang, X.; Liu, X.; Michaud, J.P.; Wu, Y.; Zhang, H.; Li, Y.; Li, Z. Molecular characterization of insulin receptor (IR) in oriental fruit moth, Grapholita molesta (Lepidoptera: Tortricidae), and elucidation of its regulatory roles in glucolipid homeostasis and metamorphosis through interaction with miR-982490. Insect Mol. Biol. 2022. [Google Scholar] [CrossRef]
- Sun, R.; Xu, Y.; Liu, J.; Yang, L.; Cui, G.; Zhong, G.; Yi, X. Proteomic profiling for ovarian development and azadirachtin ex-posure in Spodoptera litura during metamorphosis from pupae to adults. Ecotoxicol. Environ. Saf. 2022, 237, 113548. [Google Scholar] [CrossRef]
- Fu, K.-Y.; Li, Q.; Zhou, L.-T.; Meng, Q.-W.; Lü, F.-G.; Guo, W.-C.; Li, G.-Q. Knockdown of juvenile hormone acid methyl transferase severely affects the performance of Leptinotarsa decemlineata (Say) larvae and adults. Pest Manag. Sci. 2016, 72, 1231–1241. [Google Scholar] [CrossRef]
- Xu, J.-J.; Sheng, Z.-T.; Palli, S.R. Juvenile Hormone and Insulin Regulate Trehalose Homeostasis in the Red Flour Beetle, Tribolium castaneum. PLoS Genet. 2013, 9, e1003535. [Google Scholar] [CrossRef]
- Wang, X.; Hou, Y.; Saha, T.T.; Pei, G.; Raikhel, A.S.; Zou, Z. Hormone and receptor interplay in the regulation of mosquito lipid metabolism. Proc. Natl. Acad. Sci. USA 2017, 114, E2709–E2718. [Google Scholar] [CrossRef]
- Yang, L.; Qiu, L.; Fang, Q.; Wu, S.; Ye, G. A venom protein of ectoparasitoid Pachycrepoideus vindemiae, PvG6PDH, contributes to parasitism by inhibiting host glucose-6-phosphate metabolism. Insect Sci. 2022, 29, 399–410. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wang, J.; Wu, W.; Wang, X.; Wang, M.; Wu, F. An effective GC method for the determination of the fatty acid composition in silkworm pupae oil using a two-step methylation process. J. Serbian Chem. Soc. 2015, 80, 9–20. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Volume 14, pp. 12–21. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Gu, F.; Liu, Z.; Li, Z.; Wu, F.; Sheng, S. The Key Role of Fatty Acid Synthase in Lipid Metabolism and Metamorphic Development in a Destructive Insect Pest, Spodoptera litura (Lepidoptera: Noctuidae). Int. J. Mol. Sci. 2022, 23, 9064. https://doi.org/10.3390/ijms23169064
Song Y, Gu F, Liu Z, Li Z, Wu F, Sheng S. The Key Role of Fatty Acid Synthase in Lipid Metabolism and Metamorphic Development in a Destructive Insect Pest, Spodoptera litura (Lepidoptera: Noctuidae). International Journal of Molecular Sciences. 2022; 23(16):9064. https://doi.org/10.3390/ijms23169064
Chicago/Turabian StyleSong, Yan, Fengming Gu, Zhixiang Liu, Zongnan Li, Fu’an Wu, and Sheng Sheng. 2022. "The Key Role of Fatty Acid Synthase in Lipid Metabolism and Metamorphic Development in a Destructive Insect Pest, Spodoptera litura (Lepidoptera: Noctuidae)" International Journal of Molecular Sciences 23, no. 16: 9064. https://doi.org/10.3390/ijms23169064
APA StyleSong, Y., Gu, F., Liu, Z., Li, Z., Wu, F., & Sheng, S. (2022). The Key Role of Fatty Acid Synthase in Lipid Metabolism and Metamorphic Development in a Destructive Insect Pest, Spodoptera litura (Lepidoptera: Noctuidae). International Journal of Molecular Sciences, 23(16), 9064. https://doi.org/10.3390/ijms23169064