
Irradiated Triple-Negative Breast Cancer Co-Culture Produces a Less Oncogenic Extracellular Matrix

 , ,
, ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. One Dose of 5 Gy Does Not Alter the Viability of the Cells
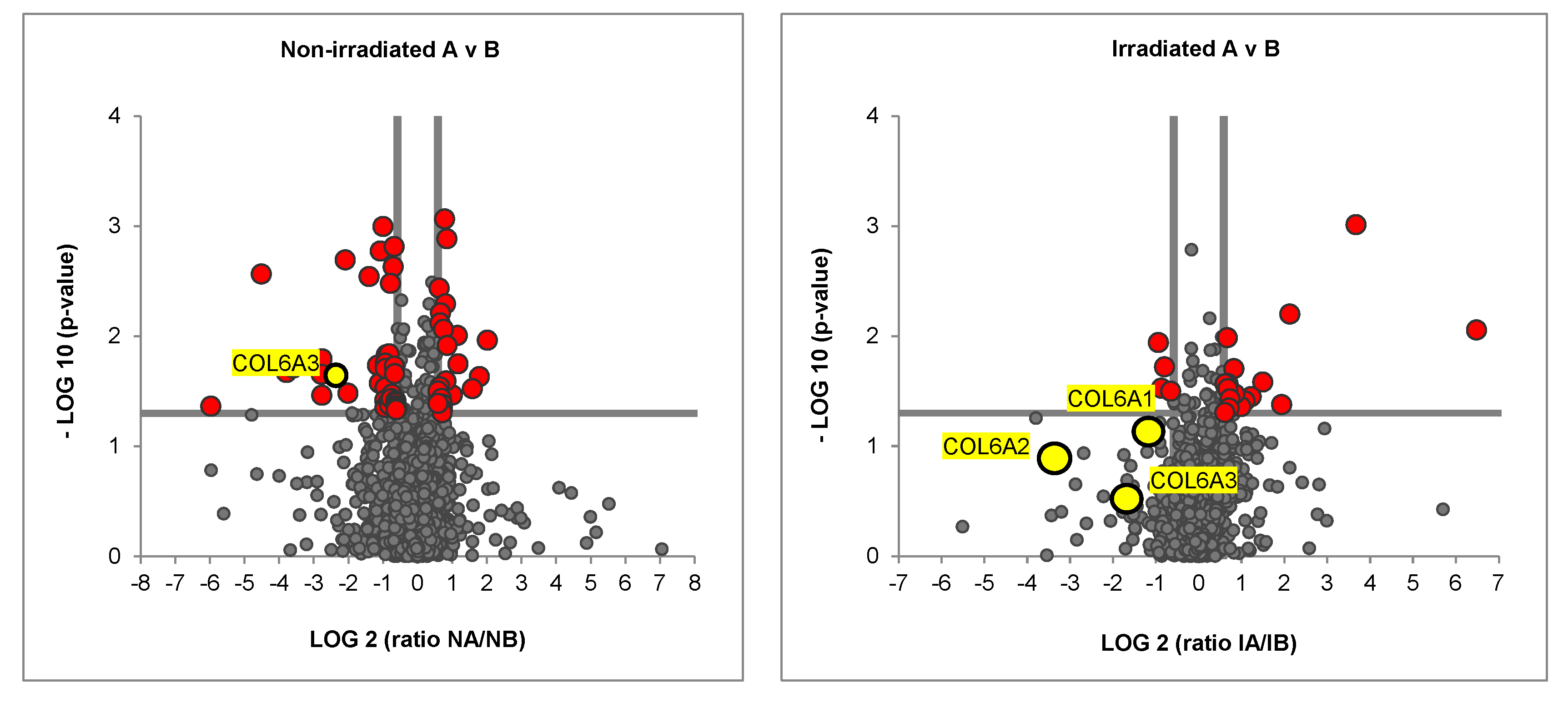
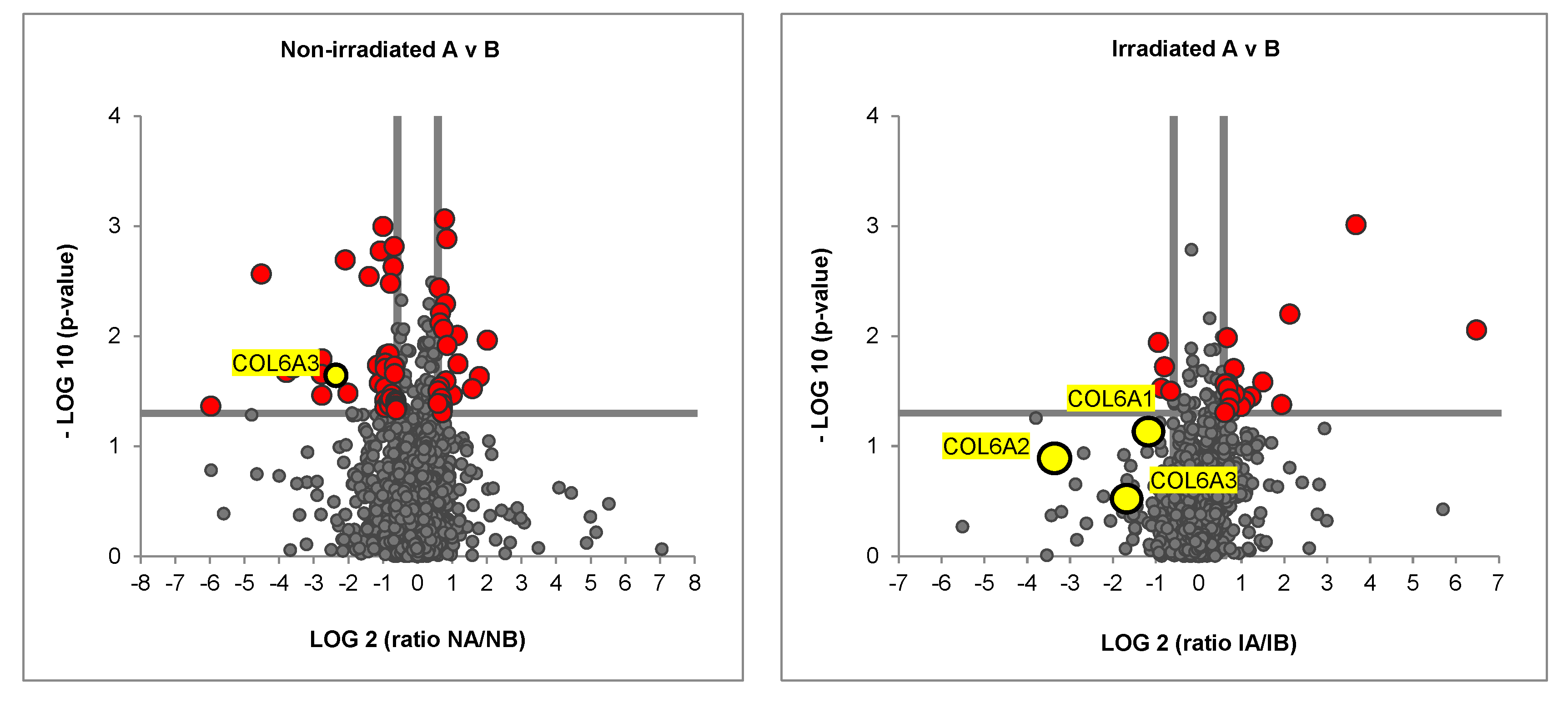
2.2. Irradiated Co-Culture (Fibroblasts, ASCs, and MDA-MB-231) Produces Matrix with Decreased Collagen Vi Isoforms
3. Discussion
4. Methods and Materials
4.1. Cell Lines and Cell Cultures
4.2. Cell Viability—Live/Dead Staining
4.3. Irradiation of Cells and Decellularization
4.4. Cell/Matrix Preparation for Mass Spectrometry and Proteomic Analysis
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Goldhirsch, A.; Winer, E.P.; Coates, A.S.; Gelber, R.D.; Piccart-Gebhart, M.; Thürlimann, B.; Senn, H.-J. Personalizing the treatment of women with early breast cancer: Highlights of the St Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2013. Ann. Oncol. 2013, 24, 2206–2223. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.J.; Naidu, S.; Topham, A.K.; Guiles, F.; Xu, Y.; McCue, P.; Schwartz, G.F.; Park, P.K.; Rosenberg, A.L.; Brill, K.; et al. Differences in breast carcinoma characteristics in newly diagnosed African–American and Caucasian patients: A single-institution compilation compared with the National Cancer Institute′s Surveillance, Epidemiology, and end results database. Cancer Interdiscip. Int. J. Am. Cancer Soc. 2007, 110, 876–884. [Google Scholar] [CrossRef]
- Wang, J.; Shi, M.; Ling, R.; Xia, Y.; Luo, S.; Fu, X.; Xiao, F.; Li, J.; Long, X.; Wang, J.; et al. Adjuvant chemotherapy and radiotherapy in triple-negative breast carcinoma: A prospective randomized controlled multi-center trial. Radiother. Oncol. 2011, 100, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Collignon, J.; Schroeder, H.; Lousberg, L.; Jerusalem, G. Triple-negative breast cancer: Treatment challenges and solutions. Breast Cancer Targets Ther. 2016, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Abdulkarim, B.S.; Cuartero, J.; Hanson, J.; Deschênes, J.; Lesniak, D.; Sabri, S. Increased Risk of Locoregional Recurrence for Women with T1-2N0 Triple-Negative Breast Cancer Treated With Modified Radical Mastectomy Without Adjuvant Radiation Therapy Compared With Breast-Conserving Therapy. J. Clin. Oncol. 2011, 29, 2852–2858. [Google Scholar] [CrossRef]
- Goodhead, D. Initial Events in the Cellular Effects of Ionizing Radiations: Clustered Damage in DNA. Int. J. Radiat. Biol. 1994, 65, 7–17. [Google Scholar] [CrossRef]
- Brett, E.; Sauter, M.; Timmins, É.; Azimzadeh, O.; Rosemann, M.; Merl-Pham, J.; Hauck, S.M.; Nelson, P.J.; Becker, K.F.; Schunn, I.; et al. Oncogenic Linear Collagen VI of Invasive Breast Cancer Is Induced by CCL5. J. Clin. Med. 2020, 9, 991. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Ponzetta, A.; Inforzato, A.; Jaillon, S. Innate immunity, inflammation and tumour progression: Double-edged swords. J. Intern. Med. 2019, 285, 524–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krisnawan, V.E.; Stanley, J.A.; Schwarz, J.K.; DeNardo, D.G. Tumor Microenvironment as a Regulator of Radiation Therapy: New Insights into Stromal-Mediated Radioresistance. Cancers 2020, 12, 2916. [Google Scholar] [CrossRef]
- Jarosz-Biej, M.; Smolarczyk, R.; Cichoń, T.; Kułach, N. Tumor microenvironment as a “game changer” in cancer radiotherapy. Int. J. Mol. Sci. 2019, 20, 3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-Negative Breast Cancer: Clinical Features and Patterns of Recurrence. Clin. Cancer Res. 2007, 13 Pt 1, 4429–4434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haffty, B.G.; Yang, Q.; Reiss, M.; Kearney, T.; Higgins, S.A.; Weidhaas, J.; Harris, L.; Hait, W.; Toppmeyer, D. Locoregional Relapse and Distant Metastasis in Conservatively Managed Triple Negative Early-Stage Breast Cancer. J. Clin. Oncol. 2006, 24, 5652–5657. [Google Scholar] [CrossRef] [PubMed]
- Taroni, P.; Quarto, G.; Pifferi, A.; Abbate, F.; Balestreri, N.; Menna, S.; Cassano, E.; Cubeddu, R. Breast Tissue Composition and Its Dependence on Demographic Risk Factors for Breast Cancer: Non-Invasive Assessment by Time Domain Diffuse Optical Spectroscopy. PLoS ONE 2015, 10, e0128941. [Google Scholar] [CrossRef] [Green Version]
- Jarosz-Biej, M.; Kamińska, N.; Matuszczak, S.; Cichoń, T.; Pamuła-Piłat, J.; Czapla, J.; Smolarczyk, R.; Skwarzyńska, D.; Kulik, K.; Szala, S. M1-like macrophages change tumor blood vessels and microenvironment in murine melanoma. PLoS ONE 2018, 13, e0191012. [Google Scholar] [CrossRef]
- Klein, D. The Tumor Vascular Endothelium as Decision Maker in Cancer Therapy. Front. Oncol. 2018, 8, 367. [Google Scholar] [CrossRef]
- Barker, H.E.; Paget, J.T.; Khan, A.A.; Harrington, K.J. The tumour microenvironment after radiotherapy: Mechanisms of resistance and recurrence. Nat. Rev. Cancer 2015, 15, 409–425. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Z.-G.; Yuan, H.; Deng, W.; Li, J.; Huang, Y.; Kim, B.Y.S.; Story, M.D.; Jiang, W. The Reciprocity between Radiotherapy and Cancer ImmunotherapyRadiosensitizing Immunotherapy. Clin. Cancer Res. 2019, 25, 1709–1717. [Google Scholar] [CrossRef] [Green Version]
- Tsoutsou, P.G.; Zaman, K.; Lluesma, S.M.; Cagnon, L.; Kandalaft, L.; Vozenin, M.-C. Emerging Opportunities of Radiotherapy Combined with Immunotherapy in the Era of Breast Cancer Heterogeneity. Front. Oncol. 2018, 8. [Google Scholar] [CrossRef]
- Weichselbaum, R.R.; Liang, H.; Deng, L.; Fu, Y.-X. Radiotherapy and immunotherapy: A beneficial liaison? Nat. Rev. Clin. Oncol. 2017, 14, 365–379. [Google Scholar] [CrossRef]
- Steward, L.T.; Gao, F.; Taylor, M.A.; Margenthaler, J.A. Impact of radiation therapy on survival in patients with triple-negative breast cancer. Oncol. Lett. 2013, 7, 548–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rave-Fränk, M.; Virsik-Köpp, P.; Pradier, O.; Nitsche, M.; Grünefeld, S.; Schmidberger, H. In vitro response of human dermal fibroblasts to X-irradiation: Relationship between radiation-induced clonogenic cell death, chromosome aberrations and markers of proliferative senescence or differentiation. Int. J. Radiat. Biol. 2001, 77, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Baaße, A.; Machoy, F.; Juerß, D.; Baake, J.; Stang, F.; Reimer, T.; Krapohl, B.D.; Hildebrandt, G. Radiation Sensitivity of Adipose-Derived Stem Cells Isolated from Breast Tissue. Int. J. Mol. Sci. 2018, 19, 1988. [Google Scholar] [CrossRef] [Green Version]
- Luan, A.; Duscher, D.; Whittam, A.J.; Paik, K.J.; Zielins, E.R.; Brett, E.A.; Atashroo, D.A.; Hu, M.S.; Lee, G.K.; Gurtner, G.C.; et al. Cell-Assisted Lipotransfer Improves Volume Retention in Irradiated Recipient Sites and Rescues Radiation-Induced Skin Changes. Stem Cells 2015, 34, 668–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, S.C.; A Khan, F.; Pant, I.; Shukla, A. Role of radiotherapy in early breast cancer: An overview. Int. J. Heal. Sci. 2007, 1, 259–264. [Google Scholar]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Merl, J.; Ueffing, M.; Hauck, S.M.; von Toerne, C. Direct comparison of MS-based label-free and SILAC quantitative proteome profiling strategies in primary retinal M üller cells. Proteomics 2012, 12, 1902–1911. [Google Scholar] [CrossRef]
- Hauck, S.M.; Dietter, J.; Kramer, R.L.; Hofmaier, F.; Zipplies, J.K.; Amann, B.; Feuchtinger, A.; Deeg, C.A.; Ueffing, M. Deciphering Membrane-Associated Molecular Processes in Target Tissue of Autoimmune Uveitis by Label-Free Quantitative Mass Spectrometry. Mol. Cell. Proteom. 2010, 9, 2292–2305. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brett, E.; Rosemann, M.; Azimzadeh, O.; Pagani, A.; Prahm, C.; Daigeler, A.; Duscher, D.; Kolbenschlag, J. Irradiated Triple-Negative Breast Cancer Co-Culture Produces a Less Oncogenic Extracellular Matrix. Int. J. Mol. Sci. 2022, 23, 8265. https://doi.org/10.3390/ijms23158265
Brett E, Rosemann M, Azimzadeh O, Pagani A, Prahm C, Daigeler A, Duscher D, Kolbenschlag J. Irradiated Triple-Negative Breast Cancer Co-Culture Produces a Less Oncogenic Extracellular Matrix. International Journal of Molecular Sciences. 2022; 23(15):8265. https://doi.org/10.3390/ijms23158265
Chicago/Turabian StyleBrett, Elizabeth, Michael Rosemann, Omid Azimzadeh, Andrea Pagani, Cosima Prahm, Adrien Daigeler, Dominik Duscher, and Jonas Kolbenschlag. 2022. "Irradiated Triple-Negative Breast Cancer Co-Culture Produces a Less Oncogenic Extracellular Matrix" International Journal of Molecular Sciences 23, no. 15: 8265. https://doi.org/10.3390/ijms23158265
APA StyleBrett, E., Rosemann, M., Azimzadeh, O., Pagani, A., Prahm, C., Daigeler, A., Duscher, D., & Kolbenschlag, J. (2022). Irradiated Triple-Negative Breast Cancer Co-Culture Produces a Less Oncogenic Extracellular Matrix. International Journal of Molecular Sciences, 23(15), 8265. https://doi.org/10.3390/ijms23158265