
Fluorescent Auxin Analogs Report Two Auxin Binding Sites with Different Subcellular Distribution and Affinities: A Cue for Non-Transcriptional Auxin Signaling


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
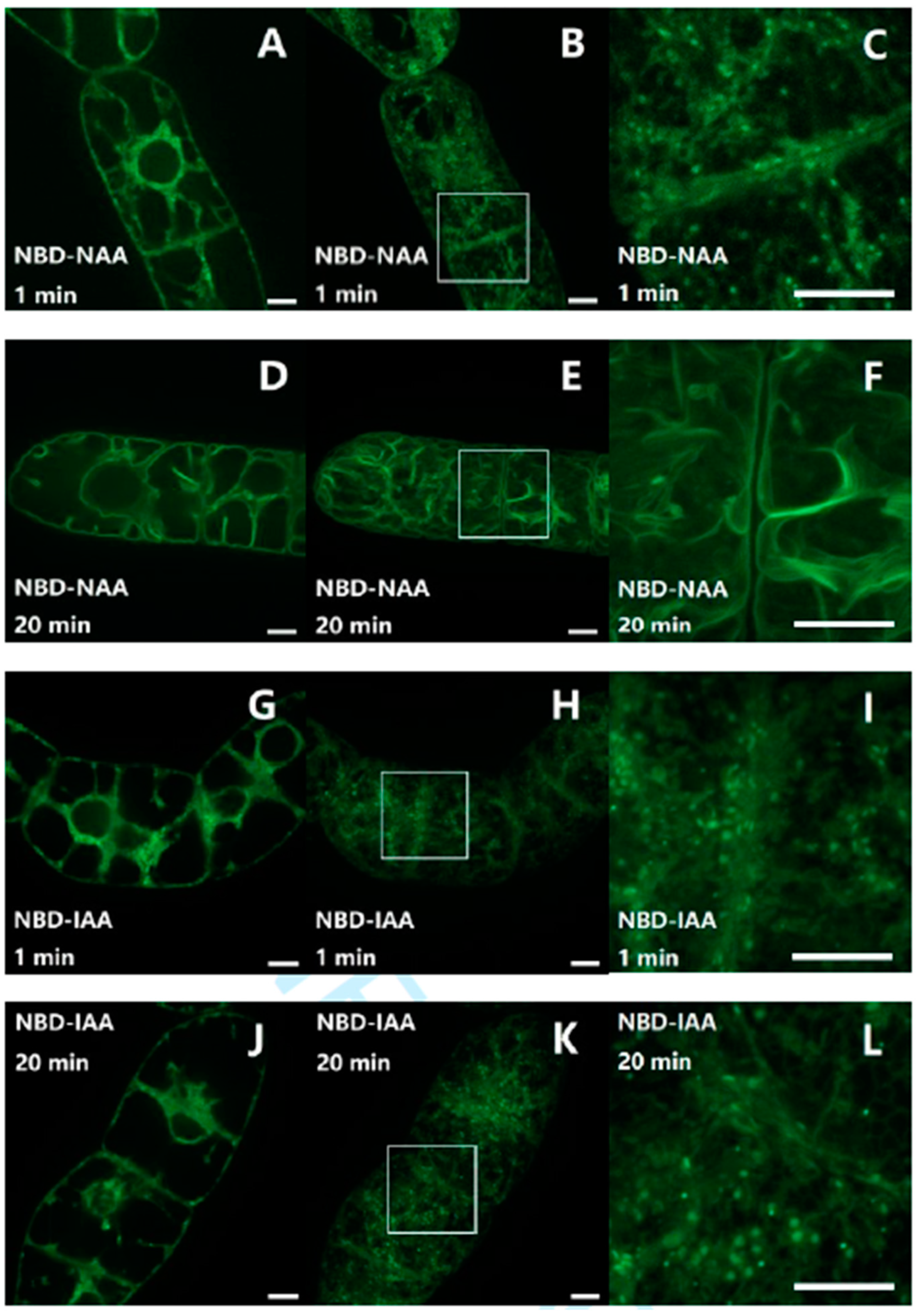
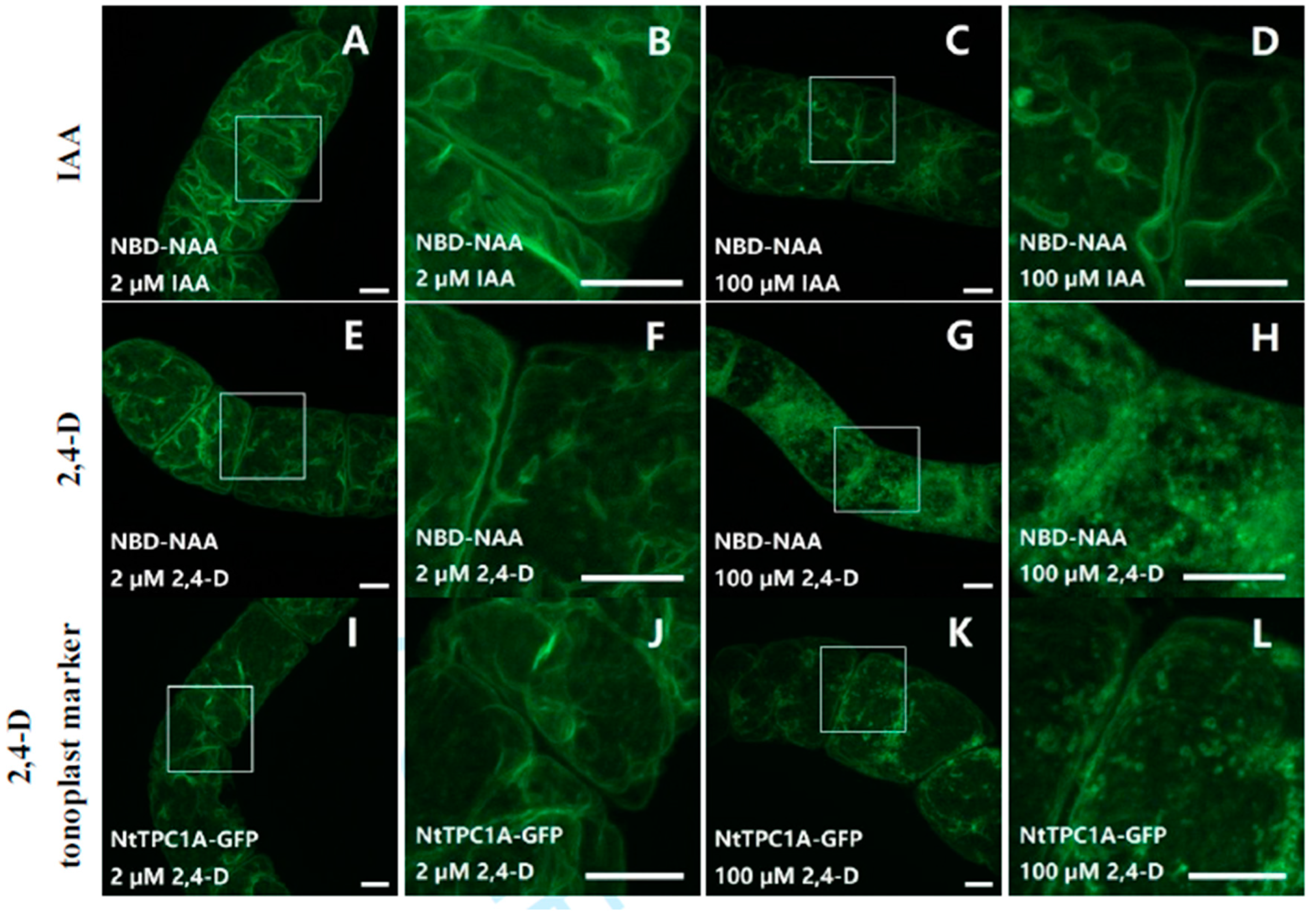
2.1. The Fluorescent Auxin-Analogs NBD-NAA and NBD-IAA Localize Differently
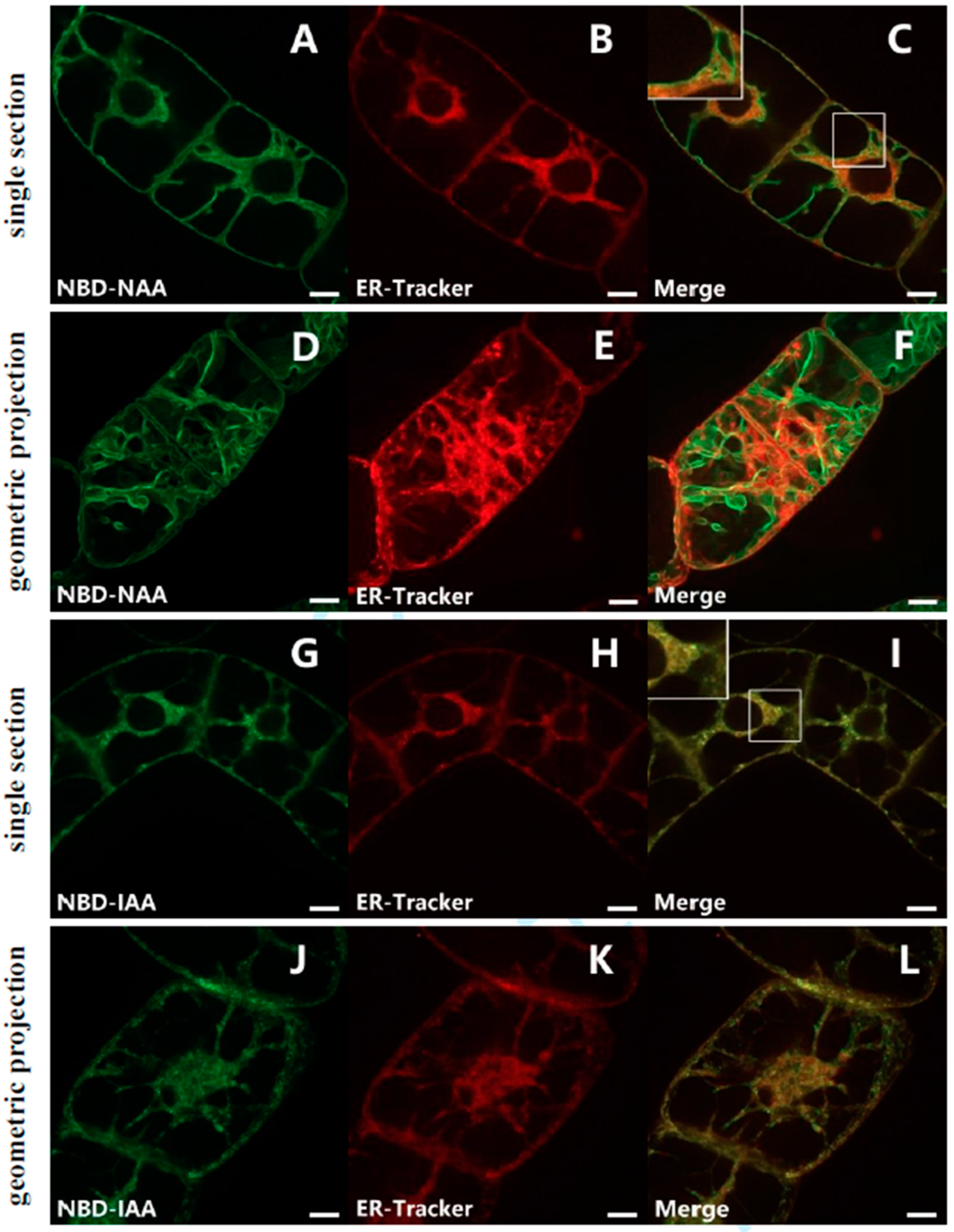
2.2. NBD-NAA Links to Endoplasmic Reticulum (ER) and Tonoplast, NBD-IAA to the ER
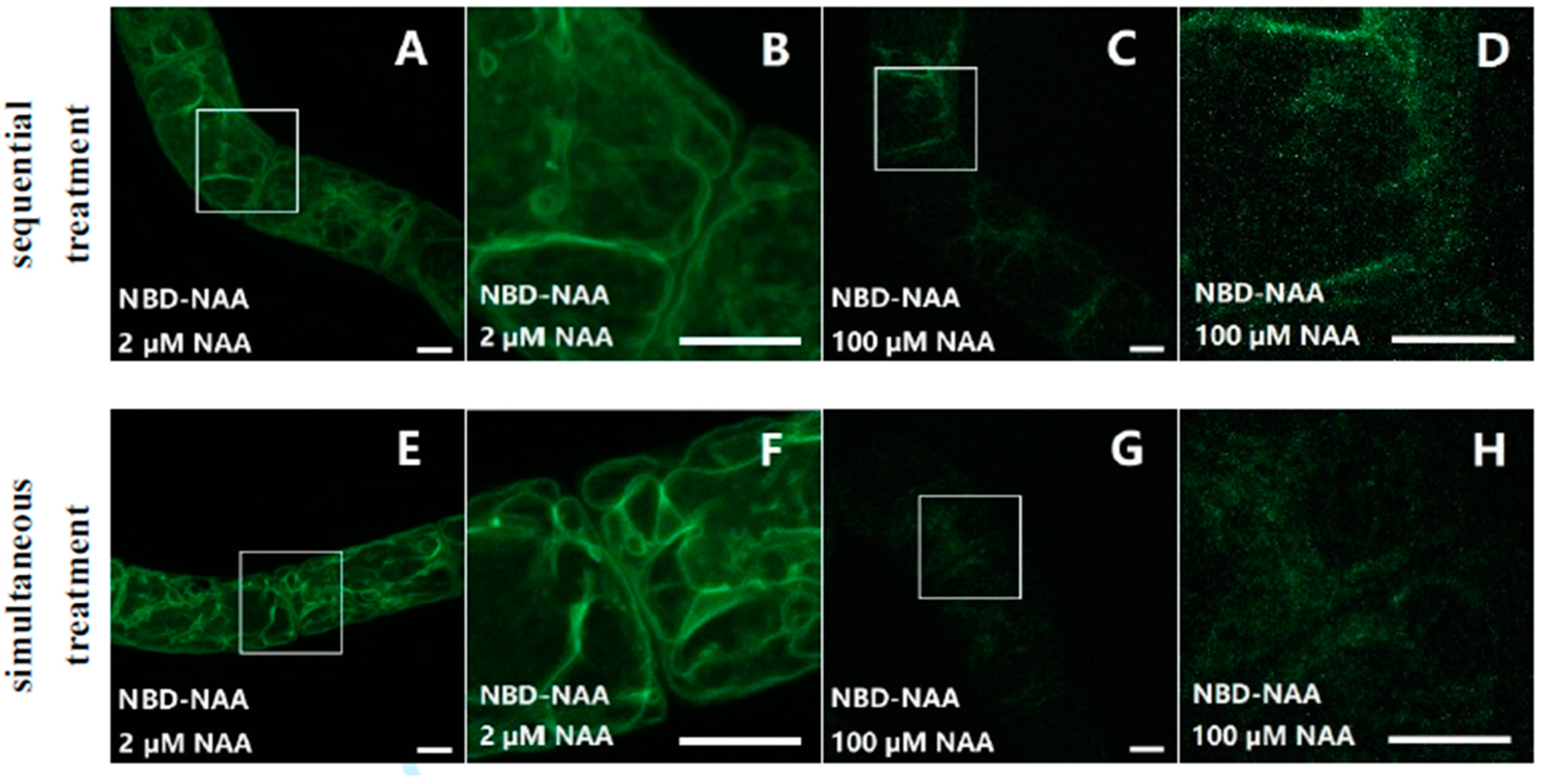
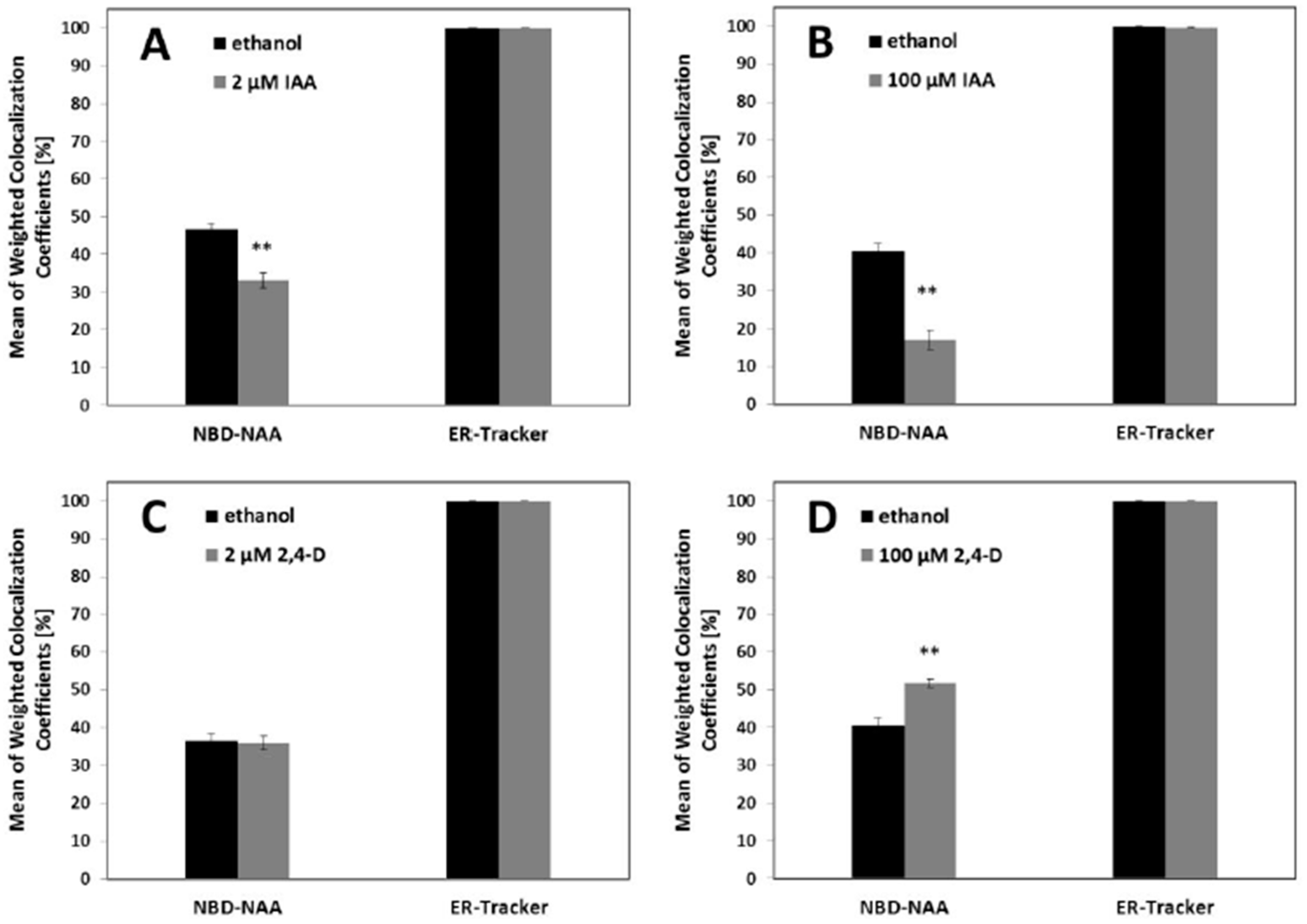
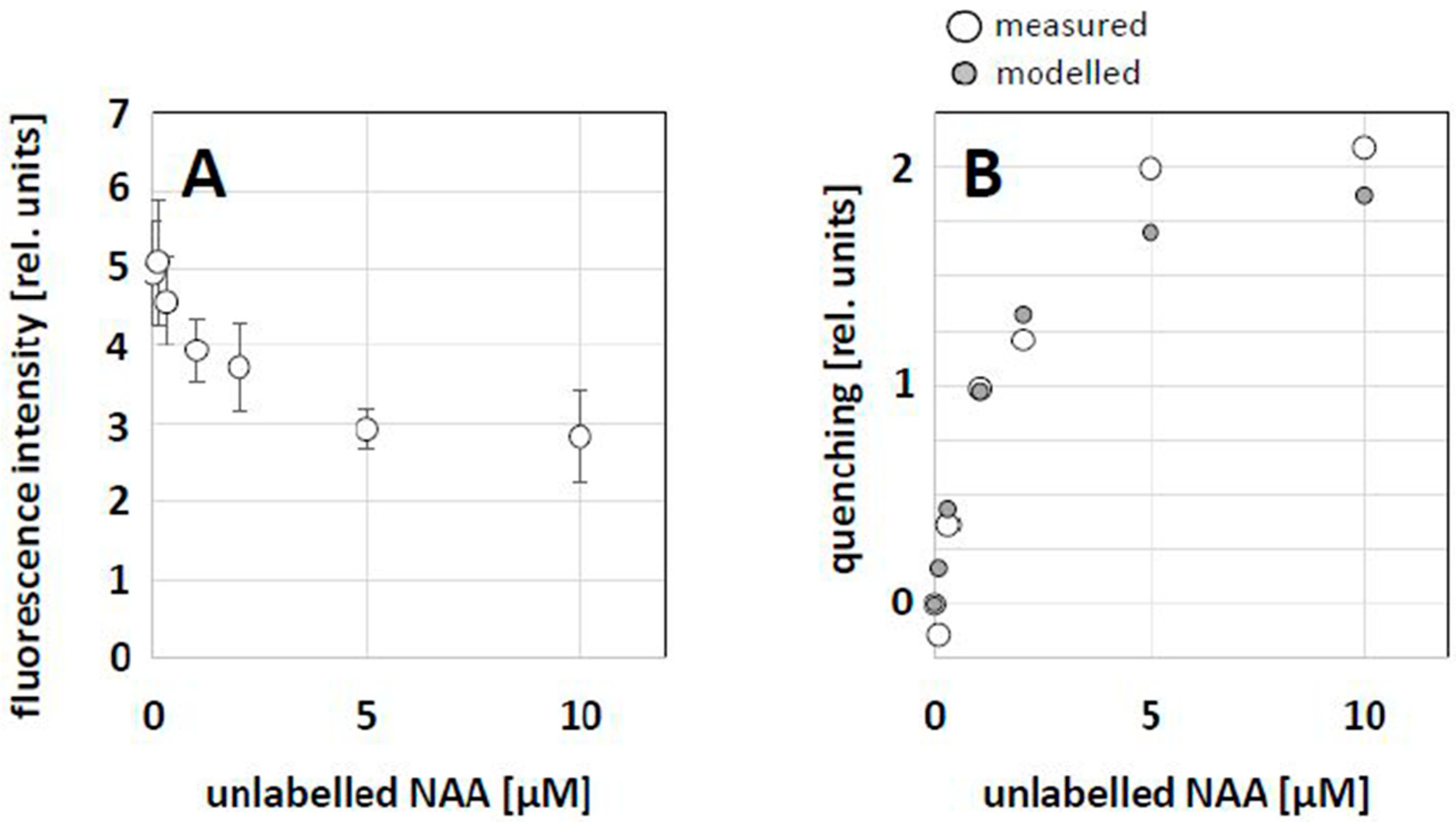
2.3. NAA Can Compete with NAA-NBD for the Same Binding Sites
2.4. Quantification of the Competition between NBD-NAA and Unlabeled NAA
3. Discussion
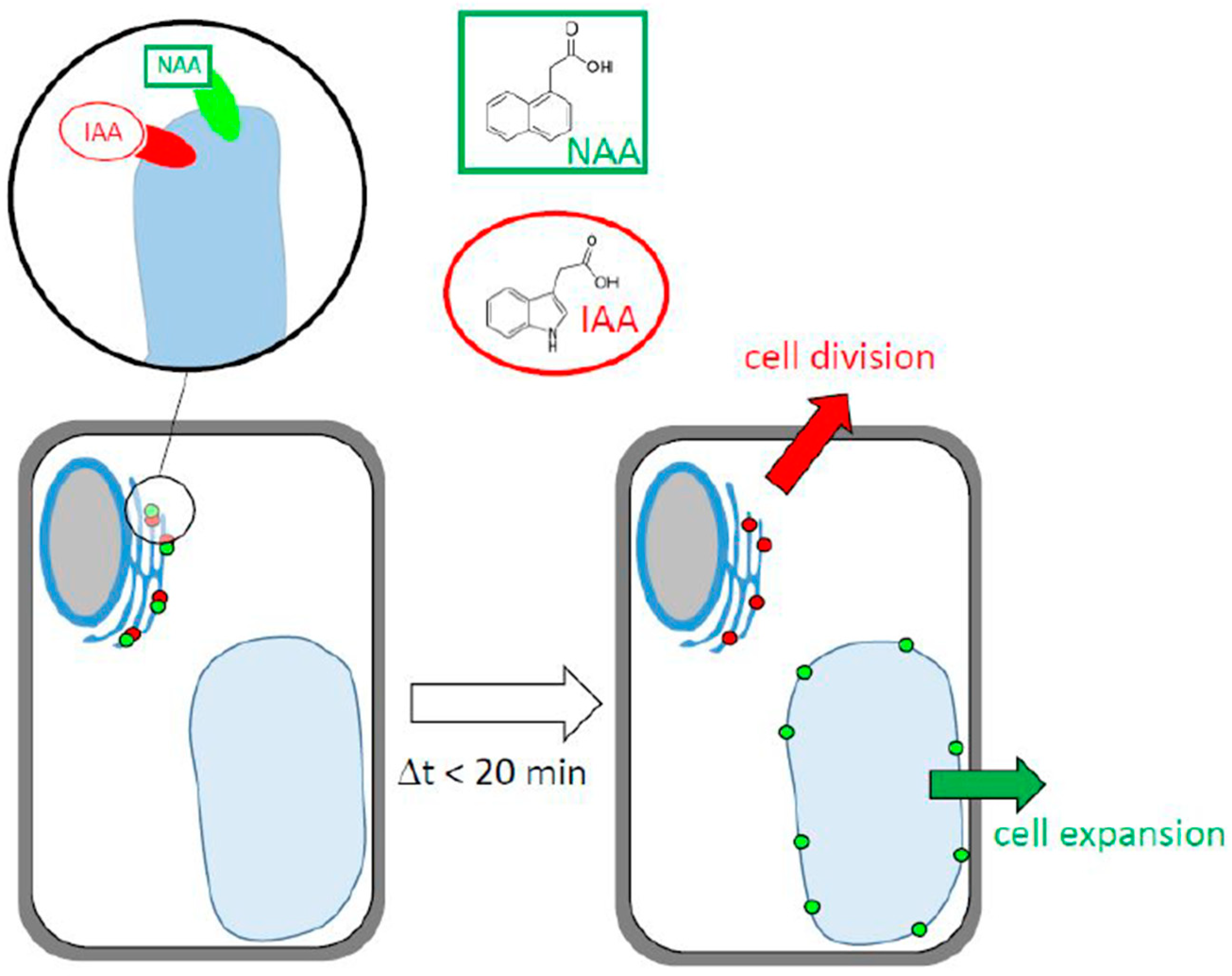
3.1. Do Fluorescent Auxin Analogs Report the Subcellular Distribution of Auxin?
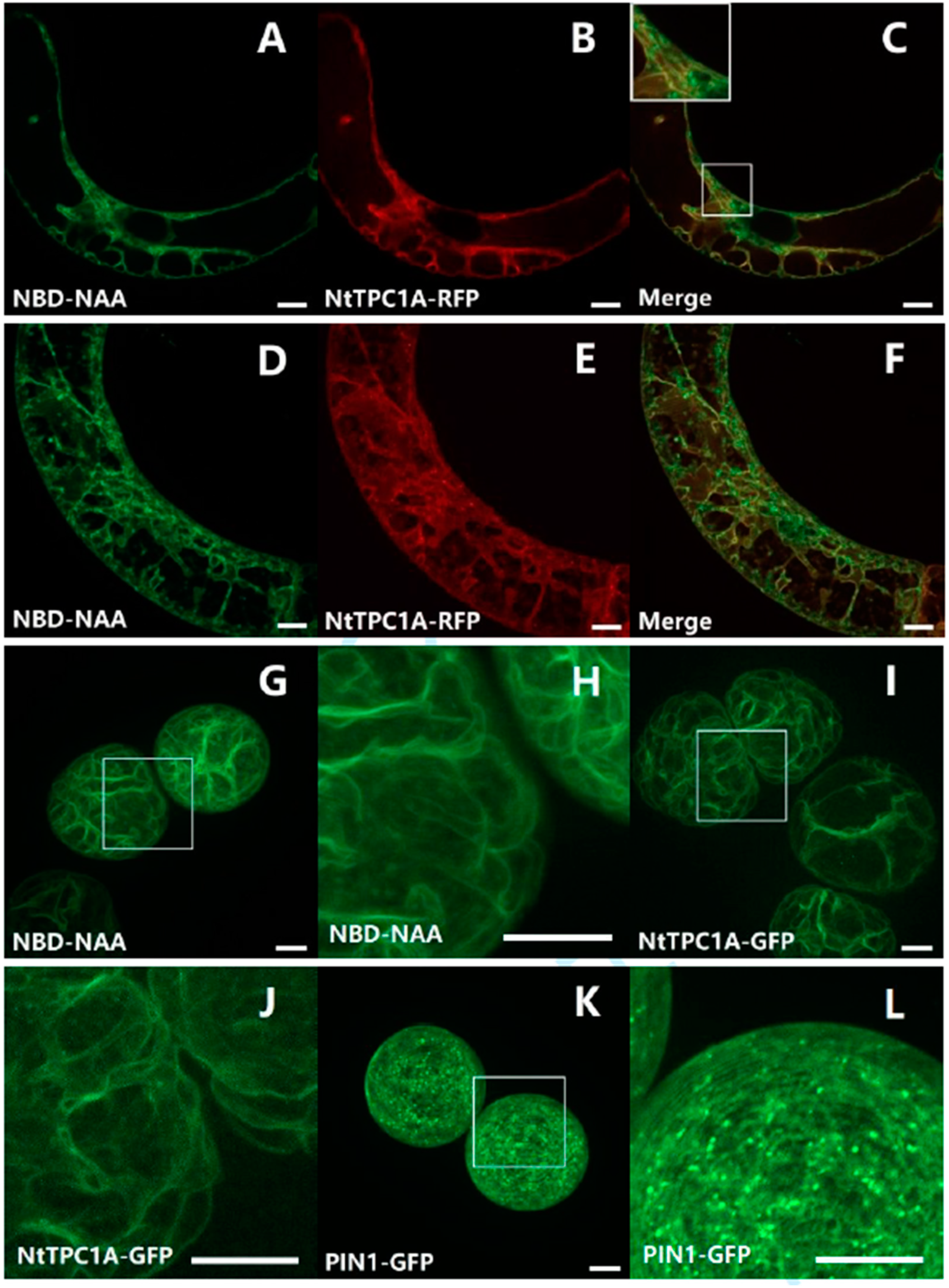
3.2. Two Perceptive Events, Two Types of Auxin Transporters?
4. Materials and Methods
4.1. Tobacco Cell Cultivation
4.2. Fluorescent Auxin Analogs
4.3. Treatment with Fluorescent Auxin Analogs
4.4. Generation of Protoplasts
4.5. Acquisition of Microscopical Images
4.6. Co-Localization Analysis
4.7. Quantitative Image Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darwin, C.; Darwin, F. Sensitiveness of plants to light: It’s transmitted effect. In The Power of Movement in Plants; Murray: London, UK, 1880; pp. 574–592. [Google Scholar]
- Went, F. On growth accelerating substances in the coleoptile of Avena sativa. Proc. Kon. Ned. Akad. Weten. 1926, 30, 10–19. [Google Scholar]
- Cholodny, N. Wuchshormone und Tropismen bei den Pflanzen. Biol. Zent. 1927, 47, 604–626. [Google Scholar]
- Thimann, K.V. On the plant growth hormone produced by Rhizopus suinus. J. Biol. Chem. 1935, 109, 279–291. [Google Scholar] [CrossRef]
- Kögl, F.; Hagen-Smit, J.; Erxleben, H. Über ein neues Auxin (“Hetero-Auxin”) aus Harn. Hoppe-Seylers Z Physiol. Chem. 1934, 228, 104–112. [Google Scholar] [CrossRef]
- Korasick, D.A.; Enders, T.A.; Strader, L.C. Auxin biosynthesis and storage forms. J. Exp. Bot. 2013, 64, 2541–2555. [Google Scholar] [CrossRef] [Green Version]
- Enders, T.A.; Strader, L.C. Auxin activity: Past, present, and future. Am. J. Bot. 2015, 102, 180–196. [Google Scholar] [CrossRef] [Green Version]
- Ljung, K.; Hull, A.K.; Kowalczyk, M.; Marchant, A.; Celenza, J.; Cohen, J.D.; Sandberg, G. Biosynthesis, conjugation, catabolism and homeostasis of indole-3-acetic acid in Arabidopsis thaliana. Plant Mol. Biol. 2002, 49, 249–272. [Google Scholar] [CrossRef] [PubMed]
- Carrier, D.J.; Abu Bakar, N.T.; Swarup, R.; Callaghan, R.; Napier, R.M.; Bennett, M.J.; Kerr, I.D. The binding of auxin to the Arabidopsis auxin influx transporter AUX1. Plant Physiol. 2008, 148, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Benková, E.; Michniewicz, M.; Sauer, M.; Teichmann, T.; Seifertová, D.; Jürgens, G.; Friml, J. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 2003, 115, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Brumos, J.; Robles, L.M.; Yun, J.; Vu, T.C.; Jackson, S.; Alonso, J.M.; Stepanova, A.N. Local auxin biosynthesis is a key regulator of plant development. Dev. Cell 2018, 47, 306–318. [Google Scholar] [CrossRef] [Green Version]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubery, P.H.; Sheldrake, A.R. Carrier-mediated auxin transport. Planta 1974, 118, 101–121. [Google Scholar] [CrossRef] [PubMed]
- Friml, J. Subcellular trafficking of PIN auxin efflux carriers in auxin transport. Eur. J. Cell Biol. 2010, 89, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Carraro, N.; Tisdale-Orr, T.E.; Clouse, R.M.; Knöller, A.S.; Spicer, R. Diversification and expression of the PIN, AUX/LAX, and ABCB families of putative auxin transporters in populus. Front. Plant Sci. 2012, 3, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisniewská, J.; Xu, J.; Seifertová, D.; Brewer, P.B.; Růžička, K.; Blilou, I.; Rouquie, D.; Benková, E.; Scheres, B.; Friml, J. Polar PIN localization directs auxin flow in plants. Science 2006, 312, 883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grieneisen, V.A.; Xu, J.; Marée, A.F.; Hogeweg, P.; Scheres, B. Auxin transport is sufficient to generate a maximum and gradient guiding root growth. Nature 2007, 449, 1008–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, H.S.; Friml, J. Auxin and other signals on the move in plants. Nat. Chem. Biol. 2009, 5, 325–332. [Google Scholar] [CrossRef]
- Bennett, M.J.; Marchant, A.; Green, H.G.; May, S.T.; Ward, S.P.; Millner, P.A.; Walker, A.R.; Schulz, B.; Feldmann, K.A. Arabidopsis AUX1 gene: A permease-like regulator of root gravitropism. Science 1996, 273, 948–950. [Google Scholar] [CrossRef]
- Péret, B.; Swarup, K.; Ferguson, A.; Seth, M.; Yang, Y.D.; Dhondt, S.; James, N.; Casimiro, I.; Perry, P.; Syed, A.; et al. AUX/LAX genes encode a family of auxin influx transporters that perform distinct functions during Arabidopsis development. Plant Cell 2012, 24, 2874–2885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarup, R.; Friml, J.; Marchant, A.; Ljung, K.; Sandberg, G.; Palme, K.; Bennett, M. Localization of the auxin permease AUX1 suggests two functionally distinct hormone transport pathways operate in the Arabidopsis root apex. Genes Dev. 2001, 15, 2648–2653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nick, P. Auxin and self organisation. In Auxin and Its Role in Plant Development; Zažímalová, E., Petrášek, J., Benková, E., Eds.; Springer: Heidelberg, Germany, 2014; pp. 291–313. [Google Scholar]
- Wang, R.; Estelle, M. Diversity and specificity: Auxin perception and signaling through the TIR1/AFB pathway. Curr. Opin. Plant Biol. 2014, 21, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.; Chen, R.; Li, P.; Yu, Y.Q.; Zheng, R.; Ge, D.F.; Zheng, W.; Wang, X.H.; Gu, Y.T.; Gelová, Z.; et al. TMK1-mediated auxin signalling regulates differential growth of the apical hook. Nature 2019, 568, 240–243. [Google Scholar] [CrossRef]
- Scherer, G.F.E. AUXIN-BINDING-PROTEIN1, the second auxin receptor: What is the significance of a two-receptor concept in plant signal transduction? J. Exp. Bot. 2011, 62, 3339–3357. [Google Scholar] [CrossRef] [Green Version]
- Monshausen, G.B.; Miller, N.D.; Murphy, A.S.; Gilroy, S. Dynamics of auxin-dependent Ca2+ and pH signaling in root growth revealed by integrating high-resolution imaging with automated computer vision-based analysis. Plant J. 2011, 65, 309–318. [Google Scholar] [CrossRef]
- Napier, R. Towards an understanding of ABP1. J. Exp. Bot. 1995, 46, 1787–1795. [Google Scholar] [CrossRef]
- Bargmann, B.O.R.; Estelle, M. Auxin perception: In the IAA of the beholder. Physiol. Plant 2014, 151, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.B.; Zhang, Y.; Zhang, D.; Dai, X.H.; Estelle, M.; Zhao, Y.D. Auxin binding protein 1 (ABP1) is not required for either auxin signaling or Arabidopsis development. Proc. Natl. Acad. Sci. USA 2015, 112, 2275–2280. [Google Scholar] [CrossRef] [Green Version]
- Dahlke, R.I.; Fraas, S.; Ullrich, K.K.; Heinemann, K.; Romeiks, M.; Rickmeyer, T.; Klebe, G.; Palme, K.; Lüthen, H.; Steffens, B. Protoplast Swelling and Hypocotyl Growth Depend on Different Auxin Signaling Pathways. Plant Physiol. 2017, 175, 982–994. [Google Scholar] [CrossRef]
- Campanoni, P.; Nick, P. Auxin-dependent cell division and cell elongation: NAA and 2,4-D activate different pathways. Plant Physiol. 2005, 137, 939–948. [Google Scholar] [CrossRef] [Green Version]
- Friml, J.; Vieten, A.; Sauer, M.; Weijers, D.; Schwarz, H.; Hamann, T.; Offringa, R.; Jürgens, G. Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 2003, 426, 147–153. [Google Scholar] [CrossRef]
- Heisler, M.G.; Ohno, C.; Das, P.; Sieber, P.; Reddy, G.V.; Long, J.A.; Meyerowitz, E.M. Patterns of auxin transport and gene expression during primordium development revealed by live imaging of the Arabidopsis inflorescence meristem. Curr. Biol. 2005, 15, 1899–1911. [Google Scholar] [CrossRef] [Green Version]
- Sauer, M.; Balla, J.; Luschnig, C.; Wisniewská, J.; Reinöhl, V.; Friml, J.; Benková, E. Canalization of auxin flow by Aux/IAA-ARF-dependent feedback regulation of PIN polarity. Genes Dev. 2006, 20, 2902–2911. [Google Scholar] [CrossRef] [Green Version]
- Krouk, G.; Lacombe, B.; Bielach, A.; Perrine-Walker, F.; Malinská, K.; Mounier, E.; Hoyerová, K.; Tillard, P.; Leon, S.; Ljung, K.; et al. Nitrate-Regulated Auxin Transport by NRT1.1 Defines a Mechanism for Nutrient Sensing in Plants. Dev. Cell 2010, 18, 927–937. [Google Scholar] [CrossRef]
- Liu, X.; Hegeman, A.D.; Gardner, G.; Cohen, J.D. Protocol: High-throughput and quantitative assays of auxin and auxin precursors from minute tissue samples. Plant Methods 2012, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Novák, O.; Hényková, E.; Sairanen, I.; Kowalczyk, M.; Pospíšil, T.; Ljung, K. Tissue-specific profiling of the Arabidopsis thaliana auxin metabolome. Plant J. 2012, 72, 523–536. [Google Scholar] [CrossRef]
- Herud-Sikimić, O.; Stiel, A.C.; Kolb, M.; Shanmugaratnam, S.; Berendzen, K.W.; Feldhaus, C.; Höcker, B.; Jürgens, G. A biosensor for the direct visualization of auxin. Nature 2021, 592, 768–772. [Google Scholar] [CrossRef]
- Jones, A.M. Location of transported auxin in etiolated maize shoots using 5-azidoindole-3-acetic Acid. Plant Physiol. 1990, 93, 1154–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, K.I.; Nakamura, S.I.; Fukunaga, S.; Nishimura, T.; Jenness, M.K.; Murphy, A.S.; Motose, H.; Nozaki, H.; Furutani, M.; Aoyama, T. Auxin transport sites are visualized in planta using fluorescent auxin analogs. Proc. Natl. Acad. Sci. USA 2014, 111, 11557–11562. [Google Scholar] [CrossRef] [Green Version]
- Kadota, Y.; Furuichi, T.; Ogasawara, Y.; Goh, T.; Higashi, K.; Muto, S.; Kuchitsu, K. Identification of putative voltage-dependent Ca2+-permeable channels involved in cryptogein-induced Ca2+ transients and defense responses in tobacco BY-2 cells. Biochem. Biophys. Res. Commun. 2004, 317, 823–830. [Google Scholar] [CrossRef]
- Kurusu, T.; Nishikawa, D.; Yamazaki, Y.; Gotoh, M.; Nakano, M.; Hamada, H.; Yamanaka, T.; Iida, K.; Nakagawa, Y.; Saji, H.; et al. Plasma membrane protein OsMCA1 is involved in regulation of hypo-osmotic shock-induced Ca2+ influx and modulates generation of reactive oxygen species in cultured rice cells. BMC Plant Biol. 2012, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Heisenberg, W. Über den anschaulichen Inhalt der quantentheoretischen Kinematik und Mechanik. Zeitschr. Phys. 1927, 43, 172–198. [Google Scholar] [CrossRef]
- Chen, J.G. Dual auxin signalling pathways control cell elongation and division. J. Plant Growth Regul. 2001, 20, 255–264. [Google Scholar] [CrossRef]
- Ullah, H.; Chen, J.G.; Young, J.C.; Im, K.H.; Sussman, R.M.; Jones, A.M. Modulation of cell proliferation by heterotrimeric G protein in Arabidopsis. Science 2001, 292, 2066–2069. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Hertel, R.; Ishikawa, H.; Evans, M.L. Species differences in ligand specificity of auxin-controlled elongation and auxin transport: Comparing Zea and Vigna. Planta 2002, 216, 293–301. [Google Scholar] [CrossRef]
- Hössel, D.; Schmeiser, C.; Hertel, R. Specificity Patterns Indicate that Auxin Exporters and Receptors are the Same Proteins. Plant Biol. 2005, 7, 41–48. [Google Scholar] [CrossRef]
- Hennig, W. Phylogenetische Systematik; Parey: Berlin, Germany, 1982. [Google Scholar]
- Geldner, N.; Friml, J.; Stierhof, Y.D.; Jürgens, G.; Palme, K. Auxin transport inhibitors block PIN1 cycling and vesicle trafficking. Nature 2001, 413, 425–428. [Google Scholar] [CrossRef]
- Lam, S.K.; Cai, Y.; Tse, Y.C.; Wang, J.; Law, A.H.Y.; Pimpl, P.; Chan, H.Y.E.; Xia, J.; Jiang, L.W. BFA-induced compartments from the Golgi apparatus and trans-Golgi network/early endosome are distinct in plant cells. Plant J. 2009, 60, 865–881. [Google Scholar] [CrossRef]
- Mravec, J.; Skůpa, P.; Bailly, A.; Hoyerová, K.; Krecek, P.; Bielach, A.; Petrásek, J.; Zhang, J.; Gaykova, V.; Stierhof, Y.D.; et al. Subcellular homeostasis of phytohormone auxin is mediated by the ER-localized PIN5 transporter. Nature 2009, 459, 1136–1140. [Google Scholar] [CrossRef] [PubMed]
- Viotti, C.; Falco, K.; Krebs, M.; Neubert, C.; Fink, F.; Lupanga, U.; Scheuring, D.; Boutté, Y.; Frescatada-Rosa, M.; Wolfenstetter, S.; et al. The endoplasmic reticulum is the main membrane source for biogenesis of the lytic vacuole in Arabidopsis. Plant Cell 2013, 25, 3434–3449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedrazzini, E.; Komarova, N.Y.; Rentsch, D.; Vitale, A. Traffic routes and signals for the tonoplast. Traffic 2013, 14, 622–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viotti, C. ER and vacuoles: Never been closer. Front. Plant Sci. 2014, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Maisch, J.; Nick, P. Actin is involved in auxin-dependent patterning. Plant Physiol. 2007, 143, 1695–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, M.X.; Ren, H.Y.; Li, J.J. An Auxin Transport Inhibitor Targets Villin-Mediated Actin Dynamics to Regulate Polar Auxin Transport. Plant Physiol. 2019, 181, 161–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.; Calderon-Villalobos, L.I.A.; Sharon, M.; Zheng, C.X.; Robinson, C.V.; Estelle, M.; Zheng, N. Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature 2007, 446, 640–645. [Google Scholar] [CrossRef]
- Yang, H.; Murphy, A.S. Functional expression and characterization of Arabidopsis ABCB, AUX 1 and PIN auxin transporters in Schizosaccharomyces pombe. Plant J. 2009, 59, 179–191. [Google Scholar] [CrossRef]
- Petrášek, J.; Mravec, J.; Bouchard, R.; Blakeslee, J.J.; Abas, M.; Seifertová, D.; Wiśniewska, J.; Tadele, Z.; Kubeš, M.; Čovanová, M.; et al. PIN proteins perform a rate-limiting function in cellular auxin efflux. Science 2006, 312, 914–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakeslee, J.J.; Bandyopadhyay, A.; Lee, O.R.; Mravec, J.; Titapiwatanakun, B.; Sauer, M.; Makam, S.N.; Cheng, Y.; Bouchard, R.; Adamec, J.; et al. Interactions among PIN-FORMED and P-glycoprotein auxin transporters in Arabidopsis. Plant Cell 2007, 19, 131–147. [Google Scholar] [CrossRef] [Green Version]
- Sokołowska, K.; Kizińska, J.; Szewczuk, Z.; Banasiak, A. Auxin conjugated to fluorescent dyes—A tool for the analysis of auxin transport pathways. Plant Biol. 2014, 16, 866–877. [Google Scholar] [CrossRef]
- Ganguly, A.; Lee, S.H.; Cho, M.; Lee, O.R.; Yoo, H.; Cho, H.T. Differential auxin-transporting activities of PIN-FORMED proteins in Arabidopsis root hair cells. Plant Physiol. 2010, 153, 1046–1061. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Dhonukshe, P.; Brewer, P.B.; Friml, J. Spatiotemporal asymmetric auxin distribution: A means to coordinate plant development. Cell Mol. Life Sci. 2006, 63, 2738–2754. [Google Scholar] [CrossRef]
- Zažímalová, E.; Křeček, P.; Skůpa, P.; Hoyerová, K.; Petrášek, J. Polar transport of the plant hormone auxin—The role of PIN-FORMED (PIN) proteins. Cell Mol. Life Sci. 2007, 64, 1621–1637. [Google Scholar] [CrossRef] [PubMed]
- Leyser, O. Auxin signaling. Plant Physiol. 2018, 76, 465–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robatzek, S.; Chinchilla, D.; Boller, T. Ligand-induced endocytosis of the pattern recognition receptor FLS2 in Arabidopsis. Genes Dev. 2006, 20, 537–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, T.; Nemoto, Y.; Hasezawa, S. Tobacco BY-2 cell line as the “Hela” cell in the cell biology of higher plants. Int. Rev. Cytol. 1992, 132, 1–30. [Google Scholar]
- Liu, Q. Mechanosensing in Plants—Mechanosensitive Calcium Channels versus the Cell Wall-Plasma Membrane-Cytoskeleton Continuum. Ph.D. Thesis, Karlsruhe Institute of Technology, Karlsruhe, Germany, 2014. Available online: https://publikationen.bibliothek.kit.edu/100004048 (accessed on 29 June 2022).
- Nick, P.; Heuing, A.; Ehmann, B. Plant chaperonins: A role in microtubule-dependent wall-formation? Protoplasma 2000, 211, 234–244. [Google Scholar] [CrossRef]
- Kuss-Wymer, C.L.; Cyr, R.J. Tobacco protoplasts differentiate into elongate cells without net microtubule depolymerization. Protoplasma 1992, 168, 64–72. [Google Scholar] [CrossRef]
- Zaban, B.; Maisch, J.; Nick, P. Dynamic actin controls polarity induction de novo in protoplasts. J. Integr. Plant Biol. 2013, 55, 142–159. [Google Scholar] [CrossRef]
- Brochhausen, L.; Maisch, J.; Nick, P. Break of symmetry in regenerating tobacco protoplasts is independent of nuclear positioning. J. Integr. Plant Biol. 2016, 58, 799–812. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Maisch, J.; Nick, P. Sensory role of actin in auxin-dependent responses of tobacco BY-2. J. Plant Physiol. 2017, 218, 6–15. [Google Scholar] [CrossRef]
- Bühler, K. Sprachtheorie: Die Darstellungsfunktion der Sprache; Gustav Fischer: Stuttgart, Germany, 1934. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Maisch, J.; Hayashi, K.-I.; Nick, P. Fluorescent Auxin Analogs Report Two Auxin Binding Sites with Different Subcellular Distribution and Affinities: A Cue for Non-Transcriptional Auxin Signaling. Int. J. Mol. Sci. 2022, 23, 8593. https://doi.org/10.3390/ijms23158593
Huang X, Maisch J, Hayashi K-I, Nick P. Fluorescent Auxin Analogs Report Two Auxin Binding Sites with Different Subcellular Distribution and Affinities: A Cue for Non-Transcriptional Auxin Signaling. International Journal of Molecular Sciences. 2022; 23(15):8593. https://doi.org/10.3390/ijms23158593
Chicago/Turabian StyleHuang, Xiang, Jan Maisch, Ken-Ichiro Hayashi, and Peter Nick. 2022. "Fluorescent Auxin Analogs Report Two Auxin Binding Sites with Different Subcellular Distribution and Affinities: A Cue for Non-Transcriptional Auxin Signaling" International Journal of Molecular Sciences 23, no. 15: 8593. https://doi.org/10.3390/ijms23158593
APA StyleHuang, X., Maisch, J., Hayashi, K.-I., & Nick, P. (2022). Fluorescent Auxin Analogs Report Two Auxin Binding Sites with Different Subcellular Distribution and Affinities: A Cue for Non-Transcriptional Auxin Signaling. International Journal of Molecular Sciences, 23(15), 8593. https://doi.org/10.3390/ijms23158593