
Helicobacter pylori, Protected from Antibiotics and Stresses Inside Candida albicans Vacuoles, Cause Gastritis in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
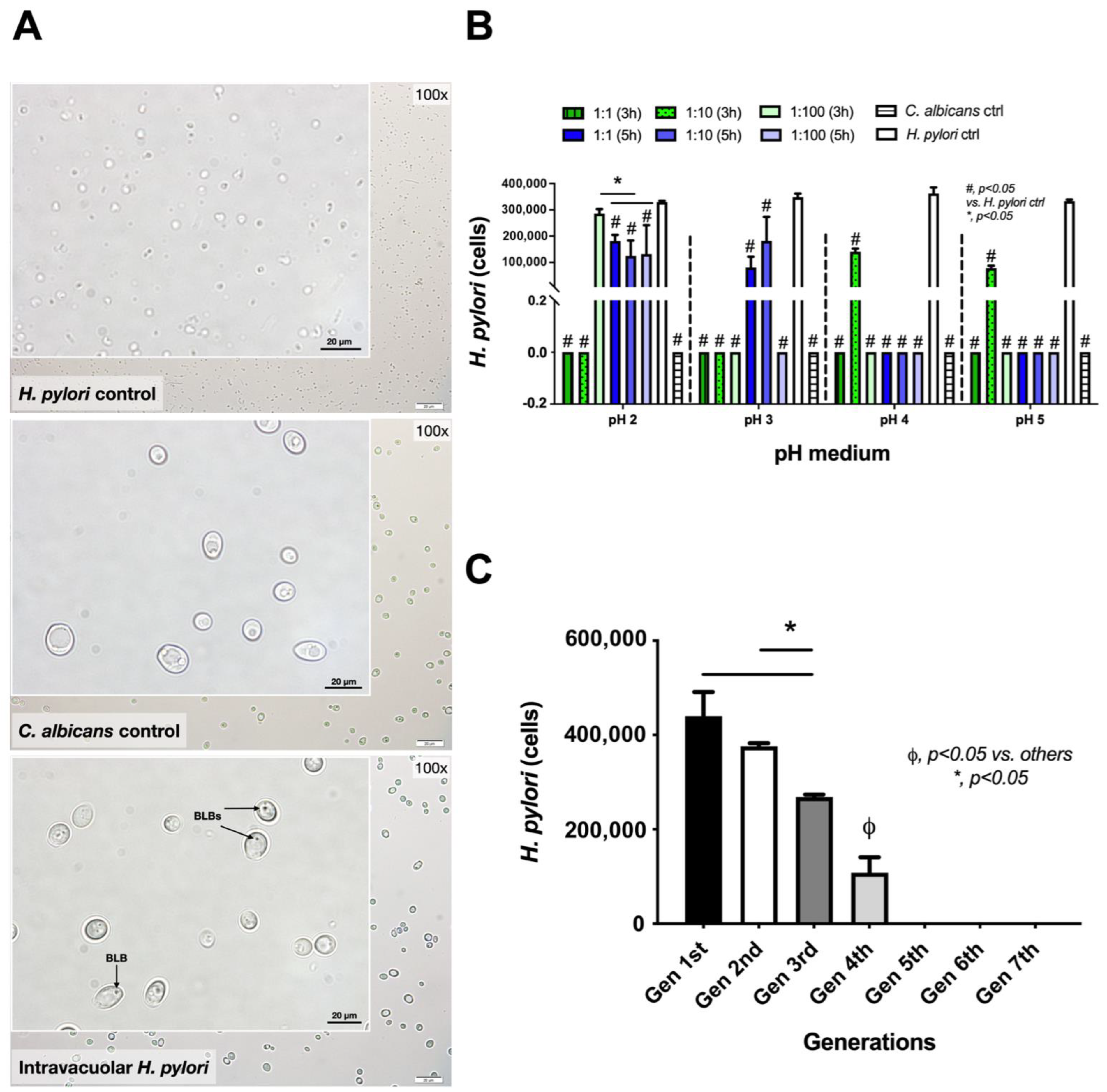
2.1. Induction of Intravacuolar H. pylori in Candida Yeast Cells
2.2. Release of Intravacuolar H. pylori from Candida Yeast Cells
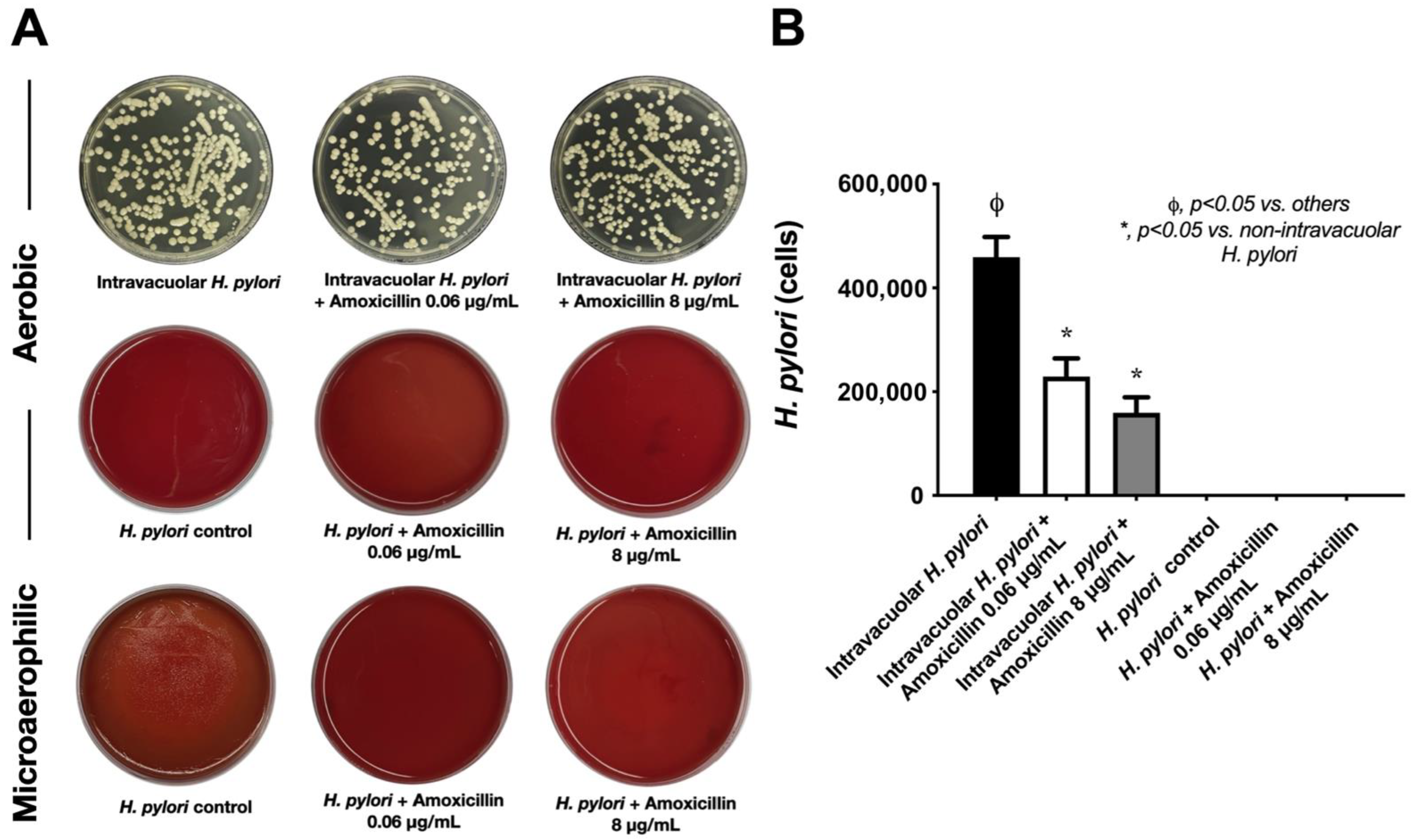
2.3. Intravacuolar H. pylori Was Protected from Antibiotics and Stress Conditions
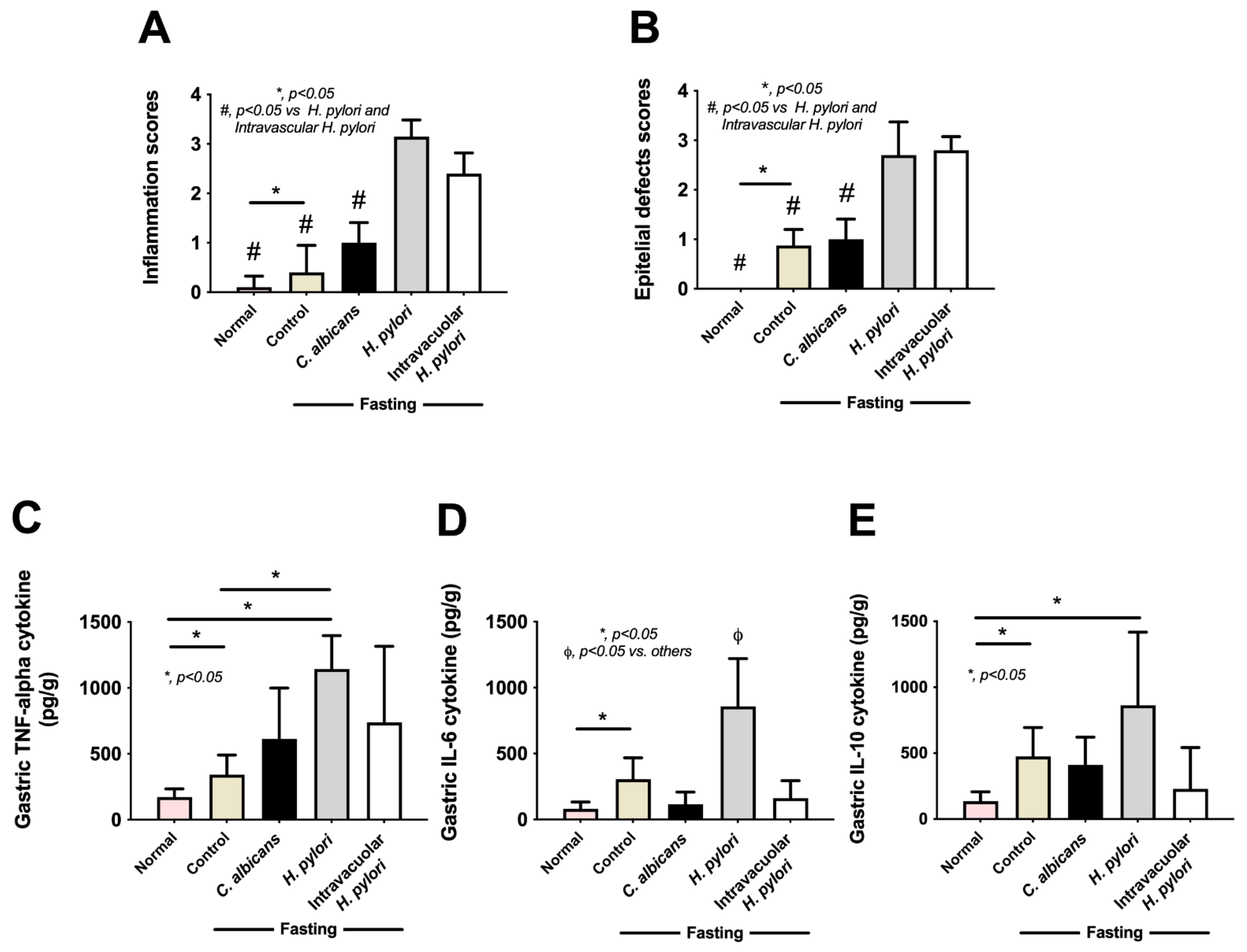
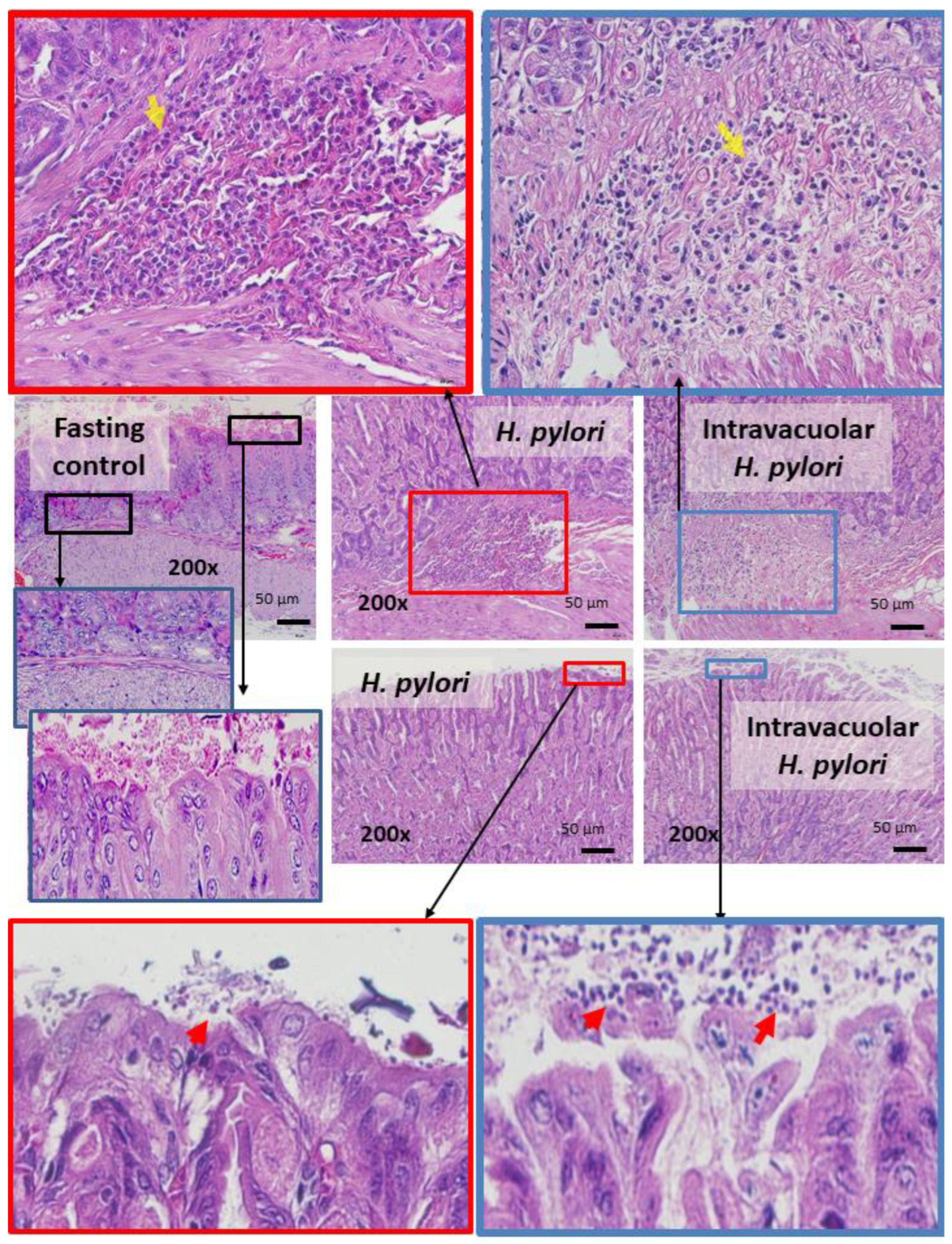
2.4. Candida Containing Intravacuolar H. pylori Caused Peptic Ulcer Disease
3. Discussion
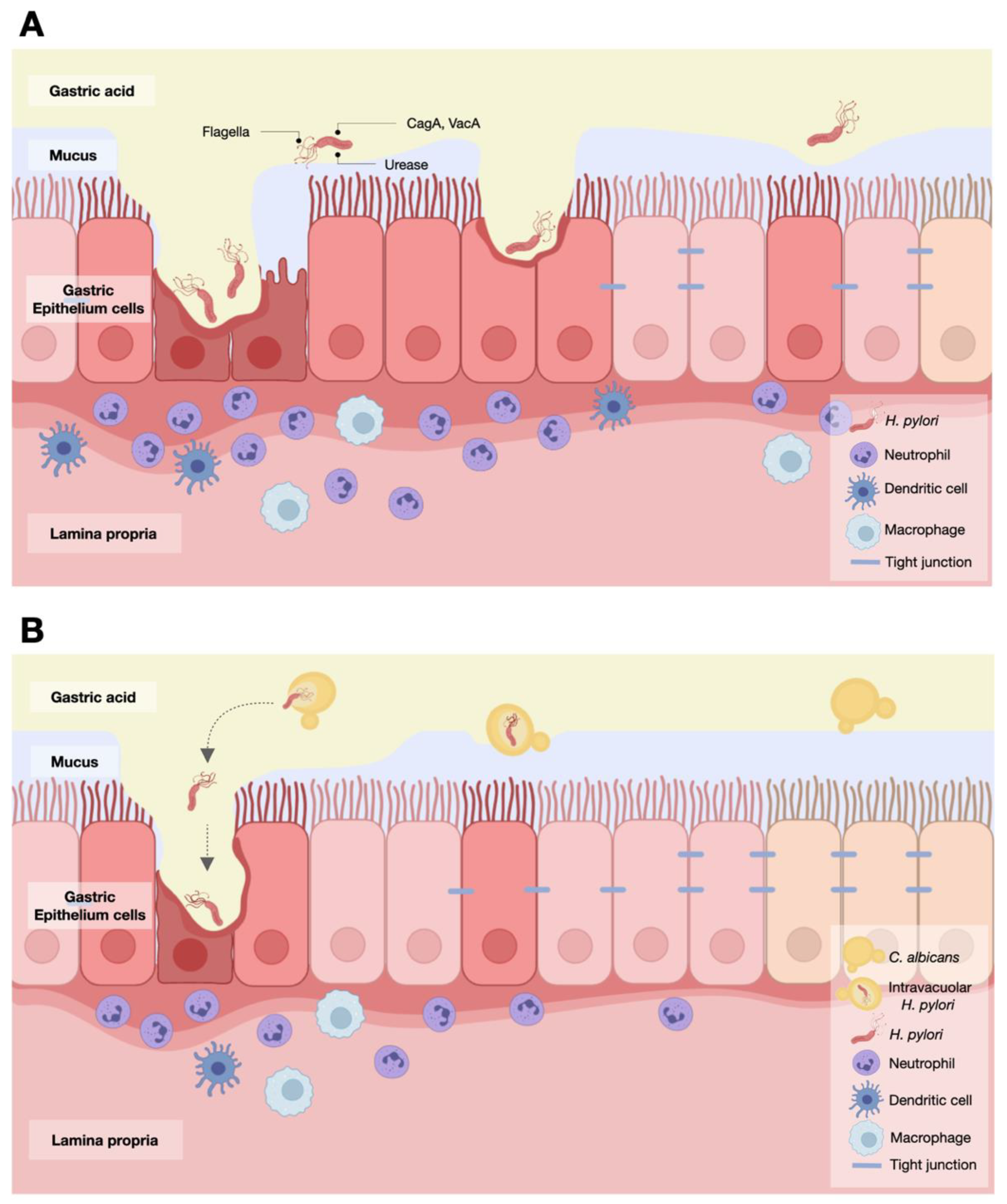
3.1. The Transfer of H. pylori Inside the Vacuoles of Candida Yeast Cells
3.2. Intravacuolar H. pylori Inside C. albicans as a Cause of Gastritis
4. Materials and Methods
4.1. Inducing Intravacuolar H. pylori in Candida Yeast Cells
4.2. Determination of Intravacuolar H. pylori Inside the Candida Yeast Cells
4.3. Antimicrobial Incubation and Stresses against Intravacuolar H. pylori
4.4. Animal and Peptic Ulcer Model
4.5. Mouse Gastric Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuna, L.; Jakab, J.; Smolic, R.; Raguz-Lucic, N.; Vcev, A.; Smolic, M. Peptic ulcer disease: A brief review of conventional therapy and herbal treatment options. J. Clin. Med. 2019, 8, 179. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, M.; Reddy, K.M.; Marsicano, E. Peptic ulcer disease and Helicobacter pylori infection. Mo. Med. 2018, 115, 219–224. [Google Scholar]
- Ramakrishnan, K.; Salinas, R.C. Peptic ulcer disease. Am. Fam. Physician 2007, 76, 1005–1012. [Google Scholar]
- Kavitt, R.T.; Lipowska, A.M.; Anyane-Yeboa, A.; Gralnek, I.M. Diagnosis and treatment of peptic ulcer disease. Am. J. Med. 2019, 132, 447–456. [Google Scholar] [CrossRef]
- Azhari, H.; Underwood, F.; King, J.; Coward, S.; Shah, S.; Chan, C.; Ho, G.; Ng, S.; Kaplan, G. The global incidence of peptic ulcer disease and its complications at the turn of the 21st century: A systematic review: 1199. Off. J. Am. Coll. Gastroenterol. ACG 2018, 113, S684–S685. [Google Scholar] [CrossRef]
- Woolf, A.; Rose, R. Gastric ulcer. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Bazzoli, F.; de Luca, L.; Graham, D.Y. Helicobacter pylori infection and the use of NSAIDs. Best Pract. Res. Clin. Gastroenterol. 2001, 15, 775–785. [Google Scholar] [CrossRef] [PubMed]
- De Brito, B.B.; da Silva, F.A.F.; Soares, A.S.; Pereira, V.A.; Santos, M.L.C.; Sampaio, M.M.; Neves, P.H.; de Melo, F.F. Pathogenesis and clinical management of Helicobacter pylori gastric infection. World J. Gastroenterol. 2019, 25, 5578–5589. [Google Scholar] [CrossRef]
- Diaconu, S.; Predescu, A.; Moldoveanu, A.; Pop, C.S.; Fierbinteanu-Braticevici, C. Helicobacter pylori infection: Old and new. J. Med. Life 2017, 10, 112–117. [Google Scholar]
- Denic, M.; Touati, E.; de Reuse, H. Review: Pathogenesis of Helicobacter pylori infection. Helicobacter 2020, 25 (Suppl. S1), e12736. [Google Scholar] [CrossRef]
- Kayali, S.; Manfredi, M.; Gaiani, F.; Bianchi, L.; Bizzarri, B.; Leandro, G.; di Mario, F.; de’Angelis Gian, L. Helicobacter pylori, transmission routes and recurrence of infection: State of the art. Acta Biomed. 2018, 89 (Suppl. S8), 72–76. [Google Scholar]
- Chang, W.L.; Yeh, Y.C.; Sheu, B.S. The impacts of H. pylori virulence factors on the development of gastroduodenal diseases. J. Biomed. Sci. 2018, 25, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massarrat, S.; Saniee, P.; Siavoshi, F.; Mokhtari, R.; Mansour-Ghanaei, F.; Khalili-Samani, S. The effect of Helicobacter pylori infection, aging, and consumption of proton pump inhibitor on fungal colonization in the stomach of dyspeptic patients. Front. Microbiol. 2016, 7, 801. [Google Scholar] [CrossRef] [PubMed]
- Roesler, B.M.; Rabelo-Gonçalves, E.M.; Zeitune, J.M. Virulence factors of Helicobacter pylori: A review. Clin. Med. Insights Gastroenterol. 2014, 7, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Shiota, S.; Suzuki, R.; Yamaoka, Y. The significance of virulence factors in Helicobacter pylori. J. Dig. Dis. 2013, 14, 341–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterbenc, A.; Jarc, E.; Poljak, M.; Homan, M. Helicobacter pylori virulence genes. World J. Gastroenterol. 2019, 25, 4870–4884. [Google Scholar] [CrossRef]
- Javed, S.; Skoog, E.C.; Solnick, J.V. Impact of Helicobacter pylori virulence factors on the host immune response and gastric pathology. Curr. Top Microbiol. Immunol. 2019, 421, 21–52. [Google Scholar]
- Dunne, C.; Dolan, B.; Clyne, M. Factors that mediate colonization of the human stomach by Helicobacter pylori. World J. Gastroenterol. 2014, 20, 5610–5624. [Google Scholar] [CrossRef]
- Wang, Z.; Ren, R.; Yang, Y. Mucosa microbiome of gastric lesions: Fungi and bacteria interactions. Prog. Mol. Biol. Transl. Sci. 2020, 171, 195–213. [Google Scholar]
- Gow, N.A.; van de Veerdonk, F.L.; Brown, A.J.; Netea, M.G. Candida albicans morphogenesis and host defence: Discriminating invasion from colonization. Nat. Rev. Microbiol. 2011, 10, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Dadar, M.; Tiwari, R.; Karthik, K.; Chakraborty, S.; Shahali, Y.; Dhama, K. Candida albicans—Biology, molecular characterization, pathogenicity, and advances in diagnosis and control—An update. Microb. Pathog. 2018, 117, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Hiengrach, P.; Visitchanakun, P.; Finkelman, M.A.; Chancharoenthana, W.; Leelahavanichkul, A. More prominent inflammatory response to pachyman than to whole-glucan particle and oat-β-glucans in dextran sulfate-induced mucositis mice and mouse injection through proinflammatory macrophages. Int. J. Mol. Sci. 2022, 23, 4026. [Google Scholar] [CrossRef] [PubMed]
- Saithong, S.; Worasilchai, N.; Saisorn, W.; Udompornpitak, K.; Bhunyakarnjanarat, T.; Chindamporn, A.; Tovichayathamrong, P.; Torvorapanit, P.; Chiewchengchol, D.; Chancharoenthana, W.; et al. Neutrophil extracellular traps in severe SARS-CoV-2 infection: A possible impact of LPS and (1→3)-β-d-glucan in blood from gut translocation. Cells 2022, 11, 1103. [Google Scholar] [CrossRef] [PubMed]
- Panpetch, W.; Somboonna, N.; Bulan, D.E.; Issara-Amphorn, J.; Worasilchai, N.; Finkelman, M.; Chindamporn, A.; Palaga, T.; Tumwasorn, S.; Leelahavanichkul, A. Gastrointestinal colonization of Candida albicans increases serum (1→3)-β-d-glucan, without candidemia, and worsens cecal ligation and puncture sepsis in murine model. Shock 2018, 49, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Saithong, S.; Saisorn, W.; Dang, C.P.; Visitchanakun, P.; Chiewchengchol, D.; Leelahavanichkul, A. Candida administration worsens neutrophil extracellular traps in renal ischemia reperfusion injury mice: An impact of gut fungi on acute kidney injury. J. Innate Immun. 2022, 1–16, Online First. [Google Scholar] [CrossRef]
- Amornphimoltham, P.; Yuen, P.S.T.; Star, R.A.; Leelahavanichkul, A. Gut leakage of fungal-derived inflammatory mediators: Part of a gut-liver-kidney axis in bacterial sepsis. Dig. Dis. Sci. 2019, 64, 2416–2428. [Google Scholar] [CrossRef]
- Issara-Amphorn, J.; Surawut, S.; Worasilchai, N.; Thim-Uam, A.; Finkelman, M.; Chindamporn, A.; Palaga, T.; Hirankarn, N.; Pisitkun, P.; Leelahavanichkul, A. The synergy of endotoxin and (1→3)-β-d-glucan, from gut translocation, worsens sepsis severity in a lupus model of Fc gamma receptor IIb-deficient mice. J. Innate Immun. 2018, 10, 189–201. [Google Scholar] [CrossRef]
- Thim-Uam, A.; Surawut, S.; Issara-Amphorn, J.; Jaroonwitchawan, T.; Hiengrach, P.; Chatthanathon, P.; Wilantho, A.; Somboonna, N.; Palaga, T.; Pisitkun, P.; et al. Leaky-gut enhanced lupus progression in the Fc gamma receptor-IIb deficient and pristane-induced mouse models of lupus. Sci. Rep. 2020, 10, 777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visitchanakun, P.; Saisorn, W.; Wongphoom, J.; Chatthanathon, P.; Somboonna, N.; Svasti, S.; Fucharoen, S.; Leelahavanichkul, A. Gut leakage enhances sepsis susceptibility in iron-overloaded β-thalassemia mice through macrophage hyperinflammatory responses. Am. J. Physiol.-Gastrointest. Liver Physiol. 2020, 318, G966–G979. [Google Scholar] [CrossRef] [PubMed]
- Hiengrach, P.; Panpetch, W.; Worasilchai, N.; Chindamporn, A.; Tumwasorn, S.; Jaroonwitchawan, T.; Wilantho, A.; Chatthanathon, P.; Somboonna, N.; Leelahavanichkul, A. Administration of Candida albicans to dextran sulfate solution treated mice causes intestinal dysbiosis, emergence and dissemination of intestinal Pseudomonas aeruginosa and lethal sepsis. Shock 2020, 53, 189–198. [Google Scholar] [CrossRef]
- Panpetch, W.; Kullapanich, C.; Dang, C.P.; Visitchanakun, P.; Saisorn, W.; Wongphoom, J.; Wannigama, D.L.; Thim-Uam, A.; Patarakul, K.; Somboonna, N.; et al. Candida administration worsens uremia-induced gut leakage in bilateral nephrectomy mice, an impact of gut fungi and organismal molecules in uremia. mSystems 2021, 6, e01187-20. [Google Scholar] [CrossRef] [PubMed]
- Panpetch, W.; Sawaswong, V.; Chanchaem, P.; Ondee, T.; Dang, C.P.; Payungporn, S.; Tumwasorn, S.; Leelahavanichkul, A. Candida administration worsens cecal ligation and puncture-induced sepsis in obese mice through gut dysbiosis enhanced systemic inflammation, impact of pathogen-associated molecules from gut translocation and saturated fatty acid. Front. Immunol. 2020, 11, 2278. [Google Scholar]
- Panpetch, W.; Hiengrach, P.; Nilgate, S.; Tumwasorn, S.; Somboonna, N.; Wilantho, A.; Chatthanathon, P.; Prueksapanich, P.; Leelahavanichkul, A. Additional Candida albicans administration enhances the severity of dextran sulfate solution induced colitis mouse model through leaky gut-enhanced systemic inflammation and gut-dysbiosis but attenuated by Lactobacillus rhamnosus L34. Gut Microbes 2020, 11, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Saithong, S.; Saisorn, W.; Visitchanakun, P.; Sae-Khow, K.; Chiewchengchol, D.; Leelahavanichkul, A. A synergy between endotoxin and (1→3)-β-d-glucan enhanced neutrophil extracellular traps in Candida administered dextran sulfate solution induced colitis in FcGRIIB-/-lupus mice, an impact of intestinal fungi in lupus. J. Inflamm. Res. 2021, 14, 2333–2352. [Google Scholar] [CrossRef] [PubMed]
- Issara-Amphorn, J.; Dang, C.P.; Saisorn, W.; Limbutara, K.; Leelahavanichkul, A. Candida administration in bilateral nephrectomy mice elevates serum (1→3)-β-d-glucan that enhances systemic inflammation through energy augmentation in macrophages. Int. J. Mol. Sci. 2021, 22, 5031. [Google Scholar] [CrossRef] [PubMed]
- Tungsanga, S.; Panpetch, W.; Bhunyakarnjanarat, T.; Udompornpitak, K.; Katavetin, P.; Chancharoenthana, W.; Chatthanathon, P.; Somboonna, N.; Tungsanga, K.; Tumwasorn, S.; et al. Uremia-induced gut barrier defect in 5/6 nephrectomized mice is worsened by Candida administration through a synergy of uremic toxin, lipopolysaccharide, and (1→3)-β-d-glucan, but is attenuated by Lacticaseibacillus rhamnosus L34. Int. J. Mol. Sci. 2022, 23, 2511. [Google Scholar] [CrossRef]
- Phuengmaung, P.; Panpetch, W.; Singkham-In, U.; Chatsuwan, T.; Chirathaworn, C.; Leelahavanichkul, A. Presence of Candida tropicalis on Staphylococcus epidermidis biofilms facilitated biofilm production and Candida dissemination: An impact of fungi on bacterial biofilms. Front. Cell. Infect. Microbiol. 2021, 11, 763239. [Google Scholar] [CrossRef] [PubMed]
- Phuengmaung, P.; Somparn, P.; Panpetch, W.; Singkham-In, U.; Wannigama, D.L.; Chatsuwan, T.; Leelahavanichkul, A. Coexistence of Pseudomonas aeruginosa with Candida albicans enhances biofilm thickness through alginate-related extracellular matrix but is attenuated by N-acetyl-l-cysteine. Front. Cell. Infect. Microbiol. 2020, 10, 594336. [Google Scholar] [CrossRef]
- Saniee, P.; Siavoshi, F.; Nikbakht Broujeni, G.; Khormali, M.; Sarrafnejad, A.; Malekzadeh, R. Localization of H. pylori within the vacuole of Candida yeast by direct immunofluorescence technique. Arch. Iran Med. 2013, 16, 705–710. [Google Scholar]
- Siavoshi, F.; Saniee, P. Vacuoles of Candida yeast as a specialized niche for Helicobacter pylori. World J. Gastroenterol. 2014, 20, 5263–5273. [Google Scholar] [CrossRef]
- Saniee, P.; Siavoshi, F.; Nikbakht Broujeni, G.; Khormali, M.; Sarrafnejad, A.; Malekzadeh, R. Immunodetection of Helicobacter pylori-specific proteins in oral and gastric Candida yeasts. Arch. Iran Med. 2013, 16, 624–630. [Google Scholar] [PubMed]
- Karczewska, E.; Wojtas, I.; Sito, E.; Trojanowska, D.; Budak, A.; Zwolinska-Wcislo, M.; Wilk, A. Assessment of co-existence of Helicobacter pylori and Candida fungi in diseases of the upper gastrointestinal tract. J. Physiol. Pharmacol. 2009, 60 (Suppl. 6), 33–39. [Google Scholar] [PubMed]
- Abrantes, P.; Africa, C.W.J. Measuring Streptococcus mutans, Streptococcus sanguinis and Candida albicans biofilm formation using a real-time impedance-based system. J. Microbiol. Methods 2020, 169, 105815. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhou, X.; Liao, B.; Zhou, Y.; Cheng, L.; Ren, B. The cross-kingdom interaction between Helicobacter pylori and Candida albicans. PLoS Pathog. 2021, 17, e1009515. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Alonzo, K.; Parra-Sepúlveda, C.; Vega, S.; Bernasconi, H.; Campos, V.L.; Smith, C.T.; Sáez, K.; García-Cancino, A. In vitro incorporation of Helicobacter pylori into Candida albicans caused by acidic pH stress. Pathogens 2020, 9, 489. [Google Scholar] [CrossRef] [PubMed]
- Matamala-Valdés, L.; Sánchez-Alonzo, K.; Parra, C.; Saez, K.; Aguayo-Reyes, A.; García, A. Detection of intracellular Helicobacter pylori in Candida. SPP from neonate oral swabs. Rev. Assoc. Méd. Bras. 2018, 64, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Alonzo, K.; Parra-Sepulveda, C.; Vergara, L.; Bernasconi, H.; Garcia-Cancino, A. Detection of Helicobacter pylori in oral yeasts from students of a Chilean university. Rev. Assoc. Méd. Bras. 2020, 66, 1509–1514. [Google Scholar] [CrossRef] [PubMed]
- Salmanian, A.H.; Siavoshi, F.; Akbari, F.; Afshari, A.; Malekzadeh, R. Yeast of the oral cavity is the reservoir of Heliobacter pylori. J. Oral Pathol. Med. 2008, 37, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Heydari, S.; Siavoshi, F.; Jazayeri, M.H.; Sarrafnejad, A.; Saniee, P. Helicobacter pylori release from yeast as a vesicle-encased or free bacterium. Helicobacter 2020, 25, e12725. [Google Scholar] [CrossRef]
- Pinto-Tomás, A.A.; Anderson, M.A.; Suen, G.; Stevensonm, D.M.; Chun, F.S.; Cleland, W.W.; Weimer, P.J.; Currie, C.R. Symbiotic nitrogen fixation in the fungus gardens of leaf-cutter ants. Science 2009, 326, 1120–1123. [Google Scholar] [CrossRef]
- Dantas Ada, S.; Day, A.; Ikeh, M.; Kos, I.; Achan, B.; Quinn, J. Oxidative stress responses in the human fungal pathogen, Candida albicans. Biomolecules 2015, 5, 142–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenço, A.; Pedro, N.A.; Salazar, S.B.; Mira, N.P. Effect of acetic acid and lactic acid at low pH in growth and azole resistance of Candida albicans and Candida glabrata. Front. Microbiol. 2019, 9, 3265. [Google Scholar] [CrossRef] [Green Version]
- Ansari, S.; Yamaoka, Y. Survival of Helicobacter pylori in gastric acidic territory. Helicobacter 2017, 22, e12386. [Google Scholar] [CrossRef]
- Clyne, M.; Dolan, B.; Reeves, E.P. Bacterial factors that mediate colonization of the stomach and virulence of Helicobacter pylori. FEMS Microbiol. Lett. 2007, 268, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Vylkova, S. Environmental pH modulation by pathogenic fungi as a strategy to conquer the host. PLoS Pathogens 2017, 13, e1006149-e. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, D.R.; Sachs, G.; Marcus, E.A. The role of acid inhibition in Helicobacter pylori eradication. F1000Res. 2016, 5, F1000, Faculty Rev-747. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.-C.; Lu, C.-W.; Lin, C.-J. Treatment of Helicobacter pylori infection: Current status and future concepts. World J. Gastroenterol. 2014, 20, 5283–5293. [Google Scholar] [CrossRef]
- Chen, M.-C.; Lei, W.-Y.; Lin, J.-S.; Yi, C.-H.; Wu, D.-C.; Hu, C.-T. Levofloxacin-amoxicillin/clavulanate-rabeprazole versus a standard seven-day triple therapy for eradication of Helicobacter pylori infection. BioMed Res. Int. 2014, 2014, 158520. [Google Scholar]
- Mazaheri Assadi, M.; Chamanrokh, P.; Whitehouse, C.A.; Huq, A. Methods for detecting the environmental coccoid form of Helicobacter pylori. Front. Public Health 2015, 3, 147. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Yang, Z.; Cheng, D.; Xie, C.; Zhu, Y.; Ge, Z.; Luo, Z.; Lu, N. Helicobacter pylori infection aggravates diet-induced insulin resistance in association with gut microbiota of mice. EBioMedicine 2016, 12, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Holland, R.L.; Bosi, K.D.; Harpring, G.H.; Luo, J.; Wallig, M.; Phillips, H.; Blanke, S.R. Chronic in vivo exposure to Helicobacter pylori VacA: Assessing the efficacy of automated and long-term intragastric toxin infusion. Sci. Rep. 2020, 10, 9307. [Google Scholar] [CrossRef] [PubMed]
- Sujan, K.; Islam, K. Effects of fasting on body weight, serum glucose and creatinine and histotexture of liver and kidney in Swiss albino mice. Res. Agric. Livest. Fish. 2020, 7, 421–430. [Google Scholar]
- Nosanchuk, J.D.; Nimrichter, L.; Casadevall, A.; Rodrigues, M.L. A role for vesicular transport of macromolecules across cell walls in fungal pathogenesis. Commun. Integr. Biol. 2008, 1, 37–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, C.P.; Leelahavanichkul, A. Over-expression of miR-223 induces M2 macrophage through glycolysis alteration and attenuates LPS-induced sepsis mouse model, the cell-based therapy in sepsis. PLoS ONE 2020, 15, e0236038. [Google Scholar]
- Visitchanakun, P.; Tangtanatakul, P.; Trithiphen, O.; Soonthornchai, W.; Wongphoom, J.; Tachaboon, S.; Srisawat, N.; Leelahavanichkul, A. Plasma miR-370-3P as a biomarker of sepsis-associated encephalopathy, the transcriptomic profiling analysis of microrna-arrays from mouse brains. Shock 2020, 54, 347–357. [Google Scholar] [CrossRef]
- Leelahavanichkul, A.; Somparn, P.; Panich, T.; Chancharoenthana, W.; Wongphom, J.; Pisitkun, T.; Hirankarn, N.; Eiam-Ong, S. Serum miRNA-122 in acute liver injury induced by kidney injury and sepsis in CD-1 mouse models. Hepatol. Res. 2015, 45, 1341–1352. [Google Scholar] [CrossRef]
- Peres da Silva, R.; Puccia, R.; Rodrigues, M.L.; Oliveira, D.L.; Joffe, L.S.; Cesar, G.V.; Nimrichter, L.; Goldenberg, S.; Alves, L.R. Extracellular vesicle-mediated export of fungal RNA. Sci. Rep. 2015, 5, 7763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, L.; Sood, P.; Lenardon, M.D.; Milne, G.; Olson, J.; Jensen, G.; Wolf, J.; Casadevall, A.; Adler-Moore, J.; Gow, N.A. The viscoelastic properties of the fungal cell wall allow traffic of AmBisome as intact liposome vesicles. mBio 2018, 9, e02383-17. [Google Scholar] [CrossRef] [Green Version]
- Deatherage, B.L.; Cookson, B.T. Membrane vesicle release in bacteria, eukaryotes, and archaea: A conserved yet underappreciated aspect of microbial life. Infect. Immun. 2012, 80, 1948–1957. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.; Sheh, A.; Feng, Y.; Muthupalani, S.; Ge, L.; Wang, C.; Kurnick, S.; Mannion, A.; Whary, M.T.; Fox, J.G. Helicobacter pylori-infected C57BL/6 mice with different gastrointestinal microbiota have contrasting gastric pathology, microbial and host immune responses. Sci. Rep. 2018, 8, 8014. [Google Scholar] [CrossRef]
- Cadamuro, A.C.T.; Rossi, A.F.T.; Maniezzo, N.M.; Silva, A.E. Helicobacter pylori infection: Host immune response, implications on gene expression and microRNAs. World J. Gastroenterol. 2014, 20, 1424–1437. [Google Scholar] [CrossRef] [Green Version]
- O’Morain, C.; Smith, S. Helicobacter pylori treatment failure: The rationale for alternative antibiotics. Digestion 2016, 93, 309–310. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.H.; Lu, C.Y.; Shih, H.Y.; Liu, C.J.; Wu, M.C.; Hu, H.M.; Hsu, W.H.; Yu, F.J.; Wu, D.C.; Kuo, F.C. CYP2C19 polymorphism influences Helicobacter pylori eradication. World J. Gastroenterol. 2014, 20, 16029–16036. [Google Scholar] [CrossRef]
- Jaka, H.; Mueller, A.; Kasang, C.; Mshana, S.E. Predictors of triple therapy treatment failure among H. pylori infected patients attending at a tertiary hospital in Northwest Tanzania: A prospective study. BMC Infect. Dis. 2019, 19, 447. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Q.; Hunt, R.H. Treatment after failure: The problem of “non-responders”. Gut 1999, 45 (Suppl. S1), I40–I44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moaddeb, A.; Fattahi, M.R.; Firouzi, R.; Derakhshandeh, A.; Farshad, S. Genotyping of the Helicobacter pylori cagA gene isolated from gastric biopsies in Shiraz, Southern Iran: A PCR-RFLP and sequence analysis approach. Jundishapur J. Microbiol. 2016, 9, e30046-e. [Google Scholar] [CrossRef] [Green Version]
- Surawut, S.; Panpetch, W.; Makjaroen, J.; Tangtanatakul, P.; Thim-Uam, A.; Wongphoom, J.; Tumwasorn, S.; Leelahavanichkul, A. Helicobacter pylori infection increased anti-dsDNA and enhanced lupus severity in symptomatic FcγRIIb-deficient lupus mice. Front. Microbiol. 2018, 9, 1488. [Google Scholar] [CrossRef]
- Rogers, A.B. Histologic scoring of gastritis and gastric cancer in mouse models. Methods Mol. Biol. 2012, 921, 189–203. [Google Scholar] [PubMed]
- Pennelli, G.; Grillo, F.; Galuppini, F.; Ingravallo, G.; Pilozzi, E.; Rugge, M.; Fiocca, R.; Fassan, M.; Mastracci, L. Gastritis: Update on etiological features and histological practical approach. Pathologica 2020, 112, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.B.; Taylor, N.S.; Whary, M.T.; Stefanich, E.D.; Wang, T.C.; Fox, J.G. Helicobacter pylori but not high salt induces gastric intraepithelial neoplasia in B6129 mice. Cancer Res. 2005, 65, 10709–10715. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hiengrach, P.; Panpetch, W.; Chindamporn, A.; Leelahavanichkul, A. Helicobacter pylori, Protected from Antibiotics and Stresses Inside Candida albicans Vacuoles, Cause Gastritis in Mice. Int. J. Mol. Sci. 2022, 23, 8568. https://doi.org/10.3390/ijms23158568
Hiengrach P, Panpetch W, Chindamporn A, Leelahavanichkul A. Helicobacter pylori, Protected from Antibiotics and Stresses Inside Candida albicans Vacuoles, Cause Gastritis in Mice. International Journal of Molecular Sciences. 2022; 23(15):8568. https://doi.org/10.3390/ijms23158568
Chicago/Turabian StyleHiengrach, Pratsanee, Wimonrat Panpetch, Ariya Chindamporn, and Asada Leelahavanichkul. 2022. "Helicobacter pylori, Protected from Antibiotics and Stresses Inside Candida albicans Vacuoles, Cause Gastritis in Mice" International Journal of Molecular Sciences 23, no. 15: 8568. https://doi.org/10.3390/ijms23158568
APA StyleHiengrach, P., Panpetch, W., Chindamporn, A., & Leelahavanichkul, A. (2022). Helicobacter pylori, Protected from Antibiotics and Stresses Inside Candida albicans Vacuoles, Cause Gastritis in Mice. International Journal of Molecular Sciences, 23(15), 8568. https://doi.org/10.3390/ijms23158568