
Kinetic Characterization of Cerium and Gallium Ions as Inhibitors of Cysteine Cathepsins L, K, and S


Abstract
:1. Introduction
2. Results
2.1. Spectroscopic Analysis of Interaction between Ions and Substrate
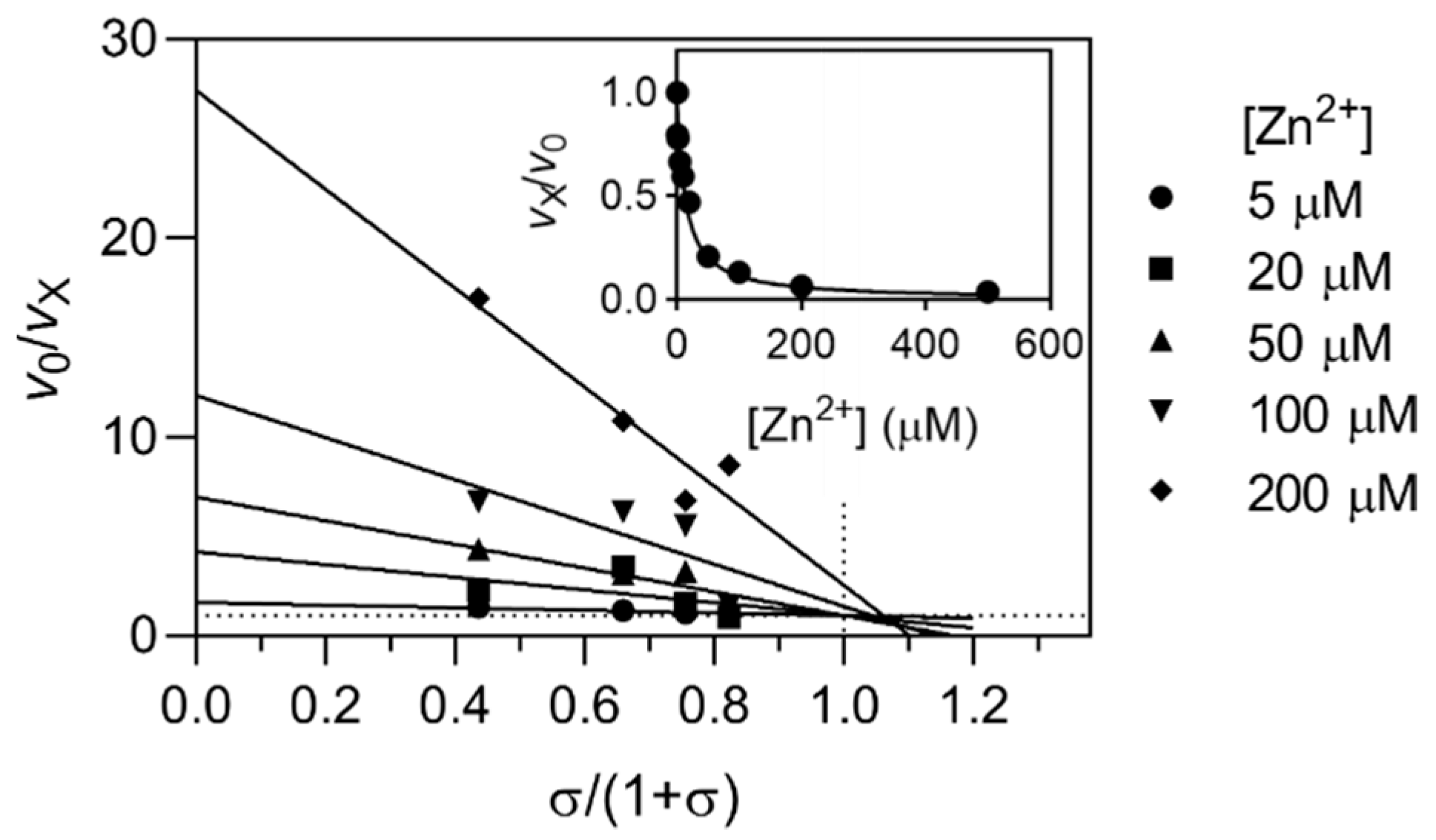
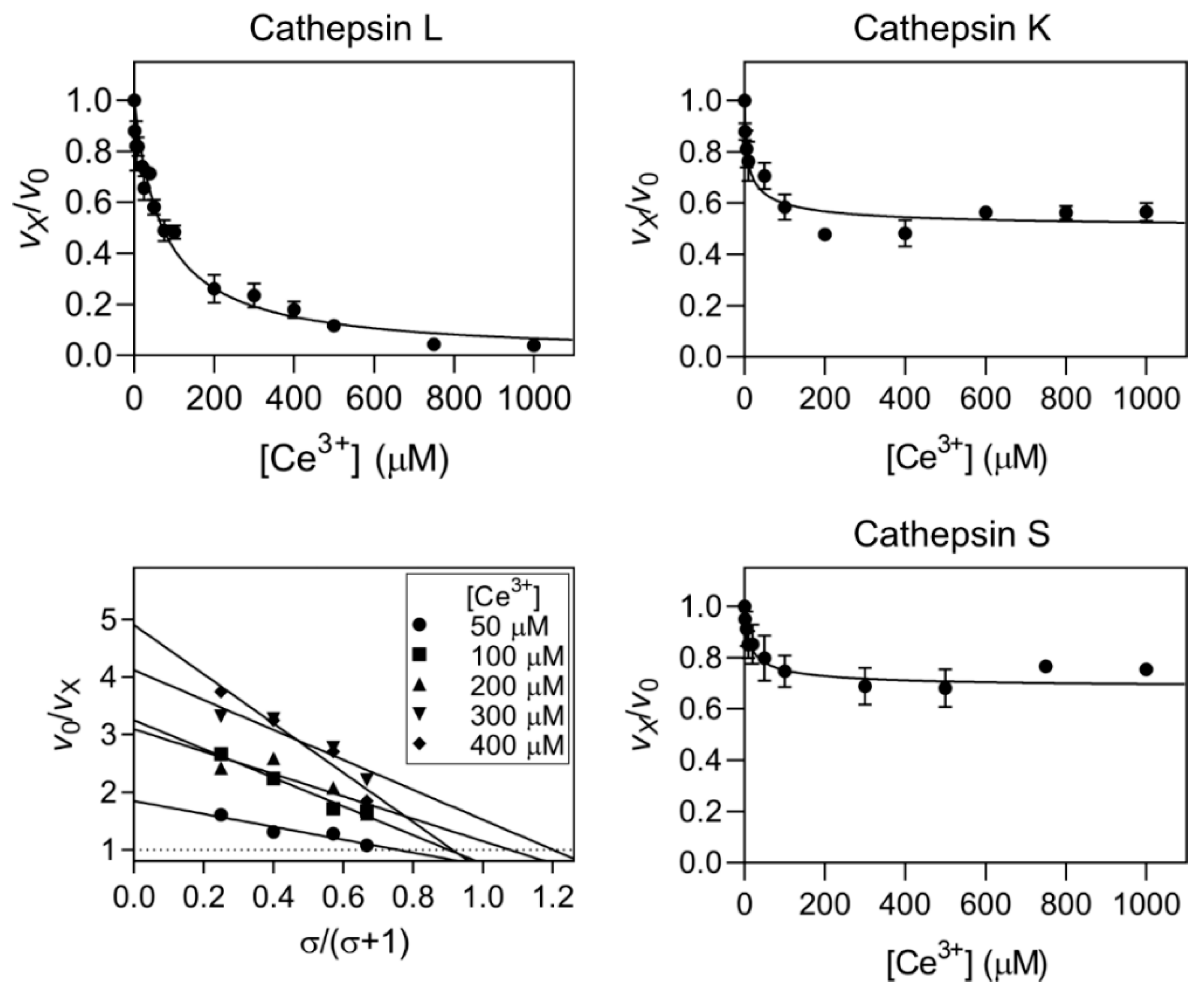
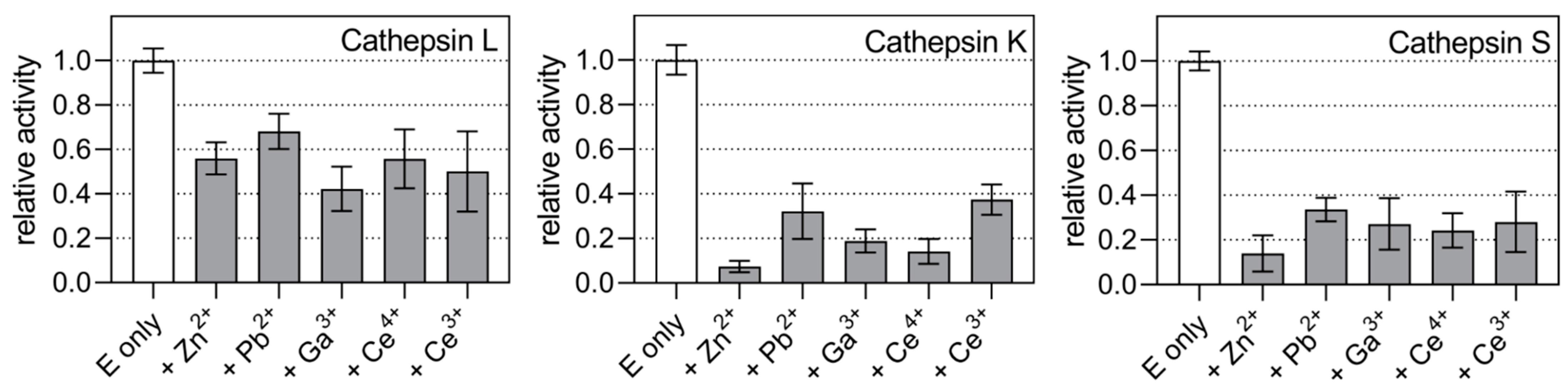
2.2. Kinetic Mechanisms of Action
2.3. Hydrolysis of Bovine Neck Ligament Elastin
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. UV-Vis Spectroscopy
4.3. Enzyme Kinetics
4.4. Elastin Degradation Assays
4.5. Kinetic Models
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Novinec, M.; Lenarčič, B. Papain-like Peptidases: Structure, Function, and Evolution. Biomol. Concepts 2013, 4, 287–308. [Google Scholar] [CrossRef]
- Turk, V.; Stoka, V.; Vasiljeva, O.; Renko, M.; Sun, T.; Turk, B.; Turk, D. Cysteine Cathepsins: From Structure, Function and Regulation to New Frontiers. Biochim. Biophys. Acta—Proteins Proteom. 2012, 1824, 68–88. [Google Scholar] [CrossRef] [PubMed]
- Karrer, K.M.; Peiffer, S.L.; Ditomas, M.E. Two Distinct Gene Subfamilies within the Family of Cysteine Protease Genes. Proc. Natl. Acad. Sci. USA 1993, 90, 3063–3067. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of Spike Glycoprotein of SARS-CoV-2 on Virus Entry and Its Immune Cross-Reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Zeng, C.; Huang, F.; Yuan, F.; Yan, J.; Zhao, Y.; Zhou, Y.; Hankey, W.; Jin, V.X.; Huang, J.; et al. Cas13d Knockdown of Lung Protease Ctsl Prevents and Treats SARS-CoV-2 Infection. Nat. Chem. Biol. 2022. [Google Scholar] [CrossRef]
- Zhu, J.; Li, L.; Drelich, A.; Chenna, B.C.; Mellott, D.M.; Taylor, Z.W.; Tat, V.; Garcia, C.Z.; Katzfuss, A.; Tseng, C.-T.K.; et al. Self-Masked Aldehyde Inhibitors of Human Cathepsin L Are Potent Anti-CoV-2 Agents. Front. Chem. 2022, 10, 867928. [Google Scholar] [CrossRef] [PubMed]
- Gelb, B.D.; Shi, G.-P.; Chapman, H.A.; Desnick, R.J. Pycnodysostosis, a Lysosomal Disease Caused by Cathepsin K Deficiency. Science 1996, 273, 1236–1238. [Google Scholar] [CrossRef]
- Riese, R.J.; Wolf, P.R.; Brömme, D.; Natkin, L.R.; Villadangos, J.A.; Ploegh, H.L.; Chapman, H.A. Essential Role for Cathepsin S in MHC Class II-Associated Invariant Chain Processing and Peptide Loading. Immunity 1996, 4, 357–366. [Google Scholar] [CrossRef]
- Dai, R.; Wu, Z.; Chu, H.Y.; Lu, J.; Lyu, A.; Liu, J.; Zhang, G. Cathepsin K: The Action in and Beyond Bone. Front. Cell Dev. Biol. 2020, 8, 433. [Google Scholar] [CrossRef]
- McClung, M.R.; O’Donoghue, M.L.; Papapoulos, S.E.; Bone, H.; Langdahl, B.; Saag, K.G.; Reid, I.R.; Kiel, D.P.; Cavallari, I.; Bonaca, M.P.; et al. Odanacatib for the Treatment of Postmenopausal Osteoporosis: Results of the LOFT Multicentre, Randomised, Double-Blind, Placebo-Controlled Trial and LOFT Extension Study. Lancet Diabetes Endocrinol. 2019, 7, 899–911. [Google Scholar] [CrossRef]
- Li, H.; Xiao, Z.; Quarles, L.D.; Li, W. Osteoporosis: Mechanism, Molecular Target and Current Status on Drug Development. Curr. Med. Chem. 2020, 28, 1489–1507. [Google Scholar] [CrossRef]
- Conaghan, P.G.; Bowes, M.A.; Kingsbury, S.R.; Brett, A.; Guillard, G.; Rizoska, B.; Sjögren, N.; Graham, P.; Jansson, Å.; Wadell, C.; et al. Disease-Modifying Effects of a Novel Cathepsin k Inhibitor in Osteoarthritis: A Randomized Controlled Trial. Ann. Intern. Med. 2020, 172, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.; Nath, S.; Lora, A.; Samaha, G.; Elgamal, Z.; Kaiser, R.; Taggart, C.; Weldon, S.; Geraghty, P. Cathepsin S: Investigating an Old Player in Lung Disease Pathogenesis, Comorbidities, and Potential Therapeutics. Respir. Res. 2020, 21, 111. [Google Scholar] [CrossRef] [PubMed]
- Yoo, Y.J.; Choi, E.; Kim, Y.; Cha, Y.; Um, E.; Kim, Y.; Kim, Y.; Lee, Y.S. Therapeutic Potential of Targeting Cathepsin S in Pulmonary Fibrosis. Biomed. Pharmacother. 2022, 145, 112245. [Google Scholar] [CrossRef]
- Moustakas, M. The Role of Metal Ions in Biology, Biochemistry and Medicine. Materials 2021, 14, 549. [Google Scholar] [CrossRef]
- Crichton, R.R. CHAPTER 1: Metal Toxicity—An Introduction. In Metal Chelation in Medicine; Royal Society of Chemistry: London, UK, 2016; pp. 1–23. [Google Scholar] [CrossRef]
- Tsvetkov, P.; Coy, S.; Petrova, B.; Dreishpoon, M.; Verma, A.; Abdusamad, M.; Rossen, J.; Joesch-Cohen, L.; Humeidi, R.; Spangler, R.D.; et al. Copper Induces Cell Death by Targeting Lipoylated TCA Cycle Proteins. Science 2022, 375, 1254–1261. [Google Scholar] [CrossRef]
- Lockwood, T.D. Cys-His Proteases Are among the Wired Proteins of the Cell. Arch. Biochem. Biophys. 2004, 432, 12–24. [Google Scholar] [CrossRef]
- Lockwood, T.D. Lysosomal Metal, Redox and Proton Cycles Influencing the CysHis Cathepsin Reaction. Metallomics 2013, 5, 110. [Google Scholar] [CrossRef]
- Panchariya, L.; Khan, W.A.; Kuila, S.; Sonkar, K.; Sahoo, S.; Ghoshal, A.; Kumar, A.; Verma, D.K.; Hasan, A.; Khan, M.A.; et al. Zinc2+ Ion Inhibits SARS-CoV-2 Main Protease and Viral Replication In Vitro. Chem. Commun. 2021, 57, 10083–10086. [Google Scholar] [CrossRef]
- Kladnik, J.; Dolinar, A.; Kljun, J.; Perea, D.; Grau-Exposito, J.; Genesca, M.; Novinec, M.; Buzon, M.J.; Turel, I. Zinc Pyrithione Is a Potent Inhibitor of PLPro and Cathepsin L Enzymes with Ex Vivo Inhibition of SARS-CoV-2 Entry and Replication. J. Enzyme Inhib. Med. Chem. 2022, 37, 2158–2168. [Google Scholar] [CrossRef]
- Meisenheimer, M.; Saenko, Y.; Eppard, E. Gallium-68: Radiolabeling of Radiopharmaceuticals for PET Imaging—A Lot to Consider. In Medical Isotopes; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Chandler, J.E.; Messer, H.H.; Ellender, G. Cytotoxicity of Gallium and Indium Ions Compared with Mercuric Ion. J. Dent. Res. 1994, 73, 1554–1559. [Google Scholar] [CrossRef] [PubMed]
- Rajeshkumar, S.; Naik, P. Synthesis and Biomedical Applications of Cerium Oxide Nanoparticles—A Review. Biotechnol. Rep. 2018, 17, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Nelson, B.C.; Johnson, M.E.; Walker, M.L.; Riley, K.R.; Sims, C.M. Antioxidant Cerium Oxide Nanoparticles in Biology and Medicine. Antioxidants 2016, 5, 15. [Google Scholar] [CrossRef]
- Benedetto, A.; Bocca, C.; Brizio, P.; Cannito, S.; Abete, M.C.; Squadrone, S. Effects of the Rare Elements Lanthanum and Cerium on the Growth of Colorectal and Hepatic Cancer Cell Lines. Toxicol. Vitr. 2018, 46, 9–18. [Google Scholar] [CrossRef]
- Ngoc, L.T.N.; Bui, V.K.H.; Moon, J.-Y.; Lee, Y.-C. In-Vitro Cytotoxicity and Oxidative Stress Induced by Cerium Aminoclay and Cerium Oxide Nanoparticles in Human Skin Keratinocyte Cells. J. Nanosci. Nanotechnol. 2019, 19, 6369–6375. [Google Scholar] [CrossRef]
- De Marzi, L.; Monaco, A.; De Lapuente, J.; Ramos, D.; Borras, M.; Di Gioacchino, M.; Santucci, S.; Poma, A. Cytotoxicity and Genotoxicity of Ceria Nanoparticles on Different Cell Lines in Vitro. Int. J. Mol. Sci. 2013, 14, 3065–3077. [Google Scholar] [CrossRef]
- Benameur, L.; Auffan, M.; Cassien, M.; Liu, W.; Culcasi, M.; Rahmouni, H.; Stocker, P.; Tassistro, V.; Bottero, J.Y.; Rose, J.; et al. DNA Damage and Oxidative Stress Induced by CeO2 Nanoparticles in Human Dermal Fibroblasts: Evidence of a Clastogenic Effect as a Mechanism of Genotoxicity. Nanotoxicology 2015, 9, 696–705. [Google Scholar] [CrossRef]
- Mayr, H.; Breugst, M.; Ofial, A.R. Farewell to the HSAB Treatment of Ambident Reactivity. Angew. Chemie Int. Ed. 2011, 50, 6470–6505. [Google Scholar] [CrossRef]
- LoPachin, R.M.; Gavin, T.; DeCaprio, A.; Barber, D.S. Application of the Hard and Soft, Acids and Bases (HSAB) Theory to Toxicant—Target Interactions. Chem. Res. Toxicol. 2012, 25, 239–251. [Google Scholar] [CrossRef]
- Cárdenas, C.; Ayers, P.W. How Reliable Is the Hard–Soft Acid–Base Principle? An Assessment from Numerical Simulations of Electron Transfer Energies. Phys. Chem. Chem. Phys. 2013, 15, 13959–13968. [Google Scholar] [CrossRef] [PubMed]
- Ayers, P.W.; Cárdenas, C. Communication: A Case Where the Hard/Soft Acid/Base Principle Holds Regardless of Acid/Base Strength. J. Chem. Phys. 2013, 138, 181106. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.G. Hard and Soft Acids and Bases—The Evolution of a Chemical Concept. Coord. Chem. Rev. 1990, 100, 403–425. [Google Scholar] [CrossRef]
- Sekhon, B.S.; Chopra, S.L. A Thermodynamic Study of the Complexation Reaction for Some Amino Acids with Cerium(III) and Yttrium(III). Thermochim. Acta 1973, 7, 151–157. [Google Scholar] [CrossRef]
- Kiani, F.; Rostami, A.A.; Gharib, F.; Sharifi, S.; Bahadory, A. Complex Formation of the Cerium(IV) Ion with Glycyl-Glycine at Different Ionic Strengths. J. Chem. Eng. Data 2011, 56, 2830–2835. [Google Scholar] [CrossRef]
- Palmier, M.O.; Van Doren, S.R. Rapid Determination of Enzyme Kinetics from Fluorescence: Overcoming the Inner Filter Effect. Anal. Biochem. 2007, 371, 43–51. [Google Scholar] [CrossRef]
- Ghose, M.; Banerjee, S.; Patra, S.; Mukherjea, K.K. Synthesis, Structure and Artificial Protease Activities of Two Cerium (III) Complexes. J. Lumin. 2016, 180, 224–233. [Google Scholar] [CrossRef]
- Williams, D.E.; Basnet, K.; Grant, K.B. Tuning Cerium(IV)-Assisted Hydrolysis of Phosphatidylcholine Liposomes under Mildly Acidic and Neutral Conditions. ChemBioChem 2015, 16, 1474–1482. [Google Scholar] [CrossRef]
- Baici, A. The Specific Velocity Plot: A Graphical Method for Determining Inhibition Parameters for Both Linear and Hyperbolic Enzyme Inhibitors. Eur. J. Biochem. 1981, 119, 9–14. [Google Scholar] [CrossRef]
- Botts, J.; Morales, M. Analytical Description of the Effects of Modifiers and of Enzyme Multivalency upon the Steady State Catalyzed Reaction Rate. Trans. Faraday Soc. 1953, 49, 696–707. [Google Scholar] [CrossRef]
- Vizovišek, M.; Vidmar, R.; Van Quickelberghe, E.; Impens, F.; Andjelković, U.; Sobotič, B.; Stoka, V.; Gevaert, K.; Turk, B.; Fonović, M. Fast Profiling of Protease Specificity Reveals Similar Substrate Specificities for Cathepsins K, L and S. Proteomics 2015, 15, 2479–2490. [Google Scholar] [CrossRef]
- Novinec, M.; Korenč, M.; Caflisch, A.; Ranganathan, R.; Lenarčič, B.; Baici, A. A Novel Allosteric Mechanism in the Cysteine Peptidase Cathepsin K Discovered by Computational Methods. Nat. Commun. 2014, 5, 3287. [Google Scholar] [CrossRef] [PubMed]
- Novinec, M.; Lenarčič, B.; Baici, A. Probing the Activity Modification Space of the Cysteine Peptidase Cathepsin K with Novel Allosteric Modifiers. PLoS ONE 2014, 9, e106642. [Google Scholar] [CrossRef] [PubMed]
- Goričan, T.; Ciber, L.; Petek, N.; Svete, J.; Novinec, M. Synthesis and Kinetic Characterization of Hyperbolic Inhibitors of Human Cathepsins K and S Based on a Succinimide Scaffold. Bioorg. Chem. 2021, 115, 105213. [Google Scholar] [CrossRef] [PubMed]
- Schenker, P.; Baici, A. Paradoxical Interactions between Modifiers and Elastase-2. FEBS J. 2010, 277, 2486–2495. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.W.; Johnson, D.A.; Barrett, A.J.; Chapman, H.A. Elastinolytic activity of human cathepsin L. Biochem. J. 1986, 233, 925–927. [Google Scholar] [CrossRef] [PubMed]
- Andrault, P.M.; Panwar, P.; Mackenzie, N.C.W.; Brömme, D. Elastolytic Activity of Cysteine Cathepsins K, S, and V Promotes Vascular Calcification. Sci. Rep. 2019, 9, 9682. [Google Scholar] [CrossRef]
- Punturieri, A.; Filippov, S.; Allen, E.; Caras, I.; Murray, R.; Reddy, V.; Weiss, S.J. Regulation of Elastinolytic Cysteine Proteinase Activity in Normal and Cathepsin K-Deficient Human Macrophages. J. Exp. Med. 2000, 192, 789–799. [Google Scholar] [CrossRef]
- Reddy, V.Y.; Zhang, Q.Y.; Weiss, S.J. Pericellular Mobilization of the Tissue-Destructive Cysteine Proteinases, Cathepsins B, L, and S, by Human Monocyte-Derived Macrophages. Proc. Natl. Acad. Sci. USA 1995, 92, 3849–3853. [Google Scholar] [CrossRef]
- Novinec, M.; Grass, R.N.; Stark, W.J.; Turk, V.; Baici, A.; Lenarčič, B. Interaction between Human Cathepsins K, L, and S and Elastins: Mechanism of Elastinolysis and Inhibition by Macromolecular Inhibitors. J. Biol. Chem. 2007, 282, 7893–7902. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Panwar, P.; O’Donoghue, A.J.; Cui, H.; Guido, R.V.C.; Craik, C.S.; Brömme, D. Structural Requirements for the Collagenase and Elastase Activity of Cathepsin K and Its Selective Inhibition by an Exosite Inhibitor. Biochem. J. 2015, 465, 163–173. [Google Scholar] [CrossRef]
- Andrault, P.M.; Panwar, P.; Brömme, D. Characterization of Cathepsin S Exosites That Govern Its Elastolytic Activity. Biochem. J. 2020, 477, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, J.Y.; Chauret, N.; Cromlish, W.; Desmarais, S.; Duong, L.T.; Falgueyret, J.P.; Kimmel, D.B.; Lamontagne, S.; Léger, S.; LeRiche, T.; et al. The Discovery of Odanacatib (MK-0822), a Selective Inhibitor of Cathepsin K. Bioorg. Med. Chem. Lett. 2008, 18, 923–928. [Google Scholar] [CrossRef]
- Tato, M.; Kumar, S.V.; Liu, Y.; Mulay, S.R.; Moll, S.; Popper, B.; Eberhard, J.N.; Thomasova, D.; Rufer, A.C.; Gruner, S.; et al. Cathepsin S Inhibition Combines Control of Systemic and Peripheral Pathomechanisms of Autoimmune Tissue Injury. Sci. Rep. 2017, 7, 2775. [Google Scholar] [CrossRef] [PubMed]
- Alessio, E. Thirty Years of the Drug Candidate NAMI-A and the Myths in the Field of Ruthenium Anticancer Compounds: A Personal Perspective. Eur. J. Inorg. Chem. 2017, 2017, 1549–1560. [Google Scholar] [CrossRef]
- Li, Z.; Kienetz, M.; Cherney, M.M.; James, M.N.G.; Brömme, D. The Crystal and Molecular Structures of a Cathepsin K:Chondroitin Sulfate Complex. J. Mol. Biol. 2008, 383, 78–91. [Google Scholar] [CrossRef]
- Novinec, M. Computational Investigation of Conformational Variability and Allostery in Cathepsin K and Other Related Peptidases. PLoS ONE 2017, 12, e0182387. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.; Bush, N.; Burns, Z.; Doherty, G.; Foley, T.; Milone, M.; Maki, K.L.; Cromer, M. Modeling the Kinetic Behavior of Reactive Oxygen Species with Cerium Dioxide Nanoparticles. Biomolecules 2019, 9, 447. [Google Scholar] [CrossRef] [PubMed]
- Bilal, B.A.; Müller, E. Thermodynamic Study of Ce4+/Ce3+ Redox Reaction in Aqueous Solutions at Elevated Temperatures: 1. Reduction Potential and Hydrolysis Equilibria of Ce4+ in HClO4 Solutions. Z. Naturforsch.—Sect. A J. Phys. Sci. 1992, 47, 974–984. [Google Scholar] [CrossRef]
- Brown, P.L.; Ekberg, C. Theory. In Hydrolysis of Metal Ions; Wiley Online Books: Hoboken, NJ, USA, 2016; pp. 7–30. ISBN 9783527656189. [Google Scholar]
- Höllriegl, V.; González-Estecha, M.; Trasobares, E.M.; Giussani, A.; Oeh, U.; Herraiz, M.A.; Michalke, B. Measurement of Cerium in Human Breast Milk and Blood Samples. J. Trace Elem. Med. Biol. 2010, 24, 193–199. [Google Scholar] [CrossRef]
- Belozerov, E.S. The Topography of Gallium in the Healthy Human Body. Bull. Exp. Biol. Med. 1966, 62, 1141–1142. [Google Scholar] [CrossRef]
- Korenč, M.; Lenarčič, B.; Novinec, M. Human Cathepsin L, a Papain-like Collagenase without Proline Specificity. FEBS J. 2015, 282, 4328–4340. [Google Scholar] [CrossRef] [PubMed]



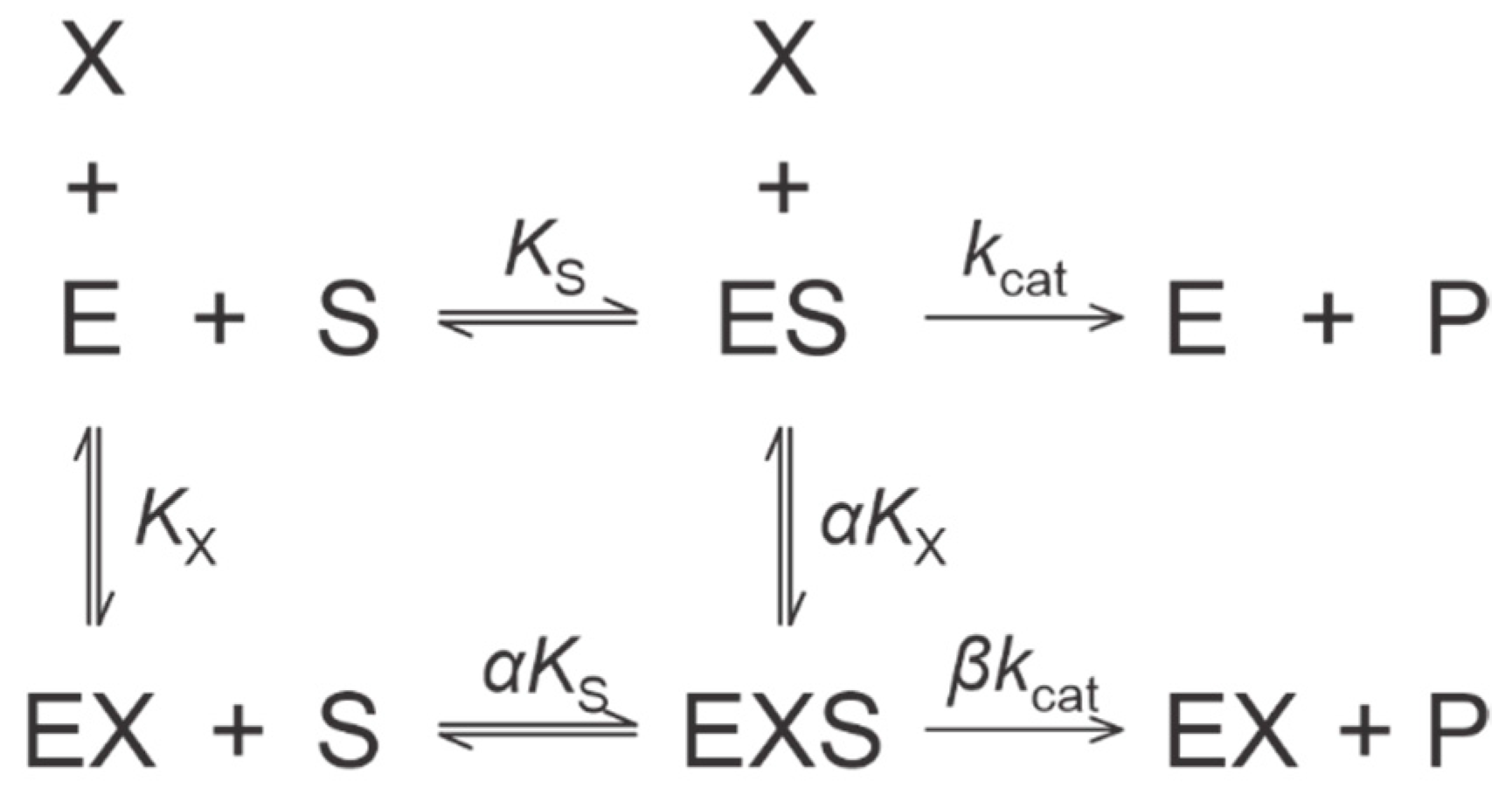

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ion | Enzyme | Mechanism | KX (µM) | α | β |
|---|---|---|---|---|---|
| Zn2+ | Cathepsin L | Linear specific | 7.5 ± 0.9 | ||
| Cathepsin K | Linear specific | 7.3 ± 0.3 | |||
| Cathepsin S | Linear specific | 2.9 ± 0.5 | |||
| Pb2+ | Cathepsin L | Linear specific | 26 ± 12 | ||
| Cathepsin K | Linear specific | 135 ± 13 | |||
| Cathepsin S | Linear specific | 50 ± 5 | |||
| Ga3+ | Cathepsin L | Linear specific | 0.60 ± 0.04 | ||
| Cathepsin K | Hyperbolic mixed | 4.1 ± 0.8 | 11.5 ± 1.0 | 2.4 ± 1.4 | |
| Cathepsin S | Hyperbolic mixed | 3.7 ± 0.6 | 11.3 ± 1.0 | 3.3 ± 0.5 | |
| Ce4+ | Cathepsin L | Linear specific | 0.42 ± 0.03 | ||
| Cathepsin K | Hyperbolic mixed | 1.3 ± 0.2 | 3.3 ± 0.6 | 0.73 ± 0.04 | |
| Cathepsin S | Hyperbolic catalytic | 1.2 ± 0.2 | 0.18 ± 0.01 | 0.18 ± 0.01 | |
| Ce3+ | Cathepsin L | Linear specific | 12 ± 6 | ||
| Ion | Enzyme | Mechanism | EC50 (µM) | h | |
| Ce3+ | Cathepsin K | Hyperbolic | 11 ± 6 | 0.6 ± 0.2 | |
| Cathepsin S | Hyperbolic | 17 ± 5 | 0.7 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novinec, M.; Bembič, P.; Janković, M.; Kisilak, M.; Kljun, J.; Turel, I. Kinetic Characterization of Cerium and Gallium Ions as Inhibitors of Cysteine Cathepsins L, K, and S. Int. J. Mol. Sci. 2022, 23, 8993. https://doi.org/10.3390/ijms23168993
Novinec M, Bembič P, Janković M, Kisilak M, Kljun J, Turel I. Kinetic Characterization of Cerium and Gallium Ions as Inhibitors of Cysteine Cathepsins L, K, and S. International Journal of Molecular Sciences. 2022; 23(16):8993. https://doi.org/10.3390/ijms23168993
Chicago/Turabian StyleNovinec, Marko, Primož Bembič, Milica Janković, Marija Kisilak, Jakob Kljun, and Iztok Turel. 2022. "Kinetic Characterization of Cerium and Gallium Ions as Inhibitors of Cysteine Cathepsins L, K, and S" International Journal of Molecular Sciences 23, no. 16: 8993. https://doi.org/10.3390/ijms23168993
APA StyleNovinec, M., Bembič, P., Janković, M., Kisilak, M., Kljun, J., & Turel, I. (2022). Kinetic Characterization of Cerium and Gallium Ions as Inhibitors of Cysteine Cathepsins L, K, and S. International Journal of Molecular Sciences, 23(16), 8993. https://doi.org/10.3390/ijms23168993