
DNA Methyltransferases: From Evolution to Clinical Applications

 ,
,  and
and
Abstract
1. Introduction
2. DNA Methyltransferases
2.1. The Structure of MTAses
2.2. DNA Cytosine 5′MTases Biology
2.3. Regulating DNA Metiltransferases by Splicing Isoforms
2.4. DNMTs in Embryonic Development
2.5. DNA Methyltransferases Are Regulated by Chemical Compounds and ncRNAs
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oliveira, P.H.; Fang, G. Conserved DNA Methyltransferases: A Window into Fundamental Mechanisms of Epigenetic Regulation in Bacteria. Trends Microbiol. 2021, 29, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Ponger, L.; Li, W.-H. Evolutionary Diversification of DNA Methyltransferases in Eukaryotic Genomes. Mol. Biol. Evol. 2005, 22, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Campos, C.; Valente, L.M.P.; Fernandes, J.M.O. Molecular Evolution of Zebrafish Dnmt3 Genes and Thermal Plasticity of Their Expression during Embryonic Development. Gene 2012, 500, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Mosquera-Rendón, J.; Cárdenas-Brito, S.; Pineda, J.D.; Corredor, M.; Benítez-Páez, A. Evolutionary and Sequence-Based Relationships in Bacterial AdoMet-Dependent Non-Coding RNA Methyltransferases. BMC Res. Notes 2014, 7, 440. [Google Scholar] [CrossRef]
- Zhenilo, S.V.; Sokolov, A.S.; Prokhortchouk, E.B. Epigenetics of Ancient DNA. Acta Nat. 2016, 30, 72–76. [Google Scholar] [CrossRef][Green Version]
- Chédin, F. The DNMT3 Family of Mammalian De Novo DNA Methyltransferases. In Progress in Molecular Biology and Translational Science; Elsevier: Amsterdam, The Netherlands, 2011; Volume 101, pp. 255–285. [Google Scholar]
- Jurkowska, R.Z.; Jurkowski, T.P.; Jeltsch, A. Structure and Function of Mammalian DNA Methyltransferases. Chembiochem 2011, 12, 206–222. [Google Scholar] [CrossRef]
- Lin, W.; Yan, Y.; Ping, S.; Li, P.; Li, D.; Hu, J.; Liu, W.; Wen, X.; Ren, Y. Metformin-Induced Epigenetic Toxicity in Zebrafish: Experimental and Molecular Dynamics Simulation Studies. Environ. Sci. Technol. 2021, 55, 1672–1681. [Google Scholar] [CrossRef]
- Daher-Reyes, G.S.; Merchan, B.M.; Yee, K.W.L. Guadecitabine (SGI-110): An Investigational Drug for the Treatment of Myelodysplastic Syndrome and Acute Myeloid Leukemia. Expert Opin. Investig. Drugs 2019, 28, 835–849. [Google Scholar] [CrossRef]
- Vassena, R.; Dee Schramm, R.; Latham, K.E. Species-Dependent Expression Patterns of DNA Methyltransferase Genes in Mammalian Oocytes and Preimplantation Embryos. Mol. Reprod. Dev. 2005, 72, 430–436. [Google Scholar] [CrossRef]
- Goyal, R. Accuracy of DNA Methylation Pattern Preservation by the Dnmt1 Methyltransferase. Nucleic Acids Res. 2006, 34, 1182–1188. [Google Scholar] [CrossRef] [PubMed]
- Iwanami, N.; Lawir, D.-F.; Sikora, K.; O´Meara, C.; Takeshita, K.; Schorpp, M.; Boehm, T. Transgenerational Inheritance of Impaired Larval T Cell Development in Zebrafish. Nat. Commun. 2020, 11, 4505. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.A.; Hartono, S.R.; Chédin, F. Inactive DNMT3B Splice Variants Modulate De Novo DNA Methylation. PLoS ONE 2013, 8, e69486. [Google Scholar] [CrossRef] [PubMed]
- Foulks, J.M.; Parnell, K.M.; Nix, R.N.; Chau, S.; Swierczek, K.; Saunders, M.; Wright, K.; Hendrickson, T.F.; Ho, K.-K.; McCullar, M.V.; et al. Epigenetic Drug Discovery: Targeting DNA Methyltransferases. J. Biomol. Screen. 2012, 17, 2–17. [Google Scholar] [CrossRef] [PubMed]
- dos Reis, T.F.; Silva, L.P.; de Castro, P.A.; Almeida de Lima, P.B.; do Carmo, R.A.; Marini, M.M.; da Silveira, J.F.; Ferreira, B.H.; Rodrigues, F.; Malavazi, I.; et al. The Influence of Genetic Stability on Aspergillus fumigatus Virulence and Azole Resistance. G3 Genes Genomes Genet. 2018, 8, 265–278. [Google Scholar] [CrossRef]
- Li, J.; Li, C.; Lu, S. Identification and Characterization of the Cytosine-5 DNA Methyltransferase Gene Family in Salvia miltiorrhiza. PeerJ 2018, 6, e4461. [Google Scholar] [CrossRef]
- Cardoso-Júnior, C.A.M.; Eyer, M.; Dainat, B.; Hartfelder, K.; Dietemann, V. Social Context Influences the Expression of DNA Methyltransferase Genes in the Honeybee. Sci. Rep. 2018, 8, 11076. [Google Scholar] [CrossRef]
- Weisenberger, D.J.; Velicescu, M.; Cheng, J.C.; Gonzales, F.A.; Liang, G.; Jones, P.A. Role of the DNA Methyltransferase Variant DNMT3b3 in DNA Methylation. Mol. Cancer Res. 2004, 2, 62–72. [Google Scholar] [CrossRef]
- Jurkowski, T.P.; Jeltsch, A. On the Evolutionary Origin of Eukaryotic DNA Methyltransferases and Dnmt2. PLoS ONE 2011, 6, e28104. [Google Scholar] [CrossRef]
- Bheemanaik, S.; Reddy, Y.V.R.; Rao, D.N. Structure, Function and Mechanism of Exocyclic DNA Methyltransferases. Biochem. J. 2006, 399, 177–190. [Google Scholar] [CrossRef]
- Lee, J.-H.; Voo, K.S.; Skalnik, D.G. Identification and Characterization of the DNA Binding Domain of CpG-Binding Protein. J. Biol. Chem. 2001, 276, 44669–44676. [Google Scholar] [CrossRef]
- Edwards, J.R.; Yarychkivska, O.; Boulard, M.; Bestor, T.H. DNA Methylation and DNA Methyltransferases. Epigenetics Chromatin 2017, 10, 23. [Google Scholar] [CrossRef]
- Xu, T.-H.; Liu, M.; Zhou, X.E.; Liang, G.; Zhao, G.; Xu, H.E.; Melcher, K.; Jones, P.A. Structure of Nucleosome-Bound DNA Methyltransferases DNMT3A and DNMT3B. Nature 2020, 586, 151–155. [Google Scholar] [CrossRef]
- Taverna, S.D.; Li, H.; Ruthenburg, A.J.; Allis, C.D.; Patel, D.J. How Chromatin-Binding Modules Interpret Histone Modifications: Lessons from Professional Pocket Pickers. Nat. Struct. Mol. Biol. 2007, 14, 1025–1040. [Google Scholar] [CrossRef]
- Bestor, T.H. Cytosine Methylation Mediates Sexual Conflict. Trends Genet. 2003, 19, 185–190. [Google Scholar] [CrossRef]
- Fatemi, M.; Hermann, A.; Pradhan, S.; Jeltsch, A. The Activity of the Murine DNA Methyltransferase Dnmt1 Is Controlled by Interaction of the Catalytic Domain with the N-Terminal Part of the Enzyme Leading to an Allosteric Activation of the Enzyme after Binding to Methylated DNA. J. Mol. Biol. 2001, 309, 1189–1199. [Google Scholar] [CrossRef]
- Bestor, T.; Laudano, A.; Mattaliano, R.; Ingram, V. Cloning and Sequencing of a CDNA Encoding DNA Methyltransferase of Mouse Cells. J. Mol. Biol. 1988, 203, 971–983. [Google Scholar] [CrossRef]
- Jia, D.; Jurkowska, R.Z.; Zhang, X.; Jeltsch, A.; Cheng, X. Structure of Dnmt3a Bound to Dnmt3L Suggests a Model for de Novo DNA Methylation. Nature 2007, 449, 248–251. [Google Scholar] [CrossRef]
- Jain, D.; Meydan, C.; Lange, J.; Claeys Bouuaert, C.; Lailler, N.; Mason, C.E.; Anderson, K.V.; Keeney, S. Rahu Is a Mutant Allele of Dnmt3c, Encoding a DNA Methyltransferase Homolog Required for Meiosis and Transposon Repression in the Mouse Male Germline. PLOS Genet. 2017, 13, e1006964. [Google Scholar] [CrossRef]
- Barau, J.; Teissandier, A.; Zamudio, N.; Roy, S.; Nalesso, V.; Hérault, Y.; Guillou, F.; Bourc’his, D. The DNA Methyltransferase DNMT3C Protects Male Germ Cells from Transposon Activity. Science 2016, 354, 909–912. [Google Scholar] [CrossRef]
- Hung, M.-S.; Karthikeyan, N.; Huang, B.; Koo, H.-C.; Kiger, J.; Shen, C.-K.J. Drosophila Proteins Related to Vertebrate DNA (5-Cytosine) Methyltransferases. Proc. Natl. Acad. Sci. USA 1999, 96, 11940–11945. [Google Scholar] [CrossRef]
- Tweedie, S.; Ng, H.-H.; Barlow, A.L.; Turner, B.M.; Hendrich, B.; Bird, A. Vestiges of a DNA Methylation System in Drosophila Melanogaster? Nat. Genet. 1999, 23, 389–390. [Google Scholar] [CrossRef]
- Vieira, G.C.; D’Ávila, M.F.; Zanini, R.; Deprá, M.; da Silva Valente, V.L. Evolution of DNMT2 in Drosophilids: Evidence for Positive and Purifying Selection and Insights into New Protein (Pathways) Interactions. Genet. Mol. Biol. 2018, 41, 215–234. [Google Scholar] [CrossRef]
- Capuano, F.; Mülleder, M.; Kok, R.; Blom, H.J.; Ralser, M. Cytosine DNA Methylation Is Found in Drosophila melanogaster but Absent in Saccharomyces cerevisiae, Schizosaccharomyces pombe, and Other Yeast Species. Anal. Chem. 2014, 86, 3697–3702. [Google Scholar] [CrossRef]
- Simpson, V.J.; Johnson, T.E.; Hammen, R.F. Caenorhabditis elegans DNA Does Not Contain 5-Methylcytosine at Any Time during Development or Aging. Nucleic Acids Res. 1986, 14, 6711–6719. [Google Scholar] [CrossRef]
- Mouammine, A.; Collier, J. The Impact of DNA Methylation in Alphaproteobacteria: Suppressor Mutations in FgSNU66. Mol. Microbiol. 2018, 110, 1–10. [Google Scholar] [CrossRef]
- Schaefer, M.; Lyko, F. Solving the Dnmt2 Enigma. Chromosoma 2010, 119, 35–40. [Google Scholar] [CrossRef]
- Babinger, P.; Völkl, R.; Cakstina, I.; Maftei, A.; Schmitt, R. Maintenance DNA Methyltransferase (Met1) and Silencing of CpG-Methylated Foreign DNA in Volvox Carteri. Plant Mol. Biol. 2007, 63, 325–336. [Google Scholar] [CrossRef]
- Taşkin, K.M.; Özbilen, A.; Sezer, F.; Hürkan, K.; Güneş, Ş. Structure and Expression of Dna Methyltransferase Genes from Apomictic and Sexual Boechera Species. Comput. Biol. Chem. 2017, 67, 15–21. [Google Scholar] [CrossRef]
- Sugiyama, K.; Furusawa, H.; Grúz, P.; Honma, M. Functional Role of DNA Methylation at the FLO1 Promoter in Budding Yeast. FEMS Microbiol. Lett. 2017, 364, fnx221. [Google Scholar] [CrossRef]
- Manfrão-Netto, J.H.C.; Mello-de-Sousa, T.M.; Mach-Aigner, A.R.; Mach, R.L.; Poças-Fonseca, M.J. The DNA-Methyltransferase Inhibitor 5-Aza-2-Deoxycytidine Affects Humicola Grisea Enzyme Activities and the Glucose-Mediated Gene Repression. J. Basic Microbiol. 2018, 58, 144–153. [Google Scholar] [CrossRef]
- Zutz, C.; Gacek, A.; Sulyok, M.; Wagner, M.; Strauss, J.; Rychli, K. Small Chemical Chromatin Effectors Alter Secondary Metabolite Production in Aspergillus Clavatus. Toxins 2013, 5, 1723–1741. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T. One-Way Control of FWA Imprinting in Arabidopsis Endosperm by DNA Methylation. Science 2004, 303, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Kay, S.; Skowronski, D.; Hunt, B.G. Developmental DNA Methyltransferase Expression in the Fire Ant Solenopsis invicta: Fire Ant Developmental DNMT Expression. Insect Sci. 2018, 25, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Pratx, L.; Rancurel, C.; Da Rocha, M.; Danchin, E.G.J.; Castagnone-Sereno, P.; Abad, P.; Perfus-Barbeoch, L. Genome-Wide Expert Annotation of the Epigenetic Machinery of the Plant-Parasitic Nematodes Meloidogyne Spp., with a Focus on the Asexually Reproducing Species. BMC Genomics 2018, 19, 321. [Google Scholar] [CrossRef] [PubMed]
- Firmino, J.; Carballo, C.; Armesto, P.; Campinho, M.A.; Power, D.M.; Manchado, M. Phylogeny, Expression Patterns and Regulation of DNA Methyltransferases in Early Development of the Flatfish, Solea Senegalensis. BMC Dev. Biol. 2017, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Goll, M.G.; Halpern, M.E. DNA Methylation in Zebrafish. In Progress in Molecular Biology and Translational Science; Elsevier: Amsterdam, The Netherlands, 2011; Volume 101, pp. 193–218. [Google Scholar]
- Rana, A.K.; Ankri, S. Reviving the RNA World: An Insight into the Appearance of RNA Methyltransferases. Front. Genet. 2016, 7, 99. [Google Scholar] [CrossRef]
- Que, Q.; Zhang, Y.; Nelson, M.; Ropp, S.; Burbank, D.E.; Van Etten, J.L. Chlorella Virus SC-1A Encodes at Least Five Functional and One Nonfunctional DNA Methyltransferases. Gene 1997, 190, 237–244. [Google Scholar] [CrossRef]
- Kan, T.-N.J.; Li, L. Cloning, Sequencing, Overproduction, and Purification of M CviBI (GANTC) Methyltransferase from Chlorella Virus NC-1A. Gene 1992, 121, 1–7. [Google Scholar] [CrossRef]
- Decewicz, P.; Radlinska, M.; Dziewit, L. Characterization of Sinorhizobium Sp. LM21 Prophages and Virus-Encoded DNA Methyltransferases in the Light of Comparative Genomic Analyses of the Sinorhizobial Virome. Viruses 2017, 9, 161. [Google Scholar] [CrossRef]
- Zeng, C.; Wu, A.; Wang, Y.; Xu, S.; Tang, Y.; Jin, X.; Wang, S.; Qin, L.; Sun, Y.; Fan, C.; et al. Identification and Characterization of a Ribose 2′- O -Methyltransferase Encoded by the Ronivirus Branch of Nidovirales. J. Virol. 2016, 90, 6675–6685. [Google Scholar] [CrossRef]
- Case, J.B.; Ashbrook, A.W.; Dermody, T.S.; Denison, M.R. Mutagenesis of S -Adenosyl-l-Methionine-Binding Residues in Coronavirus Nsp14 N7-Methyltransferase Demonstrates Differing Requirements for Genome Translation and Resistance to Innate Immunity. J. Virol. 2016, 90, 7248–7256. [Google Scholar] [CrossRef]
- Martin, B.; Coutard, B.; Guez, T.; Paesen, G.C.; Canard, B.; Debart, F.; Vasseur, J.-J.; Grimes, J.M.; Decroly, E. The Methyltransferase Domain of the Sudan Ebolavirus L Protein Specifically Targets Internal Adenosines of RNA Substrates, in Addition to the Cap Structure. Nucleic Acids Res. 2018, 46, 7902–7912. [Google Scholar] [CrossRef]
- Li, H.; Yang, F.; Gao, B.; Yu, Z.; Liu, X.; Xie, F.; Zhang, J. Hepatitis B Virus Infection in Hepatocellular Carcinoma Tissues Upregulates Expression of DNA Methyltransferases. Int. J. Clin. Exp. Med. 2015, 8, 4175. [Google Scholar]
- Pazienza, V.; Panebianco, C.; Andriulli, A. Hepatitis Viruses Exploitation of Host DNA Methyltransferases Functions. Clin. Exp. Med. 2016, 16, 265–272. [Google Scholar] [CrossRef]
- Tao, Q. Defective de Novo Methylation of Viral and Cellular DNA Sequences in ICF Syndrome Cells. Hum. Mol. Genet. 2002, 11, 2091–2102. [Google Scholar] [CrossRef]
- Ksiaa, F.; Ziadi, S.; Gacem, R.B.; Dhiab, M.B.; Trimeche, M. Correlation between DNA Methyltransferases Expression and Epstein-Barr Virus, JC Polyomavirus and Helicobacter Pylori Infections in Gastric Carcinomas. Neoplasma 2014, 61, 710–717. [Google Scholar] [CrossRef]
- Chong, J.-M.; Sakuma, K.; Sudo, M.; Ushiku, T.; Uozaki, H.; Shibahara, J.; Nagai, H.; Funata, N.; Taniguchi, H.; Aburatani, H.; et al. Global and Non-Random CpG-Island Methylation in Gastric Carcinoma Associated with Epstein-Barr Virus. Cancer Sci. 2003, 94, 76–80. [Google Scholar] [CrossRef]
- Shamay, M.; Krithivas, A.; Zhang, J.; Hayward, S.D. Recruitment of the de Novo DNA Methyltransferase Dnmt3a by Kaposi’s Sarcoma-Associated Herpesvirus LANA. Proc. Natl. Acad. Sci. USA 2006, 103, 14554–14559. [Google Scholar] [CrossRef]
- Fang, J.-Y.; Mikovits, J.A.; Bagni, R.; Petrow-Sadowski, C.L.; Ruscetti, F.W. Infection of Lymphoid Cells by Integration-Defective Human Immunodeficiency Virus Type 1 Increases De Novo Methylation. J. Virol. 2001, 75, 9753–9761. [Google Scholar] [CrossRef]
- Tock, M.R.; Dryden, D.T. The Biology of Restriction and Anti-Restriction. Curr. Opin. Microbiol. 2005, 8, 466–472. [Google Scholar] [CrossRef]
- Seong, H.J.; Han, S.-W.; Sul, W.J. Prokaryotic DNA Methylation and Its Functional Roles. J. Microbiol. 2021, 59, 242–248. [Google Scholar] [CrossRef]
- Ouellette, M.; Gogarten, J.; Lajoie, J.; Makkay, A.; Papke, R. Characterizing the DNA Methyltransferases of Haloferax Volcanii via Bioinformatics, Gene Deletion, and SMRT Sequencing. Genes 2018, 9, 129. [Google Scholar] [CrossRef]
- Wu, J.; Jia, Q.; Wu, S.; Zeng, H.; Sun, Y.; Wang, C.; Ge, R.; Xie, W. The Crystal Structure of the Pyrococcus Abyssi Mono-Functional Methyltransferase PaTrm5b. Biochem. Biophys. Res. Commun. 2017, 493, 240–245. [Google Scholar] [CrossRef]
- Pampa, K.J.; Madan Kumar, S.; Hema, M.K.; Kumara, K.; Naveen, S.; Kunishima, N.; Lokanath, N.K. Crystal Structure of SAM-Dependent Methyltransferase from Pyrococcus horikoshii. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2017, 73, 706–712. [Google Scholar] [CrossRef]
- Maynard-Smith, M.D.; McKelvie, J.C.; Wood, R.J.; Harmer, J.E.; Ranasinghe, R.T.; Williams, C.L.; Coomber, D.M.; Stares, A.F.; Roach, P.L. Direct and Continuous Fluorescence-Based Measurements of Pyrococcus Horikoshii DNA N-6 Adenine Methyltransferase Activity. Anal. Biochem. 2011, 418, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Currie, M.A.; Brown, G.; Wong, A.; Ohira, T.; Sugiyama, K.; Suzuki, T.; Yakunin, A.F.; Jia, Z. Structural and Functional Characterization of the TYW3/Taw3 Class of SAM-Dependent Methyltransferases. RNA 2017, 23, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Bestor, T.H.; Verdine, G.L. DNA Methylransferases. Curr. Opin. Cell Biol. 1994, 6, 380–389. [Google Scholar] [CrossRef]
- Casselli, T.; Tourand, Y.; Scheidegger, A.; Arnold, W.; Proulx, A.; Stevenson, B.; Brissette, C.A. DNA Methylation by Restriction Modification Systems Affects the Global Transcriptome Profile in Borrelia burgdorferi. J. Bacteriol. 2018, 200, e00395-18. [Google Scholar] [CrossRef] [PubMed]
- Forde, B.M.; Phan, M.-D.; Gawthorne, J.A.; Ashcroft, M.M.; Stanton-Cook, M.; Sarkar, S.; Peters, K.M.; Chan, K.-G.; Chong, T.M.; Yin, W.-F.; et al. Lineage-Specific Methyltransferases Define the Methylome of the Globally Disseminated Escherichia Coli ST131 Clone. MBio 2015, 6, e01602-15. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Tanneeru, K.; Guruprasad, L. Structure and Dynamics of H. Pylori 98-10 C5-Cytosine Specific DNA Methyltransferase in Complex with S-Adenosyl- l -Methionine and DNA. Mol. Biosyst. 2016, 12, 3111–3123. [Google Scholar] [CrossRef]
- Grover, S.; Gupta, P.; Kahlon, P.S.; Goyal, S.; Grover, A.; Dalal, K.; Sabeeha, S.; Ehtesham, N.Z.; Hasnain, S.E. Analyses of Methyltransferases across the Pathogenicity Spectrum of Different Mycobacterial Species Point to an Extremophile Connection. Mol. Biosyst. 2016, 12, 1615–1625. [Google Scholar] [CrossRef]
- Brocklehurst, S.; Watson, M.; Carr, I.M.; Out, S.; Heidmann, I.; Meyer, P. Induction of Epigenetic Variation in Arabidopsis by Over-Expression of DNA Methyltransferase1 (MET1). PLoS ONE 2018, 13, e0192170. [Google Scholar] [CrossRef]
- Sun, Q.; Qiao, J.; Zhang, S.; He, S.; Shi, Y.; Yuan, Y.; Zhang, X.; Cai, Y. Changes in DNA Methylation Assessed by Genomic Bisulfite Sequencing Suggest a Role for DNA Methylation in Cotton Fruiting Branch Development. PeerJ 2018, 6, e4945. [Google Scholar] [CrossRef]
- Gianoglio, S.; Moglia, A.; Acquadro, A.; Comino, C.; Portis, E. The Genome-Wide Identification and Transcriptional Levels of DNA Methyltransferases and Demethylases in Globe Artichoke. PLoS ONE 2017, 12, e0181669. [Google Scholar] [CrossRef]
- Kumar, R.; Chauhan, P.K.; Khurana, A. Identification and Expression Profiling of DNA Methyltransferases during Development and Stress Conditions in Solanaceae. Funct. Integr. Genomics 2016, 16, 513–528. [Google Scholar] [CrossRef]
- Garg, R.; Kumari, R.; Tiwari, S.; Goyal, S. Genomic Survey, Gene Expression Analysis and Structural Modeling Suggest Diverse Roles of DNA Methyltransferases in Legumes. PLoS ONE 2014, 9, e88947. [Google Scholar] [CrossRef]
- Zhang, J.J.; Yang, H. Metabolism and Detoxification of Pesticides in Plants. Sci. Total Environ. 2021, 790, 148034. [Google Scholar] [CrossRef]
- Nai, Y.-S.; Huang, Y.-C.; Yen, M.-R.; Chen, P.-Y. Diversity of Fungal DNA Methyltransferases and Their Association With DNA Methylation Patterns. Front. Microbiol. 2021, 11, 616922. [Google Scholar] [CrossRef]
- Liu, S.-Y.; Lin, J.-Q.; Wu, H.-L.; Wang, C.-C.; Huang, S.-J.; Luo, Y.-F.; Sun, J.-H.; Zhou, J.-X.; Yan, S.-J.; He, J.-G.; et al. Bisulfite Sequencing Reveals That Aspergillus Flavus Holds a Hollow in DNA Methylation. PLoS ONE 2012, 7, e30349. [Google Scholar] [CrossRef]
- Fujihara, Y.; Miyasako, H.; Kato, K.; Hayashi, T.; Toraya, T. Molecular Cloning, Expression, and Characterization of Starfish DNA (Cytosine-5)-Methyltransferases. Biosci. Biotechnol. Biochem. 2012, 76, 1661–1671. [Google Scholar] [CrossRef]
- Dattani, A.; Sridhar, D.; Aziz Aboobaker, A. Planarian Flatworms as a New Model System for Understanding the Epigenetic Regulation of Stem Cell Pluripotency and Differentiation. Semin. Cell Dev. Biol. 2019, 87, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Riviere, G.; Wu, G.-C.; Fellous, A.; Goux, D.; Sourdaine, P.; Favrel, P. DNA Methylation Is Crucial for the Early Development in the Oyster C. Gigas. Mar. Biotechnol. 2013, 15, 739–753. [Google Scholar] [CrossRef] [PubMed]
- Pearce, K.; Cai, D.; Roberts, A.C.; Glanzman, D.L. Role of Protein Synthesis and DNA Methylation in the Consolidation and Maintenance of Long-Term Memory in Aplysia. Elife 2017, 6, e18299. [Google Scholar] [CrossRef]
- Kotsarenko, K.; Vechtova, P.; Hammerova, Z.; Langova, N.; Malinovska, L.; Wimmerova, M.; Sterba, J.; Grubhoffer, L. Newly Identified DNA Methyltransferases of Ixodes Ricinus Ticks. Ticks Tick-Borne Dis. 2020, 11, 101348. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xing, Y.; Li, Y.; Yin, C.; Ge, C.; Li, F. DNA Methyltransferases Have an Essential Role in Female Fecundity in Brown Planthopper, Nilaparvata Lugens. Biochem. Biophys. Res. Commun. 2015, 464, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Wang, C.; Nieh, J.C.; Tan, K. Inhibiting DNA Methylation Alters Olfactory Extinction but Not Acquisition Learning in Apis Cerana and Apis Mellifera. J. Insect Physiol. 2016, 90, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Biergans, S.D.; Giovanni Galizia, C.; Reinhard, J.; Claudianos, C. Dnmts and Tet Target Memory-Associated Genes after Appetitive Olfactory Training in Honey Bees. Sci. Rep. 2015, 5, 16223. [Google Scholar] [CrossRef] [PubMed]
- Roellig, D.; Bronner, M.E. The Epigenetic Modifier DNMT3A Is Necessary for Proper Otic Placode Formation. Dev. Biol. 2016, 411, 294–300. [Google Scholar] [CrossRef]
- Bahar Halpern, K.; Vana, T.; Walker, M.D. Paradoxical Role of DNA Methylation in Activation of FoxA2 Gene Expression during Endoderm Development. J. Biol. Chem. 2014, 289, 23882–23892. [Google Scholar] [CrossRef]
- Shimoda, N.; Yamakoshi, K.; Miyake, A.; Takeda, H. Identification of a Gene Required for de Novo DNA Methylation of the Zebrafishno Tail Gene. Dev. Dyn. 2005, 233, 1509–1516. [Google Scholar] [CrossRef]
- Takayama, K.; Shimoda, N.; Takanaga, S.; Hozumi, S.; Kikuchi, Y. Expression Patterns of Dnmt3aa, Dnmt3ab, and Dnmt4 during Development and Fin Regeneration in Zebrafish. Gene Expr. Patterns 2014, 14, 105–110. [Google Scholar] [CrossRef]
- Smith, T.H.L.; Collins, T.M.; McGowan, R.A. Expression of the Dnmt3 Genes in Zebrafish Development: Similarity to Dnmt3a and Dnmt3b. Dev. Genes Evol. 2011, 220, 347–353. [Google Scholar] [CrossRef]
- Wang, F.; Qin, Z.; Li, Z.; Yang, S.; Gao, T.; Sun, L.; Wang, D. Dnmt3aa but Not Dnmt3ab Is Required for Maintenance of Gametogenesis in Nile Tilapia (Oreochromis niloticus). Int. J. Mol. Sci. 2021, 22, 10170. [Google Scholar] [CrossRef]
- Giannetto, A.; Nagasawa, K.; Fasulo, S.; Fernandes, J.M.O. Influence of Photoperiod on Expression of DNA (Cytosine-5) Methyltransferases in Atlantic Cod. Gene 2013, 519, 222–230. [Google Scholar] [CrossRef]
- Rodriguez-Osorio, N.; Wang, H.; Rupinski, J.; Bridges, S.M.; Memili, E. Comparative Functional Genomics of Mammalian DNA Methyltransferases. Reprod. Biomed. Online 2010, 20, 243–255. [Google Scholar] [CrossRef][Green Version]
- Okano, M.; Xie, S.; Li, E. Cloning and Characterization of a Family of Novel Mammalian DNA (Cytosine-5) Methyltransferases. Nat. Genet. 1998, 19, 219–220. [Google Scholar] [CrossRef]
- Gujar, H.; Weisenberger, D.; Liang, G. The Roles of Human DNA Methyltransferases and Their Isoforms in Shaping the Epigenome. Genes 2019, 10, 172. [Google Scholar] [CrossRef]
- Xie, S.; Wang, Z.; Okano, M.; Nogami, M.; Li, Y.; He, W.W.; Katsuzumi, O.; En, L. Cloning, Expression and Chromosome Locations of the Human DNMT3 gene family. Gene 1999, 236, 87–95. [Google Scholar] [CrossRef]
- Plourde, K.V.; Labrie, Y.; Ouellette, G.; Pouliot, M.-C.; Durocher, F. Genome-Wide Methylation Analysis of DNMT3B Gene Isoforms Revealed Specific Methylation Profiles in Breast Cell Lines. Epigenomics 2016, 8, 1209–1226. [Google Scholar] [CrossRef]
- Aoki, A. Enzymatic Properties of de Novo-Type Mouse DNA (Cytosine-5) Methyltransferases. Nucleic Acids Res. 2001, 29, 3506–3512. [Google Scholar] [CrossRef]
- Gopalakrishna-Pillai, S.; Iverson, L.E. A DNMT3B Alternatively Spliced Exon and Encoded Peptide Are Novel Biomarkers of Human Pluripotent Stem Cells. PLoS ONE 2011, 6, e20663. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Kanai, Y.; Sakamoto, M.; Saito, H.; Ishii, H.; Hirohashi, S. Overexpression of a Splice Variant of DNA Methyltransferase 3b, DNMT3b4, Associated with DNA Hypomethylation on Pericentromeric Satellite Regions during Human Hepatocarcinogenesis. Proc. Natl. Acad. Sci. USA 2002, 99, 10060–10065. [Google Scholar] [CrossRef] [PubMed]
- Shao, G.; Zhang, R.; Zhang, S.; Jiang, S.; Liu, Y.; Zhang, W.; Zhang, Y.; Li, J.; Gong, K.; Hu, X.-R.; et al. Splice Variants DNMT3B4 and DNMT3B7 Overexpression Inhibit Cell Proliferation in 293A Cell Line. In Vitro Cell. Dev. Biol.-Anim. 2013, 49, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Robertson, K.D.; Uzvolgyi, E.; Liang, G.; Talmadge, C.; Sumegi, J.; Gonzales, F.A.; Jones, P.A. The Human DNA Methyltransferases (DNMTs) 1, 3a and 3b: Coordinate MRNA Expression in Normal Tissues and Overexpression in Tumors. Nucleic Acids Res. 1999, 27, 2291–2298. [Google Scholar] [CrossRef]
- Teodoridis, J.M.; Hall, J.; Marsh, S.; Kannall, H.D.; Smyth, C.; Curto, J.; Siddiqui, N.; Gabra, H.; McLeod, H.L.; Strathdee, G.; et al. CpG Island Methylation of DNA Damage Response Genes in Advanced Ovarian Cancer. Cancer Res. 2005, 65, 8961–8967. [Google Scholar] [CrossRef]
- Ostler, K.R.; Yang, Q.; Looney, T.J.; Zhang, L.; Vasanthakumar, A.; Tian, Y.; Kocherginsky, M.; Raimondi, S.L.; DeMaio, J.G.; Salwen, H.R.; et al. Truncated DNMT3B Isoform DNMT3B7 Suppresses Growth, Induces Differentiation, and Alters DNA Methylation in Human Neuroblastoma. Cancer Res. 2012, 72, 4714–4723. [Google Scholar] [CrossRef]
- Brambert, P.R.; Kelpsch, D.J.; Hameed, R.; Desai, C.V.; Calafiore, G.; Godley, L.A.; Raimondi, S.L. DNMT3B7 Expression Promotes Tumor Progression to a More Aggressive Phenotype in Breast Cancer Cells. PLoS ONE 2015, 10, e0117310. [Google Scholar] [CrossRef][Green Version]
- Shah, M.Y.; Vasanthakumar, A.; Barnes, N.Y.; Figueroa, M.E.; Kamp, A.; Hendrick, C.; Ostler, K.R.; Davis, E.M.; Lin, S.; Anastasi, J.; et al. DNMT3B7, a Truncated DNMT3B Isoform Expressed in Human Tumors, Disrupts Embryonic Development and Accelerates Lymphomagenesis. Cancer Res. 2010, 70, 5840–5850. [Google Scholar] [CrossRef]
- Ma, M.Z.; Lin, R.; Carrillo, J.; Bhutani, M.; Pathak, A.; Ren, H.; Li, Y.; Song, J.; Mao, L. ∆ DNMT3B4-Del Contributes to Aberrant DNA Methylation Patterns in Lung Tumorigenesis. EBioMedicine 2015, 2, 1340–1350. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Van Emburgh, B.O.; Shan, J.; Su, Z.; Fields, C.R.; Vieweg, J.; Hamazaki, T.; Schwartz, P.H.; Terada, N.; Robertson, K.D. A Novel DNMT3B Splice Variant Expressed in Tumor and Pluripotent Cells Modulates Genomic DNA Methylation Patterns and Displays Altered DNA Binding. Mol. Cancer Res. 2009, 7, 1622–1634. [Google Scholar] [CrossRef]
- Wang, J.; Walsh, G.; Liu, D.D.; Lee, J.J.; Mao, L. Expression of ΔDNMT3B Variants and Its Association with Promoter Methylation of P16 and RASSF1A in Primary Non–Small Cell Lung Cancer. Cancer Res. 2006, 66, 8361–8366. [Google Scholar] [CrossRef]
- Wang, J.; Bhutani, M.; Pathak, A.K.; Lang, W.; Ren, H.; Jelinek, J.; He, R.; Shen, L.; Issa, J.-P.; Mao, L. ΔDNMT3B Variants Regulate DNA Methylation in a Promoter-Specific Manner. Cancer Res. 2007, 67, 10647–10652. [Google Scholar] [CrossRef]
- Su, X.; Lv, C.; Qiao, F.; Qiu, X.; Huang, W.; Wu, Q.; Zhao, Z.; Fan, H. Expression Pattern and Clinical Significance of DNA Methyltransferase 3B Variants in Gastric Carcinoma. Oncol. Rep. 2010, 23, 819–826. [Google Scholar]
- Liu, Y.; Sun, L.; Fong, P.; Yang, J.; Zhang, Z.; Yin, S.; Jiang, S.; Liu, X.; Ju, H.; Huang, L.; et al. An Association between Overexpression of DNA Methyltransferase 3B4 and Clear Cell Renal Cell Carcinoma. Oncotarget 2017, 8. [Google Scholar] [CrossRef]
- Siddiqui, S.; White, M.W.; Schroeder, A.M.; DeLuca, N.V.; Leszczynski, A.L.; Raimondi, S.L. Aberrant DNMT3B7 Expression Correlates to Tissue Type, Stage, and Survival across Cancers. PLoS ONE 2018, 13, e0201522. [Google Scholar] [CrossRef]
- Hu, H.; Chen, C.; Shi, S.; Li, B.; Duan, S. The Gene Mutations and Subtelomeric DNA Methylation in Immunodeficiency, Centromeric Instability and Facial Anomalies Syndrome. Autoimmunity 2019, 52, 192–198. [Google Scholar] [CrossRef]
- Wang, X.; Bhandari, R.K. DNA Methylation Dynamics during Epigenetic Reprogramming of Medaka Embryo. Epigenetics 2019, 14, 611–622. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, Y. Role of Mammalian DNA Methyltransferases in Development. Annu. Rev. Biochem. 2020, 89, 135–158. [Google Scholar] [CrossRef]
- Gatto, S.; Gagliardi, M.; Franzese, M.; Leppert, S.; Papa, M.; Cammisa, M.; Grillo, G.; Velasco, G.; Francastel, C.; Toubiana, S.; et al. ICF-Specific DNMT3B Dysfunction Interferes with Intragenic Regulation of MRNA Transcription and Alternative Splicing. Nucleic Acids Res. 2017, 45, 5739–5756. [Google Scholar] [CrossRef]
- Li, E.; Bestor, T.H.; Jaenisch, R. Targeted Mutation of the DNA Methyltransferase Gene Results in Embryonic Lethality. Cell 1992, 69, 915–926. [Google Scholar] [CrossRef]
- Saadeh, H.; Schulz, R. Protection of CpG Islands against de Novo DNA Methylation during Oogenesis Is Associated with the Recognition Site of E2f1 and E2f2. Epigenetics Chromatin 2014, 7, 26. [Google Scholar] [CrossRef]
- Li, E.; Zhang, Y. DNA Methylation in Mammals. Cold Spring Harb. Perspect. Biol. 2014, 6, a019133. [Google Scholar] [CrossRef]
- Feinberg, A.P.; Vogelstein, B. Hypomethylation Distinguishes Genes of Some Human Cancers from Their Normal Counterparts. Nature 1983, 301, 89–92. [Google Scholar] [CrossRef]
- Feinberg, A.P.; Vogelstein, B. A Technique for Radiolabeling DNA Restriction Endonuclease Fragments to High Specific Activity. Anal. Biochem. 1983, 132, 6–13. [Google Scholar] [CrossRef]
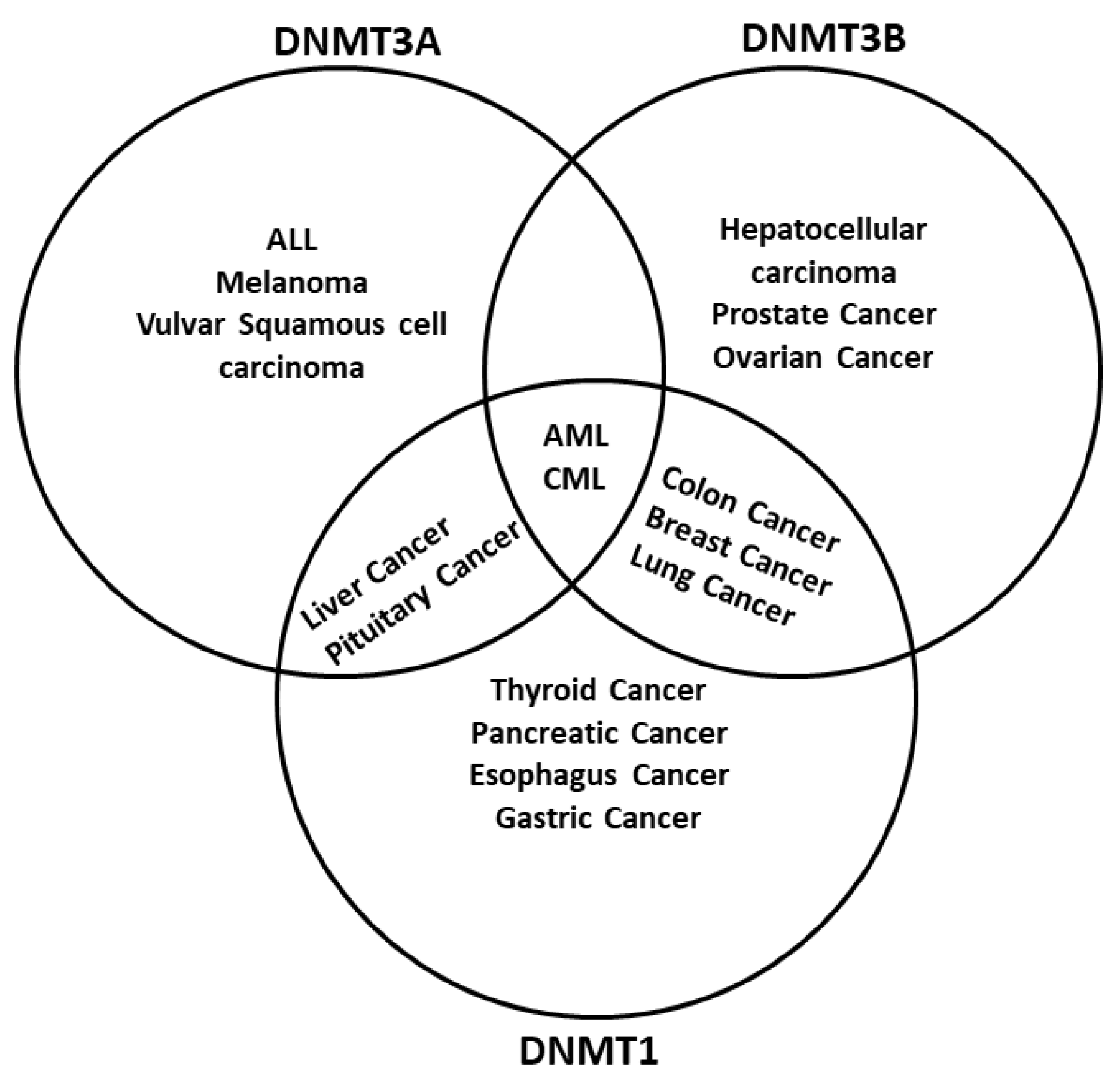
- Ley, T.J.; Ding, L.; Walter, M.J.; McLellan, M.D.; Lamprecht, T.; Larson, D.E.; Kandoth, C.; Payton, J.E.; Baty, J.; Welch, J.; et al. DNMT3A Mutations in Acute Myeloid Leukemia. N. Engl. J. Med. 2010, 363, 2424–2433. [Google Scholar] [CrossRef]
- Kanai, Y.; Ushijima, S.; Nakanishi, Y.; Sakamoto, M.; Hirohashi, S. Mutation of the DNA Methyltransferase (DNMT) 1 Gene in Human Colorectal Cancers. Cancer Lett. 2003, 192, 75–82. [Google Scholar] [CrossRef]
- Amatori, S.; Bagaloni, I.; Donati, B.; Fanelli, M. DNA Demethylating Antineoplastic Strategies: A Comparative Point of View. Genes Cancer 2010, 1, 197–209. [Google Scholar] [CrossRef]
- Huang, D.; Cui, L.; Ahmed, S.; Zainab, F.; Wu, Q.; Wang, X.; Yuan, Z. An Overview of Epigenetic Agents and Natural Nutrition Products Targeting DNA Methyltransferase, Histone Deacetylases and MicroRNAs. Food Chem. Toxicol. 2018, 123, 574–594. [Google Scholar] [CrossRef]
- Pan, Y.; Liu, G.; Zhou, F.; Su, B.; Li, Y. DNA Methylation Profiles in Cancer Diagnosis and Therapeutics. Clin. Exp. Med. 2018, 18, 1–14. [Google Scholar] [CrossRef]
- Gnyszka, A.; Jastrzebski, Z.; Flis, S. DNA Methyltransferase Inhibitors and Their Emerging Role in Epigenetic Therapy of Cancer. Anticancer Res. 2013, 33, 2989–2996. [Google Scholar]
- Datta, J.; Ghoshal, K.; Denny, W.A.; Gamage, S.A.; Brooke, D.G.; Phiasivongsa, P.; Redkar, S.; Jacob, S.T. A New Class of Quinoline-Based DNA Hypomethylating Agents Reactivates Tumor Suppressor Genes by Blocking DNA Methyltransferase 1 Activity and Inducing Its Degradation. Cancer Res. 2009, 69, 4277–4285. [Google Scholar] [CrossRef] [PubMed]
- Rilova, E.; Erdmann, A.; Gros, C.; Masson, V.; Aussagues, Y.; Poughon-Cassabois, V.; Rajavelu, A.; Jeltsch, A.; Menon, Y.; Novosad, N.; et al. Design, Synthesis and Biological Evaluation of 4-Amino-N-(4-Aminophenyl)Benzamide Analogues of Quinoline-Based SGI-1027 as Inhibitors of DNA Methylation. Chemmedchem 2014, 9, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Zwergel, C.; Schnekenburger, M.; Sarno, F.; Battistelli, C.; Manara, M.C.; Stazi, G.; Mazzone, R.; Fioravanti, R.; Gros, C.; Ausseil, F.; et al. Identification of a Novel Quinoline-Based DNA Demethylating Compound Highly Potent in Cancer Cells. Clin. Epigenetics 2019, 11, 68. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Liu, X.; Zeng, Y.; Liu, J.; Wu, F. DNA Methyltransferase Inhibitors Combination Therapy for the Treatment of Solid Tumor: Mechanism and Clinical Application. Clin. Epigenetics 2021, 13, 166. [Google Scholar] [CrossRef]
- Ahuja, N.; Sharma, A.R.; Baylin, S.B. Epigenetic Therapeutics: A New Weapon in the War Against Cancer. Annu. Rev. Med. 2016, 67, 73–89. [Google Scholar] [CrossRef]
- Bhat, S.A.; Ahmad, S.M.; Mumtaz, P.T.; Malik, A.A.; Dar, M.A.; Urwat, U.; Shah, R.A.; Ganai, N.A. Long Non-Coding RNAs: Mechanism of Action and Functional Utility. Non-Coding RNA Res. 2016, 1, 43–50. [Google Scholar] [CrossRef]
- Klisovic, R.B.; Stock, W.; Cataland, S.; Klisovic, M.I.; Liu, S.; Blum, W.; Green, M.; Odenike, O.; Godley, L.; Burgt, J.V.; et al. A Phase I Biological Study of MG98, an Oligodeoxynucleotide Antisense to DNA Methyltransferase 1, in Patients with High-Risk Myelodysplasia and Acute Myeloid Leukemia. Clin. Cancer Res. 2008, 14, 2444–2449. [Google Scholar] [CrossRef]
- Winquist, E.; Knox, J.; Ayoub, J.-P.; Wood, L.; Wainman, N.; Reid, G.K.; Pearce, L.; Shah, A.; Eisenhauer, E. Phase II Trial of DNA Methyltransferase 1 Inhibition with the Antisense Oligonucleotide MG98 in Patients with Metastatic Renal Carcinoma: A National Cancer Institute of Canada Clinical Trials Group Investigational New Drug Study. Investig. New Drugs 2006, 24, 159–167. [Google Scholar] [CrossRef]
- Plummer, R.; Vidal, L.; Griffin, M.; Lesley, M.; de Bono, J.; Coulthard, S.; Sludden, J.; Siu, L.L.; Chen, E.X.; Oza, A.M.; et al. Phase I Study of MG98, an Oligonucleotide Antisense Inhibitor of Human DNA Methyltransferase 1, Given as a 7-Day Infusion in Patients with Advanced Solid Tumors. Clin. Cancer Res. 2009, 15, 3177–3183. [Google Scholar] [CrossRef]
- Mazzoccoli, L.; Robaina, M.C.; Apa, A.G.; Bonamino, M.; Pinto, L.W.; Queiroga, E.; Bacchi, C.E.; Klumb, C.E. MiR-29 Silencing Modulates the Expression of Target Genes Related to Proliferation, Apoptosis and Methylation in Burkitt Lymphoma Cells. J. Cancer Res. Clin. Oncol. 2018, 144, 483–497. [Google Scholar] [CrossRef]
- Wang, L.; Huang, J.; Wu, C.; Huang, L.; Cui, J.; Xing, Z.; Zhao, C. Downregulation of MiR-29b Targets DNMT3b to Suppress Cellular Apoptosis and Enhace Proliferation in Pancreatic Cancer. Mol. Med. Rep. 2018, 17, 2113–2120. [Google Scholar] [CrossRef]
- Zhang, S.; Pei, M.; Li, Z.; Li, H.; Liu, Y.; Li, J. Double-negative Feedback Interaction between DNA Methyltransferase 3A and MicroRNA-145 in the Warburg Effect of Ovarian Cancer Cells. Cancer Sci. 2018, 109, 2734–2745. [Google Scholar] [CrossRef]
- Qi, D.; Li, J.; Que, B.; Su, J.; Li, M.; Zhang, C.; Yang, M.; Zhou, G.; Ji, W. Long Non-Coding RNA DBCCR1-003 Regulate the Expression of DBCCR1 via DNMT1 in Bladder Cancer. Cancer Cell Int. 2016, 16, 81. [Google Scholar] [CrossRef]
- Jones, R.; Wijesinghe, S.; Wilson, C.; Halsall, J.; Liloglou, T.; Kanhere, A. A Long Intergenic Non-Coding RNA Regulates Nuclear Localization of DNA Methyl Transferase-1. Iscience 2021, 24, 102273. [Google Scholar] [CrossRef]
- Somasundaram, S.; Forrest, M.E.; Moinova, H.; Cohen, A.; Varadan, V.; LaFramboise, T.; Markowitz, S.; Khalil, A.M. The DNMT1-Associated LincRNA DACOR1 Reprograms Genome-Wide DNA Methylation in Colon Cancer. Clin. Epigenetics 2018, 10, 127. [Google Scholar] [CrossRef]
- Guo, X.; Chen, Z.; Zhao, L.; Cheng, D.; Song, W.; Zhang, X. Long Non-Coding RNA-HAGLR Suppressed Tumor Growth of Lung Adenocarcinoma through Epigenetically Silencing E2F1. Exp. Cell Res. 2019, 382, 111461. [Google Scholar] [CrossRef]
- Li, W.; Zheng, J.; Deng, J.; You, Y.; Wu, H.; Li, N.; Lu, J.; Zhou, Y. Increased Levels of the Long Intergenic Non-Protein Coding RNA POU3F3 Promote DNA Methylation in Esophageal Squamous Cell Carcinoma Cells. Gastroenterology 2014, 146, 1714–1726. [Google Scholar] [CrossRef]
- Yan, J.; Guo, X.; Xia, J.; Shan, T.; Gu, C.; Liang, Z.; Zhao, W.; Jin, S. MiR-148a Regulates MEG3 in Gastric Cancer by Targeting DNA Methyltransferase 1. Med. Oncol. 2014, 31, 879. [Google Scholar] [CrossRef]
- Braconi, C.; Kogure, T.; Valeri, N.; Huang, N.; Nuovo, G.; Costinean, S.; Negrini, M.; Miotto, E.; Croce, C.M.; Patel, T. MicroRNA-29 Can Regulate Expression of the Long Non-Coding RNA Gene MEG3 in Hepatocellular Cancer. Oncogene 2011, 30, 4750–4756. [Google Scholar] [CrossRef]
- Zhang, P.; Sun, H.; Yang, B.; Luo, W.; Liu, Z.; Wang, J.; Zuo, Y. MiR-152 Regulated Glioma Cell Proliferation and Apoptosis via Runx2 Mediated by DNMT1. Biomed. Pharmacother. 2017, 92, 690–695. [Google Scholar] [CrossRef]
- Zhang, Z.; Tang, H.; Wang, Z.; Zhang, B.; Liu, W.; Lu, H.; Xiao, L.; Liu, X.; Wang, R.; Li, X.; et al. MiR-185 Targets the DNA Methyltransferases 1 and Regulates Global DNA Methylation in Human Glioma. Mol. Cancer 2011, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lei, Q.; Yu, Z.; Xu, G.; Tang, H.; Wang, W.; Wang, Z.; Li, G.; Wu, M. MiR-101 Reverses the Hypomethylation of the LMO3 Promoter in Glioma Cells. Oncotarget 2015, 6, 7930. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-H.; Hsu, W.-L.; Tseng, Y.-J.; Liu, D.-W.; Weng, C.-F. Involvement of DNMT 3B Promotes Epithelial-Mesenchymal Transition and Gene Expression Profile of Invasive Head and Neck Squamous Cell Carcinomas Cell Lines. BMC Cancer 2016, 16, 431. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
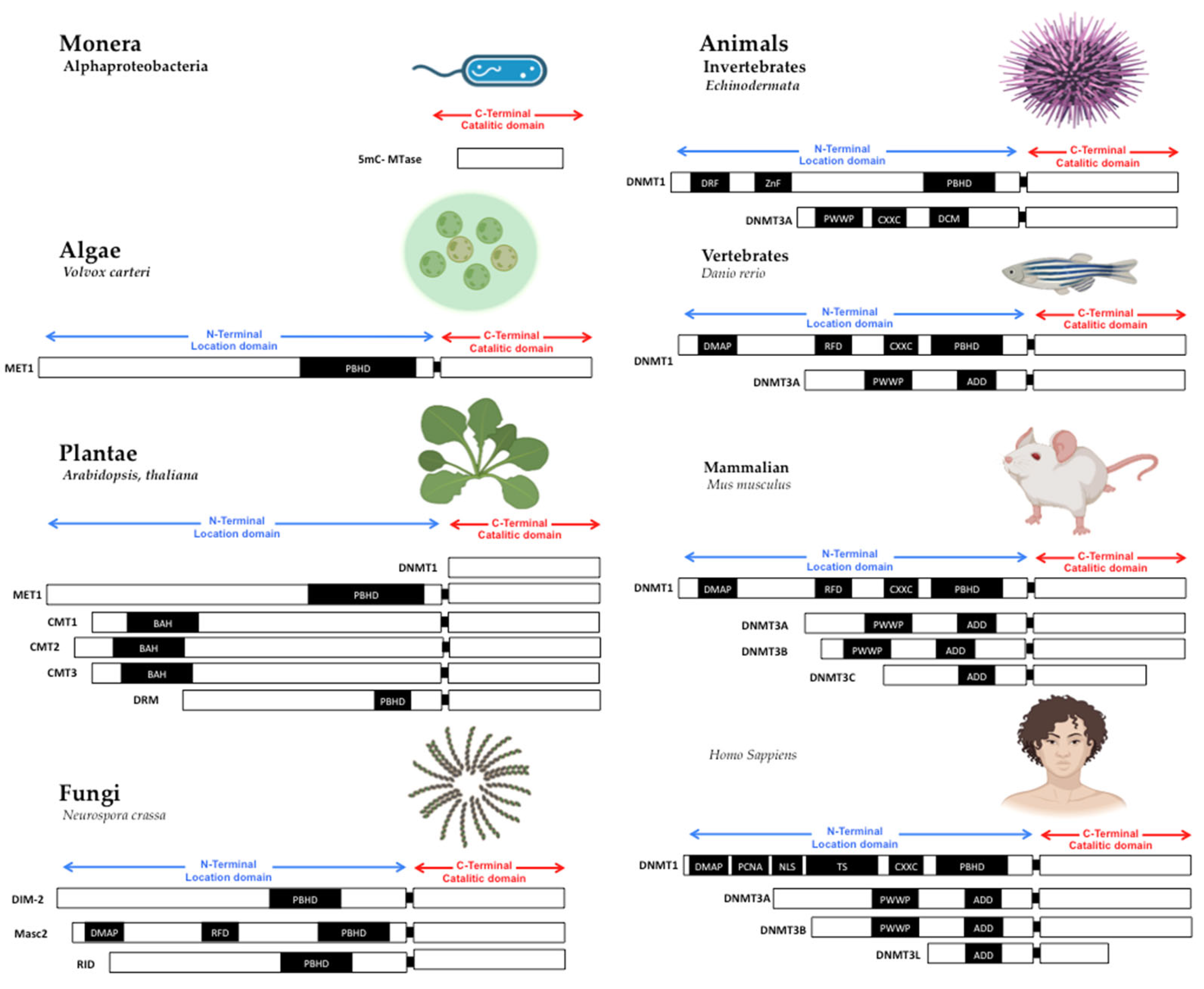
| Orthologs | DNA Methyltransferases Know in Each Taxonomic Group | Paralogs of DNA-Methyltransferases | Function Associated with Each Taxonomic Group | References |
|---|---|---|---|---|
| Monera | Alphaproteobacteria | 5mC-MTase. | Restriction modification (RM), bacteriophage’s sequence silence | [36] |
| Protists | Diatomeas and Choanoflagellates. | MET1, CMT3, and DRM2. | Not experimentally probed | [37] |
| Algae | Chlorella, spp., Clamydomonas; Volvox carteri. | MET1, CMT1 and 3, and DMR. | Genomic imprinting and development | [38,39] |
| Fungi | Humicola grisea; budding yeast; Neurospora sp.; Aspergillus clavatus. | DIM-2, Masc1 and 2, and RID. | Genomic repression. | [40,41,42] |
| Plantae | Arabidopsis thaliana | MET1, CMT, DMR DNMT1, 2, and 3. | Genomic imprinting, reproduction, and development. | [25,43] |
| Animals invertebrates | Nematoda, Equinoderma, Paracentrotus lividus, Solea seleganensis, Gadus morhua, Arthropoda | DNMT1 and 3 | Repetitive sequence repression, Development, Gene regulation, and epigenetic memory. | [44,45] |
| Animals vertebrates | Fishes and mammals | DNMT1, 2 and 3 | Repetitive sequence repression, Development, Gene regulation, epigenetic memory, and Cancer progression. | [3,46,47] |
| Orthologs within DNA Methylation Detected | DNMT Reported In Specie | References |
|---|---|---|
| Saccharomyces cerevisiae, | - | [34] |
| Caenorhabditis elegans | - | [34,35,45] |
| Drosophila melanogaster | DNMT2 | [31,32,33] |
| DNMT3B Isoform | Type of Biomarker | Type of Pathology | References |
|---|---|---|---|
| DNMT3B3 | Diagnostic | Cirrhosis Gastric cancer Hepatitis Liver cancer Ovarian carcinoma | [104,107,115] |
| DNMT3B4 | Diagnostic | Helicobacter pylori infection Renal cancer | [13,104,105,116] |
| DNMT3B7 | Diagnostic | Breast cancer | [105,108,109,110,117] |
| ΔDNMT3B4 | Diagnostic | Hyperplasia Lung cancer | [111] |
| ncRNAs | DNMT Deregulated in Cancer | Type of Cancer | Type of Deregulation | Reference |
|---|---|---|---|---|
| DBCCR1-003 | DNMT1 | Bladder | Down | [145] |
| linc-POU3F3 | DNMT1, 3A, and 3B | ESCC | Up | [149] |
| miR-148a | DNMT1 | Gastric | Down | [150] |
| miR-29a | DNMT1 | Liver | Down | [151] |
| miR-152 | DNMT1 | Glioma | Down | [152] |
| miR-185 | DNMT1 | Glioma | Down | [153] |
| miR-145 | DNMT3A | Ovarian | Down | [144] |
| miR-101 | DNMT3A | Glioma | Down | [154] |
| miR-29 | DNMT3B | Burkitt | Down | [142] |
| miR-29b | DNMT3B | Lymphoma, pancreatic, head and neck cell line cancer | Down | [143,155] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Castillo Falconi, V.M.; Torres-Arciga, K.; Matus-Ortega, G.; Díaz-Chávez, J.; Herrera, L.A. DNA Methyltransferases: From Evolution to Clinical Applications. Int. J. Mol. Sci. 2022, 23, 8994. https://doi.org/10.3390/ijms23168994
Del Castillo Falconi VM, Torres-Arciga K, Matus-Ortega G, Díaz-Chávez J, Herrera LA. DNA Methyltransferases: From Evolution to Clinical Applications. International Journal of Molecular Sciences. 2022; 23(16):8994. https://doi.org/10.3390/ijms23168994
Chicago/Turabian StyleDel Castillo Falconi, Victor M., Karla Torres-Arciga, Genaro Matus-Ortega, José Díaz-Chávez, and Luis A. Herrera. 2022. "DNA Methyltransferases: From Evolution to Clinical Applications" International Journal of Molecular Sciences 23, no. 16: 8994. https://doi.org/10.3390/ijms23168994
APA StyleDel Castillo Falconi, V. M., Torres-Arciga, K., Matus-Ortega, G., Díaz-Chávez, J., & Herrera, L. A. (2022). DNA Methyltransferases: From Evolution to Clinical Applications. International Journal of Molecular Sciences, 23(16), 8994. https://doi.org/10.3390/ijms23168994