
A Pilot Study on Peptidylarginine Deiminases and Protein Deimination in Animal Cancers across Vertebrate Species




Abstract
:1. Introduction
2. Results
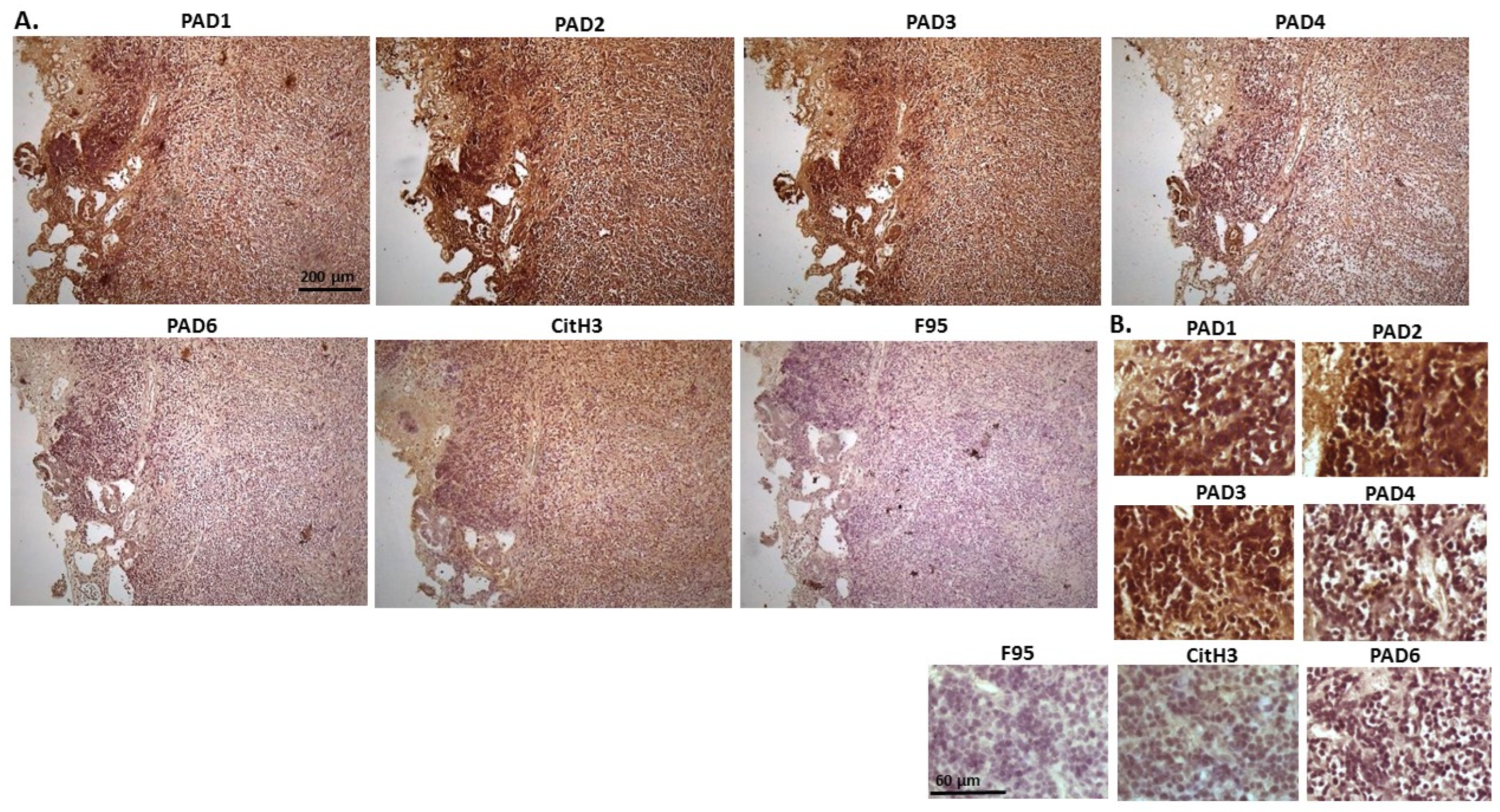
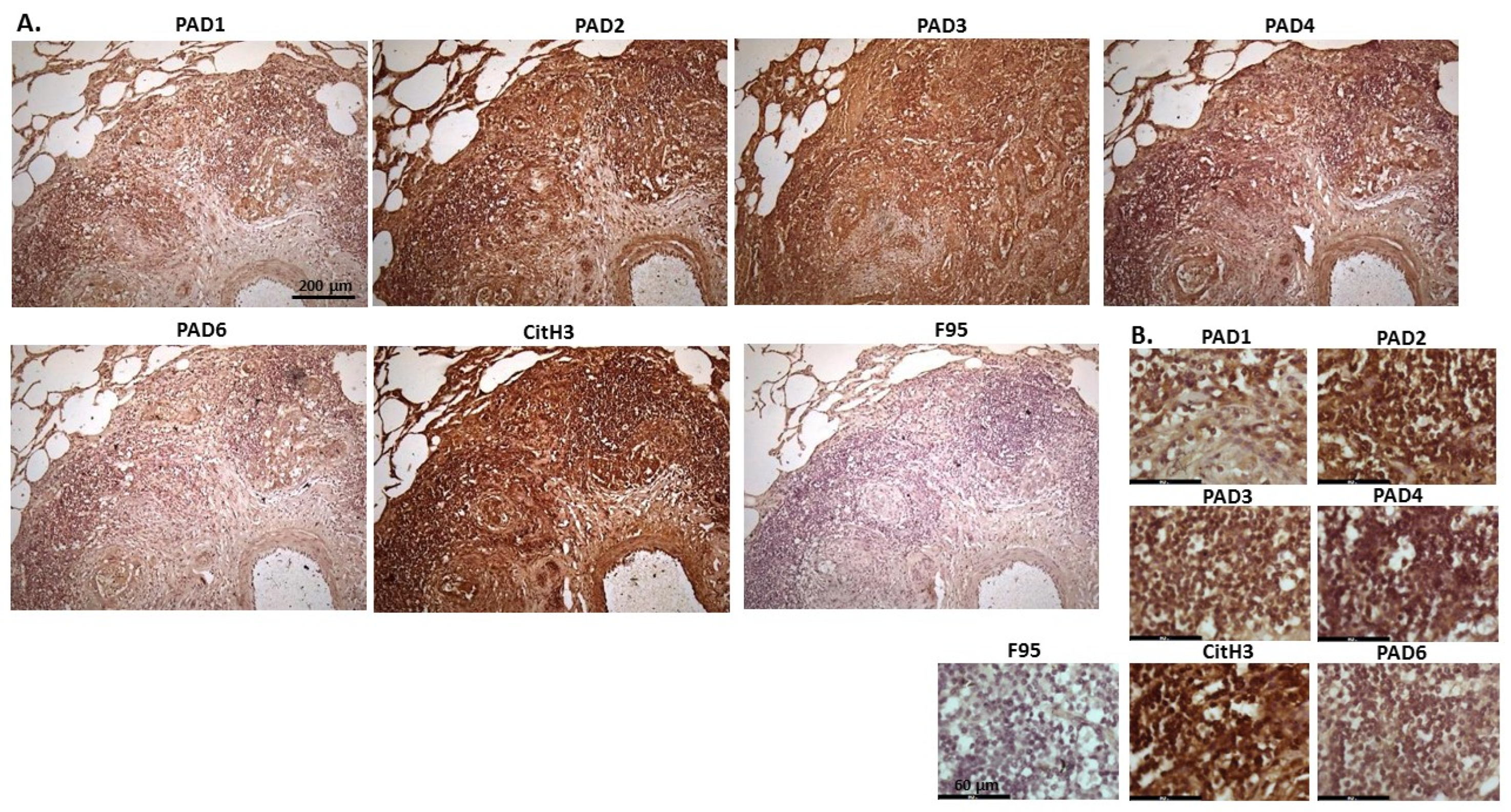
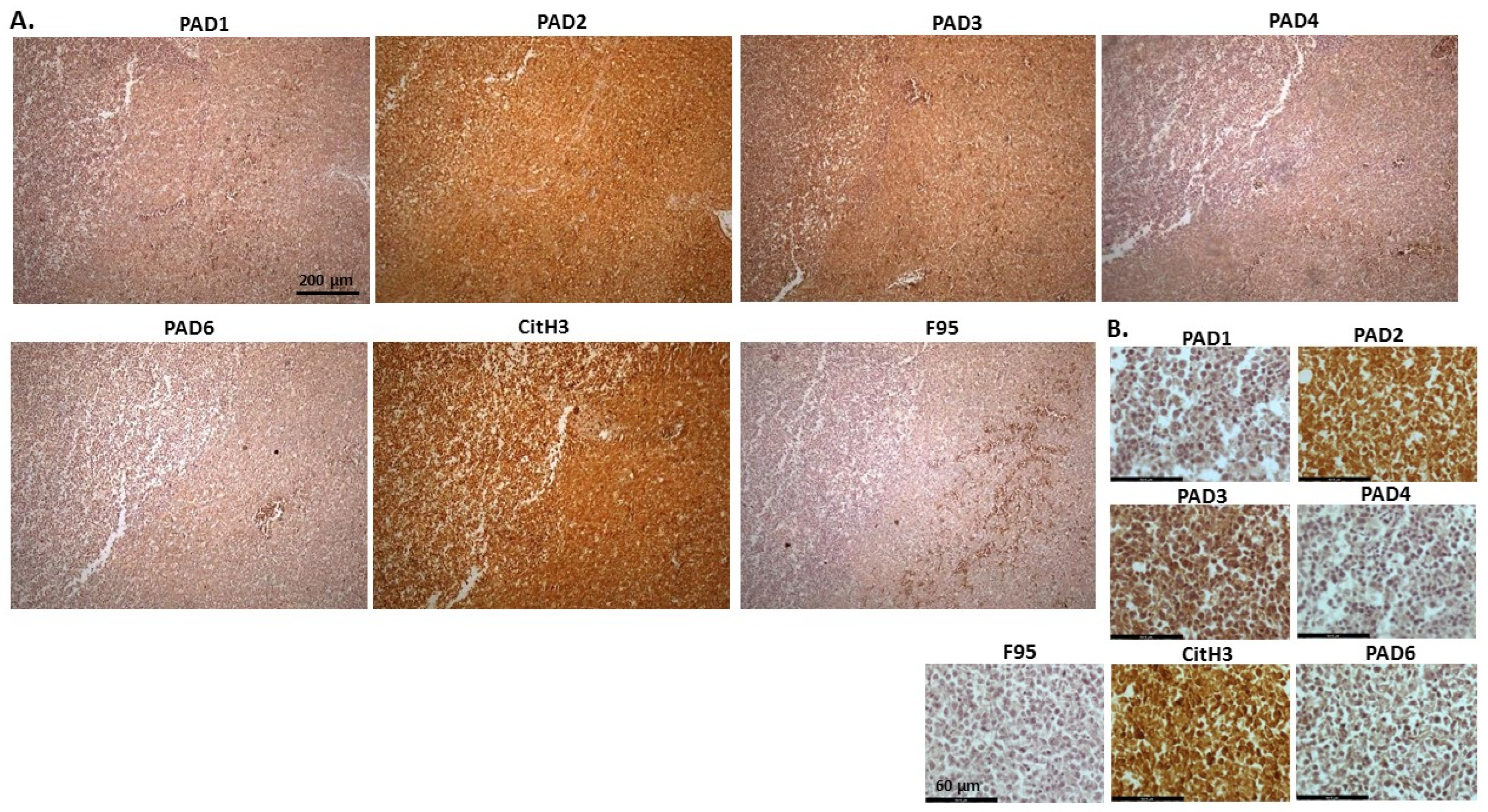
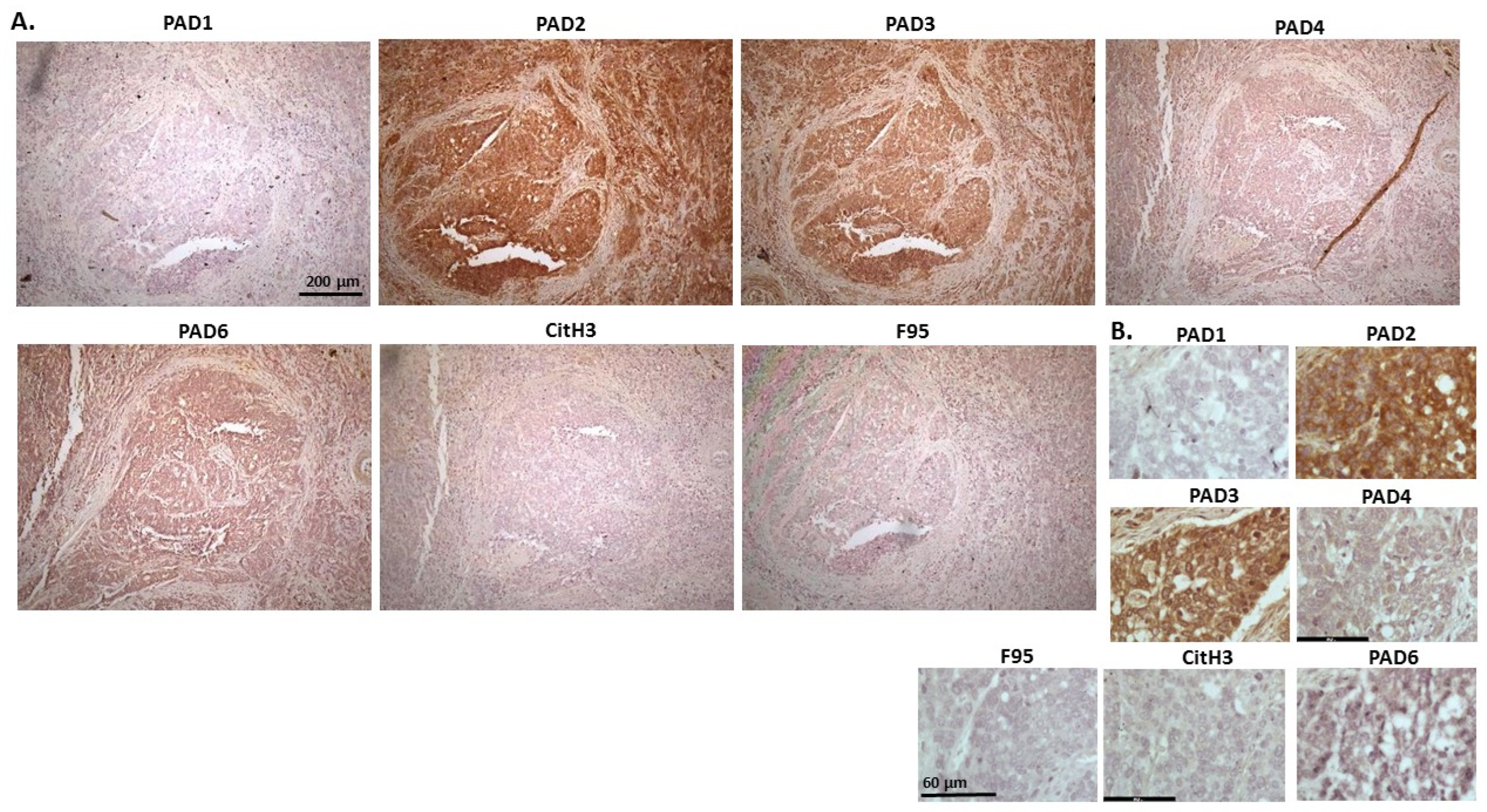
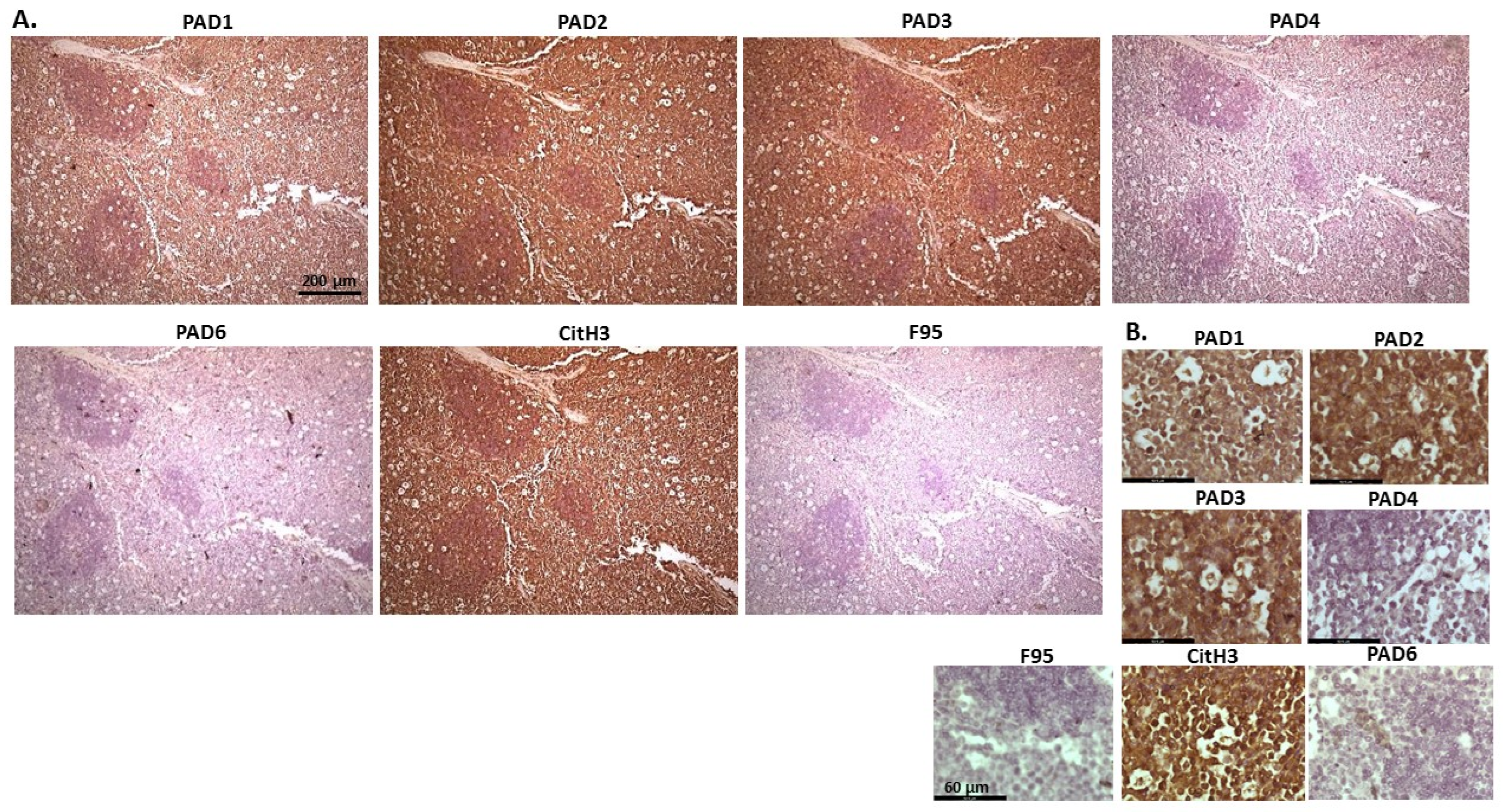
2.1. Detection of Protein Citrullination/Deimination, Deiminated Histone H3 and PAD Isoforms in Animal Cancer Tissues
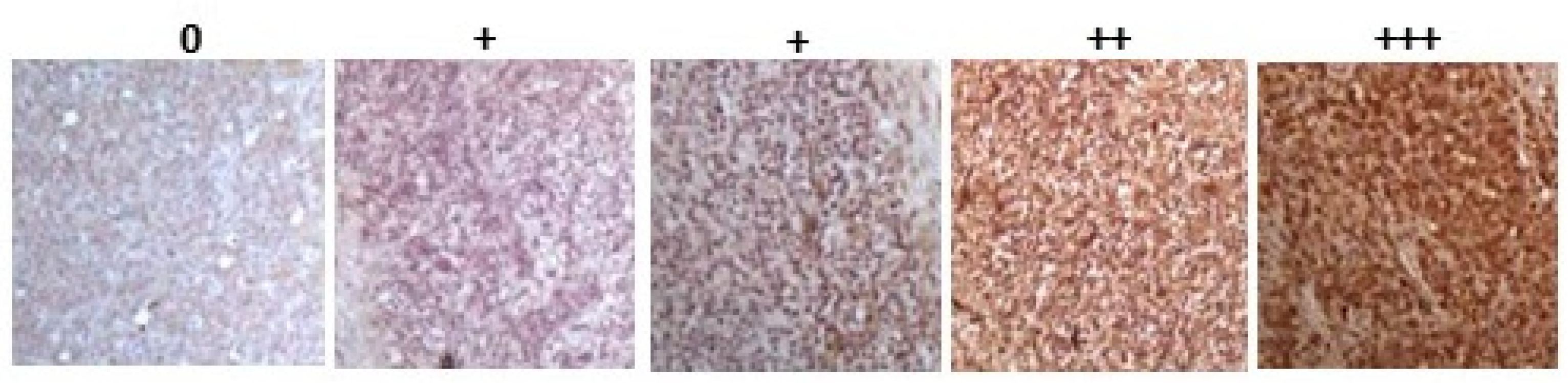

{kind=link}
{kind=link}
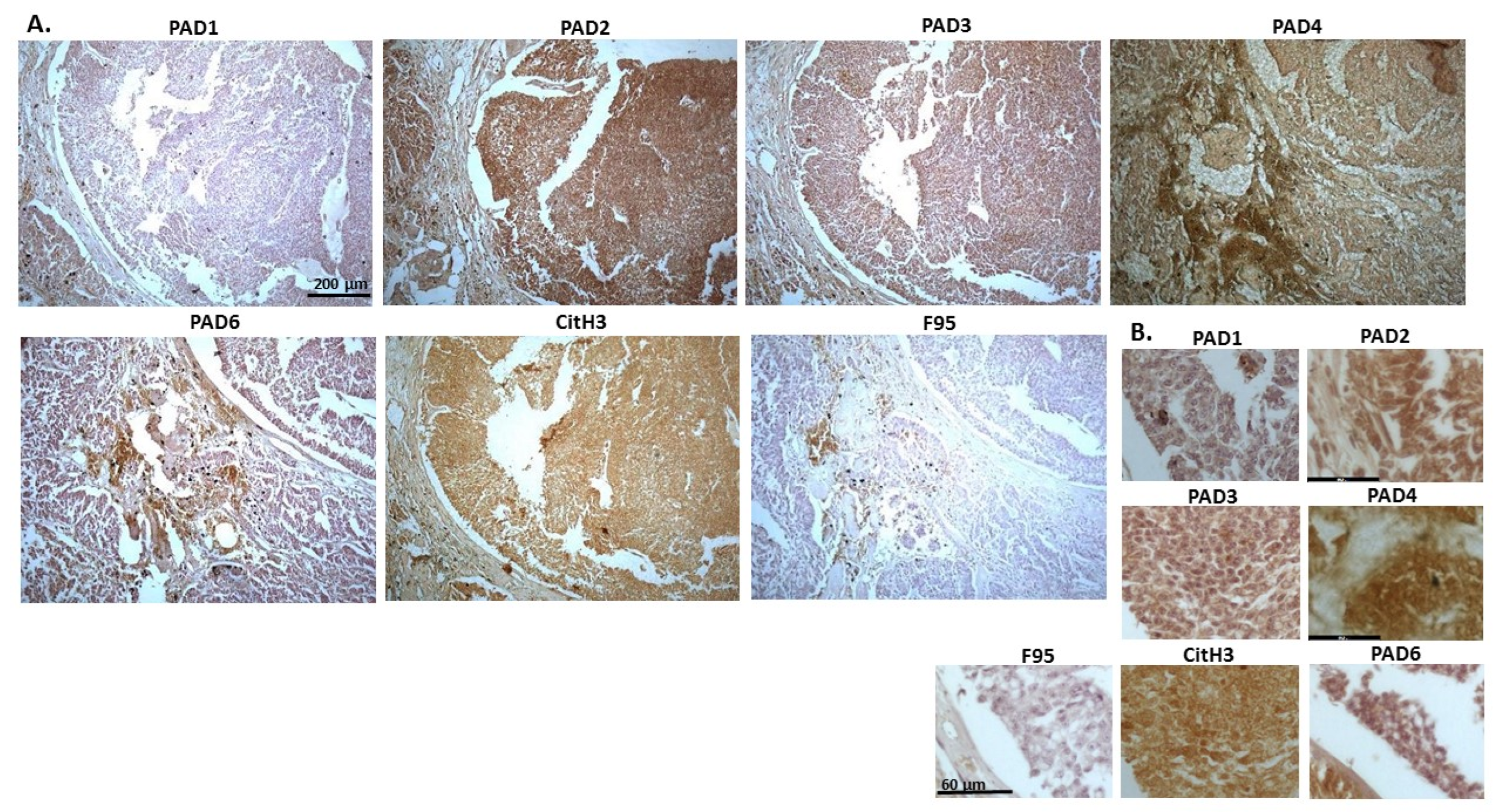{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
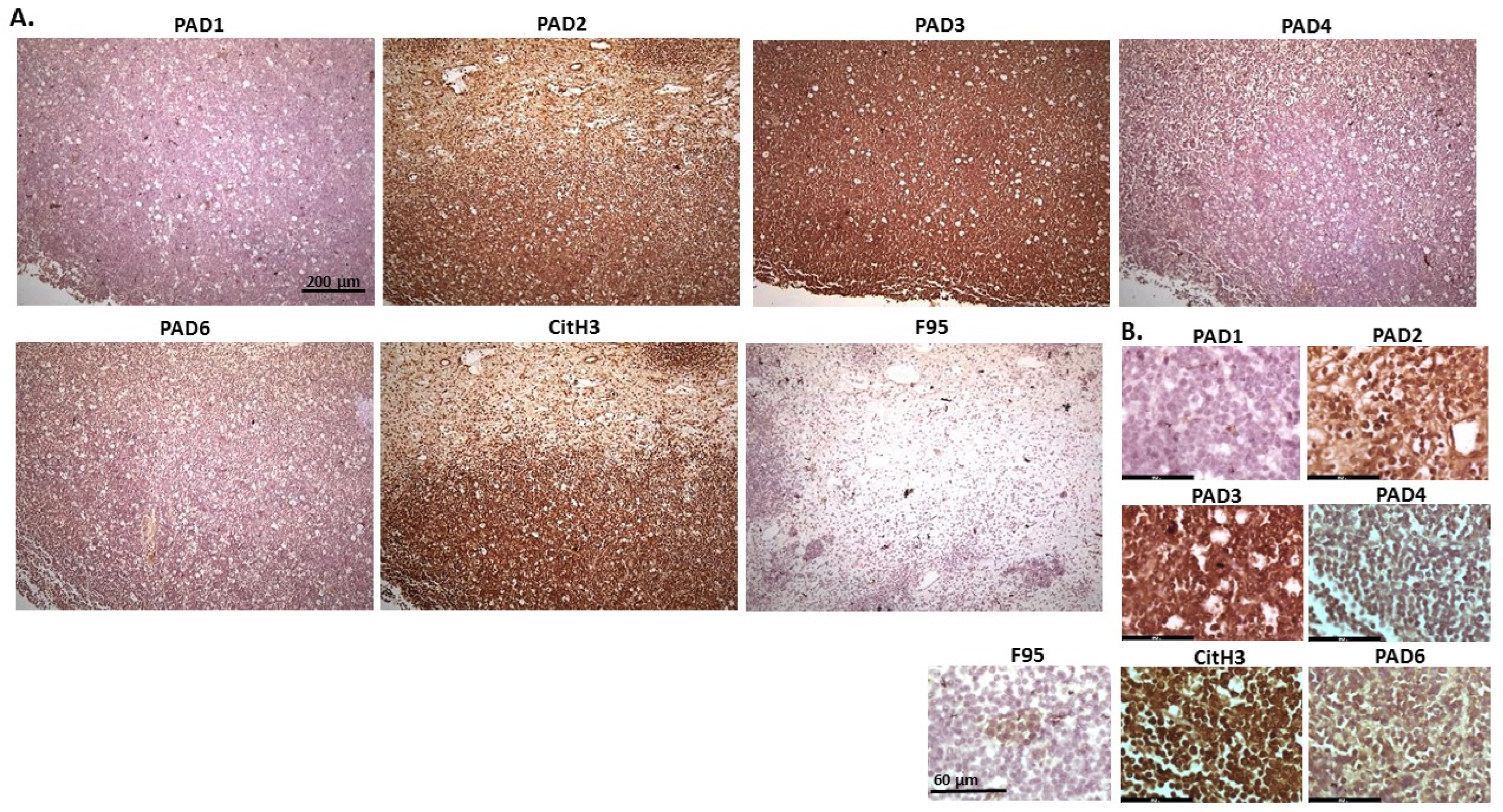{kind=link}
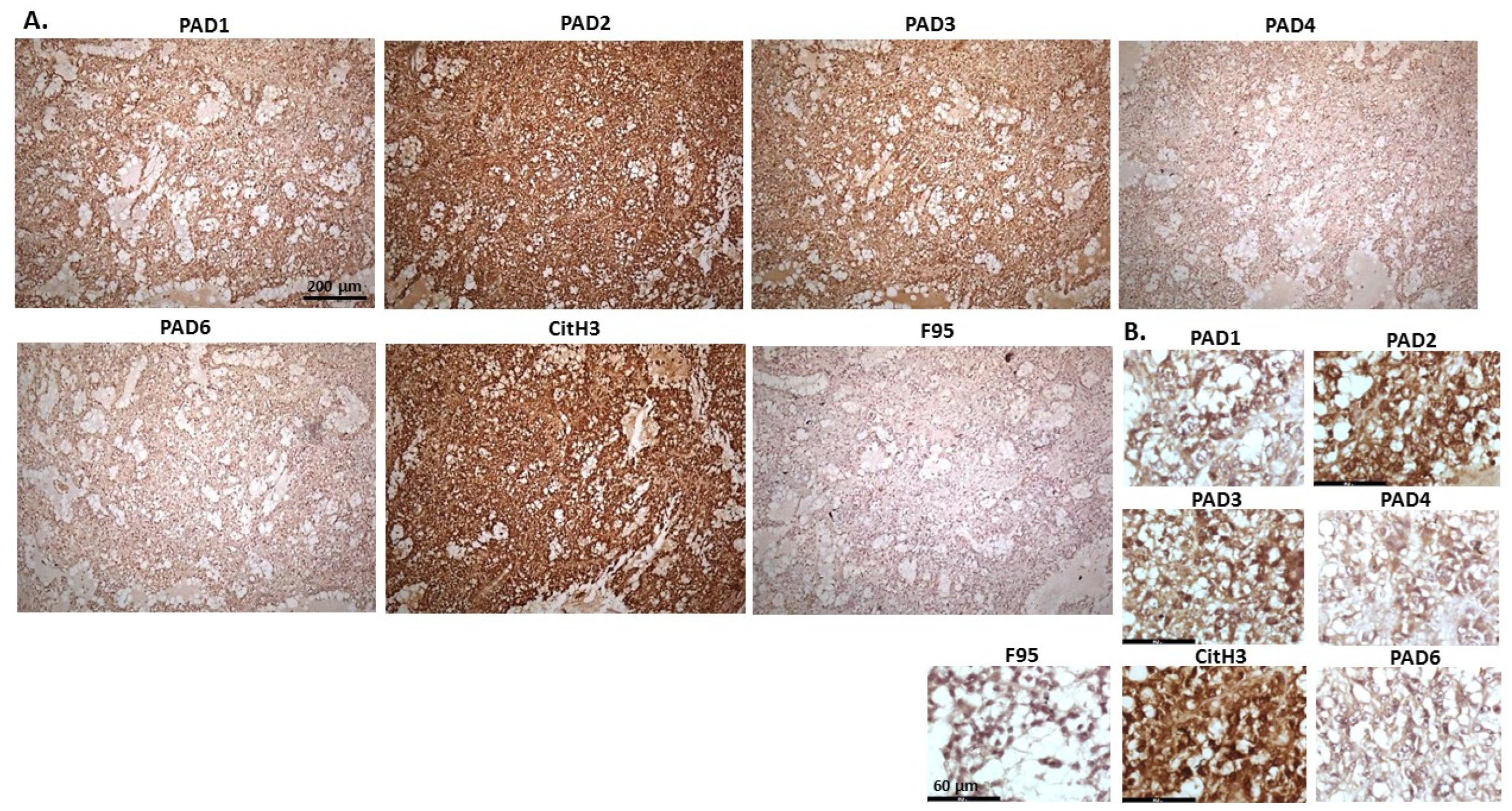{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Tumour | F95 | CitH3 | PAD1 | PAD2 | PAD3 | PAD4 | PAD6 |
|---|---|---|---|---|---|---|---|---|
| Rabbit (Oryctolagus cuniculus) | Mammary adenocarcinoma | + | ++ | ++ | +++ | +++ | ++ | ++ |
| Dog (Canis lupus familiaris) | (1) Mixed germ cell-sex cord stromal tumour, testicle. (2) Squamous cell tumour; origin of tissue unknown | (+) | ++ | + | +++ | ++ | ++ | + |
| Regent parrot (Polytelis anthopeplus) | Lymphoma, liver and spleen | + | +++ | + | ++ | +++ | + | + |
| Domestic duck (Anas platyrhynchos domesticus) | Malignant round cell tumour, liver; probably lymphoma | + | +++ | ++ | +++ | +++ | + | + |
| Mink (Neovision vision) | Malignant renal tumour, possibly nephroblastoma. Tumour in kidney with metastasis to lung and liver | (+) | +++ | (+) | +++ | ++ | + | ++ |
| Mink (Neovision vision) | Malignant renal tumour, probably nephroblastoma. Extensive necrosis and haemorrhage in tumour mass. | + | +++ | ++ | +++ | ++ | + | + |
| Cat (Felis catus) | Pulmonary carcinoma | + | +++ | ++ | +++ | +++ | ++ | + |
| Horse (Equus caballus) | Carcinoma in liver (ovary and mesenteric lymph node) | (+) | (+) | 0 | +++ | +++ | + | ++ |
| Cattle (Bos Taurus) | Neuroendocrine tumour (metastasis to the heart) | (+) | +++ | ++ | +++ | +++ | + | ++ |
| * Dog (Canis lupus familiaris) | * Anaplastic mammary carcinoma | ++ | +++ | ++ | +++ | ++ | ++ | ++ |
| Sheep (Ovis aries) | Malignant round cell tumour, skin | (+) | +++ | (+) | +++ | + | (+) | (+) |
| Hamster (Mesocricetus auratus) | Lymphoma, mesentery/small intestine | (+) | + | ++ | +++ | +++ | + | (+) |
| Dog (Canis lupus familiaris) | Scirrhous adenocarcinoma, mammary gland | + | +++ | ++ | +++ | ++ | + | + |
| Horse (Equus caballus) | Penile squamous cell carcinoma | 0 | +++ | (+) | +++ | +++ | ++ | + |
| Cattle (Bos Taurus) | Metastatic carcinoma, lungs and lymph nodes | 0 | +++ | + | +++ | ++ | ++ | + |
| Cattle (Bos Taurus) | Metastatic carcinoma, lungs and lymph nodes | (+) | +++ | ++ | +++ | +++ | ++ | + |
| Cat (Felis catus) | Malignant tumour, kidneys; prob. Lymphoma | + | +++ | + | +++ | +++ | + | + |
| Dog Canis lupus familiaris | Granulosa cell tumour, ovary | + | +++ | ++ | +++ | ++ | + | + |
| Cattle (Bos Taurus) | Lymphoma, lymph node | 0 | +++ | ++ | +++ | +++ | 0 | 0 |
| Reindeer (Rangifer tarandus) | Multicentric malignant tumour, probably mesothelioma | + | +++ | ++ | +++ | ++(+) | ++ | ++ |
| Pig (Sus scrofa domesticus) | Lymphoma (lymph node) | (+) | +++ | (+) | +++ | +++ | + | + |
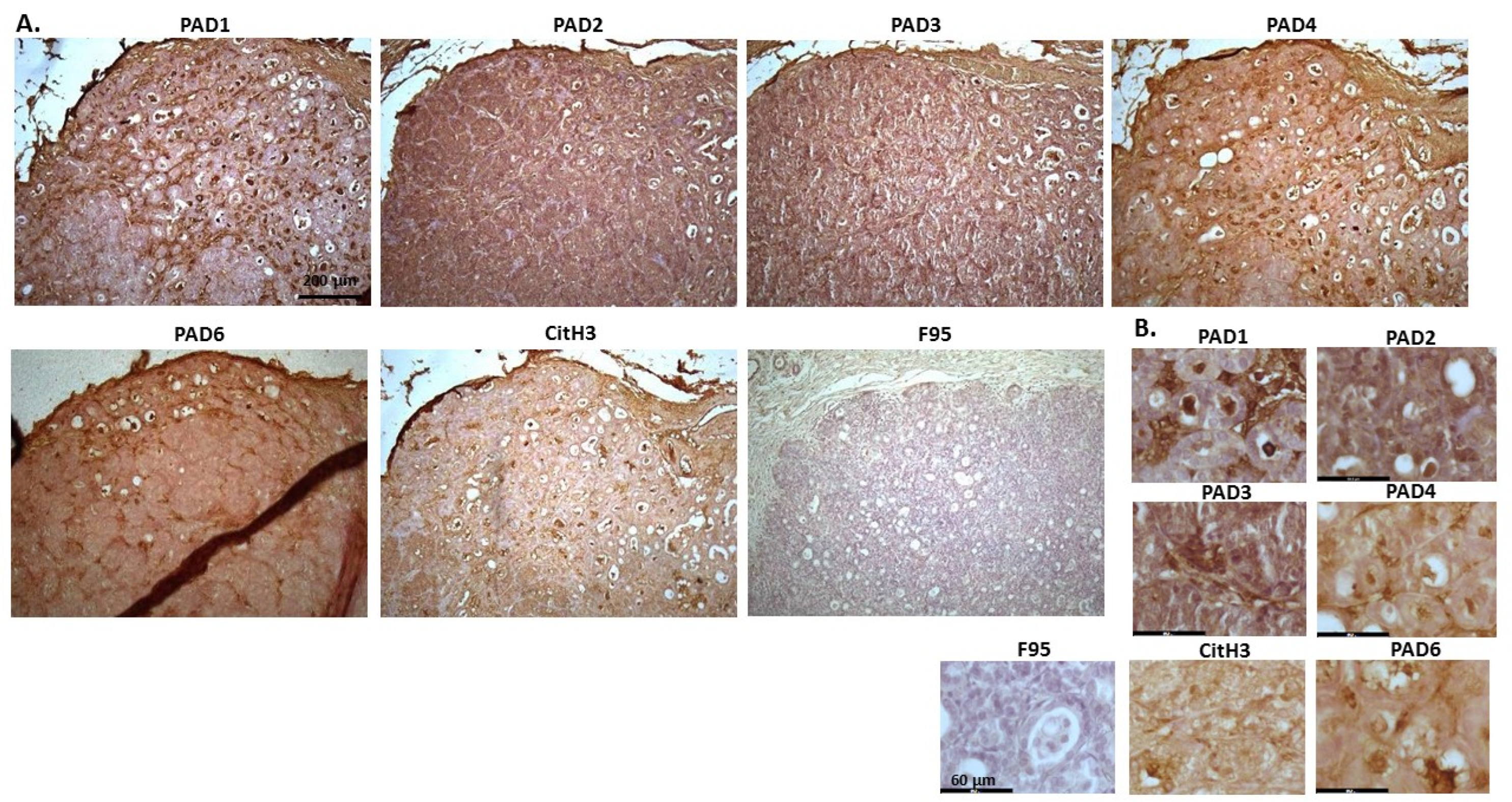

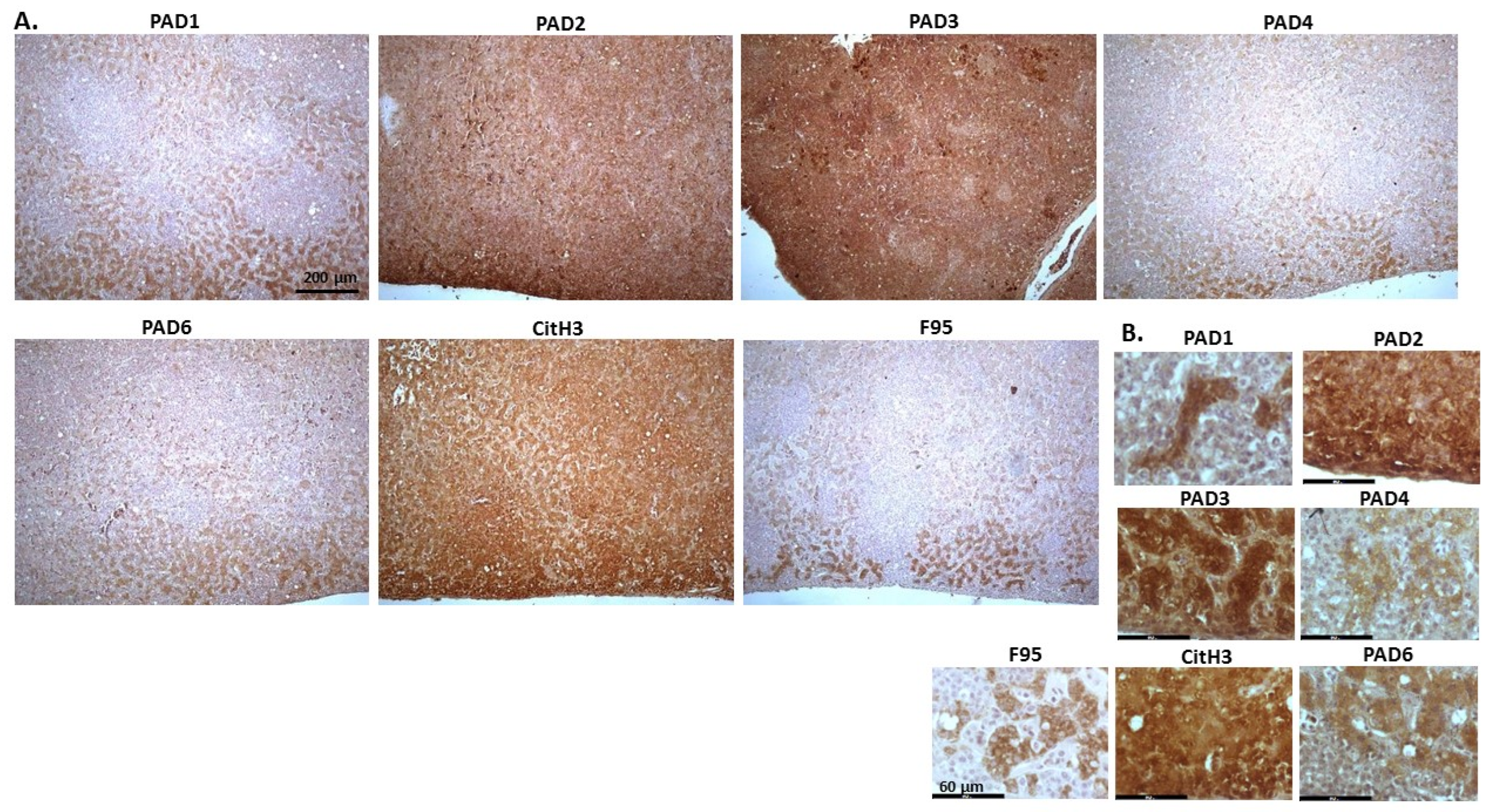
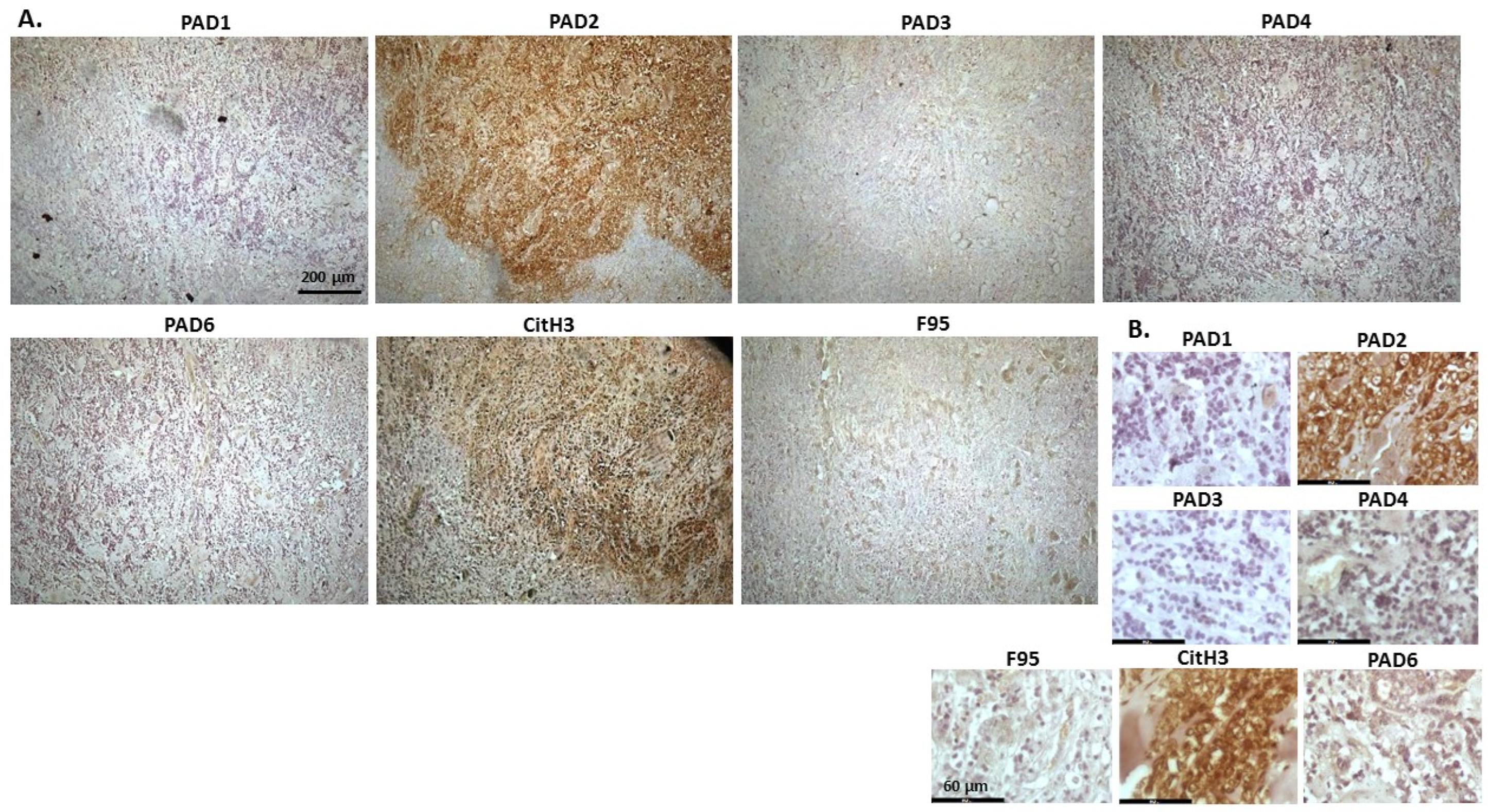
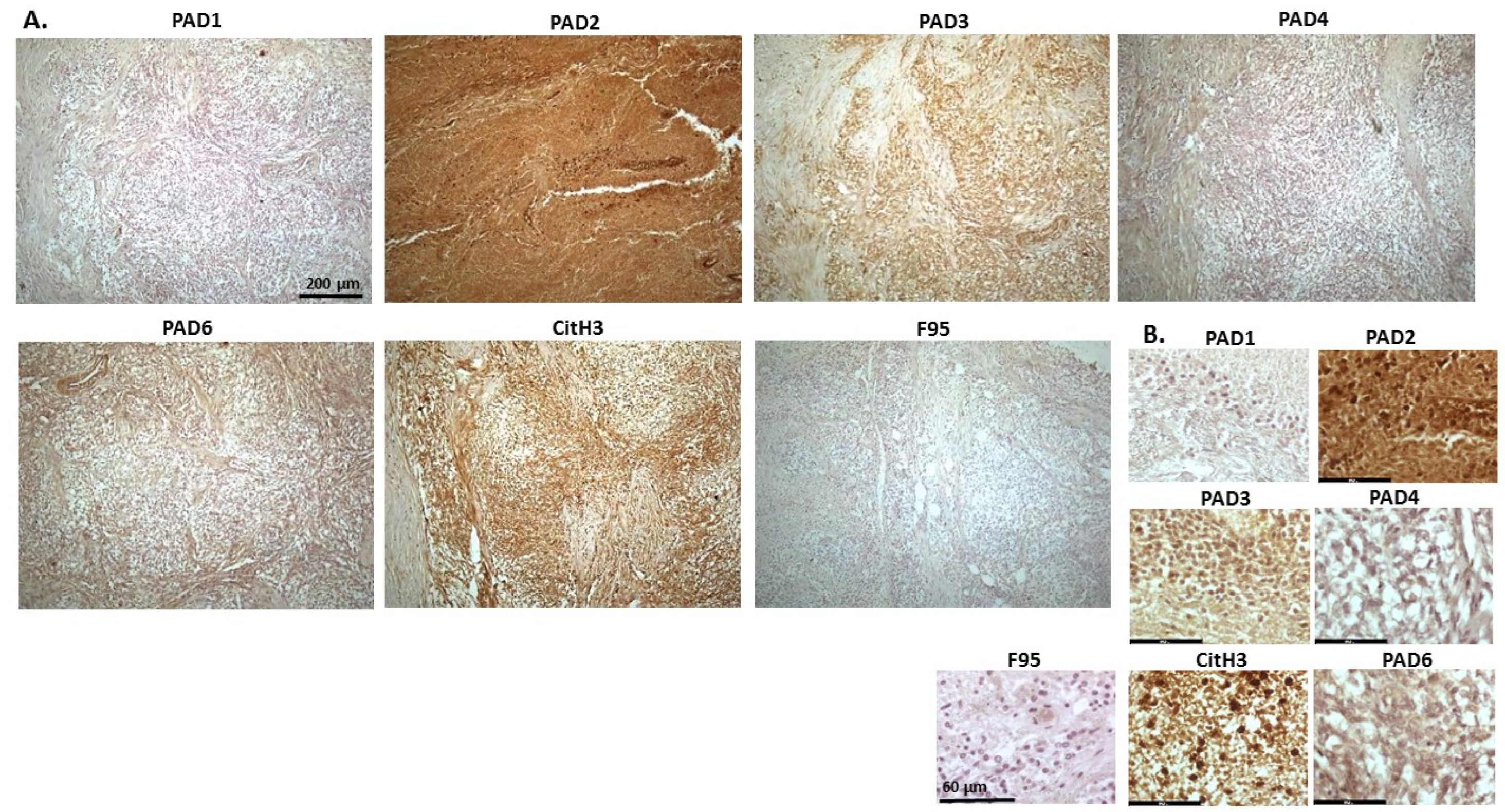
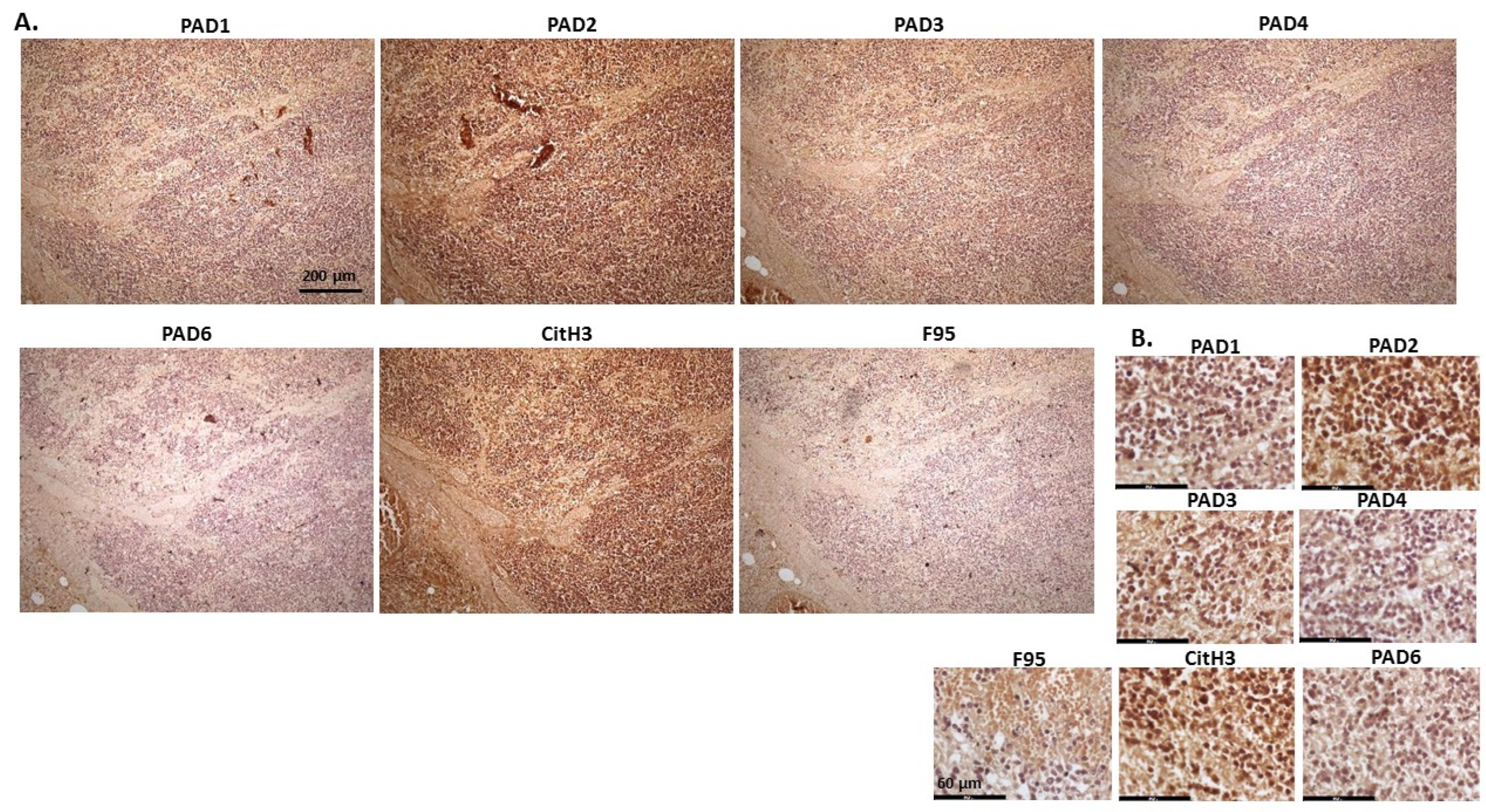
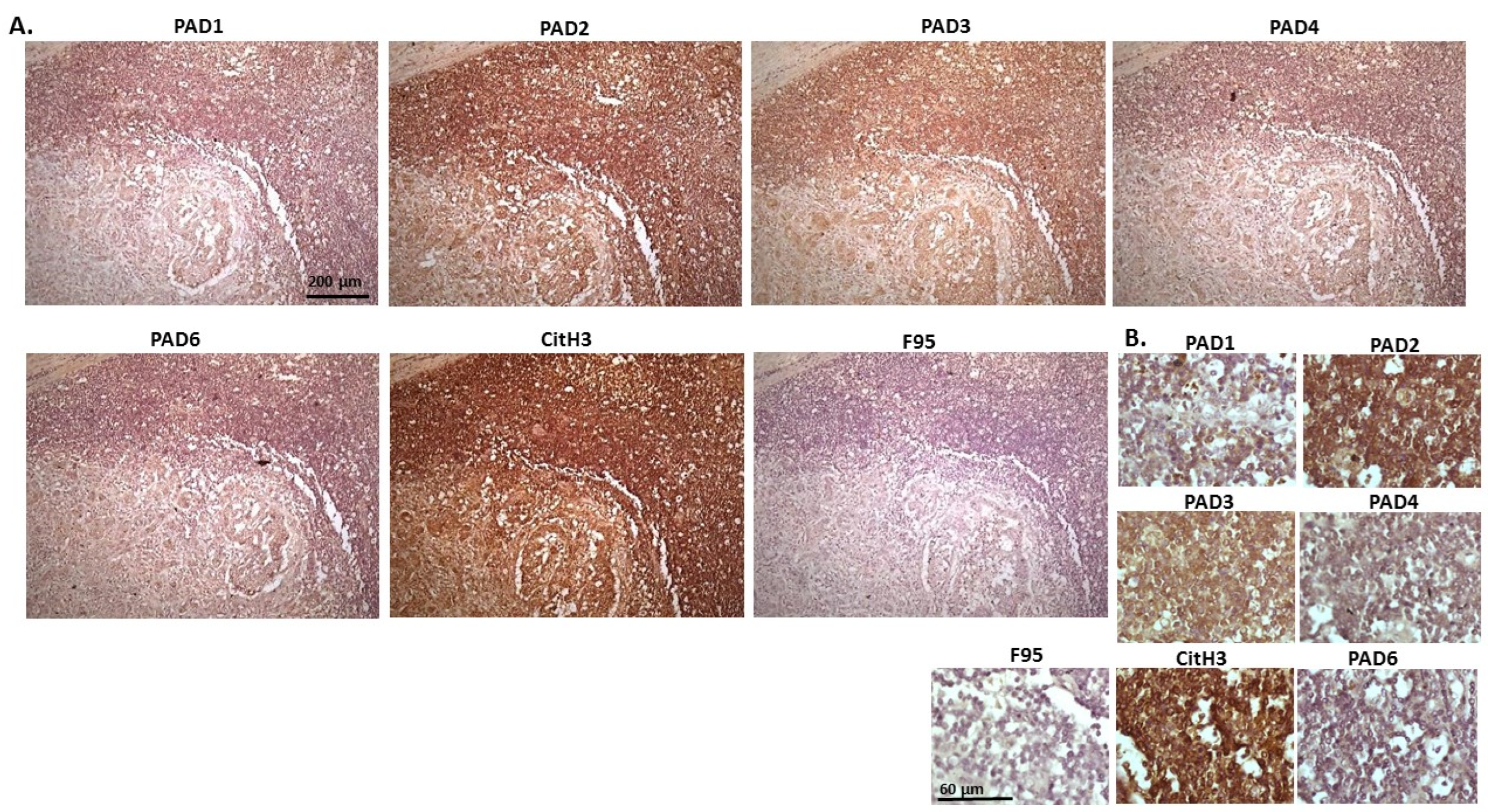
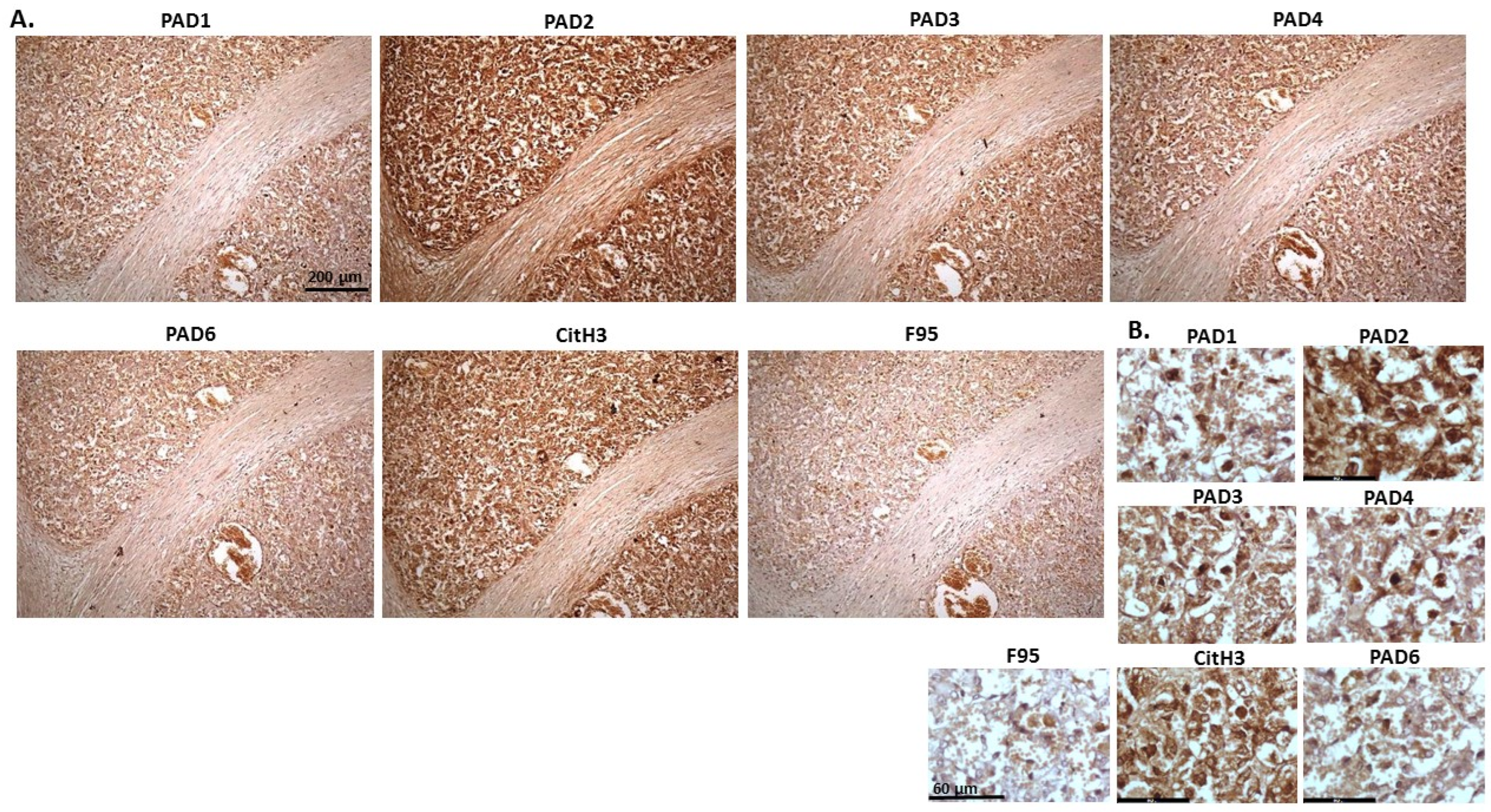
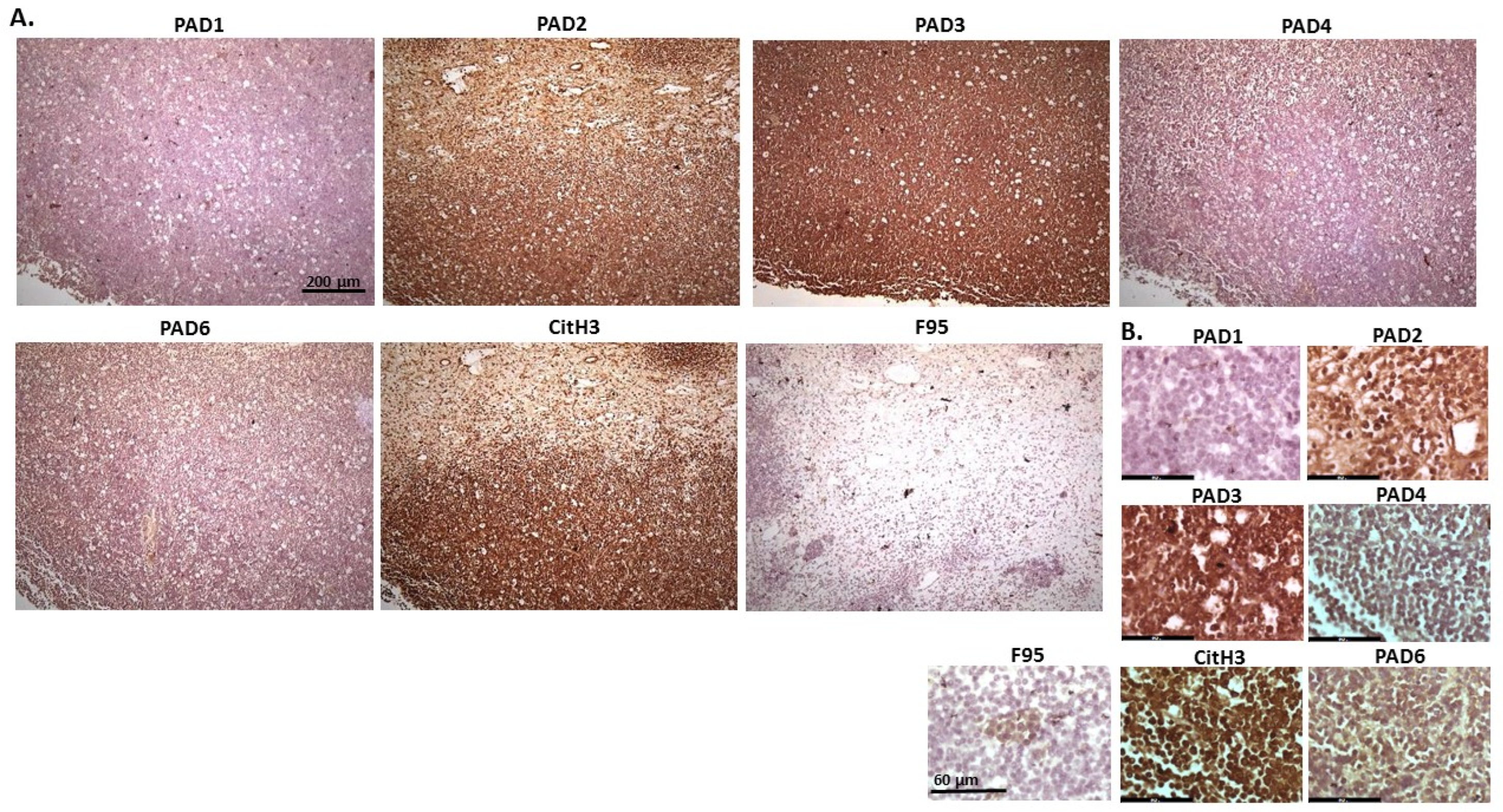
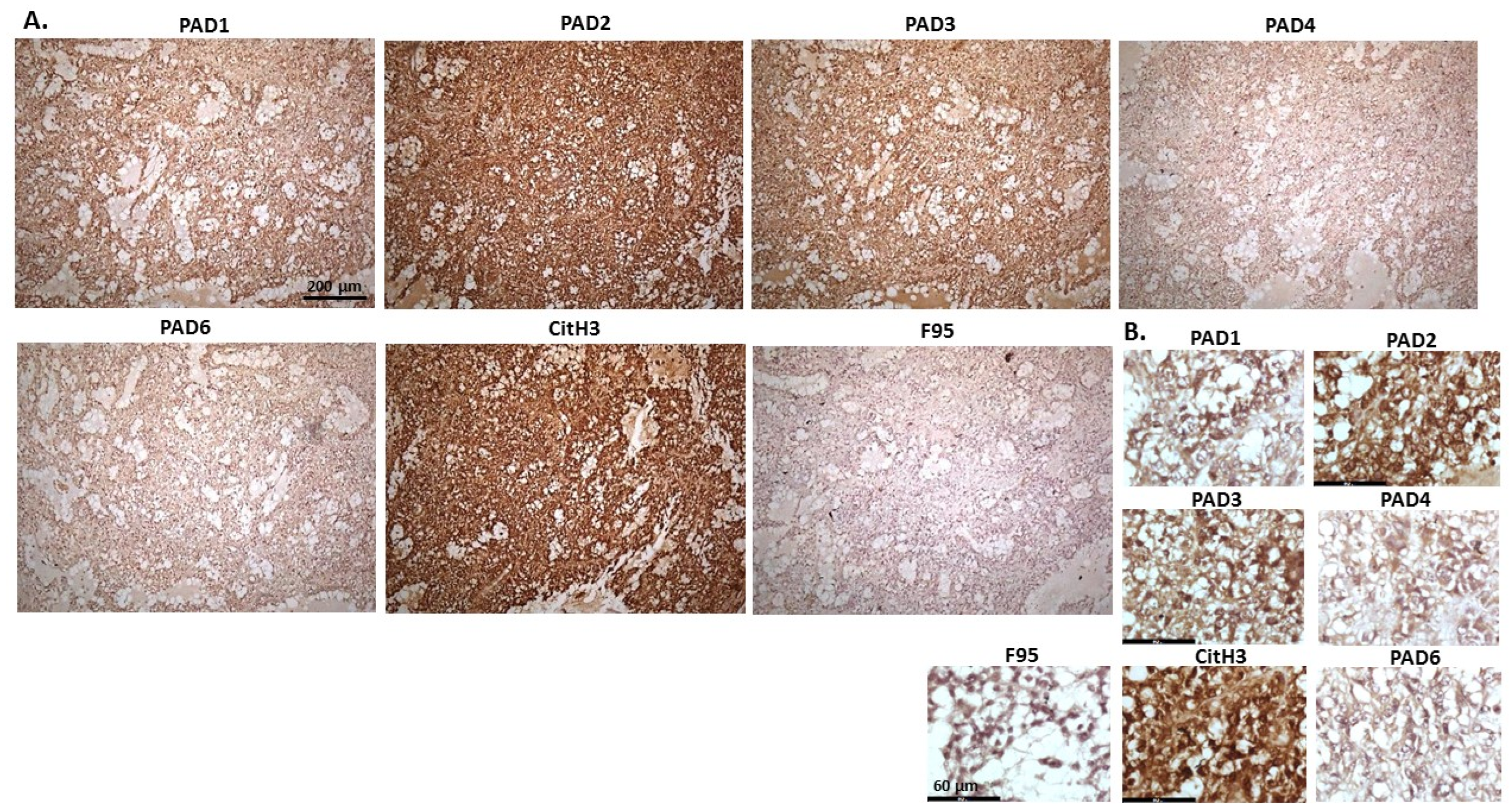
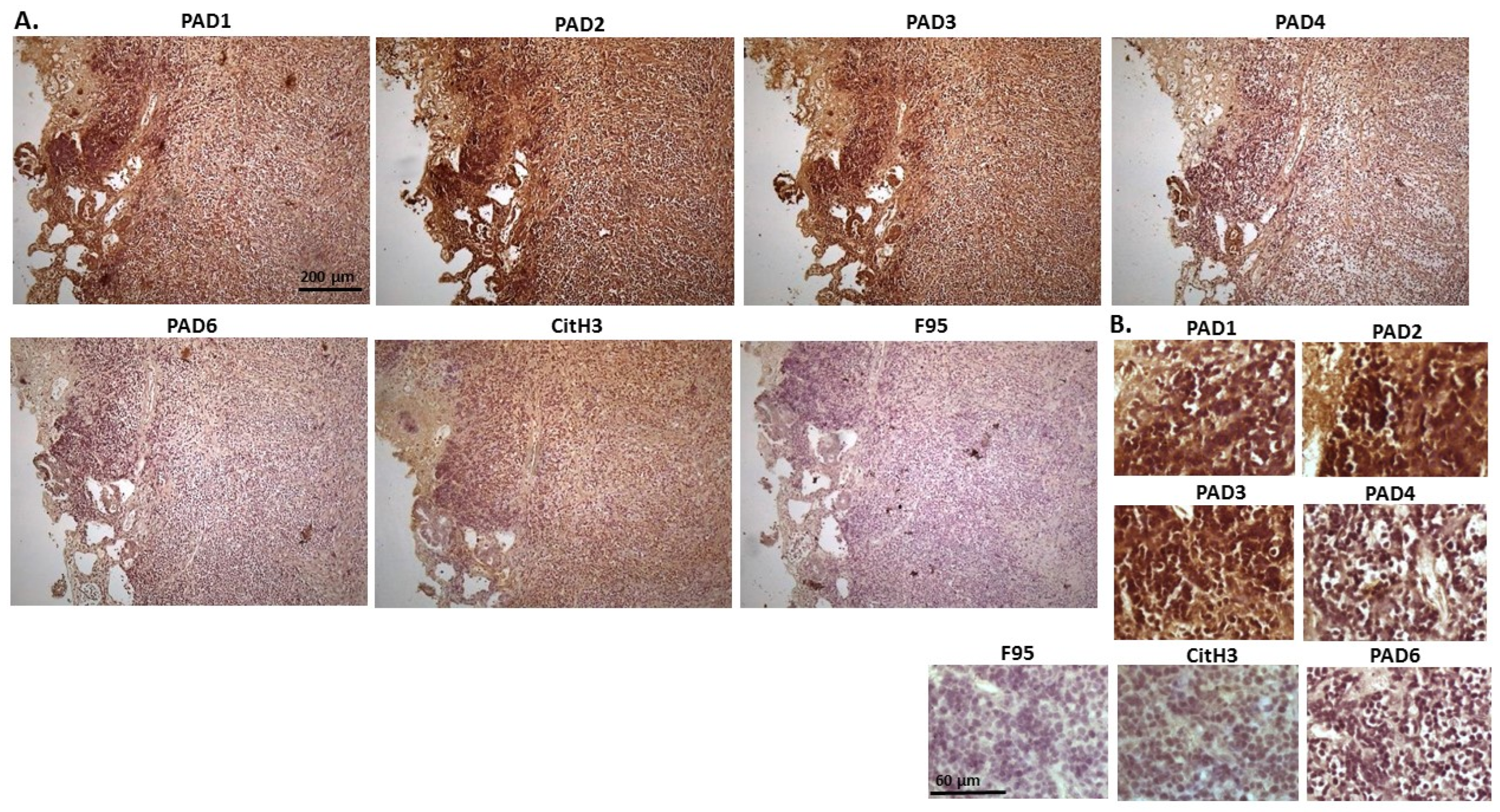
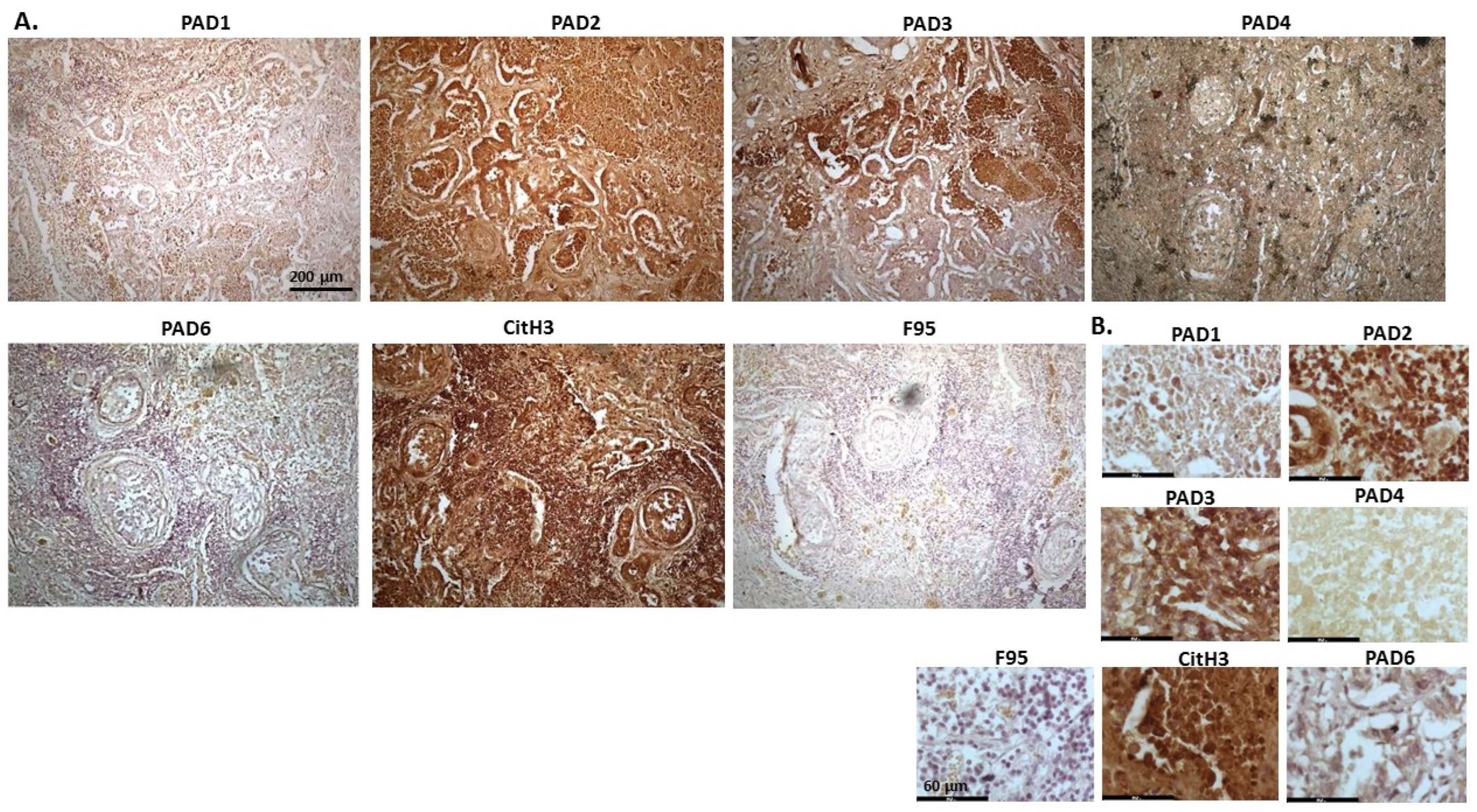
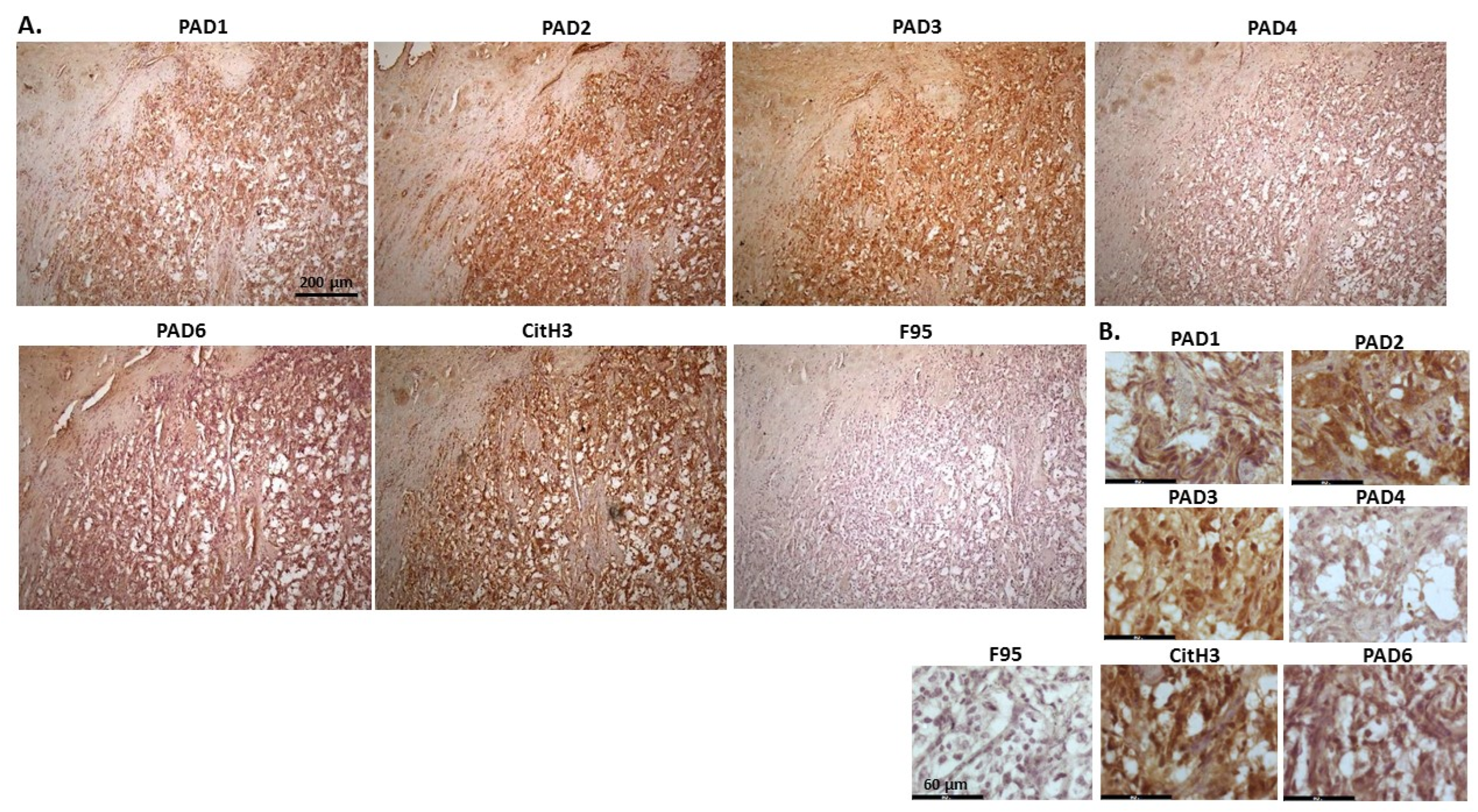
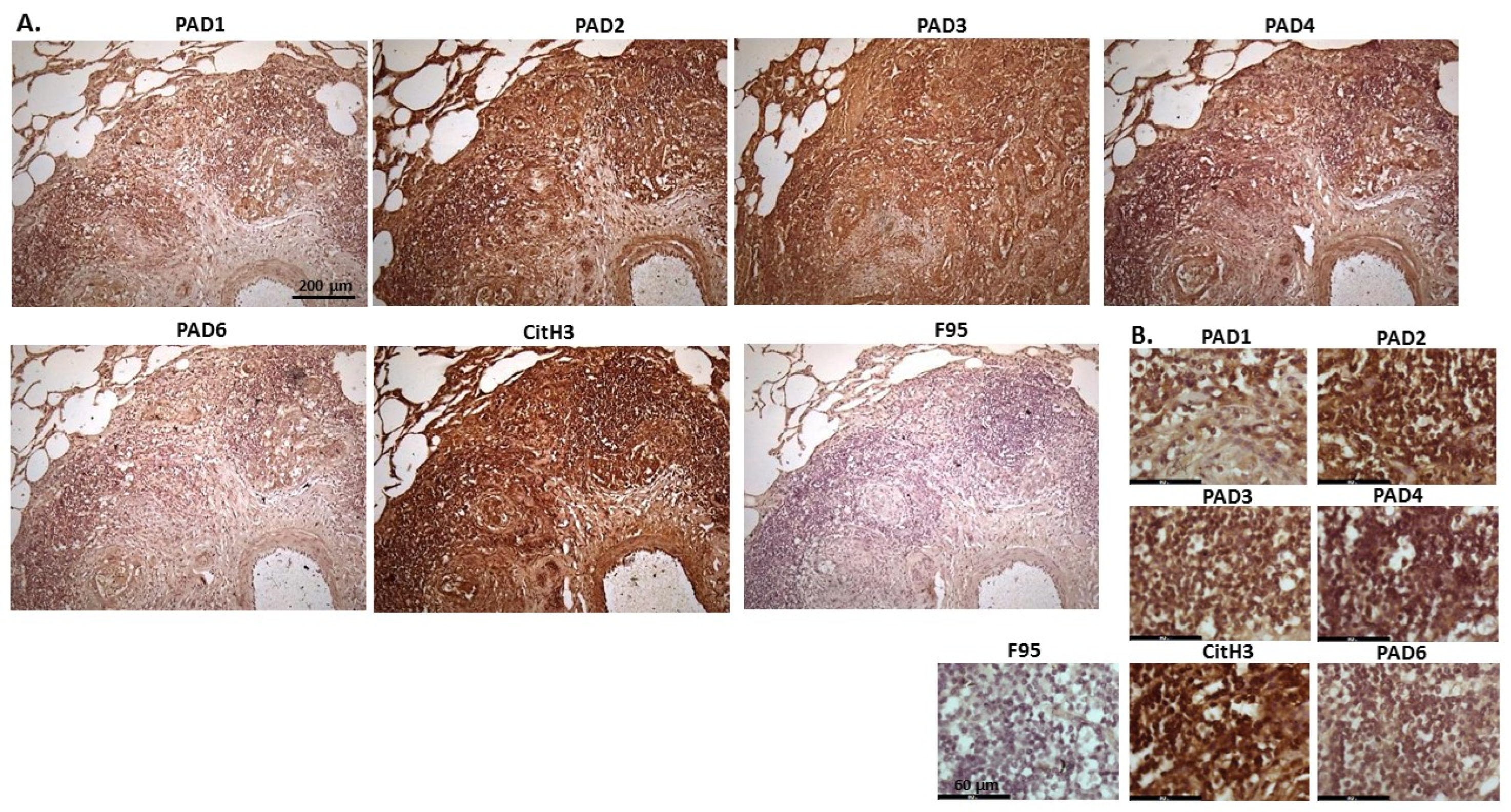


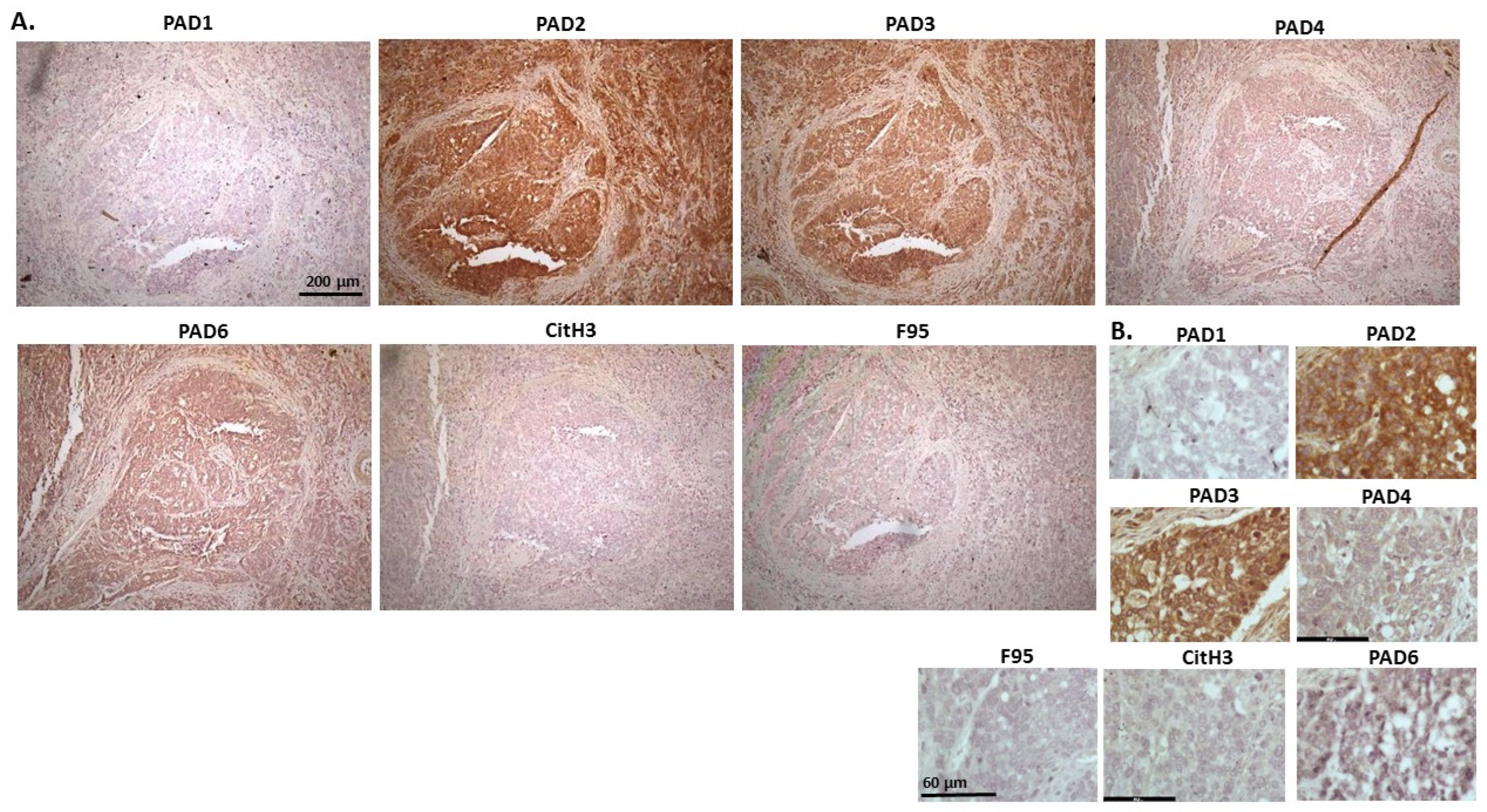
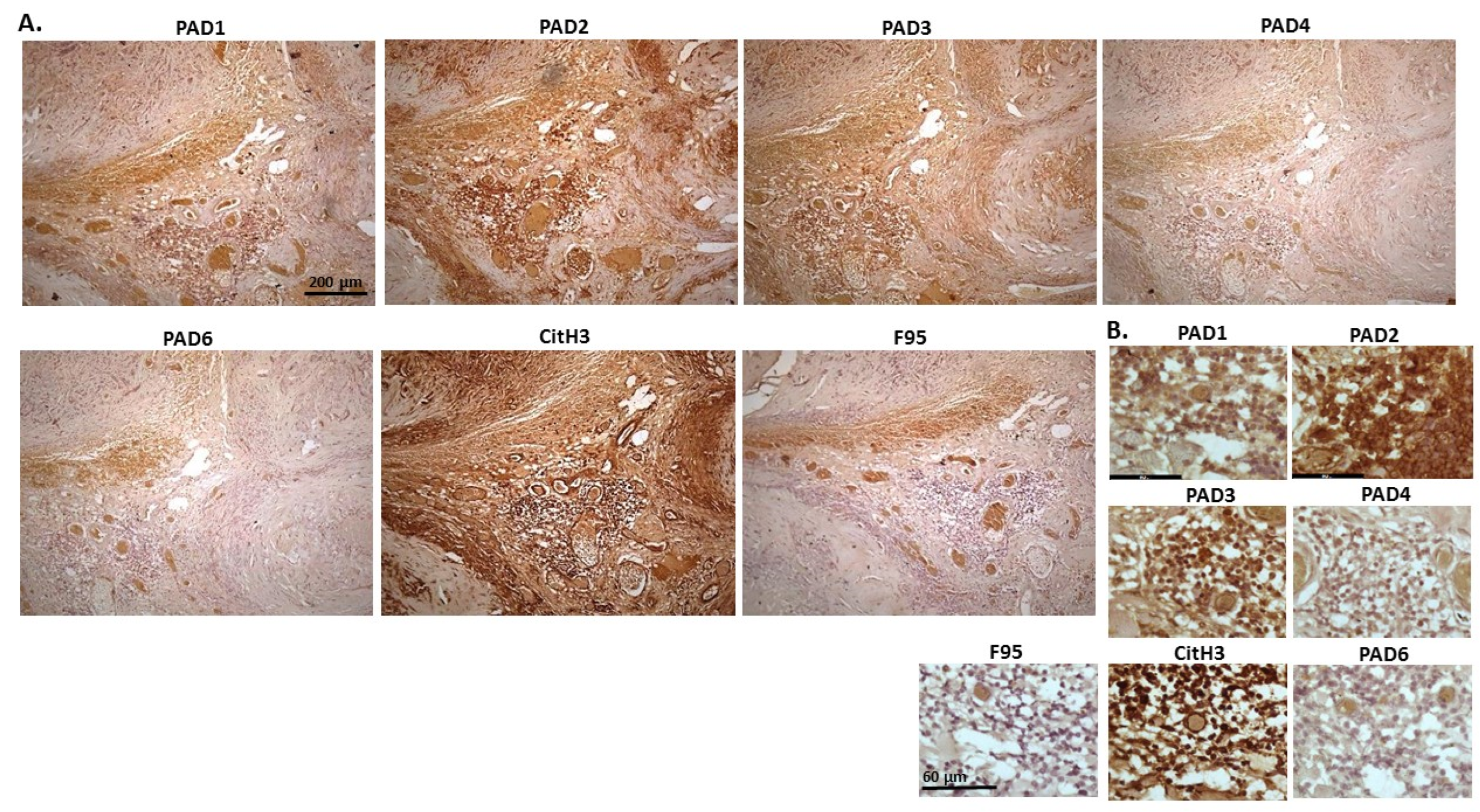
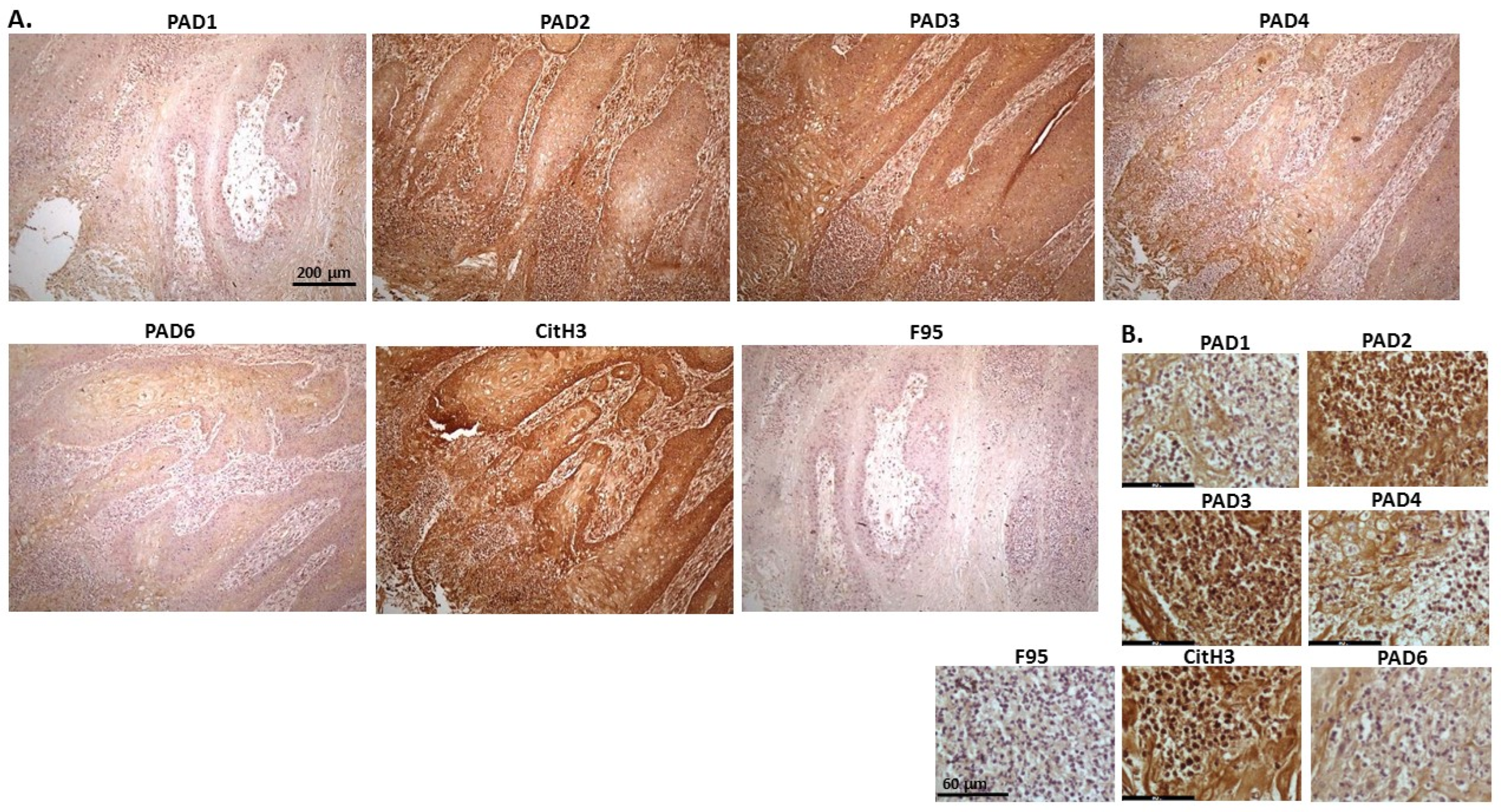
2.2. Phylogeny Tree Construction for PAD Isozymes in the Species under Study

3. Discussion
4. Materials and Methods
4.1. Tissue Sections from Animal Cancers
4.2. Immunohistochemical Analysis
4.3. Phylogenetic Tree Construction of PAD Isozymes in the Animal Species under Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohanan, S.; Cherrington, B.D.; Horibata, S.; McElwee, J.L.; Thompson, P.R.; Coonrod, S.A. Potential role of peptidylarginine deiminase enzymes and protein citrullination in cancer pathogenesis. Biochem. Res. Int. 2012, 2012, 895343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, S.; Gallagher, M.; Kholia, S.; Kosgodage, U.S.; Hristova, M.; Hardy, J.; Inal, J.M. Peptidylarginine deiminases-roles in cancer and neurodegeneration and possible avenues for therapeutic intervention via modulation of exosome and microvesicle (EMV) release? Int. J. Mol. Sci. 2017, 18, 1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brentville, V.A.; Vankemmelbeke, M.; Metheringham, R.L.; Durrant, L.G. Post-translational modifications such as citrullination are excellent targets for cancer therapy. Semin. Immunol. 2020, 47, 101393. [Google Scholar] [CrossRef] [PubMed]
- Vossenaar, E.R.; Zendman, A.J.; van Venrooij, W.J.; Pruijn, G.J. PAD, a growing family of citrullinating enzymes: Genes, features and involvement in disease. Bioessays 2003, 25, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Magnadóttir, B.; Hayes, P.; Hristova, M.; Bragason, B.T.; Nicholas, A.P.; Dodds, A.W.; Guðmundsdóttir, S.; Lange, S. Post-translational protein deimination in cod (Gadus morhua L.) ontogeny novel roles in tissue remodelling and mucosal immune defences? Dev. Comp. Immunol. 2018, 87, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Rast, J.P.; D’Alessio, S.; Kraev, I.; Lange, S. Post-translational protein deimination signatures in sea lamprey (Petromyzon marinus) plasma and plasma-extracellular vesicles. Dev. Comp. Immunol. 2021, 125, 104225. [Google Scholar] [CrossRef] [PubMed]
- D’Alessio, S.; Buckley, K.M.; Kraev, I.; Hayes, P.; Lange, S. Extracellular vesicle signatures and post-translational protein deimination in purple sea urchin (Strongylocentrotus purpuratus) coelomic fluid-novel insights into echinodermata biology. Biology 2021, 10, 866. [Google Scholar] [CrossRef] [PubMed]
- Cummings, T.F.M.; Gori, K.; Sanchez-Pulido, L.; Gavriilidis, G.; Moi, D.; Wilson, A.R.; Murchison, E.; Dessimoz, C.; Ponting, C.P.; Christophorou, M.A. Protein citrullination was introduced into animals by horizontal gene transfer from cyanobacteria. Mol. Biol. Evol. 2022, 39, msab317. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Thompson, P.R. Protein arginine deiminases (PADs): Biochemistry and chemical biology of protein citrullination. Acc. Chem. Res. 2019, 52, 818–832. [Google Scholar] [CrossRef] [PubMed]
- Méchin, M.C.; Takahara, H.; Simon, M. Deimination and peptidylarginine deiminases in skin physiology and diseases. Int. J. Mol. Sci. 2020, 21, 566. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liu, X.; Zhang, M.; Li, T.; Muth, A.; Thompson, P.R.; Coonrod, S.A.; Zhang, X. Peptidylarginine deiminase 1-catalyzed histone citrullination is essential for early embryo development. Sci. Rep. 2016, 6, 38727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, S.; Gong, X.; Zhang, X.; Salinger, A.J.; Zheng, L.; Sen, S.; Weerapana, E.; Zhang, X.; Thompson, P.R. Halogen bonding increases the potency and isozyme selectivity of protein arginine deiminase 1 inhibitors. Angew. Chem. Int. Ed. Engl. 2019, 58, 12476–12480. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Liu, X.; Li, F.; Miao, L.; Li, T.; Xu, B.; An, X.; Muth, A.; Thompson, P.R.; Coonrod, S.A.; et al. PAD1 promotes epithelial-mesenchymal transition and metastasis in triple-negative breast cancer cells by regulating MEK1-ERK1/2-MMP2 signaling. Cancer Lett. 2017, 409, 30–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cau, L.; Takahara, H.; Thompson, P.R.; Serre, G.; Méchin, M.C.; Simon, M. Peptidylarginine deiminase inhibitor Cl-amidine attenuates cornification and interferes with the regulation of autophagy in reconstructed human epidermis. J. Investig. Dermatol. 2019, 139, 1889–1897.e4. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.; Gögel, S.; Leung, K.Y.; Vernay, B.; Nicholas, A.P.; Causey, C.P.; Thompson, P.R.; Greene, N.D.; Ferretti, P. Protein deiminases: New players in the developmentally regulated loss of neural regenerative ability. Dev. Biol. 2011, 355, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Lange, S.; Rocha-Ferreira, E.; Thei, L.; Mawjee, P.; Bennett, K.; Thompson, P.R.; Subramanian, V.; Nicholas, A.P.; Peebles, D.; Hristova, M.; et al. Peptidylarginine deiminases: Novel drug targets for prevention of neuronal damage following hypoxic ischemic insult (HI) in neonates. J. Neurochem. 2014, 130, 555–562. [Google Scholar] [CrossRef]
- Subramanian, V.; Nicholas, A.P.; Thompson, P.R.; Ferretti, P. Modulation of calcium-induced cell death in human neural stem cells by the novel peptidylarginine deiminase-AIF pathway. Biochim. Biophys. Acta 2014, 1843, 1162–1171. [Google Scholar]
- Kosgodage, U.S.; Uysal-Onganer, P.; MacLatchy, A.; Kraev, I.; Chatterton, N.P.; Nicholas, A.P.; Inal, J.M.; Lange, S. Peptidylarginine deiminases post-translationally deiminate prohibitin and modulate extracellular vesicle release and microRNAs in glioblastoma multiforme. Int. J. Mol. Sci. 2018, 20, 103. [Google Scholar] [CrossRef] [Green Version]
- Uysal-Onganer, P.; MacLatchy, A.; Mahmoud, R.; Kraev, I.; Thompson, P.R.; Inal, J.M.; Lange, S. Peptidylarginine deiminase isozyme-specific PAD2, PAD3 and PAD4 inhibitors differentially modulate extracellular vesicle signatures and cell invasion in two glioblastoma multiforme cell lines. Int. J. Mol. Sci. 2020, 21, 1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uysal-Onganer, P.; D’Alessio, S.; Mortoglou, M.; Kraev, I.; Lange, S. Peptidylarginine deiminase inhibitor application, using Cl-amidine, PAD2, PAD3 and PAD4 isozyme-specific inhibitors in pancreatic cancer cells, reveals roles for PAD2 and PAD3 in cancer invasion and modulation of extracellular vesicle signatures. Int. J. Mol. Sci. 2021, 22, 1396. [Google Scholar] [CrossRef]
- Foulquier, C.; Sebbag, M.; Clavel, C.; Chapuy-Regaud, S.; Al Badine, R.; Méchin, M.C.; Vincent, C.; Nachat, R.; Yamada, M.; Takahara, H.; et al. Peptidyl arginine deiminase type 2 (PAD-2) and PAD-4 but not PAD-1, PAD-3, and PAD-6 are expressed in rheumatoid arthritis synovium in close association with tissue inflammation. Arthritis Rheum. 2007, 56, 3541–3553. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, R.; Gan, Y.; Ying, S. The roles of PAD2- and PAD4-mediated protein citrullination catalysis in cancers. Int. J. Cancer 2021, 148, 267–276. [Google Scholar] [CrossRef]
- Esposito, G.; Vitale, A.M.; Leijten, F.P.; Strik, A.M.; Koonen-Reemst, A.M.; Yurttas, P.; Robben, T.J.; Coonrod, S.; Gossen, J.A. Peptidylarginine deiminase (PAD) 6 is essential for oocyte cytoskeletal sheet formation and female fertility. Mol. Cell. Endo-crinol. 2007, 273, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurttas, P.; Vitale, A.M.; Fitzhenry, R.J.; Cohen-Gould, L.; Wu, W.; Gossen, J.A.; Coonrod, S.A. Role for PADI6 and the cytoplasmic lattices in ribosomal storage in oocytes and translational control in the early mouse embryo. Development 2008, 135, 2627–2636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kan, R.; Yurttas, P.; Kim, B.; Jin, M.; Wo, L.; Lee, B.; Gosden, R.; Coonrod, S.A. Regulation of mouse oocyte microtubule and organelle dynamics by PADI6 and the cytoplasmic lattices. Dev. Biol. 2011, 350, 311–322. [Google Scholar] [CrossRef] [Green Version]
- Horibata, S.; Coonrod, S.A.; Cherrington, B.D. Role for peptidylarginine deiminase enzymes in disease and female reproduction. J. Reprod. Dev. 2012, 58, 274–282. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Morency, E.; Li, T.; Qin, H.; Zhang, X.; Zhang, X.; Coonrod, S. Role for PADI6 in securing the mRNA-MSY2 complex to the oocyte cytoplasmic lattices. Cell Cycle 2017, 16, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Shi, Y.; Fu, J.; Yu, M.; Feng, R.; Sang, Q.; Liang, B.; Chen, B.; Qu, R.; Li, B.; et al. Mutations in PADI6 cause female infertility characterized by early embryonic arrest. Am. J. Hum. Genet. 2016, 99, 744–752. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Zheng, Y.; Xu, B.; Ma, F.; Li, C.; Zhang, X.; Wang, Y.; Chang, X. Investigating the expression, effect and tumorigenic pathway of PADI2 in tumors. OncoTargets Ther. 2017, 10, 1475–1485. [Google Scholar] [CrossRef] [Green Version]
- Cantarino, N.; Musulen, E.; Valero, V.; Peinado, M.A.; Perucho, M.; Moreno, V.; Forcales, S.V.; Douet, J.; Buschbeck, M. Downregulation of the deiminase PADI2 is an early event in colorectal carcinogenesis and indicates poor prognosis. Mol. Cancer Res. 2016, 14, 841–848. [Google Scholar] [CrossRef] [Green Version]
- Cherrington, B.D.; Zhang, X.; McElwee, J.L.; Morency, E.; Anguish, L.J.; Coonrod, S.A. Potential role for PAD2 in gene regulation in breast cancer cells. PLoS ONE 2012, 7, e41242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuzhalin, A.E. Citrullination in cancer. Cancer Res. 2019, 79, 1274–1284. [Google Scholar] [CrossRef] [Green Version]
- Cherrington, B.D.; Morency, E.; Struble, A.M.; Coonrod, S.A.; Wakshlag, J.J. Potential role for peptidylarginine deiminase 2 (PAD2) in citrullination of canine mammary epithelial cell histones. PLoS ONE 2010, 5, e11768. [Google Scholar] [CrossRef] [Green Version]
- Ledet, M.M.; Anderson, R.; Harman, R.; Muth, A.; Thompson, P.R.; Coonrod, S.A.; Van de Walle, G.R. BB-Cl-Amidine as a novel therapeutic for canine and feline mammary cancer via activation of the endoplasmic reticulum stress pathway. BMC Cancer 2018, 18, 412. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, A.P.; Whitaker, J.N. Preparation of a monoclonal antibody to citrullinated epitopes: Its characterization and some applications to immunohistochemistry in human brain. Glia 2002, 37, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Hamam, H.J.; Palaniyar, N. Post-translational modifications in NETosis and NETs-mediated diseases. Biomolecules 2019, 9, 369. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Lin, C.; Deng, H.; Strnad, J.; Bernabei, L.; Vogl, D.T.; Burke, J.J.; Nefedova, Y. A novel peptidylarginine deiminase 4 (PAD4) inhibitor BMS-P5 blocks formation of neutrophil extracellular traps and delays progression of multiple myeloma. Mol. Cancer Ther. 2020, 19, 1530–1538. [Google Scholar] [CrossRef] [PubMed]
- Francischetti, I.M.B.; Alejo, J.C.; Sivanandham, R.; Davies-Hill, T.; Fetsch, P.; Pandrea, I.; Jaffe, E.S.; Pittaluga, S. Neutrophil and eosinophil extracellular traps in hodgkin lymphoma. Hemasphere 2021, 5, e633. [Google Scholar] [CrossRef]
- Baecklund, E.; Backlin, C.; Rönnelid, J.; Toes, R.; Huizinga, T.; Åhlin, E.; Askling, J.; Hochberg, F.H.; Klareskog, L.; Kay, J.; et al. Anti-cyclic citrullinated peptide antibodies, other common autoantibodies, and smoking as risk factors for lymphoma in patients with rheumatoid arthritis. Scand. J. Rheumatol. 2018, 47, 270–275. [Google Scholar] [CrossRef]
- Assmann, G.; Shihadeh, K.; Poeschel, V.; Murawski, N.; Conigliarou, J.; Ong, M.F.; Pfreundschuh, M. Prevalence of anti-citrullinated protein antibodies (ACPA) in patients with diffuse large B-cell lymphoma (DLBCL): A case-control study. PLoS ONE 2014, 9, e88177. [Google Scholar] [CrossRef] [Green Version]
- Bagchi, A.; Mills, A.A. The quest for the 1p36 tumor suppressor. Cancer Res. 2008, 68, 2551–2556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellsworth, R.E.; Vertrees, A.; Love, B.; Hooke, J.A.; Ellsworth, D.L.; Shriver, C.D. Chromosomal alterations associated with the transition from in situ to invasive breast cancer. Ann. Surg. Oncol. 2008, 15, 2519–2525. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Han, J.; Pang, L.; Zhao, Y.; Yang, Y.; Shen, Z. Increased PADI4 expression in blood and tissues of patients with malignant tumors. BMC Cancer 2009, 9, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherrington, B.D.; Mohanan, S.; Diep, A.N.; Fleiss, R.; Sudilovsky, D.; Anguish, L.J.; Coonrod, S.A.; Wakshlag, J.J. Comparative analysis of peptidylarginine deiminase-2 expression in canine, feline and human mammary tumours. J. Comp. Pathol. 2012, 147, 139–146. [Google Scholar] [CrossRef]
- Xin, J.; Song, X. Role of peptidylarginine deiminase type 4 in gastric cancer. Exp. Ther. Med. 2016, 12, 3155–3160. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Song, G.; Zhang, X.; Feng, T.; Pan, J.; Chen, W.; Yang, M.; Bai, X.; Pang, Y.; Yu, J.; et al. PADI2-mediated citrullination promotes prostate cancer progression. Cancer Res. 2017, 77, 5755–5768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulivi, P.; Mercatali, L.; Casoni, G.L.; Scarpi, E.; Bucchi, L.; Silvestrini, R.; Sanna, S.; Monteverde, M.; Amadori, D.; Poletti, V.; et al. Multiple marker detection in peripheral blood for NSCLC diagnosis. PLoS ONE 2013, 8, e57401. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chang, X.; Yuan, G.; Zhao, Y.; Wang, P. Expression of peptidylarginine deiminase type 4 in ovarian tumors. Int. J. Biol. Sci. 2010, 6, 454–464. [Google Scholar] [CrossRef]
- Li, G.; Young, C.H.; Snow, B.; Christensen, A.O.; Demoruelle, M.K.; Nemmara, V.V.; Thompson, P.R.; Rothfuss, H.M.; Cherrington, B.D. Identification and characterization of the lactating mouse mammary gland citrullinome. Int. J. Mol. Sci. 2020, 21, 2634. [Google Scholar] [CrossRef] [Green Version]
- Beato, M.; Sharma, P. Peptidyl arginine deiminase 2 (PADI2)-mediated arginine citrullination modulates transcription in cancer. Int. J. Mol. Sci. 2020, 21, 1351. [Google Scholar] [CrossRef] [Green Version]
- Stacey, S.N.; Gudbjartsson, D.F.; Sulem, P.; Bergthorsson, J.T.; Kumar, R.; Thorleifsson, G.; Sigurdsson, A.; Jakobsdottir, M.; Sigurgeirsson, B.; Benediktsdottir, K.R.; et al. Common variants on 1p36 and 1q42 are associated with cutaneous basal cell carcinoma but not with melanoma or pigmentation traits. Nat. Genet. 2008, 40, 1313–1318. [Google Scholar] [CrossRef] [PubMed]
- van Beers, J.J.; Zendman, A.J.; Raijmakers, R.; Stammen-Vogelzangs, J.; Pruijn, G.J. Peptidylarginine deiminase expression and activity in PAD2 knock-out and PAD4-low mice. Biochimie 2013, 95, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Witalison, E.E.; Thompson, P.R.; Hofseth, L.J. Protein arginine deiminases and associated citrullination: Physiological functions and diseases associated with dysregulation. Curr. Drug Targets 2015, 16, 700–710. [Google Scholar] [CrossRef]
- Zhu, D.; Zhang, Y.; Wang, S. Histone citrullination: A new target for tumors. Mol. Cancer 2021, 20, 90. [Google Scholar] [CrossRef] [PubMed]
- Albrengues, J.; Shields, M.A.; Ng, D.; Park, C.G.; Ambrico, A.; Poindexter, M.E.; Upadhyay, P.; Uyeminami, D.L.; Pommier, A.; Küttner, V.; et al. Neutrophil extracellular traps produced during inflammation awaken dormant cancer cells in mice. Science 2018, 361, eaao4227. [Google Scholar] [CrossRef] [Green Version]
- Berger-Achituv, S.; Brinkmann, V.; Abed, U.A.; Kühn, L.I.; Ben-Ezra, J.; Elhasid, R.; Zychlinsky, A. A proposed role for neutrophil extracellular traps in cancer immunoediting. Front. Immunol. 2013, 4, 48. [Google Scholar] [CrossRef] [Green Version]
- Tohme, S.; Yazdani, H.O.; Al-Khafaji, A.B.; Chidi, A.P.; Loughran, P.; Mowen, K.; Wang, Y.; Simmons, R.L.; Huang, H.; Tsung, A. Neutrophil extracellular traps promote the development and progression of liver metastases after surgical stress. Cancer Res. 2016, 76, 1367–1380. [Google Scholar] [CrossRef] [Green Version]
- Cools-Lartigue, J.; Spicer, J.; McDonald, B.; Gowing, S.; Chow, S.; Giannias, B.; Bourdeau, F.; Kubes, P.; Ferri, L. Neutrophil extracellular traps sequester circulating tumor cells and promote metastasis. J. Clin. Investig. 2013, 123, 3446–3458. [Google Scholar] [CrossRef]
- Magnadóttir, B.; Bragason, B.T.; Bricknell, I.R.; Bowden, T.; Nicholas, A.P.; Hristova, M.; Guðmundsdóttir, S.; Dodds, A.W.; Lange, S. Peptidylarginine deiminase and deiminated proteins are detected throughout early halibut ontogeny—Complement components C3 and C4 are post-translationally deiminated in halibut (Hippoglossus hippoglossus L.). Dev. Comp. Immunol. 2019, 92, 1–19. [Google Scholar] [CrossRef]
| Animal (Species Name) | Carcinoma Type | Sample Type |
|---|---|---|
| Rabbit (Oryctolagus cuniculus) | Mammary adenocarcinoma | Biopsy from the mammary gland of a four-year old female rabbit/doe. |
| Dog (Canis lupus familiaris) | (1) Mixed germ cell-sex cord stromal tumour, testicle; (2) Squamous cell tumour; origin of tissue unknown | Biopsies from seven-year old dog, male: (1) mixed tumour in testicle (based on morphological assessment); (2) Bladder. |
| Regent parrot (Polytelis anthopeplus) | Lymphoma, liver and spleen | Cadaver, male bird. Tumour in various organs, including liver, spleen, heart, kidney, adipose tissue and skin. Diagnosis: disseminated malignant round cell tumour, probable lymphoma. |
| Domestic duck (Anas platyrhynchos domesticus) | Malignant round cell tumour, liver; probably lymphoma | Organ samples. Tumour, liver; probably lymphoma—not verified by IHC; tumour cells only seen in the liver |
| Mink (Neovision vision) | Malignant renal tumour, possibly nephroblastoma | Cadaver, adult mink, male. Tumour in kidney with metastasis to lung and liver. |
| Cat (Felis catus) | Pulmonary carcinoma | Cadaver of 13-year old male cat. Primary lung tumour suspected as no other primary tumour found. Metastasis to heart. |
| Horse (Equus caballus) | Carcinoma in liver (ovary and mesenteric lymph node) | Organs from an 18-year old mare. Carcinoma in liver; probable primary bile duct carcinoma but tumour metastasis to the liver could not be ruled out. |
| Cattle (Bos Taurus) | Neuroendocrine tumour (metastasis to the heart) | Heart with tumour located on the surface of the right ventricle. Origin of the tumour uncertain (can be from chemoreceptors at the carotid or aortic body, from C-cells in the thyroid gland (ultimobranchial tumour or adrenal gland medulla (pheochromocytoma—neuroendocrine tumour). |
| Dog (Canis lupus familiaris) | Anaplastic mammary carcinoma | Biopsy from a five-year old bitch. |
| Sheep (Ovis aries) | Malignant round cell tumour, skin | Two-year old ewe. Skin with tumour was received; origin of this highly malignant tumour is unknown. |
| Hamster (Mesocricetus auratus) | Lymphoma, mesentery/small intestine | Cadaver of a hamster with tumour masses in mesentery. Multicentric lymphoma caused by Hamster Polyomavirus (HaPV). |
| Dog (Canis lupus familiaris) | Scirrhous adenocarcinoma, mammary gland | Biopsy from tumour in mammary gland of an eight-year old bitch |
| Horse (Equus caballus) | Squamous cell carcinoma, penile. | Samples taken from penile tumour. |
| Cattle (Bos taurus) | Metastatic carcinoma, lungs and lymph nodes | Organ samples with tumour in lungs and lymph nodes; origin of the primary tumour unknown |
| Cat (Felis catus) | Malignant tumour, kidneys; probably lymphoma | Kidneys from an eight-month old male cat. Tumour in both kidneys—probably lymphoma. |
| Dog (Canis lupus familiaris) | Granulosa cell tumour, ovary | Samples from ovary of a five-year-old bitch. |
| Cattle (Bos Taurus) | Lymphoma, lymph node | Organ samples from a cow, including kidney and lymph nodes. |
| Mink (Neovision vision) | Malignant renal tumour, probably nephroblastoma | Cadaver of a male mink. Extensive necrosis and haemorrhage in tumour mass. |
| Reindeer (Rangifer tarandus) | Multicentric malignant tumour, probably mesothelioma | Organ samples from reindeer, female. Tumour masses in the abdominal cavity. Spleen and diaphragm with tumour. |
| Pig (Sus scrofa domesticus) | Lymphoma (lymph node) | Organ sample from a slaughtered pig. Lymphoma in lymph node and kidney. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inal, J.M.; Hristova, M.; Lange, S. A Pilot Study on Peptidylarginine Deiminases and Protein Deimination in Animal Cancers across Vertebrate Species. Int. J. Mol. Sci. 2022, 23, 8697. https://doi.org/10.3390/ijms23158697
Inal JM, Hristova M, Lange S. A Pilot Study on Peptidylarginine Deiminases and Protein Deimination in Animal Cancers across Vertebrate Species. International Journal of Molecular Sciences. 2022; 23(15):8697. https://doi.org/10.3390/ijms23158697
Chicago/Turabian StyleInal, Jameel M., Mariya Hristova, and Sigrun Lange. 2022. "A Pilot Study on Peptidylarginine Deiminases and Protein Deimination in Animal Cancers across Vertebrate Species" International Journal of Molecular Sciences 23, no. 15: 8697. https://doi.org/10.3390/ijms23158697
APA StyleInal, J. M., Hristova, M., & Lange, S. (2022). A Pilot Study on Peptidylarginine Deiminases and Protein Deimination in Animal Cancers across Vertebrate Species. International Journal of Molecular Sciences, 23(15), 8697. https://doi.org/10.3390/ijms23158697