Abstract
Pituitary Adenylate Cyclase-Activating Peptide (PACAP) and Vasoactive Intestinal Peptide (VIP) are neuropeptides involved in a diverse array of physiological and pathological processes through activating the PACAP subfamily of class B1 G protein-coupled receptors (GPCRs): VIP receptor 1 (VPAC1R), VIP receptor 2 (VPAC2R), and PACAP type I receptor (PAC1R). VIP and PACAP share nearly 70% amino acid sequence identity, while their receptors PAC1R, VPAC1R, and VPAC2R share 60% homology in the transmembrane regions of the receptor. PACAP binds with high affinity to all three receptors, while VIP binds with high affinity to VPAC1R and VPAC2R, and has a thousand-fold lower affinity for PAC1R compared to PACAP. Due to the wide distribution of VIP and PACAP receptors in the body, potential therapeutic applications of drugs targeting these receptors, as well as expected undesired side effects, are numerous. Designing selective therapeutics targeting these receptors remains challenging due to their structural similarities. This review discusses recent discoveries on the molecular mechanisms involved in the selectivity and signaling of the PACAP subfamily of receptors, and future considerations for therapeutic targeting.
1. Introduction—Physiological Roles of the PACAP and VIP Receptors
Pituitary Adenylate Cyclase-Activating Peptide (PACAP) and Vasoactive Intestinal Peptide (VIP) are neuropeptides involved in a multitude of physiological and pathological processes. VIP is a 28 amino acid peptide that is widely expressed in various cell types in the central (CNS) and peripheral nervous systems [1]. Centrally, VIP promotes neuronal survival [2] and regulates glycogen metabolism in astrocytes [3], while peripherally, VIP functions as a vasodilator leading to non-adrenergic and non-cholinergic relaxation of vascular and non-vascular smooth muscle through co-transmission with nitric oxide and carbon monoxide [4,5]. VIP is also co-transmitted with acetylcholine in exocrine glands [5,6,7], and works as a secretagogue by stimulating prolactin secretion in the pituitary and catecholamine release in the adrenal medulla [8,9]. In the immune system, VIP is an immunoregulator that controls T-lymphocyte trafficking and inhibits interleukin-2 (IL2) production [10]. VIP also stimulates electrolyte secretion in the jejunum [11,12], and plays a protective role against oxidant injury [13].
Similar to VIP, PACAP is widely expressed and is involved in numerous physiological processes, mediated both centrally and peripherally [14,15,16]. PACAP exists in two isoforms from proteolysis of the same precursor protein in the body, PACAP38 and PACAP27, which differ by eleven amino acid residues in the C-terminus [17,18]. Both PACAP38 and PACAP27 are bioactive, however, PACAP38 is more than 100-fold more abundant than PACAP27 in neuronal tissues [14,15]. PACAP plays important roles in the CNS as a neurotransmitter, being involved in learning and memory [19], and control of circadian rhythms [20]. Additionally, PACAP exerts a neuroprotective effect in responses to stress [21], including during brain injury and ischaemia [22,23]. Peripherally, PACAP modulates control of vasodilation [24,25], and secretion of insulin [26], adrenaline and anterior pituitary hormone [27,28].
PACAP and VIP exert their physiological effects by activating the PACAP subfamily of G protein-coupled receptors (GPCRs) that consist of VIP receptor 1 (VPAC1R), VIP receptor 2 (VPAC2R) and the PACAP type I receptor (PAC1R). These receptors are distributed in the central and peripheral nervous systems, endocrine system, and immune system, and mediate the physiological effects of the peptides described above, including control of circadian rhythm [29,30,31], immune regulation [32,33], and insulin secretion [34]. A summary of the distribution and physiological or potential therapeutic roles of these receptors is described below (Table 1).

Table 1.
Distribution and physiological or therapeutic role of the PACAP subfamily of receptors.
Due to their extensive, yet discrete, distribution and diverse physiological roles, the PACAP subfamily receptors are attractive targets for the development of therapeutics, particularly for the treatment of chronic inflammation, nociceptive pain, and neurodegeneration (Table 1) [33,85,86].
The PAC1R is involved in the regulation of pleiotropic neurological functions including promotion of neural survival and synaptic plasticity, nociceptive pain, and regulating the hypothalamic–pituitary–adrenal axial stress response, which makes it an attractive target for nervous system disorders including migraine [85,102,103], secondary injury in traumatic brain injury [84], and post-traumatic stress disorder [81,83]. The PAC1R is expressed in regions of the trigeminal-autonomic system that are associated with migraine, namely the sphenopalatine autonomic ganglia neurons and spinal trigeminal nucleus [104]. PAC1R activation mediates intracranial nociceptive activation of the central trigeminal-vascular neurons and induces neurogenic dural vasodilation contributing to migraine pathophysiology [102,103]. PACAP/PAC1R signaling is also involved in the stress mechanisms of the body [105,106]. PACAP and PAC1R are highly expressed in stress-associated brain regions in the hypothalamic and limbic structures [14,73,82]. PAC1R signaling modulates corticosterone and corticotropin-releasing hormone levels [107,108], with PACAP and PAC1R knockout mice displaying reduced anxiety-like phenotypes and blunted corticosterone response [83,109]. In women, PACAP serum levels are directly correlated to PTSD symptoms [83]. These sex-specific effects are speculated to be associated with oestrogen-dependent regulation of PACAP systems within the bed nucleus of stria terminalis in the extended amygdala [83].
On the other hand, VPAC receptors are attractive targets for immune and inflammatory conditions. VPAC1Rs are expressed constitutively, or in the case of VPAC2Rs, following induction, in T-lymphocytes and macrophages in the immune system [53,54,55,56]. In homeostatic states, the VPAC1 and VPAC2 receptors play an important role in preserving the equilibrium of pro- and anti-inflammatory responses through the regulation of various inflammatory mediators in immune cells [53,54,56]. While in pathological inflammatory states, these receptors can promote anti-inflammatory outcomes by inhibiting the pro-inflammatory Th1 and Th17 responses and stimulating Th2 and Treg responses [110,111,112]. As such, there is significant interest in targeting these receptors for the treatment of chronic inflammatory diseases including Crohn’s [113], Sjogren’s syndrome [114], COPD [45], autoimmune encephalitis [115], and rheumatoid arthritis [116].
2. Pharmacology of PACAP and VIP Receptors
To coordinate the diverse physiological outcomes mediated by the activation of the PACAP subfamily receptors by their cognate peptide ligands, these receptors activate multiple signaling pathways through the coupling of different G proteins, as well as through the interactions with a myriad of scaffolding and accessory proteins to stimulate additional signaling pathways.
2.1. Binding Characteristics of Natural Peptides Differentiating the PACAP Subfamily of Receptors
The PACAP subfamily receptors were initially differentiated by their distinct peptide binding affinity profiles. Early studies in tissues using 125I-labelled PACAP identified two types of PACAP receptors. The first type, which was classified as the PACAP type I receptor or PAC1R, had a high binding affinity for PACAP (Kd of approximately 0.5 nM) with a lower affinity for VIP (Kd > 500 nM) [117,118,119,120]. In contrast, the second type of PACAP receptors had a similar affinity (Kd of approximately 0.5 nM) for both VIP and PACAP and were subsequently classified as PACAP type II receptors [14,118,119]. Later, it was discovered that the type II PACAP receptor subclass contained two distinct types of receptors [121,122]. One (later named VPAC1R) had a lower affinity for helodermin, an exogenous peptide derived from the venom of the lizard Heloderma suspectum [123,124], compared to VIP and PACAP [125], and the other (VPAC2R) had a similar affinity toward all three peptides [59,126].
Other peptide agonists of the VPAC receptors were later identified, including peptide histidine isoleucine (PHI) in rodents and peptide histidine methionine (PHM) in humans, which are derived from post-translational processing of the VIP precursor following mRNA translation and were revealed to be biologically active [127,128,129]. These peptides share almost 50% sequence homology to VIP and bind with lower affinity to the VPAC1R and VPAC2R receptors compared to VIP [130,131]. Maxadilan, a PAC1R selective agonist that was originally isolated from the sand-fly (Lutzomyia longipalpis) salivary gland [132], shares no significant sequence similarity to VIP and PACAP, but can also bind and activate the PAC1R with a similar affinity and potency to PACAP [133,134].
Selective Peptide Analogues of PAC1R, VPAC1R, and VPAC2R
Several of the natural peptides described above are able to interact with multiple members of the PACAP receptor subfamily. However, non-selective binding may lead to undesirable side effects in clinical practice, hence the development of receptor-selective ligands may improve therapeutic utility. Before the determination of the PACAP-bound VPAC1R, VPAC2R, and PAC1R structures through cryo-electron microscopy (cryo-EM) [135,136,137,138,139], peptide analogues were designed and generated through systematic residue scanning of the peptide sequences seeking improved receptor selectivity. These analogues were subsequently screened in in vitro assays for binding and receptor signaling. From this process, several peptide analogues were identified that exhibited improved selectivity within the PACAP subfamily of receptors (Table 2).
Table 2.
VPAC1R, VPAC2R, or PAC1R selective peptide analogues.
By categorizing receptor-selective analogues by their modifications to the natural peptide, several patterns emerge that provide a foundation for structure-activity relationship analysis and drug design. Firstly, deletion or modification of the N-terminal residues in VIP and PACAP analogues can be used to generate antagonists as seen with PACAP (6-38), PG 97-269, and PG 99–465 [145,147,149,152] (Table 3). As observed in general for endogenous Class B1 GPCR peptides, the N-terminal region is essential for receptor activation. The N-terminal truncation of multiple endogenous peptides of class B1 GPCRs, including corticotropin-releasing factor (CRF), the CRF analogue urotensin, the glucagon-like peptide-1 receptor (GLP-1R) agonist exendin, calcitonin, parathyroid hormone (PTH)-related peptide (PTHrP), glucagon and secretin results in the production of antagonists to their respective receptors [153,154,155,156,157,158,159].
Table 3.
Deletion (grey) or modification (red) of the N-terminal residues in VIP and PACAP analogues can be used to generate receptor-selective peptide antagonists.
Secondly, at the C-terminal end, modifications to peptide length play a role in VPAC1R/VPAC2R selectivity (Table 4). RO 25-1553 was initially developed as a long-acting, VPAC2R selective, VIP analogue to overcome some of the deficiencies in the natural peptide for clinical bronchodilatory action; namely the short half-life in vivo due to Thr7P-Asp8P and Ser25P-Ile26P cleavage. The VIP analogue RO 25-1553 introduced a lactam bridge between the amino acid residues 21 and 25, along with an elongated C-terminal tail (Asp25P-Leu26P-Lys27P-Lys28P-Gly29P-Gly30P-Thr31P), generating a peptide with a long duration of bronchodilatory activity, compared with the natural peptide [160]. PG 96-249a, a RO 25-1553 analogue where the lactam bridge between positions 21 and 25 is absent, retained VPAC2R selectivity [147], suggesting that the elongation of the C-terminus rather than the lactam bridge was involved in VPAC2R selectivity. This improvement in VPAC2R selectivity with C-terminal peptide length is also observed when extending the C-terminal end of a VIP analogue from 28 to 31 residues with the equivalent PACAP38 residues [161]. On the other hand, [Arg16]-PACAP (1–23), a C-terminally truncated analogue was selective for VPAC1R, while the full-length [Arg16]-PACAP (1–27) was non-selective between VPAC1 and VPAC2R [143].
Table 4.
Truncation (grey) and elongation (red) peptide length contribute to VPAC1R/VPAC2R-selective peptide analogues.
Thirdly, when analyzing common residues where modifications alter receptor selectivity, position 22 on the C-terminal end of VIP appears to play a role in VPAC1R/VPAC2R selectivity [141]. Alanine scanning revealed that the aromatic Tyr residue at position 22 was important for high-affinity VPAC2R binding, while it was not as crucial for VPAC1R binding [141]. These empirical observations in conjunction with emerging structural data provide guidance for future peptide and potentially small molecule drug design for VIP and PACAP receptor selective therapeutics.
2.2. Signaling Characteristics of PAC1R, VPAC1R, and VPAC2R
Similar to other class B1 GPCRs, the PACAP family receptors predominantly couple to Gs protein to activate the adenylate cyclase (AC) to produce cAMP (3′,5′-cyclic adenosine monophosphate) (Figure 1). However, some members of this receptor family also couple to Gq/11 and Gi/o proteins to induce activation of phospholipase C (PLC) and intracellular calcium (Ca2+) mobilization [162,163,164,165]. Many class B1 GPCRs also signal through scaffolding proteins such as β-arrestin and A-kinase anchoring proteins (AKAPs) [166,167]. Furthermore, certain receptors in this family can associate with receptor activity-modifying proteins (RAMPs) to modulate receptor signaling and trafficking [168,169,170,171,172].
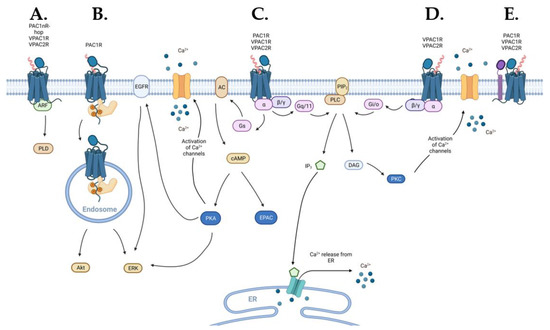
Figure 1.
Schematic diagram of the signaling pathways activated by the PACAP subfamily of receptors (blue): (A) VPACRs and the PAC1R splice variant PAC1nR-hop can engage ADP-ribosylation factor (ARF) signaling, (B) endosomal signaling may be activated by β-arrestin (light-orange) recruitment to PAC1R, (C) VPAC1R, VPAC2R, and PAC1R directly engage Gs- and Gq/11-coupled pathways, (D) VPAC1R and VPAC2R also directly activate Gi/o-coupled pathways, and (E) PACAP subfamily of receptors can interact, in a receptor-specific manner, with receptor activity-modifying protein (RAMP) (purple) that can alter receptor signaling or trafficking. Abbreviations used in this figure: ARF—ADP-ribosylation factor, PLD—phospholipase D, ERK—extracellular signal-regulated kinase, EGFR—epidermal growth factor receptor, AC—adenylate cyclase, cAMP—3′,5′-cyclic adenosine monophosphate, PKA—protein kinase A, EPAC—exchange protein directly activated by cAMP, Ca2+—calcium, PLC—phospholipase C, PIP2—phosphatidylinositol 4,5-bisphosphate, IP3—inositol 1,4,5-trisphosphate, DAG—diacylglycerol, ER—endoplasmic reticulum, PKC—protein kinase C.
2.2.1. Alternative Splicing
Some class B1 receptors, including the CRF, calcitonin, and PTH1R, undergo alternative splicing events resulting in receptor variants that can exhibit diverse signaling outcomes [173]. Alternative splicing is a regulatory mechanism during the pre-mRNA splicing process that results in the production of multiple mRNA variants from a single gene [174]. PAC1R undergoes extensive alternative splicing resulting in a host of receptor variants known as splice isoforms [175,176,177,178,179], while reports of this phenomenon occurring in the VPAC receptors are limited [173,180]. In humans, these splice isoforms occur in the N-terminal extracellular domain (ECD) and/or the intracellular loop 3 (ICL3) of PAC1R with the most prevalent and common variant containing a full-length N-terminal ECD and no ICL3 insertions (PAC1R null, a.k.a. PAC1nR) (Figure 2) [176,181].
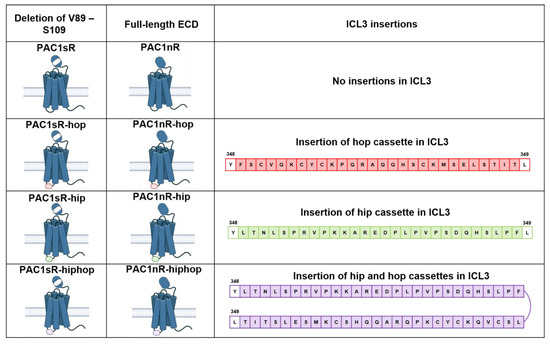
Figure 2.
Schematic illustration of PAC1R splice isoforms occurring in the N-terminal extracellular domain (ECD) and/or intracellular loop 3 (ICL3) in humans. N-terminal ECD splice variation arises from the presence (PAC1R null a.k.a. PAC1nR) or deletion (PAC1R-short a.k.a. PAC1sR) of 21 amino acids (residues V89 to S109) from the N-terminal ECD. ICL3 splice variation arises from the inclusion of one or two of the cassette exons, the “hip” cassette (red) and the “hop” cassette (green) of 28 amino acids each, between residues Y348 and L349 of the PAC1R ICL3. Inclusion of the hip cassette in PAC1R leads to the splice isoform PAC1nR-hip or PAC1sR-hip. Inclusion of the hop cassette leads to the PAC1nR-hop or PAC1sR-hop splice isoform, while the inclusion of both hip and hop cassettes leads to PAC1nR-hiphop or PAC1sR-hiphop (purple).
Splice isoforms involving the N-terminal ECD including the variant with a truncation within the ECD, PAC1R short (PAC1sR), can affect ligand-binding specificity and affinity, while isoforms of ICL3 may alter G protein coupling and receptor trafficking, enriching complexity in PAC1R signaling [175,178,182,183,184,185]. Splice isoforms from alternative splicing events at ICL3 are characterized by the inclusion of one or two of the cassette exons, exons 14 (the “hip” cassette) and 15 (the “hop” cassette) of 28 amino acids each [180]. The inclusion of the hip cassette in PAC1R leads to the splice isoform PAC1R-hip [176]. Inclusion of the hop cassette leads to the PAC1R-hop, while the inclusion of both hip and hop cassettes leads to PAC1R-hiphop [181,186]. While these splice variants may have distinct pharmacological profiles, the physiological implications of these have not been studied in detail.
A summary of the expression of the PAC1R splice isoforms in the nervous system is published in Blechman et al. [180]; the spatiotemporal differences in isoform expression during embryonic and postnatal development suggest a role of the splice isoforms in the regulation of neurodevelopmental processes. While limited studies have characterized PAC1R splice isoform expression in humans, studies characterizing isoform expression in other species during development have been performed. In studies in rats, the hop, hiphop, and short PAC1R splice isoforms were expressed in the developing brain with an expression of the hop and short isoform increasing from embryonic days 10 to 21 and the hiphop variant being detected from embryonic day 17 to birth [89]. Rat cortical expression of the hip, hop and hiphop PAC1R splice variants was highest immediately after birth with expression levels decreasing significantly one month later [187]. In the rat retina, expression of the null and hip isoforms was the highest immediately after birth, with expression decreasing overtime suggesting that these isoforms might be involved in early stage retinal development, while the hop isoform expression increased at day 10–20 postnatal suggesting a role in later-phase retinal development [188]. Interestingly in both these studies, the expression of VPAC1R and VPAC2R increased over time with cortical expression increasing significantly one month postnatal and retinal expression peaking at days 10–15 after birth for VPAC1R and days 5–15 after birth for VPAC2R [187,188]. Additionally, the PAC1-hop isoform was induced in response to stress in zebrafish [189]. An age-dependent effect in hop isoform-deficient zebrafish was observed, where the hop isoform-deficient larvae exhibited increased anxiety-like behavior, while the adult hop isoform-deficient zebrafish exhibited the opposite response to novelty stress, suggesting a role of this PAC1R isoform in stress adaptation response development [190]. However, species differences must be taken into account when extrapolating these findings to humans. In one study, where isoform expression in the rat and human placenta was studied, expression of the short, the hip or hop and the, hiphop PAC1R isoforms were detected in the rat placenta, while only the hop isoform was detected in the human placenta [191].
2.2.2. Signaling via Gs Coupling
All three receptors in the PACAP subfamily couple to the Gs protein resulting in the activation of AC and production of cAMP (Figure 1C) [146,178,192]. cAMP is a nucleotide that acts as a second messenger in various signaling pathways. cAMP can activate numerous effector proteins including protein kinase A (PKA) and exchange proteins activated by cAMP (EPAC).
PAC1R-induced cAMP production is associated with broad roles in the nervous system including the promotion of neuronal survival, regulation of cerebellar development, neuritogenesis, and the enhancement of N-methyl-D-aspartate (NMDA) receptor activity through PKA signaling [193,194,195,196,197]. PAC1R-induced cAMP activation of EPAC has been associated with neuronal excitability and differentiation through activation of Rap1 [198,199]. In addition, PAC1R-induced cAMP signaling is associated with neuronal differentiation independently of PKA and EPAC activation through an effector termed the neuritogenic cAMP sensor [200,201]. This signaling pathway involved the activation of extracellular signal-regulated kinase (ERK) by the neuritogenic cAMP sensor [200].
For the VPAC receptors, VPAC2R signaling by cAMP/PKA promotes circadian clock gene expression through cAMP-response element binding (CREB) phosphorylation [202], enhances synaptic transmission to hippocampal CA1 pyramidal cells [203], and through activating ERK phosphorylation, regulates anterior pituitary cell secretion and proliferation [204]. In the immune system, VPAC1R inhibits TNFα production through cAMP/PKA activation in macrophages [205], and is associated with pro-inflammatory exocytosis of secretory vesicles and granules from monocytes through cAMP/EPAC activation [206].
2.2.3. Signaling via Gq/11 and Gi/o Coupling
All three receptors of the PACAP subfamily also couple to Gq/11 resulting in the activation of PLC (Figure 1C) [178,192,207]. PLC cleaves phosphatidylinositol 4,5-bisphosphate to form diacylglycerol (DAG) and inositol 1,4,5-trisphosphate (IP3). In turn, DAG activates phosphokinase C (PKC), while IP3 stimulates intracellular Ca2+ release from the endoplasmic reticulum [177,180,208].
Gq/11 coupling plays an important role in PAC1R-modulated NMDA receptor activity and neuroplasticity in the CNS. While both PAC1R-mediated Gs/cAMP/PKA signaling and PKC signaling through Gq/11 are involved in the enhancement of NMDA receptor-mediated activity, PLC knockout resulted in a major reduction in the PAC1R-activated NMDA receptor-mediated excitatory postsynaptic currents despite a similar level of cAMP/PKA activation, suggesting Gq/11/PKC signaling is the dominant signaling pathway in modulation of NMDA receptors in hippocampal CA1 neurons [196,209]. PAC1R/PKC signaling can also activate ERK pathways for adaptative neuronal responses [210,211]. Additionally, in rat cerebral cortical slices, PAC1R stimulates PLC-IP3/DAG signaling in astrocytes [209,212].
While PAC1R activation of PLC has thus far been reported to exclusively involve Gq/11-mediated signaling, VPAC1R and VPAC2R can interact with both Gi/o and Gq/11 proteins to elicit PLC activation [207,213]. However, this response appears to be species and cell-type dependent. VPAC1R couples to Gi3 in rat alveolar macrophages [214], while PLC activation is exclusively mediated through Gq/11 by human VPAC1R and VPAC2R when overexpressed in COS-7 cells [207]. In gastric muscle cells, VPAC2R stimulates nitric oxide synthase activity through Gi1-2 [215]. While in the pancreas, VPAC2R activates PLC and mobilizes intracellular Ca2+ through a Gi-activated mechanism [216].
2.2.4. Non-G Protein Signaling
In addition to direct G protein-mediated signaling, the VPAC receptors and certain variants of the PAC1R can signal via G protein-dissociated mechanisms [217,218,219].
ADP-Ribosylation Factor
The VPAC1R and VPAC2R stimulate phospholipase D (PLD) activation through ADP-ribosylation factor (ARF) (Figure 1A) [217]. Moreover, the PAC1R stimulated PLD exclusively through the PAC1nR-hop1 variant [217,218]. The physiological significance of this pathway in the PACAP subfamily of receptors is yet to be established however, PLD activation has been associated with physiological responses including a respiratory burst in neutrophils, regulation of transport and endocytosis, and changes in cell morphology and motility [220,221].
Endosomal Signaling
β-arrestins can also provide a means for G protein-dissociated signaling by acting as scaffolds to facilitate multiple interactions between GPCRs and cytoplasmic signaling proteins for initiation of endosomal signaling [222]. Endosomal signaling by the PAC1R can provide distinct spatial and temporal control of cellular signaling (Figure 1B). In guinea pig cardiac neurons, PAC1R internalization and endocytosis modulate neuronal excitability via PKA/ERK activation [100,223,224,225], while PAC1R-activated endosomal ERK signaling in the amygdala regulates nociceptive hypersensitivity responses [75]. In the hypothalamus, PAC1R trafficking is involved in regulating feeding behavior and energy homeostasis [226]. In primary sympathetic neurons, the PAC1R splice variant, PAC1nR-hop1 can activate endosomal PI3k/Akt signaling cascades to facilitate neuronal survival following growth factor withdrawal [227]. While the PAC1R has been extensively documented to undergo additional signaling following receptor internalization, to date endosomal signaling has not been reported for the VPACRs [228].
2.2.5. Additional Downstream Signal Transduction Pathways
Ion Channels
As described above, cytosolic Ca2+ concentrations can be altered by Ca2+ release through transporter proteins in the endoplasmic reticulum via PLC-dependent mechanisms. Additionally, the PACAP receptor subfamily can modulate intracellular Ca2+ concentrations through the activation of ion channels. Downstream of G protein-mediated PKA and PKC activation, these kinases can modulate ionic currents to control intrinsic neuronal excitability and function following PACAP and VIP receptor activation. The PAC1R also modulates extracellular Ca2+ influx following PKC activation via voltage-gated Ca2+ channels [229,230,231,232], while PAC1R/PKA signaling can inhibit potassium (K+) channels, promoting neuronal survival [193,194]. In hippocampal nerve terminals, VPAC1R inhibits voltage-gated Ca2+ channel-dependent GABA release through Gi/o-mediated PKC activation, while VPAC2 enhances voltage-gated Ca2+ channel-dependent GABA release through Gs-mediated PKA and PKC activation [233].
In the periphery, PAC1R-stimulated PKA signaling can activate Ca2+ channels and inhibit voltage-gated potassium (K+) channels leading to insulin secretion in pancreatic β-cells [234]. In addition, PAC1R/PLC signaling in PDGFRα+ interstitial cells may be involved in mediating the inhibitory regulation of colonic contractions through small conductance Ca2+-activated K+ channels 3 [235].
Transactivation of Epidermal Growth Factor Receptor (EGFR)
Through PKA signaling, the PAC1R also transactivates EGFR, resulting in the activation of ERK signaling that promotes cell survival and proliferation [96]. In diabetic retinopathy, this pathway improves cell survival following hyperglycaemic injury [96], while in growth factor-deprived human corneal endothelial cells, it promotes wound healing and improves corneal endothelial barrier integrity [93,236]. Activation of insulin-like growth factor-1 (IGF-1) in cortical neurons transactivates PAC1R, and this mechanism was important for its anti-apoptotic activity [236]. In cortical neurons, PAC1R transactivated EGFR through PKA signaling, however EGFR transactivation only had a minor contribution to the anti-apoptotic actions of IGF-1 [236]. On the other hand, PAC1R-mediated EGFR activation may promote the proliferation of certain lung cancer cells by PKC-dependent EGFR activation [237], and VPAC1 signalling can transactivate EGFR and human epidermal growth factor receptor 2 (HER2) through PKA and Src signalling in breast cancer cells [238].
MAPK/ERK Signal Transduction
Aside from modulating ion currents, receptor-mediated activation of the protein kinases, PKA and PKC, can activate ERK signaling. ERK belongs to a family of serine/threonine kinases known as mitogen-activated protein kinases (MAPK) that are important effectors of GPCRs that regulate cellular growth, division, differentiation, and apoptosis [239,240]. All three members of the PACAP receptor subfamily can activate ERK signaling via PKA activation [200,204,241]. However, PKA and PKC-independent ERK activation also occurs following PAC1R activation in astrocytes, promoting cell proliferation [242]. Moreover, MAPK/ERK signaling can be activated through both plasma membrane and endosomal signaling pathways resulting in greater spatiotemporal diversity in cellular signalling [243]. PAC1R activation at the plasma membrane activates the cAMP/PKA and PKC signaling cascades to engage MAP kinase/ERK for neuroplasticity responses [219,244]. In pancreatic β-cells, PAC1R induces transient activation of ERK in both the nuclear and cytosolic compartments via plasma membrane signaling as well as sustained ERK activation in the cytosol, mediated through β-arrestin1 dependent receptor trafficking as described above [219,244].
2.2.6. Receptor Activity-Modifying Proteins (RAMPs)
RAMPs, consisting of RAMP1, 2, and 3, are a family of single transmembrane spanning accessory proteins that associate with many GPCRs and have the potential to modulate their function [245]. As a result of this interaction, RAMPs can act as chaperones to enhance the expression of receptors at the cell surface, alter agonist selectivity of some receptors, influence G protein coupling, and control the trafficking of receptor to endosomal compartments and recycling and degradation pathways [246,247,248].
Using a suspension bead array assay the PAC1R was shown to associate with all three RAMPs [249], albeit the functional consequences, if any, of these interactions are currently unknown. VPAC1R and VPAC2R can also interact with each member of the RAMP family, translocating all three RAMPS to the cell surface (Figure 1E) [168,250]. The physiological relevance of VPAC receptor-RAMP oligomerization remains to be determined; however, functional differences have been observed in VPACR signaling in recombinant systems [164]. The interaction between VPAC1R and RAMP2 enhanced phosphoinositide turnover compared to VPAC1R alone in response to VIP with no change in cAMP stimulation [168]. For VPAC2R, association with RAMP1 and RAMP2 enhanced basal Gi/o coupling [250]. However, it must be noted that the VPACR RAMP association can be influenced by cell background as VPAC2R did not promote RAMP trafficking to the cell surface when studied in CHO cells, but was able to promote RAMP trafficking in HEK293 cells [168]. As RAMPs are broadly, but cell specifically expressed in peripheral tissues and in the nervous system and engage with GPCRs differentially in a cell line background-dependent manner [248,251,252], hence investigating VPACR–RAMP complexes in native tissue to observe properties of ligand specificity, G protein coupling, and receptor desensitization and internalization is required to understand the physiological relevance of RAMPs for modulating the function of the PACAP receptor family.
2.3. Receptor Desensitisation and Recycling
Ligand-activated receptors can induce signaling from various intracellular compartments [253,254,255]. Endocytosis of the PACAP subfamily of receptors is initiated by the phosphorylation of serine/threonine residues in the intracellular and C-terminal receptor regions [256,257,258,259]. These serine/threonine residues serve as potential phosphorylation sites for protein kinase A (PKA), PKC, and GPCR kinases (GRKs). There are seven GRKs, of which four (GRK2, 3, 5, and 6) are ubiquitously expressed [260]. Phosphorylation by GRKs decouples G protein interaction and thus terminates initial G protein signaling. Phosphorylation of the receptor increases its affinity for β-arrestin recruitment which further facilitates uncoupling of the GPCR from G proteins and can mediate receptor endocytosis as mentioned above [224,261].
PAC1R desensitization and termination of PKC signaling are mediated by GRK3 in human retinoblastoma cells [262], whereas in gastric smooth muscle cells VPAC2R desensitization is mediated by GRK2 [215,263,264]. GRK2, 3, 5, and 6 are all recruited by VPAC1R in HEK293 cells with overexpression of these GRKs, attenuating cAMP production [265]. VPAC1R phosphorylation induces β-arrestin recruitment, however, studies in recombinant cells revealed that overexpression of β-arrestin induced only a minor decrease in cAMP production, suggesting that the mechanism of VPAC1R desensitization may be β-arrestin-independent or VPAC1R may continue to undergo cAMP signaling following β-arrestin recruitment [265,266]. Both VPAC1R and VPAC2R are internalized rapidly following agonist exposure, however, these receptors exhibit distinct trafficking profiles in recombinant systems; VPAC2R is recycled to the cell surface within two hours of receptor internalization, while VPAC1R is not [257,267].
GRK2/3 facilitates β-arrestin1 and β-arrestin2 recruitment to the PAC1R, with different functional consequences. β-arrestin2 recruitment mediates PAC1R internalization and ERK phosphorylation, while PAC1R-β-arrestin1 complexes remain localized at the plasma membrane [268]. In that same study using cortical neuronal cells, silencing of β-arrestin1 increased PAC1R-mediated ERK phosphorylation [268]. However, β-arrestin1 was required for long-lasting ERK1/2 activation by PAC1R in pancreatic islets and for ERK1/2 activation by the PAC1R hop splice variants in HEK293T cells suggesting that the effect of β-arrestin1 on signalling of the PAC1R is cell type-dependent [219,269].
2.4. Understanding of PACAP and VIP Signaling and Regulation That Needs to Be Addressed for Disease-Focused Therapies
As described above, the PACAP subfamily of receptors can activate diverse signaling pathways that lead to their pleiotropic effects throughout the body. However, the link between signaling and regulatory mechanisms and the physiological consequences of these mechanisms remain poorly understood. These receptors remain attractive targets for inflammatory and immune diseases, and disorders in the nervous system. However, there are currently no drugs on the market that specifically target PACAP/VIP receptors. While PACAP has been implicated in migraine, current efforts to antagonize PACAP-PAC1R signaling through the development of monoclonal antibodies to PACAP38 or PAC1R for treatment-resistant migraines [85,270,271,272] have not been successful. In phase II clinical trials, the PAC1R monoclonal antibody, AMG301 failed to exhibit efficacy for migraine treatment, despite promising results in preclinical studies [273]. While multiple factors could contribute to this clinical outcome, such as failure to reach the target site of action and inability to block specific signaling pathways associated with the disease, the pharmacological complexities described above including the existence of multiple PAC1R splice isoforms, non-selective PACAP-VPAC receptor signaling and endosomal signaling, present challenges for developing novel therapies. Furthermore, there is increasing evidence that, in addition to forming hetero-oligomers with RAMPs, the PACAP subfamily of receptors can form dimers with themselves and other receptors [172,274,275]. The higher order oligomerization of these receptors may have significant effects on their function. If validated in vivo, this could provide novel therapeutic design strategies. Overall, a lack of understanding of the importance of these factors may impact the therapeutic outcomes of drugs targeting this receptor family. A more detailed investigation into these complexities, including cell and disease-specific expression of receptor splice variants and subtypes, oligomeric complexes, and key signal transduction/regulatory proteins will likely be required for successful clinical translation [168,207,213,214,250,276,277].
Beyond the intricacies of structural variations and potential higher order oligomeric complexes, the ability of the PACAP subfamily to activate multiple downstream signaling pathways enables the potential identification of “biased agonists” that selectively control signaling and thus initiate specific cellular and physiological outcomes. Biased agonism refers to the ability of individual ligands acting at the same receptor to produce distinct profiles of downstream signaling [166,278]. This is of particular therapeutic interest as it means different ligands binding to the same GPCR may preferentially activate beneficial pathways while minimizing those that may result in unwanted side effects [279]. Biased agonism has been observed at various Class B GPCRs, the most extensively studied being the GLP-1R. Through coupling to distinct G proteins (Gs, Gq, Gi.o) and to β-arrestins, GLP-1R is able to activate a plethora of important signaling pathways in pancreatic β-cells including cAMP production, ERK1/2 phosphorylation, and intracellular Ca2+ mobilization [280,281]. One of the earliest reports of biased agonism was at the PAC1R, where both the PACAP27 and PACAP38 stimulated cAMP production in the PAC1nR receptor isoform, while only PACAP38 stimulated IP3 production [178]. However, later studies have not necessarily replicated these findings, highlighting the complexities in PACAP subfamily signaling that may influence pharmacological outcomes and mask biased agonism [181,182].
Moreover, pharmacological probes, such as PACAP and VIP, used to study the PACAP/VIP receptors often exhibit some degree of affinity for multiple members within the subfamily. The heterogeneous distribution of these receptors in the body also complicates the attribution of specific receptors of the subfamily to a pharmacological effect, and consequently the physiological outcomes. For example, receptor cross-talk can mask the presence of receptor synergism or antagonism that may potentially occur. Hence, in addition to the therapeutic benefit of next-generation receptor-selective ligands, there is also the potential gain from developing more specific probes for future pharmacological studies.
3. Molecular Activation of PACAP and VIP Receptors
A better molecular understanding of the mechanisms behind ligand recognition and receptor activation of the PACAP receptor subfamily can guide the development of more effective clinical candidates and probes. GPCRs share the structural composition of an N-terminal extracellular domain (ECD), seven transmembrane (TM) helices embedded in the lipid bilayer membrane, three extracellular and three intracellular loops connecting the TM helices, and a C-terminal intracellular domain. The class B1 GPCR subfamily, including the VPAC1R, VPAC2R, and PAC1R, are characterized by an N-terminal ECD consisting of approximately 120 amino acid residues and three conserved disulphide bonds that form the core of the ECD structure [282]. The ECD is important for peptide recognition and binding to position the peptide optimally for interaction with the TM core, and subsequent receptor activation [135].
The recent advances in structural biology, particularly the emergence of cryo-electron microscopy (cryo-EM) for GPCR structure determination, have improved our molecular understanding of ligand-occupied GPCRs coupled with their canonical G protein transducer [282]. Breakthroughs in protein complex stabilization strategies, including the use of nanobodies, single-chain fragment antibodies and G protein modifications, such as dominant-negative mutation (DN), mini Gα proteins, and nanoBiT tethering have facilitated the formation and purification of receptor-G protein complexes suitable for structure determination [136,283,284,285,286]. This has been complemented by significant improvements in cryo-EM grid preparation, microscopy, and detector hardware, imaging strategies, and data processing for the three-dimensional visualization of GPCR complexes [282,287,288,289,290,291] and non-GPCRs [292,293].
Before the determination of the full-length PACAP38-bound PAC1R–Gs structure by cryo-EM, there had been some uncertainty on the binding mode of peptides to the N-terminal ECD of the PACAP subfamily of receptors [135]. The crystal structures of the isolated ECD of PAC1sR (PDB: 3N94) and VPAC2R (PDB: 2X57) (Table 5) demonstrated a similar ECD configuration to the crystal structures of other class B1 GPCRs, including the glucose-dependent insulinotropic polypeptide receptor (GIPR) PDB: 2QKH [294] and GLP-1R PDB: 3C5T [295]).
Table 5.
List of structures of the ECD region of the PACAP subfamily of receptors determined through X-ray crystallography and NMR.
In these structures, three conserved disulphide bonds facilitate folding of the ECD in a sandwich-like configuration consisting of an N-terminal α-helix, two antiparallel β-sheets (β1–β2) and (β3–β4), and a C-terminal α-helix. The first disulphide bond links the N-terminal α-helix and β1–β2 sheet, the second the two β-sheets while the final bond between C77ECD and C113ECD connects the β3–β4 sheet to the C-terminal helix. The peptide-binding clefts in these ECD structures resides between the two central anti-parallel β-sheets, consistent with other class B1 GPCRs ECD structures [297]. In contrast, the NMR structure of PACAP(6–38) complexed to PAC1sR (PDB: 2JOD) exhibited differences to the crystal structure in the topology of the area between β3 and β4 and a distinct peptide-binding mode compared to other class B1 GPCRs [298]. While the first full-length active cryo-EM structure of the PACAP receptor subfamily, the PACAP38-bound PAC1R complex (PDB: 6P9Y), had limited resolution in the ECD, this structure, and subsequently structures of VPAC1R and VPAC2R, confirmed that the peptide binding mode of the PACAP/VIP receptors is similar to other class B1 GPCRs (Figure 3), and the binding mode observed in the crystal structure of the PAC1R ECD [135].
3.1. Structural Characteristics of the PACAP Subfamily of Receptors
Structures of PAC1, VPAC1, and VPAC2 receptors determined by cryo-EM are consistent with the proposed class B1 GPCR two-domain activation model, where the C-terminal portion of the peptide agonist binds to the peptide-binding cleft within the ECD [135,299], while the N-terminal helix of the peptide interacts with the receptor core. The N-terminal ECD of the receptor forms a horseshoe-like configuration around the peptide, extending towards the receptor core to form interactions with the top of ECL1 [135,136,137,300,301,302,303,304,305,306,307,308,309].

Figure 3.
Comparisons of the binding mode of representative peptides bound to Class B1 GPCRs reveal a shared peptide binding mode where the N-terminus of the peptide interacts with the transmembrane (TM) core and the C-terminus of the peptide interacts with the N-terminal extracellular domain (ECD) and extracellular loops (ECLs) of the receptor [135,136,137,300,301,302,303,304,305,306,307,308,309]. The highest variability of the structures is in the extracellular domains and peptide C-termini (as displayed in the structure overlay in the first panel. Structures were aligned by receptor chains and displayed as ribbons (licorice style) using Chimera X 1.3 [310,311]. G protein subunits are not displayed for clarity. The following models lack extracellular domains due to low resolution: PDB 6VN7, 6P9Y, 7D68, and 6P9X.

Due to the greater flexibility and mobility of the ECD regions, ICLs and ECLs, these regions remain relatively less well-resolved compared to the TM domains. Details of the agonist, global resolution, and receptor and G protein modifications of each of the structures of the PACAP subfamily of receptors determined by cryo-EM are listed below (Table 6). In addition to the differing resolutions achieved in the structures, some of the domains/loops are missing in the PDB models due to the limited EM-map resolution in these regions. Namely, for the PACAP27-bound VPAC2R (PDB: 6VN7) and PACAP38-bound PAC1nR (PDB: 6P9Y) complexed with DNGαs structures, the N-terminal ECD, ECL1, and ICL3 are not modeled in the final deposited PDB file. While for the PACAP38-bound PAC1nR complexed with mini-Gαs structure (PDB: 6LPB), ICL1, ECL1 and ICL3 are absent. For the PACAP38- (PDB: 6M1I) and maxadilan-bound (PDB: 6M1H) PAC1sR and PAC27-bound VPAC2R (7VQX) structures, ICL3 is not modeled in the PDB files.
Table 6.
Cryo-EM structures of active, peptide-bound, Gs protein-coupled, PAC1R, VPAC1R, and VPAC2R as of June 2022. The PAC1R isoforms solved include PAC1R-null (PAC1nR) with the full-length ECD and PAC1R-short (PAC1sR) with the truncated N-terminal ECD.
Thus far, the published cryo-EM structures have been solved with either PACAP isoforms or maxadilan. Hence the binding characteristics of these are described below.
3.2. Characteristics of the PACAP-Bound VPAC1R, VPAC2R, and PAC1R Structures
Of the six structures determined by cryo-EM to date, five were PACAP-bound. The three PACAP-PAC1R structures (PDB: 6P9Y, 6LPB, and 6M1I) share an agreement on the PACAP binding mode (Figure 4A,B). Of these structures, two are of the receptor splice variant with the full-length ECD, PAC1nR [131,135], while the other is of PAC1sR, missing twenty residues (Val89ECD–Ser109ECD) in the N-terminal ECD [134]. While this region of the ECD is critical for the binding affinity of VIP, with an almost hundred-fold improvement in binding affinity observed for PAC1sR compared to PAC1nR, the two receptor variants share similar PACAP binding affinities [175]. The additional residues in the ECD that are present in the PAC1nR splice variant have not been resolved in any of the cryo-EM structures to date, most likely due to high flexibility.
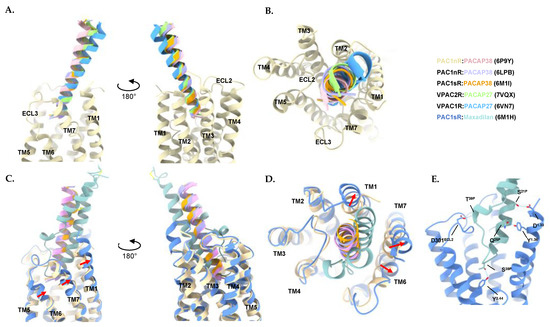
Figure 4.
Comparison of peptide binding mode to receptors of the PACAP subfamily. (A) Side views and (B) Top view of the superimposed structures of PACAP bound to VPAC1R, VPAC2R, and PAC1R. Only the transmembrane (TM) helices of the receptor chain of PAC1nR (PDB: 6P9Y) are shown for reference/clarity. (C) Side views and (D) Top view of the superimposed structures of maxadilan and PACAP bound to PAC1R reveals TM1, 6, and 7 shifted outward to accommodate maxadilan (PDB: 6M1H) when compared to the PACAP38-bound PAC1nR (PDB: 6P9Y). Movement of the TMs indicated by red arrows. Only the TM helices of the receptor chain of the PACAP38-bound PAC1nR (PDB: 6P9Y) and the TM helices of the receptor chain for the maxadilan-bound PAC1sR (PDB: 6M1H) are shown for reference/clarity. (E) Helix 1 of maxadilan forms hydrogen bonds with TM1 in the peptide binding pocket while the loop of maxadilan and helix 2 forms hydrogen bonds with ECL2 and TM3. Structure is shown as a side view with a focus on the receptor core. Hydrogen bonds calculated with ChimeraX 1.3. Structures were aligned by receptor chains and displayed as ribbons, with residues involved in hydrogen bonds (dotted lines) shown as sticks using ChimeraX 1.3 [310,311]. Colors for the peptides and receptors are shown in the figure. Peptide residues are denoted with superscript P, extracellular loop residues are denoted as ECL. Receptor transmembrane residues are denoted using the Wootten numbering system.
Comparison between the PACAP-bound PAC1R, VPAC1R, and VPAC2R structures indicates that the N-terminus of the peptide is inserted into the receptor core at a similar orientation and angle for each receptor. By contrast, greater differences are observed in the C-terminal half of PACAP interacting with the ECL1, the extracellular tip of TM1, and the ECD (Figure 4A,B).
While the PAC1R structures were determined bound to PACAP38, the VPAC1R and VPAC2R were bound to PACAP27. However, due to the high mobility of the PACAP C-terminus in PACAP38-bound structures, only the first 27 residues were modeled. When comparing these structures, the interactions of the first three residues of the relevant PACAP with the receptor core of the VPAC1R, VPAC2R, and PAC1R (Figure 5A–C), from studies of alanine substituted peptides, the N-terminal His1P (superscript is the residue number of the peptide) and Asp3P residues are crucial for binding affinity and biological activity of the peptide for all three receptor subtypes [142,312,313]. This is reflected in the extensive, conserved, hydrophobic contacts between His1P and the receptor TM3 and TM5 and the conserved salt bridge between Asp3P and Arg2.60 (the superscript number refers to the Wootten et al. [314], class B numbering system) that is conserved across all receptors of the PACAP subfamily (Figure 5). At the N-terminal end of PACAP, His1P forms extensive hydrophobic interactions in the TM bundle with TM3 (Val3.40, Val3.13) and TM5 (Trp5.36) of all three receptors. However, subtle differences in interactions are observed where His1P also forms a hydrogen bond with Gln3.37 of TM3 in VPAC2R, while no hydrogen bond was observed for the equivalent His3.37 residue in PAC1R (Figure 5B). Asp3P forms a salt bridge with Arg2.60 and hydrophobic contacts with TM7 (Leu7.43) that are shared in all three receptors (Figure 5C). The residues at position 1 and position 3 in PACAP, His1P, and Asp3P, respectively are highly conserved in class B1 GPCR peptides. On the other hand, at position 2, Ser2P of PACAP forms a hydrogen bond with Tyr3.44 lower down (from the extracellular face of the receptor) on PAC1R TM3 resulting in a deeper insertion of the peptide into the PAC1R core than observed with the VPACRs (Figure 5B). For VPAC2R, Ser2P forms a hydrogen bond with TM7 (Glu7.42) that is not observed in VPAC1R nor PAC1R (Figure 5B).
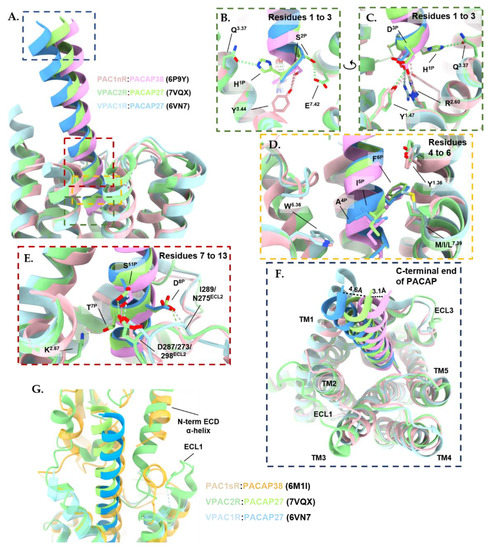
Figure 5.
Comparison of PACAP in the binding pocket of VPAC1R (blue; PDB: 6VN7), VPAC2R (green; PDB: 7VQX) and PAC1nR (pink; PDB: 6P9Y). Structures were aligned by receptor chains and displayed as ribbons, with residues involved in hydrogen bonds (dotted lines) and hydrophobic interactions shown as a stick; calculated with ChimeraX 1.3 [310,311]. Extracellular domain (ECD) residues of the receptor were removed for clarity. Peptide residues are denoted with superscript P, extracellular loop residues are denoted as ECL. Receptor transmembrane (TM) residues are denoted using the Wootten numbering system. (A) Overview of comparison highlighting the regions of higher magnification displayed in panels (B–E) with matching colored dashed rectangles. (B,C) Close-up view of the hydrogen bonds formed between the first three residues of PACAP and the binding pocket of VPAC1R (blue), VPAC2R (green), and PAC1nR (pink). (D) Close-up view (front and back side view) of PACAP residues 4 to 6 that interact with conserved hydrophobic side chains in TM1 (Y1.36), TM5 (W5.36) and TM7 (M/I/L7.39) of the peptide binding pocket of VPAC1R (blue), VPAC2R (green) and PAC1nR (pink). (E) Close-up view of hydrogen bonds formed between PACAP from residue 7 to 13 and the binding pocket of VPAC1R (blue), VPAC2R (green), and PAC1nR (pink). (F) Close-up view of the C-terminal end of PACAP peptide that is shifted 4.6 Å towards the receptor core in VPAC2R compared to VPAC1R, while the C-terminal end of PACAP peptide is shifted 3.1 Å towards ECL3 in PAC1R compared to VPAC2R. Distance between the α-carbon of Leu27P in PACAP peptide between the three structures calculated with ChimeraX 1.3. (G) The N-terminal α-helix of the VPAC2R ECD (green; PDB: 7VQX) adopts a unique conformation that deeply inserts into a cleft between PACAP27 and the predicted position of ECL1 in the model compared to the modeled ECD in the PACAP38-bound PAC1sR (yellow; PDB: 6M1I). Due to limited resolution in the N-terminal ECD and ECL1, these regions are not modeled in the PACAP27-bound VPAC1R structure (blue; PDB: 6VN7).
The PACAP residues Gly4P, Ile5P, and Phe6P make extensive interactions with the hydrophobic pocket residues of TM1 and TM7 (Figure 5D). Many of these interactions are conserved between the three receptors. Specifically, Ile5P makes hydrophobic contacts with the equivalent residue at 7.39 in all three receptors, while the aromatic residue of Phe6P interacts with Tyr1.36, Val1.39 and Tyr1.43 residues in TM1 and Leu7.43 in TM7 (Figure 5D). Phe6P of PACAP is crucial for binding affinity and biological activity at all three PACAP receptor subtypes as reflected by the loss of biological activity with alanine substitution [142,312,313,315]. However, the replacement of Phe6P with bulky hydrophobic groups modulates the specificity of PACAP analogues for different receptors to some extent [312,313,316,317,318]. For example, the substitution of Phe6P in PACAP with an amino acid with two aromatic rings in the case of [4,4′-biphenylalanine6]-PACAP27 resulted in an approximately 10-fold reduction in VPAC1R -mediated Ca2+ mobilization potency compared to PACAP27, whilst retaining similar potency at the PAC1R [312]. On the other hand, substitution with the two aromatic ring amino acids, 1-naphthylalanine6P resulted in a 3-fold reduction in potency in Ca2+ mobilization at the PAC1R and VPAC1R, and a 10-fold reduction in potency at the VPAC2R, compared with PACAP27 [312]. Gly4P is in closer proximity to Trp5.36 of TM5 in PAC1R compared to the VPACRs (Figure 5D). In VIP, the equivalent residue at position 4 is an alanine and at position 5 is a valine. Due to steric hindrance, the additional methyl group in alanine and the different steric profile of the branched alkyl side of valine compared with glycine and isoleucine, respectively, may restrict the binding of VIP to PAC1R, which may contribute to the selectivity of VIP for the VPAC receptors [318,319]. The substitution of PACAP residues Gly4P and Ile5P with Ala4P and Val5P of VIP reduced PACAP affinity for PAC1R when tested in guinea pigs and the rat brain [320,321], suggesting these residues are important for receptor selectivity.
In the cryo-EM structures, high-resolution features are observed for the TM regions, suggesting the peptide bound-TM core is relatively stable, hence side-chain rotamers could be accurately modeled [135,136,137,138,139]. In contrast, the ECLs and ECD are relatively mobile, hence these regions are not as well resolved in the cryo-EM maps, limiting the accuracy of the modeling for side chain rotamers in these regions. Nonetheless, ECL2 is better resolved than ECL1 and ECL3 in many of these structures. Ser11P contributes to polar interactions with ECL2 (with Asp287ECL2, Asp273ECL2, and Asp298ECL2 of VPAC1R, VPAC2R, and PAC1R, respectively) for all three receptors (Figure 5E). In contrast, the interaction between Asp8P of PACAP with the ECL2 residues Ile289ECL2 and Asn275ECL2 of VPAC1R is not observed in PAC1R. On the other hand, Lys2.67 in VPAC2R forms a hydrogen bond with Thr7P in VPAC2R, while this interaction is not observed in PAC1R and VPAC1R (Figure 5E) [135,136,137].
The C-terminal end of the peptide of the VPAC2-bound PACAP27 was shifted by 4.6Å toward the TMD core compared to VPAC1R-bound PACAP27 when comparing the α-carbon atom of Leu27P (Figure 5F). In comparison, the C terminus of PACAP27-bound VPAC2R moved towards ECL1 by 3.1 Å when compared with residue Leu27P of the PACAP38-bound PAC1nR (PDB: 6P9Y) (Figure 5G) [135,136,137]. The PACAP38-bound VPAC1R (PDB: 6VN7) and PACAP38-bound PAC1nR (PDB: 6P9Y) structures did not include modeling of ECL1 and the ECD due to limited resolution in these regions, however, Xu, et al., 2022 reported that the N-terminal α-helix of the VPAC2R ECD adopts a unique conformation, relative to other class B1 GPCRs, and compared to the modeled ECD in the PACAP38-bound PAC1sR (PDB: 6M1I). In VPAC2R, the N-terminal α-helix inserts deeply into a cleft between PACAP27 and the predicted, more open, position of ECL1 in the model, stabilizing the peptide-receptor interface (Figure 5G) [137]. Nonetheless, the limited resolution in the N-terminal ECD and ECLs and the accuracy of modeling rotamers in this region must be taken into consideration when interpreting interactions between the peptide and the receptor ECD and ECL1.
3.3. Characteristics of the Maxadilan-Bound PAC1R Structure
The PAC1R-selective agonist maxadilan is a 61-amino acid peptide that consists of two α-helices (helix 1 and helix 2) that form a v-shaped conformation held together by a disulphide bond between Cys14P–Cys51P [133,138,322,323]. Maxadilan binds to the same binding pocket as PACAP on PAC1R (Figure 4C). However, to accommodate the larger 3D conformation of maxadilan, TM1, 6 and 7 of PAC1R are shifted outward compared to the PACAP38-bound structures (Figure 4C,D) [138]. Helix 1 of maxadilan interacts with TM1 of PAC1sR on one side of the receptor, with the residues Ser21P and Gln25P of helix 1 of maxadilan forming hydrogen bonds with Asp1471.33 and Tyr1501.36 of TM1. On the other side of the receptor binding pocket, the loop of maxadilan and helix 2 forms hydrogen bonds with TM3 (Ser33P with Tyr2413.44) and ECL2 (Thr39P-Asp301ECL2) (Figure 4E). At the receptor core, the loop of maxadilan is reported to form hydrophobic interactions with TM6 (Phe34P with Phe3696.56 and Ala3706.57) and TM7 (Ala32P with Leu3867.43) [138].
Truncated forms of maxadilan, max.D.4 (missing amino acids 24–42) and M65 (missing amino acids 25–41) act as PAC1R-specific antagonists [133,148,324]. Both max.D.4 and M65 have residues in helix 1 and the loop of maxadilan removed. These regions of maxadilan interact with the binding site in the maxadilan-bound PAC1sR structure, suggesting their importance for agonist activity. Namely, the hydrogen bonds formed between helix 1 of maxadilan and TM1 (Gln25P-Tyr1501.36) and between the loop of maxadilan and ECL2 (Ser33P-Tyr2413.44 and Thr39P-Asp301ECL2) as well as hydrophobic contacts between Ala32P and Leu3867.43 of TM7 and the amide-Pi stack interactions between Phe34P and TM6 (Phe3696.56 and Ala3706.57) would be predicted to be lost for the antagonist maxadilan analogues [138].
3.4. Activation Characteristics of the PACAP Subfamily of Receptors
While the PACAP receptors can potentially couple to Gq/11 and Gi/o proteins and recruit β-arrestins for the activation of downstream signaling [214,215,268], currently all active structures of the PACAP receptor subfamily are in complex with Gs. Hence, the elucidation of structures of these receptors with non-Gs transducer proteins will be important in understanding the molecular mechanisms behind the diversity in transducer signaling. The Gs-coupled, active structures of the PACAP receptor subfamily members display similar characteristics to other active class B1 GPCRs [135,282,299,325]. As there are currently no inactive state structures of PACAP subfamily receptors, activation mechanisms are inferred using the inactive structures of other class B GPCRs and are generally limited to global activation mechanisms observed for class B1 GPCRs.
Deep engagement of peptide agonists with the TM bundle of class B1 GPCRs above a central polar network is proposed to promote receptor activation, with two additional layers of conserved polar networks within the TM bundle involved in this process. For the PACAP family receptors, the central conserved polar network (top layer) is formed by Arg2.60, Asn3.43, His6.52 and Gln7.49 [282,314,325,326,327], the middle being the HETx polar network consisting of residues His2.50, Glu3.50, Thr6.42 and Tyr7.57 [282,325], and the bottom at the cytoplasmic face, consisting of the TM2-6-7-8 network formed by Arg2.46, Arg6.37, Asn7.61, Glu8.41 [282,325] (Figure 6).
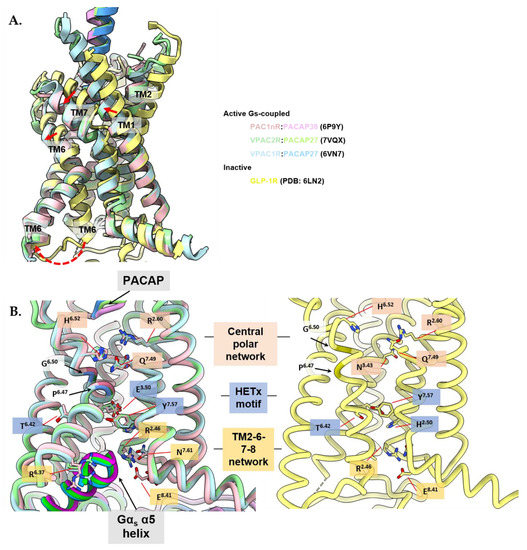
Figure 6.
The active receptor conformation is similar between members of the PACAP subfamily of receptors. Active, Gs-coupled PACAP-bound receptors are displayed: VPAC1R (blue; PDB: 6VN7), VPAC2R (green; PDB: 7VQX), and PAC1nR (pink; PDB: 6P9Y) and compared to the inactive crystal structure of full length human GLP1 receptor (GLP1-R) in complex with Fab fragment (Fab7F38) (yellow; PDB: 6LN2). Structures were aligned by receptor chains and displayed as ribbons as side views, with important residues shown as sticks using ChimeraX 1.3 [310,311]. Gαs-α5 helix colored in darker colors corresponding to the respective receptor chains (B, left panel). The Fab fragment in inactive GLP1-R was omitted for clarity. (A) Comparison between active structures of the PACAP subfamily of receptors and the full-length inactive structure of GLP1-R indicate inward movement of TM1 and outward movement of TM6 and 7 upon receptor activation with TM6 undergoing a kink at the Pro6.47xxGly6.50 motif. Movement of TMs indicated by red arrows. The G protein was omitted for clarity. (B) Comparison between conserved residues in the central polar network (pink shading-top), HETx motif (blue shading-middle) and TM2-6-7-8 network (yellow shading-bottom) between the active PACAP subfamily of receptor structures (left panel) and the inactive GLP1-R structure (right panel) indicate rearrangement of the conserved polar networks for TM6 to kink at the Pro6.47xxGly6.50 motif and accommodate the α5 helix of Gαs at the cytoplasmic face.
The binding of peptide agonists leads to reorganization of the ECLs and outward movement of the TM6/ECL3/TM7 and inward movement of TM1 (Figure 6A) [282,325,328,329,330,331]. The peptide and G protein binding pockets are allosterically connected, hence both facilitate the process of receptor activation [332]. Interactions in the central polar network are reorganized by peptide and G protein engagement, inducing the unraveling of TM6 around the Pro6.47xxGly6.50 motif, resulting in TM6 undergoing an approximate 90° kink [325,333]. Gln7.49 from the central polar network reorganizes to interact with Tyr6.53, and Asn5.50 forms hydrogen bonds with Leu3.47, while residues Phe5.54, Ile5.57 and Ile5.58 form extensive hydrophobic contacts with the main chains of TM3 and TM6 that stabilize the active conformation [136,282,334]. The activated conformation(s) is further stabilized through coordinated water networks across the TM domain [300,335,336].
The outward movement of the intracellular half of TM6 and a bend of the extracellular portion of TM7 towards TM6 results in rearrangement in the lower polar networks (Figure 6B) [282,325]. The destabilization of the HETx motif (His2.50, Glu3.50, Thr6.42 and Tyr7.57) breaks ground state interactions between Thr6.42 and Tyr7.57, and the Arg6.40–Glu8.41 interaction is lost [282]. Intracellular hydrogen bonds in the TM2-6-7-helix 8 polar network that are present in inactive state class B1 GPCR structures are broken in the peptide and G protein bound PAC1R, VPAC1R and VPA2R structures, enabling the insertion of the C-terminal α5-helix of the Gαs subunit into the core of the receptors, where the α5 helix of Gs forms conserved interactions with TM2, TM3, and ICL2 enabling G protein activation [282,325].
3.5. Allosteric Modulation
Due to the high degree of conservation in the TM bundle of different PACAP subfamily receptors where the peptide agonist activation domains bind, the identification of allosteric ligands may be an additional strategy that could be used for developing more selective therapeutics [337]. Allosteric ligands bind to a spatially distinct binding site than the endogenous agonists and therefore have the potential to modulate the receptor response to these agonists. Since allosteric sites are likely to be less evolutionarily conserved compared to the natural orthosteric sites, targeting an allosteric site could engender receptor selectivity or alter the signaling/regulatory profile of peptide-bound receptors [338].
Allosteric ligands resulting in positive effects on the affinity and/or efficacy of the orthosteric ligand response are known as positive allosteric modulators (PAMs), while allosteric ligands resulting in negative effects on the orthosteric ligands are known as negative allosteric modulators (NAMs) [338]. Only limited studies have attempted to discover allosteric ligands within the PACAP subfamily of receptors, however, this may be an avenue to explore for future drug design.
From in silico screening of the ZINC15 drug library, ticagrelor was proposed as a potential allosteric modulator for the PACAP subfamily of receptors. Ticagrelor, an approved antiplatelet medicine, was proposed as a NAM for inhibiting VIP-induced calcium mobilization in CHO cells expressing either VPAC1R or VPAC2R, with weak selectivity towards the VPAC2R [339]. Using molecular dynamics simulations with an inactive homology model of VPAC1R that was generated based on the GLP1-R crystal structure (PDB: 5VEW) [340,341], the allosteric ticagrelor binding site was predicted to be located in the intracellular region of the TM bundle, in a pocket formed by Arg6.37, Arg6.40, Arg2.46 and Asn8.47, and therefore proposed to prevent conformational changes in the region of the tyrosine Thr6.42-Tyr7.57 toggle switch required for VPAC1 and VPAC2 receptor activation [339]. The TM helical bundle of class B1 GPCRs is a region that has attracted much attention in allosteric drug design as modulation of the TMs, in particular TM6, could be targeted to stabilize or restrict an active conformational receptor state [337]. NNC0640 and MK-0893 are NAMs that bind to TM6 in the inactive glucagon receptor (GCGR) crystal structures (PDB: 5XEZ and 5EE7 respectively), restricting the outward movement of the TM6 intracellular domain [329,342].
Positive allosteric modulators of the PAC1R have also been reported with doxycycline and minocycline proposed to enhance PACAP binding to PAC1R by binding to a site in the N-terminal ECD [343]. Doxycycline and minocycline belong to the antimicrobial class of tetracyclines and promote the expression of the plasminogen activators: tissue plasminogen activator (tPA) and urokinase plasminogen activator (uPA) via PAC1R activation in RT4 Schwann cells and augment PACAP27-induced cAMP stimulation in CHO cells overexpressing PAC1R [344]. While the 3D configuration of the N-terminal ECD is conserved across class B1 GPCRs and within the PACAP receptor subfamily, the amino acid composition is relatively diverse [337]. Hence, targeting regions of the ECD provides opportunities for the development of subtype-selective PACAP receptor allosteric modulators. This may be particularly true for VPAC2R, where the PACAP27-bound VPAC2R Gs structure (PDB: 7VQX) demonstrated that the N-terminal α-helix of the ECD adopts a unique conformation by inserting into a cleft between the peptide and ECL1 to stabilize the peptide-receptor interface [137].
4. Concluding Remarks
Historically, the predominant strategy for the discovery of clinical candidates for the PACAP receptor subfamily has been based on structural modifications to endogenous peptides. Over the last decade, there has been considerable progress in the determination of the three-dimensional structures of these receptors using cryo-EM, in complex with agonists and G proteins, for the identification of key regions for peptide recognition and selectivity, which complements earlier X-ray crystallography on receptor subdomains. These recent structures advance our understanding of previously reported peptide and receptor structural-activity relationships from empirical drug development and provide insight into key regions of the peptide binding site to enable strategies for rational, structure-guided peptide design.
While the ECD, where the peptide C-terminus initially recognizes class B1 GPCRs, remains less well-resolved than other regions, the resolution of structures determined through cryo-EM has drastically improved over the past five years through advances in sample preparation, data collection, and data processing, and is predicted to continue improving with sub-2.5 Å cryo-EM structure determination being increasingly routine for class B1 GPCR–G protein complexes [288]. Moreover, direct information on 3D conformational dynamics captured during vitrification can increasingly be extracted through sophisticated analytical methods [304,345] and such dynamics play critical roles in peptide recognition and modes of receptor activation that may not be apparent from high-resolution consensus reconstructions [304,332,346,347]. This information can be integrated with other methods for interrogating GPCR dynamics, including hydrogen deuterium exchange mass spectrometry (HDX-MS) [348], single-molecule FRET studies [349] and electron paramagnetic resonance (EPR) [350,351] and holds promise for further enhancing peptide drug design [348,352]. This emerging knowledge on the different conformational states and structural features responsible for activation transition from inactive and apo-state GPCRs could be utilized in guiding the design of peptides of distinct pharmacology and selectivity for different receptors within the PACAP subfamily as well as identifying allosteric binding sites for new compounds [348,353].
The next question is: how do we use this information to develop better peptide drugs and selectively target signaling pathways activated by PACAP receptors for specific physiological outcomes? As described above, the PACAP subfamily of receptors can activate diverse signaling pathways leading to their pleiotropic effects throughout the body and are attractive targets for inflammatory and immune diseases, and disorders in the nervous system. To coordinate the diverse physiological outcomes mediated by the activation of the PACAP subfamily of receptors, these receptors activate multiple signaling pathways through the coupling of different G proteins (e.g., Gs, Gq/11, and Gi/o proteins) as well as through associating with β-arrestin and RAMPs to modulate receptor signaling and trafficking [168,178,192,207,250,268]. Additionally, splice variants within the PAC1R, as well as the potential for biased agonism through these receptor variants, may contribute to differential transducer coupling and downstream signaling, encoding for functional selectivity [178,180].
Biased peptide agonists, which have been broadly observed across class B1 GPCRs, have therapeutic potential through coupling distinct profiles of transducers that produce beneficial clinical effects, while reducing undesirable side effects [166,280,354]. All PACAP subfamily receptor structures determined to date have been Gαs protein-coupled with modifications that stabilize the receptor G protein complex. Hence future structural studies with non-Gs transducers will determine whether signaling bias can be structurally explained for the rational design of biased peptides for PACAP receptor subtypes. Furthermore, short half-lives, lack of oral bioavailability, and difficulty in penetrating the blood–brain barrier are challenges observed with many peptides. As many of the therapeutic targets of the PACAP receptor subfamily are located in the CNS, the engineering of peptides that can selectively bind to the PACAP receptors and cross the blood–brain barrier remains a major challenge.
Author Contributions
J.L., writing—original draft preparation; All authors, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by an Australian Government Research Training Program (RTP) Scholarship through Monash University (J.L.). P.M.S. is an NHMRC Senior Principal Research Fellow (#1154434) and Director of the ARC Industrial Transformation Training Centre for Cryo-electron Microscopy of Membrane Proteins (CCeMMP; #IC200100052). D.W. is an NHMRC Senior Research Fellow (#1155302) and Monash Node Leader for CCeMMP. P.Z. is an ARC Future fellow (#FT200100218). This work was partly funded by NIH grant (#GM132095), ARC grant (#DP210101504) and NHMRC grants (#1150083 & #1159006).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Schematic illustrations of PACAP receptor subfamily signaling in Figure 1 and PAC1R alternative splicing in Figure 2 were created with BioRender.com (accessed on 29 June 2022). Molecular graphics and analyses were performed with UCSF ChimeraX, developed by the Resource for Biocomputing, Visualization, and Informatics at the University of California, San Francisco, with support from the National Institutes of Health R01-GM129325 and the Office of Cyber Infrastructure and Computational Biology, National Institute of Allergy and Infectious Diseases [310,311].
Conflicts of Interest
P.M.S. and D.W. are shareholders in Septerna Inc. All other authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Mutt, V.; Said, S.I. Structure of the porcine vasoactive intestinal octacosapeptide: The amino-acid sequence. Use of kallikrein in its determination. Eur. J. Biochem. 1974, 42, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Brenneman, D.E.; Eiden, L.E. Vasoactive intestinal peptide and electrical activity influence neuronal survival. Proc. Natl. Acad. Sci. USA 1986, 83, 1159–1162. [Google Scholar] [CrossRef] [PubMed]
- Sorg, O.; Magistretti, P.J. Vasoactive intestinal peptide and noradrenaline exert long-term control on glycogen levels in astrocytes: Blockade by protein synthesis inhibition. J. Neurosci. 1992, 12, 4923–4931. [Google Scholar] [CrossRef] [PubMed]
- Said, S.I.; Mutt, V. Isolation from Porcine-Intestinal Wall of a Vasoactive Octacosapeptide Related to Secretin and to Glucagon. Eur. J. Biochem. 1972, 28, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Fahrenkrug, J. Transmitter Role of Vasoactive Intestinal Peptide. Pharmacol. Toxicol. 1993, 72, 354–363. [Google Scholar] [CrossRef]
- Behar, J.; Guenard, V.; Walsh, J.H.; Biancani, P. VIP and acetylcholine: Neurotransmitters in esophageal circular smooth muscle. Am. J. Physiol. 1989, 257, G380–G385. [Google Scholar] [CrossRef]
- Lundberg, J.M.; Anggård, A.; Fahrenkrug, J.; Hökfelt, T.; Mutt, V. Vasoactive intestinal polypeptide in cholinergic neurons of exocrine glands: Functional significance of coexisting transmitters for vasodilation and secretion. Proc. Natl. Acad. Sci. USA 1980, 77, 1651–1655. [Google Scholar] [CrossRef]
- Matsushita, N.; Kato, Y.; Shimatsu, A.; Katakami, H.; Yanaihara, N.; Imura, H. Effects of VIP, TRH, GABA and dopamine on prolactin release from superfused rat anterior pituitary cells. Life Sci. 1983, 32, 1263–1269. [Google Scholar] [CrossRef]
- Malhotra, R.K.; Wakade, A.R. Vasoactive intestinal polypeptide stimulates the secretion of catecholamines from the rat adrenal gland. J. Physiol. 1987, 388, 285–294. [Google Scholar] [CrossRef]
- Ganea, D. Regulatory effects of vasoactive intestinal peptide on cytokine production in central and peripheral lymphoid organs. Adv. Neuroimmunol. 1996, 6, 61–74. [Google Scholar] [CrossRef]
- Coupar, I.M. Stimulation of sodium and water secretion without inhibition of glucose absorption in the rat jejunum by vasoactive intestinal peptide (VIP). Clin. Exp. Pharmacol. Physiol. 1976, 3, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Racusen, L.C.; Binder, H.J. Alteration of large intestinal electrolyte transport by vasoactive intestinal polypeptide in the rat. Gastroenterology 1977, 73, 790–796. [Google Scholar] [CrossRef]
- Steingart, R.A.; Solomon, B.; Brenneman, D.E.; Fridkin, M.; Gozes, I. VIP and peptides related to activity-dependent neurotrophic factor protect PC12 cells against oxidative stress. J. Mol. Neurosci. MN 2000, 15, 137–145. [Google Scholar] [CrossRef]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.C.; Hashimoto, H.; Galas, L.; et al. Pituitary Adenylate Cyclase-Activating Polypeptide and Its Receptors: 20 Years after the Discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- Arimura, A.; Somogyvári-Vigh, A.; Miyata, A.; Mizuno, K.; Coy, D.H.; Kitada, C. Tissue Distribution of PACAP as Determined by RIA: Highly Abundant in the Rat Brain and Testes. Endocrinology 1991, 129, 2787–2789. [Google Scholar] [CrossRef]
- Shioda, S.; Shuto, Y.; Somogyvári-Vigh, A.; Legradi, G.; Onda, H.; Coy, D.H.; Nakajo, S.; Arimura, A. Localization and gene expression of the receptor for pituitary adenylate cyclase-activating polypeptide in the rat brain. Neurosci. Res. 1997, 28, 345–354. [Google Scholar] [CrossRef]
- Miyata, A.; Arimura, A.; Dahl, R.R.; Minamino, N.; Uehara, A.; Jiang, L.; Culler, M.D.; Coy, D.H. Isolation of a novel 38 residue-hypothalamic polypeptide which stimulates adenylate cyclase in pituitary cells. Biochem. Biophys. Res. Commun. 1989, 164, 567–574. [Google Scholar] [CrossRef]
- Miyata, A.; Jiang, L.; Dahl, R.D.; Kitada, C.; Kubo, K.; Fujino, M.; Minamino, N.; Arimura, A. Isolation of a neuropeptide corresponding to the N-terminal 27 residues of the pituitary adenylate cyclase activating polypeptide with 38 residues (PACAP38). Biochem. Biophys. Res. Commun. 1990, 170, 643–648. [Google Scholar] [CrossRef]
- Telegdy, G.; Kokavszky, K. The action of pituitary adenylate cyclase activating polypeptide (PACAP) on passive avoidance learning. The role of transmitters. Brain Res. 2000, 874, 194–199. [Google Scholar] [CrossRef]
- Cagampang, F.R.; Piggins, H.D.; Sheward, W.J.; Harmar, A.J.; Coen, C.W. Circadian changes in PACAP type 1 (PAC1) receptor mRNA in the rat suprachiasmatic and supraoptic nuclei. Brain Res. 1998, 813, 218–222. [Google Scholar] [CrossRef]
- Ohtaki, H.; Satoh, A.; Nakamachi, T.; Yofu, S.; Dohi, K.; Mori, H.; Ohara, K.; Miyamoto, K.; Hashimoto, H.; Shintani, N.; et al. Regulation of oxidative stress by pituitary adenylate cyclase-activating polypeptide (PACAP) mediated by PACAP receptor. J. Mol. Neurosci. MN 2010, 42, 397–403. [Google Scholar] [CrossRef]
- Uchida, D.; Arimura, A.; Somogyvári-Vigh, A.; Shioda, S.; Banks, W.A. Prevention of ischemia-induced death of hippocampal neurons by pituitary adenylate cyclase activating polypeptide. Brain Res. 1996, 736, 280–286. [Google Scholar] [CrossRef]
- Miyamoto, K.; Tsumuraya, T.; Ohtaki, H.; Dohi, K.; Satoh, K.; Xu, Z.; Tanaka, S.; Murai, N.; Watanabe, J.; Sugiyama, K.; et al. PACAP38 Suppresses Cortical Damage in Mice with Traumatic Brain Injury by Enhancing Antioxidant Activity: MN. J. Mol. Neurosci. 2014, 54, 370–379. [Google Scholar] [CrossRef]
- Whalen, E.J.; Johnson, A.K.; Lewis, S.J. Hemodynamic actions of systemically injected pituitary adenylate cyclase activating polypeptide-27 in the rat. Eur. J. Pharmacol. 1999, 365, 205–215. [Google Scholar] [CrossRef]
- Bruch, L.; Bychkov, R.; Kästner, A.; Bülow, T.; Ried, C.; Gollasch, M.; Baumann, G.; Luft, F.C.; Haller, H. Pituitary adenylate-cyclase-activating peptides relax human coronary arteries by activating K(ATP) and K(Ca) channels in smooth muscle cells. J. Vasc. Res. 1997, 34, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Filipsson, K.; Pacini, G.; Scheurink, A.J.; Ahrén, B. PACAP stimulates insulin secretion but inhibits insulin sensitivity in mice. Am. J. Physiol. 1998, 274, E834–E842. [Google Scholar] [CrossRef]
- Arimura, A. Perspectives on pituitary adenylate cyclase activating polypeptide (PACAP) in the neuroendocrine, endocrine, and nervous systems. Jpn. J. Physiol. 1998, 48, 301–331. [Google Scholar] [CrossRef]
- Rawlings, S.R.; Hezareh, M. Pituitary adenylate cyclase-activating polypeptide (PACAP) and PACAP/vasoactive intestinal polypeptide receptors: Actions on the anterior pituitary gland. Endocr. Rev. 1996, 17, 4–29. [Google Scholar] [CrossRef][Green Version]
- Patton, A.P.; Edwards, M.D.; Smyllie, N.J.; Hamnett, R.; Chesham, J.E.; Brancaccio, M.; Maywood, E.S.; Hastings, M.H. The VIP-VPAC2 neuropeptidergic axis is a cellular pacemaking hub of the suprachiasmatic nucleus circadian circuit. Nat. Commun. 2020, 11, 3394. [Google Scholar] [CrossRef]
- Shen, S.; Spratt, C.; Sheward, W.J.; Kallo, I.; West, K.; Morrison, C.F.; Coen, C.W.; Marston, H.M.; Harmar, A.J. Overexpression of the human VPAC2 receptor in the suprachiasmatic nucleus alters the circadian phenotype of mice. Proc. Natl. Acad. Sci. USA 2000, 97, 11575–11580. [Google Scholar] [CrossRef]
- Harmar, A.J.; Marston, H.M.; Shen, S.; Spratt, C.; West, K.M.; Sheward, W.J.; Morrison, C.F.; Dorin, J.R.; Piggins, H.D.; Reubi, J.-C.; et al. The VPAC2 Receptor Is Essential for Circadian Function in the Mouse Suprachiasmatic Nuclei. Cell 2002, 109, 497–508. [Google Scholar] [CrossRef]
- Abad, C.; Jayaram, B.; Becquet, L.; Wang, Y.; O’Dorisio, M.S.; Waschek, J.A.; Tan, Y.V. VPAC1 receptor (Vipr1)-deficient mice exhibit ameliorated experimental autoimmune encephalomyelitis, with specific deficits in the effector stage. J. Neuroinflammation 2016, 13, 169. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.; Ganea, D. Vasoactive intestinal peptide: A neuropeptide with pleiotropic immune functions. Amino Acids 2013, 45, 25–39. [Google Scholar] [CrossRef]
- Tsutsumi, M.; Claus, T.H.; Liang, Y.; Li, Y.; Yang, L.; Zhu, J.; Dela Cruz, F.; Peng, X.; Chen, H.; Yung, S.L.; et al. A potent and highly selective VPAC2 agonist enhances glucose-induced insulin release and glucose disposal: A potential therapy for type 2 diabetes. Diabetes 2002, 51, 1453–1460. [Google Scholar] [CrossRef]
- Hirabayashi, T.; Nakamachi, T.; Shioda, S. Discovery of PACAP and its receptors in the brain. J. Headache Pain 2018, 19, 28. [Google Scholar] [CrossRef]
- Usdin, T.B.; Bonner, T.I.; Mezey, E. Two receptors for vasoactive intestinal polypeptide with similar specificity and complementary distributions. Endocrinology 1994, 135, 2662–2680. [Google Scholar] [CrossRef]
- Ishihara, T.; Shigemoto, R.; Mori, K.; Takahashi, K.; Nagata, S. Functional expression and tissue distribution of a novel receptor for vasoactive intestinal polypeptide. Neuron 1992, 8, 811–819. [Google Scholar] [CrossRef]
- Stangerup, I.; Hannibal, J. Localization of Vasoactive Intestinal Polypeptide Receptor 1 (VPAC1) in Hypothalamic Neuroendocrine Oxytocin Neurons; A Potential Role in Circadian Prolactin Secretion. Front. Neuroanat. 2020, 14, 579466. [Google Scholar] [CrossRef]
- Nunan, R.; Sivasathiaseelan, H.; Khan, D.; Zaben, M.; Gray, W. Microglial VPAC1R mediates a novel mechanism of neuroimmune-modulation of hippocampal precursor cells via IL-4 release. Glia 2014, 62, 1313–1327. [Google Scholar] [CrossRef]
- Reubi, J.C. In vitro evaluation of VIP/PACAP receptors in healthy and diseased human tissues. Clinical implications. Ann. N. Y. Acad. Sci. 2000, 921, 1–25. [Google Scholar] [CrossRef]
- Yokota, C.; Kawai, K.; Ohashi, S.; Watanabe, Y.; Yamashita, K. PACAP stimulates glucose output from the perfused rat liver. Peptides 1995, 16, 55–60. [Google Scholar] [CrossRef]
- Sekiguchi, Y.; Kasai, K.; Hasegawa, K.; Suzuki, Y.; Shimoda, S.-I. Glycogenolytic activity of Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) in vivo and in vitro. Life Sci. 1994, 55, 1219–1228. [Google Scholar] [CrossRef]
- Ichikawa, S.; Sreedharan, S.P.; Owen, R.L.; Goetzl, E.J. Immunochemical localization of type I VIP receptor and NK-1-type substance P receptor in rat lung. Am. J. Physiol. 1995, 268, L584–L588. [Google Scholar] [CrossRef]
- Saga, T.; Said, S.I. Vasoactive intestinal peptide relaxes isolated strips of human bronchus, pulmonary artery, and lung parenchyma. Trans. Assoc. Am. Physicians 1984, 97, 304–310. [Google Scholar]
- Burian, B.; Storka, A.; Marzluf, B.A.; Yen, Y.-C.; Lambers, C.; Robibaro, B.; Vonbank, K.; Mosgoeller, W.; Petkov, V. Vasoactive intestinal peptide (VIP) receptor expression in monocyte-derived macrophages from COPD patients. Peptides 2010, 31, 603–608. [Google Scholar] [CrossRef]
- Beaubien, B.B.; Tippins, J.R.; Morris, H.R. Platelet-activating factor stimulation of peptidoleukotriene release: Inhibition by vasoactive polypeptide. Biochem. Biophys. Res. Commun. 1984, 125, 105–108. [Google Scholar] [CrossRef]
- Coles, S.J.; Said, S.I.; Reid, L.M. Inhibition by vasoactive intestinal peptide of glycoconjugate and lysozyme secretion by human airways in vitro. Am. Rev. Respir. Dis. 1981, 124, 531–536. [Google Scholar]
- Miotto, D.; Boschetto, P.; Bononi, I.; Zeni, E.; Cavallesco, G.; Fabbri, L.M.; Mapp, C.E. Vasoactive intestinal peptide receptors in the airways of smokers with chronic bronchitis. Eur. Respir. J. 2004, 24, 958–963. [Google Scholar] [CrossRef]
- Jayawardena, D.; Guzman, G.; Gill, R.K.; Alrefai, W.A.; Onyuksel, H.; Dudeja, P.K. Expression and localization of VPAC1, the major receptor of vasoactive intestinal peptide along the length of the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G16–G25. [Google Scholar] [CrossRef]
- Fung, C.; Unterweger, P.; Parry, L.J.; Bornstein, J.C.; Foong, J.P. VPAC1 receptors regulate intestinal secretion and muscle contractility by activating cholinergic neurons in guinea pig jejunum. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G748–G758. [Google Scholar] [CrossRef]
- Moody, T.W.; Gozes, I. Vasoactive intestinal peptide receptors: A molecular target in breast and lung cancer. Curr. Pharm. Des. 2007, 13, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Zia, H.; Hida, T.; Jakowlew, S.; Birrer, M.; Gozes, Y.; Reubi, J.C.; Fridkin, M.; Gozes, I.; Moody, T.W. Breast cancer growth is inhibited by vasoactive intestinal peptide (VIP) hybrid, a synthetic VIP receptor antagonist. Cancer Res. 1996, 56, 3486–3489. [Google Scholar] [PubMed]
- Delgado, M.; Pozo, D.; Ganea, D. The significance of vasoactive intestinal peptide in immunomodulation. Pharmacol. Rev. 2004, 56, 249–290. [Google Scholar] [CrossRef]
- Delgado, M.; Martinez, C.; Johnson, M.C.; Gomariz, R.P.; Ganea, D. Differential expression of vasoactive intestinal peptide receptors 1 and 2 (VIP-R1 and VIP-R2) mRNA in murine lymphocytes. J. Neuroimmunol. 1996, 68, 27–38. [Google Scholar] [CrossRef]
- Kaltreider, H.B.; Ichikawa, S.; Byrd, P.K.; Ingram, D.A.; Kishiyama, J.L.; Sreedharan, S.P.; Warnock, M.L.; Beck, J.M.; Goetzl, E.J. Upregulation of neuropeptides and neuropeptide receptors in a murine model of immune inflammation in lung parenchyma. Am. J. Respir. Cell Mol. Biol. 1997, 16, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Lara-Marquez, M.L.; O’Dorisio, M.S.; O’Dorisio, T.M.; Shah, M.H.; Karacay, B. Selective Gene Expression and Activation-Dependent Regulation of Vasoactive Intestinal Peptide Receptor Type 1 and Type 2 in Human T Cells. J. Immunol. 2001, 166, 2522–2530. [Google Scholar] [CrossRef]
- Sheward, W.J.; Lutz, E.M.; Harmar, A.J. The distribution of vasoactive intestinal peptide2 receptor messenger RNA in the rat brain and pituitary gland as assessed by in situ hybridization. Neuroscience 1995, 67, 409–418. [Google Scholar] [CrossRef]
- Lee, S.H.; Cox, C.L. Excitatory actions of vasoactive intestinal peptide on mouse thalamocortical neurons are mediated by VPAC2 receptors. J. Neurophysiol. 2006, 96, 858–871. [Google Scholar] [CrossRef]
- Lutz, E.M.; Sheward, W.J.; West, K.M.; Morrow, J.A.; Fink, G.; Harmar, A.J. The VIP2 receptor: Molecular characterisation of a cDNA encoding a novel receptor for vasoactive intestinal peptide. FEBS Lett. 1993, 334, 3–8. [Google Scholar] [CrossRef]
- Ago, Y.; Hayata, A.; Hashimoto, H. Pathophysiological implication of the VPAC2 receptor in psychiatric disorders. Nihon Yakurigaku Zasshi. Folia Pharmacol. Jpn. 2018, 151, 249–253. [Google Scholar] [CrossRef][Green Version]
- Ago, Y.; Asano, S.; Hashimoto, H.; Waschek, J.A. Probing the VIPR2 Microduplication Linkage to Schizophrenia in Animal and Cellular Models. Front. Neurosci. 2021, 15, 717490. [Google Scholar] [CrossRef] [PubMed]
- Vacic, V.; McCarthy, S.; Malhotra, D.; Murray, F.; Chou, H.H.; Peoples, A.; Makarov, V.; Yoon, S.; Bhandari, A.; Corominas, R.; et al. Duplications of the neuropeptide receptor gene VIPR2 confer significant risk for schizophrenia. Nature 2011, 471, 499–503. [Google Scholar] [CrossRef]
- Rangon, C.M.; Goursaud, S.; Medja, F.; Lelièvre, V.; Mounien, L.; Husson, I.; Brabet, P.; Jégou, S.; Janet, T.; Gressens, P. VPAC2 receptors mediate vasoactive intestinal peptide-induced neuroprotection against neonatal excitotoxic brain lesions in mice. J. Pharmacol. Exp. Ther. 2005, 314, 745–752. [Google Scholar] [CrossRef]
- Ago, Y.; Hayata-Takano, A.; Kawanai, T.; Yamauchi, R.; Takeuchi, S.; Cushman, J.D.; Rajbhandari, A.K.; Fanselow, M.S.; Hashimoto, H.; Waschek, J.A. Impaired extinction of cued fear memory and abnormal dendritic morphology in the prelimbic and infralimbic cortices in VPAC2 receptor (VIPR2)-deficient mice. Neurobiol. Learn. Mem. 2017, 145, 222–231. [Google Scholar] [CrossRef]
- Harmar, A.J.; Sheward, W.J.; Morrison, C.F.; Waser, B.; Gugger, M.; Reubi, J.C. Distribution of the VPAC2 Receptor in Peripheral Tissues of the Mouse. Endocrinology 2004, 145, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Fizanne, L.; Sigaudo-Roussel, D.; Saumet, J.L.; Fromy, B. Evidence for the involvement of VPAC1 and VPAC2 receptors in pressure-induced vasodilatation in rodents. J. Physiol. 2004, 554, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Ottesen, B.; Wagner, G.; Virag, R.; Fahrenkrug, J. Penile erection: Possible role for vasoactive intestinal polypeptide as a neurotransmitter. Br. Med. J. (Clin. Res. Ed.) 1984, 288, 9–11. [Google Scholar] [CrossRef]
- Inagaki, N.; Yoshida, H.; Mizuta, M.; Mizuno, N.; Fujii, Y.; Gonoi, T.; Miyazaki, J.; Seino, S. Cloning and functional characterization of a third pituitary adenylate cyclase-activating polypeptide receptor subtype expressed in insulin-secreting cells. Proc. Natl. Acad. Sci. USA 1994, 91, 2679–2683. [Google Scholar] [CrossRef]
- Groneberg, D.A.; Hartmann, P.; Dinh, Q.T.; Fischer, A. Expression and Distribution of Vasoactive Intestinal Polypeptide Receptor VPAC2 mRNA in Human Airways. Lab. Investig. 2001, 81, 749–755. [Google Scholar] [CrossRef]
- Yin, J.; Wang, L.; Yin, N.; Tabuchi, A.; Kuppe, H.; Wolff, G.; Kuebler, W.M. Vasodilatory Effect of the Stable Vasoactive Intestinal Peptide Analog RO 25-1553 in Murine and Rat Lungs. PLoS ONE 2013, 8, e75861. [Google Scholar] [CrossRef]
- St Hilaire, R.C.; Murthy, S.N.; Kadowitz, P.J.; Jeter, J.R., Jr. Role of VPAC1 and VPAC2 in VIP mediated inhibition of rat pulmonary artery and aortic smooth muscle cell proliferation. Peptides 2010, 31, 1517–1522. [Google Scholar] [CrossRef] [PubMed]
- Goetzl, E.J.; Voice, J.K.; Shen, S.; Dorsam, G.; Kong, Y.; West, K.M.; Morrison, C.F.; Harmar, A.J. Enhanced delayed-type hypersensitivity and diminished immediate-type hypersensitivity in mice lacking the inducible VPAC2 receptor for vasoactive intestinal peptide. Proc. Natl. Acad. Sci. USA 2001, 98, 13854–13859. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, H.; Nogi, H.; Mori, K.; Ohishi, H.; Shigemoto, R.; Yamamoto, K.; Matsuda, T.; Mizuno, N.; Nagata, S.; Baba, A. Distribution of the mRNA for a pituitary adenylate cyclase-activating polypeptide receptor in the rat brain: An in situ hybridization study. J. Comp. Neurol. 1996, 371, 567–577. [Google Scholar] [CrossRef]
- Shioda, S.; Nakai, Y.; Nakajo, S.; Nakaya, K.; Arimura, A. Pituitary Adenylate Cyclase-Activating Polypeptide and Its Type I Receptors in the Rat Hypothalamus: Neuroendocrine Interactionsa. Ann. N. Y. Acad. Sci. 1996, 805, 670–676. [Google Scholar] [CrossRef]
- Missig, G.; Mei, L.; Vizzard, M.A.; Braas, K.M.; Waschek, J.A.; Ressler, K.J.; Hammack, S.E.; May, V. Parabrachial Pituitary Adenylate Cyclase-Activating Polypeptide Activation of Amygdala Endosomal Extracellular Signal–Regulated Kinase Signaling Regulates the Emotional Component of Pain. Biol. Psychiatry 2017, 81, 671–682. [Google Scholar] [CrossRef]
- Hashimoto, H.; Kunugi, A.; Arakawa, N.; Shintani, N.; Fujita, T.; Kasai, A.; Kawaguchi, C.; Morita, Y.; Hirose, M.; Sakai, Y.; et al. Possible involvement of a cyclic AMP-dependent mechanism in PACAP-induced proliferation and ERK activation in astrocytes. Biochem. Biophys. Res. Commun. 2003, 311, 337–343. [Google Scholar] [CrossRef]
- Vu, J.P.; Goyal, D.; Luong, L.; Oh, S.; Sandhu, R.; Norris, J.; Parsons, W.; Pisegna, J.R.; Germano, P.M. PACAP intraperitoneal treatment suppresses appetite and food intake via PAC1 receptor in mice by inhibiting ghrelin and increasing GLP-1 and leptin. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G816–G825. [Google Scholar] [CrossRef]
- Morley, J.E.; Horowitz, M.; Morley, P.M.; Flood, J.F. Pituitary adenylate cyclase activating polypeptide (PACAP) reduces food intake in mice. Peptides 1992, 13, 1133–1135. [Google Scholar] [CrossRef]
- Otto, C.; Martin, M.; Wolfer, D.P.; Lipp, H.P.; Maldonado, R.; Schütz, G. Altered emotional behavior in PACAP-type-I-receptor-deficient mice. Brain Res. Mol. Brain Res. 2001, 92, 78–84. [Google Scholar] [CrossRef]
- Roman, C.W.; Lezak, K.R.; Hartsock, M.J.; Falls, W.A.; Braas, K.M.; Howard, A.B.; Hammack, S.E.; May, V. PAC1 receptor antagonism in the bed nucleus of the stria terminalis (BNST) attenuates the endocrine and behavioral consequences of chronic stress. Psychoneuroendocrinology 2014, 47, 151–165. [Google Scholar] [CrossRef]
- Stroth, N.; Liu, Y.; Aguilera, G.; Eiden, L.E. Pituitary Adenylate Cyclase-Activating Polypeptide Controls Stimulus-Transcription Coupling in the Hypothalamic-Pituitary-Adrenal Axis to Mediate Sustained Hormone Secretion During Stress. J. Neuroendocrinol. 2011, 23, 944–955. [Google Scholar] [CrossRef] [PubMed]
- Hammack, S.E.; Roman, C.W.; Lezak, K.R.; Kocho-Shellenberg, M.; Grimmig, B.; Falls, W.A.; Braas, K.; May, V. Roles for Pituitary Adenylate Cyclase-Activating Peptide (PACAP) Expression and Signaling in the Bed Nucleus of the Stria Terminalis (BNST) in Mediating the Behavioral Consequences of Chronic Stress. J. Mol. Neurosci. 2010, 42, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Ressler, K.J.; Mercer, K.B.; Bradley, B.; Jovanovic, T.; Mahan, A.; Kerley, K.; Norrholm, S.D.; Kilaru, V.; Smith, A.K.; Myers, A.J.; et al. Post-traumatic stress disorder is associated with PACAP and the PAC1 receptor. Nature 2011, 470, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Toth, D.; Tamas, A.; Reglodi, D. The Neuroprotective and Biomarker Potential of PACAP in Human Traumatic Brain Injury. Int. J. Mol. Sci. 2020, 21, 827. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Beltrán, E.; Correnti, E.; Deen, M.; Kamm, K.; Kelderman, T.; Papetti, L.; Vigneri, S.; MaassenVanDenBrink, A.; Edvinsson, L.; On behalf of the European Headache Federation School of Advanced Studies (EHF-SAS). PACAP38 and PAC1 receptor blockade: A new target for headache? J. Headache Pain 2018, 19, 64. [Google Scholar] [CrossRef] [PubMed]
- Schaler, A.W.; Runyan, A.M.; Clelland, C.L.; Sydney, E.J.; Fowler, S.L.; Figueroa, H.Y.; Shioda, S.; Santa-Maria, I.; Duff, K.E.; Myeku, N. PAC1 receptor-mediated clearance of tau in postsynaptic compartments attenuates tau pathology in mouse brain. Sci. Transl. Med. 2021, 13, 595. [Google Scholar] [CrossRef]
- Han, P.; Caselli, R.J.; Baxter, L.; Serrano, G.; Yin, J.; Beach, T.G.; Reiman, E.M.; Shi, J. Association of pituitary adenylate cyclase-activating polypeptide with cognitive decline in mild cognitive impairment due to Alzheimer disease. JAMA Neurol. 2015, 72, 333–339. [Google Scholar] [CrossRef]
- Basille, M.; Cartier, D.; Vaudry, D.; Lihrmann, I.; Fournier, A.; Freger, P.; Gallo-Payet, N.; Vaudry, H.; Gonzalez, B. Localization and characterization of pituitary adenylate cyclase-activating polypeptide receptors in the human cerebellum during development. J. Comp. Neurol. 2006, 496, 468–478. [Google Scholar] [CrossRef]
- Basille, M.; Vaudry, D.; Coulouarn, Y.; Jegou, S.; Lihrmann, I.; Fournier, A.; Vaudry, H.; Gonzalez, B. Comparative distribution of pituitary adenylate cyclase-activating polypeptide (PACAP) binding sites and PACAP receptor mRNAs in the rat brain during development. J. Comp. Neurol. 2000, 425, 495–509. [Google Scholar] [CrossRef]
- Nicot, A.; DiCicco-Bloom, E. Regulation of Neuroblast Mitosis is Determined by PACAP Receptor Isoform Expression. Proc. Natl. Acad. Sci. USA 2001, 98, 4758–4763. [Google Scholar] [CrossRef]
- Yan, Y.; Zhou, X.; Pan, Z.; Ma, J.; Waschek, J.A.; DiCicco-Bloom, E. Pro- and anti-mitogenic actions of pituitary adenylate cyclase-activating polypeptide in developing cerebral cortex: Potential mediation by developmental switch of PAC1 receptor mRNA isoforms. J. Neurosci. 2013, 33, 3865–3878. [Google Scholar] [CrossRef] [PubMed]
- Hirose, M.; Hashimoto, H.; Iga, J.; Shintani, N.; Nakanishi, M.; Arakawa, N.; Shimada, T.; Baba, A. Inhibition of self-renewal and induction of neural differentiation by PACAP in neural progenitor cells. Ann. N. Y. Acad. Sci 2006, 1070, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Castrogiovanni, P.; Saccone, S.; Federico, C.; Reibaldi, M.; Russo, A.; Bonfiglio, V.; Avitabile, T.; Longo, A.; et al. PACAP through EGFR transactivation preserves human corneal endothelial integrity. J. Cell. Biochem. 2019, 120, 10097–10105. [Google Scholar] [CrossRef] [PubMed]
- Nakamachi, T.; Matkovits, A.; Seki, T.; Shioda, S. Distribution and protective function of pituitary adenylate cyclase-activating polypeptide in the retina. Front. Endocrinol. 2012, 3, 145. [Google Scholar] [CrossRef] [PubMed]
- Nakamachi, T.; Ohtaki, H.; Seki, T.; Yofu, S.; Kagami, N.; Hashimoto, H.; Shintani, N.; Baba, A.; Mark, L.; Lanekoff, I.; et al. PACAP suppresses dry eye signs by stimulating tear secretion. Nat. Commun. 2016, 7, 12034. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Bucolo, C.; D’Agata, V. Protective effect of PACAP-38 on retinal pigmented epithelium in an in vitro and in vivo model of diabetic retinopathy through EGFR-dependent mechanism. Peptides 2019, 119, 170108. [Google Scholar] [CrossRef]
- Xu, Z.; Ohtaki, H.; Watanabe, J.; Miyamoto, K.; Murai, N.; Sasaki, S.; Matsumoto, M.; Hashimoto, H.; Hiraizumi, Y.; Numazawa, S.; et al. Pituitary adenylate cyclase-activating polypeptide (PACAP) contributes to the proliferation of hematopoietic progenitor cells in murine bone marrow via PACAP-specific receptor. Sci. Rep. 2016, 6, 22373. [Google Scholar] [CrossRef]
- Lamouche, S.; Yamaguchi, N. Role of PAC(1) receptor in adrenal catecholamine secretion induced by PACAP and VIP in vivo. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R510–R518. [Google Scholar] [CrossRef]
- Jamen, F.; Persson, K.; Bertrand, G.; Rodriguez-Henche, N.; Puech, R.; Bockaert, J.; Ahrén, B.; Brabet, P. PAC1 receptor–deficient mice display impaired insulinotropic response to glucose and reduced glucose tolerance. J. Clin. Investig. 2000, 105, 1307–1315. [Google Scholar] [CrossRef]
- Merriam, L.A.; Barstow, K.L.; Parsons, R.L. Pituitary adenylate cyclase-activating polypeptide enhances the hyperpolarization-activated nonselective cationic conductance, Ih, in dissociated guinea pig intracardiac neurons. Regul. Pept. 2004, 123, 123–133. [Google Scholar] [CrossRef]
- Ojala, J.; Tooke, K.; Hsiang, H.; Girard, B.M.; May, V.; Vizzard, M.A. PACAP/PAC1 Expression and Function in Micturition Pathways. J. Mol. Neurosci. 2019, 68, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Akerman, S.; Goadsby, P.J. Neuronal PAC1 receptors mediate delayed activation and sensitization of trigeminocervical neurons: Relevance to migraine. Sci. Transl. Med. 2015, 7, 308ra157. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, L.; Tajti, J.; Szalárdy, L.; Vécsei, L. PACAP and its role in primary headaches. J. Headache Pain 2018, 19, 21. [Google Scholar] [CrossRef] [PubMed]
- Hensley, K.; Pretorius, J.; Chan, B.; Page, K.; Liu, H.; Choi, C.; Shi, D.; Xu, C.; Edvinsson, L.; Miller, S. PAC1 receptor mRNA and protein distribution in rat and human trigeminal and sphenopalatine ganglia, spinal trigeminal nucleus and in dura mater. Cephalalgia Int. J. Headache 2019, 39, 827–840. [Google Scholar] [CrossRef]
- Ross, R.A.; Hoeppner, S.S.; Hellberg, S.N.; O’Day, E.B.; Rosencrans, P.L.; Ressler, K.J.; May, V.; Simon, N.M. Circulating PACAP peptide and PAC1R genotype as possible transdiagnostic biomarkers for anxiety disorders in women: A preliminary study. Neuropsychopharmacology 2020, 45, 1125–1133. [Google Scholar] [CrossRef]
- Stroth, N.; Holighaus, Y.; Ait-Ali, D.; Eiden, L.E. PACAP: A master regulator of neuroendocrine stress circuits and the cellular stress response. Ann. N. Y. Acad. Sci. 2011, 1220, 49–59. [Google Scholar] [CrossRef]
- Agarwal, A.; Halvorson, L.M.; Legradi, G. Pituitary adenylate cyclase-activating polypeptide (PACAP) mimics neuroendocrine and behavioral manifestations of stress: Evidence for PKA-mediated expression of the corticotropin-releasing hormone (CRH) gene. Brain Res. Mol. Brain Res. 2005, 138, 45–57. [Google Scholar] [CrossRef]
- Norrholm, S.D.; Das, M.; Légrádi, G. Behavioral effects of local microinfusion of pituitary adenylate cyclase activating polypeptide (PACAP) into the paraventricular nucleus of the hypothalamus (PVN). Regul. Pept. 2005, 128, 33–41. [Google Scholar] [CrossRef]
- Hashimoto, H.; Shintani, N.; Tanaka, K.; Mori, W.; Hirose, M.; Matsuda, T.; Sakaue, M.; Miyazaki, J.; Niwa, H.; Tashiro, F.; et al. Altered psychomotor behaviors in mice lacking pituitary adenylate cyclase-activating polypeptide (PACAP). Proc. Natl. Acad. Sci. USA 2001, 98, 13355–13360. [Google Scholar] [CrossRef]
- Voice, J.K.; Dorsam, G.; Lee, H.; Kong, Y.; Goetzl, E.J. Allergic diathesis in transgenic mice with constitutive T cell expression of inducible vasoactive intestinal peptide receptor. FASEB J. 2001, 15, 2489–2496. [Google Scholar] [CrossRef]
- Villanueva-Romero, R.; Gutiérrez-Cañas, I.; Carrión, M.; Pérez-García, S.; Seoane, I.V.; Martínez, C.; Gomariz, R.P.; Juarranz, Y. The Anti-Inflammatory Mediator, Vasoactive Intestinal Peptide, Modulates the Differentiation and Function of Th Subsets in Rheumatoid Arthritis. J. Immunol. Res. 2018, 2018, 6043710. [Google Scholar] [CrossRef] [PubMed]
- Martínez, C.; Juarranz, Y.; Gutiérrez-Cañas, I.; Carrión, M.; Pérez-García, S.; Villanueva-Romero, R.; Castro, D.; Lamana, A.; Mellado, M.; González-Álvaro, I.; et al. A Clinical Approach for the Use of VIP Axis in Inflammatory and Autoimmune Diseases. Int. J. Mol. Sci. 2020, 21, 65. [Google Scholar] [CrossRef] [PubMed]
- Abad, C.; Martinez, C.; Juarranz, M.G.; Arranz, A.; Leceta, J.; Delgado, M.; Gomariz, R.P. Therapeutic effects of vasoactive intestinal peptide in the trinitrobenzene sulfonic acid mice model of Crohn’s disease. Gastroenterology 2003, 124, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Lodde, B.M.; Mineshiba, F.; Wang, J.; Cotrim, A.P.; Afione, S.; Tak, P.P.; Baum, B.J. Effect of human vasoactive intestinal peptide gene transfer in a murine model of Sjögren’s syndrome. Ann. Rheum. Dis. 2006, 65, 195–200. [Google Scholar] [CrossRef]
- Li, H.; Mei, Y.; Wang, Y.; Xu, L. Vasoactive intestinal polypeptide suppressed experimental autoimmune encephalomyelitis by inhibiting T helper 1 responses. J. Clin. Immunol. 2006, 26, 430–437. [Google Scholar] [CrossRef]
- Delgado, M.; Abad, C.; Martinez, C.; Leceta, J.; Gomariz, R.P. Vasoactive intestinal peptide prevents experimental arthritis by downregulating both autoimmune and inflammatory components of the disease. Nat. Med. 2001, 7, 563–568. [Google Scholar] [CrossRef]
- Suda, K.; Smith, D.M.; Ghatei, M.A.; Bloom, S.R. Investigation of the interaction of VIP binding sites with VIP and PACAP in human brain. Neurosci. Lett. 1992, 137, 19–23. [Google Scholar] [CrossRef]
- Lam, H.-C.; Takahashi, K.; Ghatei, M.A.; Kanse, S.M.; Polak, J.M.; Bloom, S.R. Binding sites of a novel neuropeptide pituitary-adenylate-cyclase-activating polypeptide in the rat brain and lung. Eur. J. Biochem. 1990, 193, 725–729. [Google Scholar] [CrossRef]
- Gottschall, P.E.; Tatsuno, I.; Miyata, A.; Arimura, A. Characterization and Distribution of Binding Sites for the Hypothalamic Peptide, Pituitary Adenylate Cyclase-Activating Polypeptide. Endocrinology 1990, 127, 272–277. [Google Scholar] [CrossRef]
- Cauvin, A.; Buscail, L.; Gourlet, P.; De Neef, P.; Gossen, D.; Arimura, A.; Miyata, A.; Coy, D.H.; Robberecht, P.; Christophe, J. The novel VIP-like hypothalamic polypeptide PACAP interacts with high affinity receptors in the human neuroblastoma cell line NB-OK. Peptides 1990, 11, 773–777. [Google Scholar] [CrossRef]
- Robberecht, P.; Waelbroeck, M.; De Neef, P.; Tastenoy, M.; Gourlet, P.; Cogniaux, J.; Christophe, J. A new type of functional VIP receptor has an affinity for helodermin in human SUP-T1 lymphoblasts. FEBS Lett. 1988, 228, 351–355. [Google Scholar] [CrossRef]
- Robberecht, P.; De Neef, P.; Gourlet, P.; Cauvin, A.; Coy, D.H.; Christophe, J. Pharmacological characterization of the novel helodermin/VIP receptor present in human SUP-T1 lymphoma cell membranes. Regul. Pept. 1989, 26, 117–126. [Google Scholar] [CrossRef]
- Hoshino, M.; Yanaihara, C.; Hong, Y.M.; Kishida, S.; Katsumaru, Y.; Vandermeers, A.; Vandermeers-Piret, M.C.; Robberecht, P.; Christophe, J.; Yanaihara, N. Primary structure of helodermin, a VIP-secretin-like peptide isolated from Gila monster venom. FEBS Lett. 1984, 178, 233–239. [Google Scholar] [CrossRef]
- Raufman, J.P.; Jensen, R.T.; Sutliff, V.E.; Pisano, J.J.; Gardner, J.D. Actions of Gila monster venom on dispersed acini from guinea pig pancreas. Am. J. Physiol.-Gastrointest. Liver Physiol. 1982, 242, G470–G474. [Google Scholar] [CrossRef]
- Robberecht, P.; Waelbroeck, M.; Dehaye, J.-P.; Winand, J.; Vandermeers, A.; Vandermeers-Piret, M.-C.; Christophe, J. Evidence that helodermin, a newly extracted peptide from Gila monster venom, is a member of the secretin/VIP/PHI family of peptides with an original pattern of biological properties. FEBS Lett. 1984, 166, 277–282. [Google Scholar] [CrossRef]
- Amiranoff, B.; Vauclin-Jacques, N.; Boige, N.; Rouyer-Fessard, C.; Laburthe, M. Interaction of Gila monster venom with VIP receptors in intestinal epithelium of human: A comparison with rat. FEBS Lett. 1983, 164, 299–302. [Google Scholar] [CrossRef]
- Itoh, N.; Obata, K.-i.; Yanaihara, N.; Okamoto, H. Human preprovasoactive intestinal polypeptide contains a novel PHI-27-like peptide, PHM-27. Nature 1983, 304, 547–549. [Google Scholar] [CrossRef]
- Tatemoto, K.; Mutt, V. Isolation and characterization of the intestinal peptide porcine PHI (PHI-27), a new member of the glucagon--secretin family. Proc. Natl. Acad. Sci. USA 1981, 78, 6603–6607. [Google Scholar] [CrossRef]
- Bodner, M.; Fridkin, M.; Gozes, I. Coding sequences for vasoactive intestinal peptide and PHM-27 peptide are located on two adjacent exons in the human genome. Proc. Natl. Acad. Sci. USA 1985, 82, 3548–3551. [Google Scholar] [CrossRef]
- Laburthe, M.; Amiranoff, B.; Boige, N.; Rouyer-Fessard, C.; Tatemoto, K.; Moroder, L. Interaction of GRF with VIP receptors and stimulation of adenylate cyclase in rat and human intestinal epithelial membranes: Comparison with PHI and secretin. FEBS Lett. 1983, 159, 89–92. [Google Scholar] [CrossRef]
- Couvineau, A.; Rouyer-Fessard, C.; Maoret, J.J.; Gaudin, P.; Nicole, P.; Laburthe, M. Vasoactive intestinal peptide (VIP)1 receptor. Three nonadjacent amino acids are responsible for species selectivity with respect to recognition of peptide histidine isoleucineamide. J. Biol. Chem. 1996, 271, 12795–12800. [Google Scholar] [CrossRef]
- Moro, O.; Lerner, E.A. Maxadilan, the vasodilator from sand flies, is a specific pituitary adenylate cyclase activating peptide type I receptor agonist. J. Biol. Chem. 1997, 272, 966–970. [Google Scholar] [CrossRef] [PubMed]
- Uchida, D.; Tatsuno, I.; Tanaka, T.; Hirai, A.; Saito, Y.; Moro, O.; Tajima, M. Maxadilan Is a Specific Agonist and Its Deleted Peptide (M65) Is a Specific Antagonist for PACAP Type 1 Receptor. Ann. N. Y. Acad. Sci. 1998, 865, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Watanabe, M.; Yada, T. Cytosolic Ca2+ responses to sub-picomolar and nanomolar PACAP in pancreatic beta-cells are mediated by VPAC2 and PAC1 receptors. Regul. Pept. 2004, 123, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.-L.; Belousoff, M.J.; Zhao, P.; Koole, C.; Fletcher, M.M.; Truong, T.T.; Julita, V.; Christopoulos, G.; Xu, H.E.; Zhang, Y.; et al. Toward a Structural Understanding of Class B GPCR Peptide Binding and Activation. Mol. Cell 2020, 77, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Shen, D.-d.; Zhou, X.E.; Bi, P.; Liu, Q.-f.; Tan, Y.-x.; Zhuang, Y.-w.; Zhang, H.-b.; Xu, P.-y.; Huang, S.-J.; et al. Cryo-EM structure of an activated VIP1 receptor-G protein complex revealed by a NanoBiT tethering strategy. Nat. Commun. 2020, 11, 4121. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Feng, W.; Zhou, Q.; Liang, A.; Li, J.; Dai, A.; Zhao, F.; Yan, J.; Chen, C.-W.; Li, H.; et al. A distinctive ligand recognition mechanism by the human vasoactive intestinal polypeptide receptor 2. Nat. Commun. 2022, 13, 2272. [Google Scholar] [CrossRef]
- Wang, J.; Song, X.; Zhang, D.; Chen, X.; Li, X.; Sun, Y.; Li, C.; Song, Y.; Ding, Y.; Ren, R.; et al. Cryo-EM structures of PAC1 receptor reveal ligand binding mechanism. Cell Res. 2020, 30, 436–445. [Google Scholar] [CrossRef]
- Kobayashi, K.; Shihoya, W.; Nishizawa, T.; Kadji, F.M.N.; Aoki, J.; Inoue, A.; Nureki, O. Cryo-EM structure of the human PAC1 receptor coupled to an engineered heterotrimeric G protein. Nat. Struct. Mol. Biol. 2020, 27, 274–280. [Google Scholar] [CrossRef]
- Tams, J.W.; Jørgensen, R.M.; Holm, A.; Fahrenkrug, J. Creation of a Selective Antagonist and Agonist of the Rat VPAC1 Receptor Using a Combinatorial Approach with Vasoactive Intestinal Peptide 6–23 as Template. Mol. Pharmacol. 2000, 58, 1035–1041. [Google Scholar] [CrossRef]
- Gourlet, P.; Vandermeers-Piret, M.C.; Rathé, J.; De Neef, P.; Cnudde, J.; Robberecht, P.; Waelbroeck, M. Vasoactive intestinal peptide modification at position 22 allows discrimination between receptor subtypes. Eur. J. Pharmacol. 1998, 348, 95–99. [Google Scholar] [CrossRef]
- Nicole, P.; Lins, L.; Rouyer-Fessard, C.; Drouot, C.; Fulcrand, P.; Thomas, A.; Couvineau, A.; Martinez, J.; Brasseur, R.; Laburthe, M. Identification of key residues for interaction of vasoactive intestinal peptide with human VPAC1 and VPAC2 receptors and development of a highly selective VPAC1 receptor agonist. Alanine scanning and molecular modeling of the peptide. J. Biol. Chem. 2000, 275, 24003–24012. [Google Scholar] [CrossRef] [PubMed]
- Van Rampelbergh, J.; Juarranz, M.G.; Perret, J.; Bondue, A.; Solano, R.M.; Delporte, C.; De Neef, P.; Robberecht, P.; Waelbroeck, M. Characterization of a novel VPAC(1) selective agonist and identification of the receptor domains implicated in the carboxyl-terminal peptide recognition. Br. J. Pharmacol. 2000, 130, 819–826. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gourlet, P.; Vandermeers, A.; Vertongen, P.; Rathe, J.; De Neef, P.; Cnudde, J.; Waelbroeck, M.; Robberecht, P. Development of high affinity selective VIP1 receptor agonists. Peptides 1997, 18, 1539–1545. [Google Scholar] [CrossRef]
- Gourlet, P.; de Neef, P.; Cnudde, J.; Waelbroeck, M.; Robberecht, P. In Vitro Properties of a High Affinity Selective Antagonist of the VIP1 Receptor. Peptides 1997, 18, 1555–1560. [Google Scholar] [CrossRef]
- Xia, M.; Sreedharan, S.P.; Bolin, D.R.; Gaufo, G.O.; Goetzl, E.J. Novel cyclic peptide agonist of high potency and selectivity for the type II vasoactive intestinal peptide receptor. J. Pharmacol. Exp. Ther. 1997, 281, 629–633. [Google Scholar]
- Moreno, D.; Gourlet, P.; De Neef, P.; Cnudde, J.; Waelbroeck, M.; Robberecht, P. Development of selective agonists and antagonists for the human vasoactive intestinal polypeptide VPAC2 receptor. Peptides 2000, 21, 1543–1549. [Google Scholar] [CrossRef]
- Tatsuno, I.; Uchida, D.; Tanaka, T.; Saeki, N.; Hirai, A.; Saito, Y.; Moro, O.; Tajima, M. Maxadilan specifically interacts with PAC1 receptor, which is a dominant form of PACAP/VIP family receptors in cultured rat cortical neurons. Brain Res. 2001, 889, 138–148. [Google Scholar] [CrossRef]
- Gourlet, P.; Vandermeers, A.; Vandermeers-Piret, M.-C.; Rathé, J.; De Neef, P.; Robberecht, P. C-Terminally shortened pituitary adenylate cyclase-activating peptides (PACAP) discriminate PACAP I, PACAP II-VIP1 and PACAP II-VIP2 recombinant receptors. Regul. Pept. 1996, 62, 125–130. [Google Scholar] [CrossRef]
- Gourlet, P.; Vandermeers, A.; Vandermeers-Piret, M.-C.; Rathé, J.; De Neef, P.; Robberecht, P. Fragments of pituitary adenylate cyclase activating polypeptide discriminate between type I and II recombinant receptors. Eur. J. Pharmacol. 1995, 287, 7–11. [Google Scholar] [CrossRef]
- Robberecht, P.; Gourlet, P.; De Neef, P.; Woussen-Colle, M.-C.; Vandermeers-Piret, M.-C.; Vandermeers, A.; Christophe, J. Structural requirements for the occupancy of pituitary adenylate-cyclase-activating-peptide (PACAP) receptors and adenylate cyclase activation in human neuroblastoma NB-OK-1 cell membranes. Eur. J. Biochem. 1992, 207, 239–246. [Google Scholar] [CrossRef]
- Turner, J.T.; Jones, S.B.; Bylund, D.B. A fragment of vasoactive intestinal peptide, VIP(10-28), is an antagonist of VIP in the colon carcinoma cell line, HT29. Peptides 1986, 7, 849–854. [Google Scholar] [CrossRef]
- Göke, R.; Fehmann, H.C.; Linn, T.; Schmidt, H.; Krause, M.; Eng, J.; Göke, B. Exendin-4 is a high potency agonist and truncated exendin-(9-39)-amide an antagonist at the glucagon-like peptide 1-(7-36)-amide receptor of insulin-secreting beta-cells. J. Biol. Chem. 1993, 268, 19650–19655. [Google Scholar] [CrossRef]
- Pozvek, G.; Hilton, J.M.; Quiza, M.; Houssami, S.; Sexton, P.M. Structure/function relationships of calcitonin analogues as agonists, antagonists, or inverse agonists in a constitutively activated receptor cell system. Mol. Pharmacol. 1997, 51, 658–665. [Google Scholar] [CrossRef]
- Carter, P.H.; Jüppner, H.; Gardella, T.J. Studies of the N-Terminal Region of a Parathyroid Hormone-Related Peptide(1–36) Analog: Receptor Subtype-Selective Agonists, Antagonists, and Photochemical Cross-Linking Agents1. Endocrinology 1999, 140, 4972–4981. [Google Scholar] [CrossRef] [PubMed]
- Ohta, N.; Mochizuki, T.; Hoshino, M.; Jun, L.; Kobayashi, H.; Yanaihara, N. Adrenocorticotropic hormone-releasing activity of urotensin I and its fragments in vitro. J. Pept. Res. Off. J. Am. Pept. Soc. 1997, 50, 178–183. [Google Scholar] [CrossRef]
- Rivier, J.; Rivier, C.; Vale, W. Synthetic competitive antagonists of corticotropin-releasing factor: Effect on ACTH secretion in the rat. Science 1984, 224, 889–891. [Google Scholar] [CrossRef]
- Yang, B.; Gelfanov, V.M.; Perez-Tilve, D.; DuBois, B.; Rohlfs, R.; Levy, J.; Douros, J.D.; Finan, B.; Mayer, J.P.; DiMarchi, R.D. Optimization of Truncated Glucagon Peptides to Achieve Selective, High Potency, Full Antagonists. J. Med. Chem. 2021, 64, 4697–4708. [Google Scholar] [CrossRef]
- Dong, M.; Harikumar, K.G.; Raval, S.R.; Milburn, J.E.; Clark, C.; Alcala-Torano, R.; Mobarec, J.C.; Reynolds, C.A.; Ghirlanda, G.; Christopoulos, A.; et al. Rational development of a high-affinity secretin receptor antagonist. Biochem. Pharm. 2020, 177, 113929. [Google Scholar] [CrossRef]
- O’Donnell, M.; Garippa, R.J.; Rinaldi, N.; Selig, W.M.; Simko, B.; Renzetti, L.; Tannu, S.A.; Wasserman, M.A.; Welton, A.; Bolin, D.R. Ro 25-1553: A novel, long-acting vasoactive intestinal peptide agonist. Part I: In vitro and in vivo bronchodilator studies. J. Pharmacol. Exp. Ther. 1994, 270, 1282–1288. [Google Scholar]
- Yung, S.L.; Dela Cruz, F.; Hamren, S.; Zhu, J.; Tsutsumi, M.; Bloom, J.W.; Caudle, M.; Roczniak, S.; Todd, T.; Lemoine, L.; et al. Generation of highly selective VPAC2 receptor agonists by high throughput mutagenesis of vasoactive intestinal peptide and pituitary adenylate cyclase-activating peptide. J. Biol. Chem. 2003, 278, 10273–10281. [Google Scholar] [CrossRef] [PubMed]
- Qiao, A.; Han, S.; Li, X.; Li, Z.; Zhao, P.; Dai, A.; Chang, R.; Tai, L.; Tan, Q.; Chu, X.; et al. Structural basis of Gs and Gi recognition by the human glucagon receptor. Science 2020, 367, 1346–1352. [Google Scholar] [CrossRef] [PubMed]
- Mayo, K.E.; Miller, L.J.; Bataille, D.; Dalle, S.; Göke, B.; Thorens, B.; Drucker, D.J. International Union of Pharmacology. XXXV. The Glucagon Receptor Family. Pharmacol. Rev. 2003, 55, 167. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.H.; Gromada, J.; Bouchelouche, P.; Whitmore, T.; Jelinek, L.; Kindsvogel, W.; Nishimura, E. Glucagon-mediated Ca2+ signaling in BHK cells expressing cloned human glucagon receptors. Am. J. Physiol. 1998, 274, C1552–C1562. [Google Scholar] [CrossRef]
- Bringhurst, F.R.; Juppner, H.; Guo, J.; Urena, P.; Potts, J.T., Jr.; Kronenberg, H.M.; Abou-Samra, A.B.; Segre, G.V. Cloned, stably expressed parathyroid hormone (PTH)/PTH-related peptide receptors activate multiple messenger signals and biological responses in LLC-PK1 kidney cells. Endocrinology 1993, 132, 2090–2098. [Google Scholar] [CrossRef]
- Wootten, D.; Christopoulos, A.; Marti-Solano, M.; Babu, M.M.; Sexton, P.M. Mechanisms of signalling and biased agonism in G protein-coupled receptors. Nat. Reviews. Mol. Cell Biol. 2018, 19, 638–653. [Google Scholar] [CrossRef]
- Yuliantie, E.; van der Velden, W.J.C.; Labroska, V.; Dai, A.; Zhao, F.; Darbalaei, S.; Deganutti, G.; Xu, T.; Zhou, Q.; Yang, D.; et al. Insights into agonist-elicited activation of the human glucose-dependent insulinotropic polypeptide receptor. Biochem. Pharmacol. 2021, 192, 114715. [Google Scholar] [CrossRef]
- Christopoulos, A.; Christopoulos, G.; Morfis, M.; Udawela, M.; Laburthe, M.; Couvineau, A.; Kuwasako, K.; Tilakaratne, N.; Sexton, P.M. Novel Receptor Partners and Function of Receptor Activity-modifying Proteins. J. Biol. Chem. 2003, 278, 3293–3297. [Google Scholar] [CrossRef]
- Christopoulos, G.; Perry, K.J.; Morfis, M.; Tilakaratne, N.; Gao, Y.; Fraser, N.J.; Main, M.J.; Foord, S.M.; Sexton, P.M. Multiple Amylin Receptors Arise from Receptor Activity-Modifying Protein Interaction with the Calcitonin Receptor Gene Product. Mol. Pharmacol. 1999, 56, 235–242. [Google Scholar] [CrossRef]
- Hay, D.; Christopoulos, G.; Christopoulos, A.; Sexton, P. Amylin receptors: Molecular composition and pharmacology. Biochem. Soc. Trans. 2004, 32, 865–867. [Google Scholar] [CrossRef]
- McLatchie, L.M.; Fraser, N.J.; Main, M.J.; Wise, A.; Brown, J.; Thompson, N.; Solari, R.; Lee, M.G.; Foord, S.M. RAMPs regulate the transport and ligand specificity of the calcitonin-receptor-like receptor. Nature 1998, 393, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Harikumar, K.G.; Simms, J.; Christopoulos, G.; Sexton, P.M.; Miller, L.J. Molecular Basis of Association of Receptor Activity-Modifying Protein 3 with the Family B G Protein-Coupled Secretin Receptor. Biochemistry 2009, 48, 11773–11785. [Google Scholar] [CrossRef] [PubMed]
- Furness, S.G.; Wootten, D.; Christopoulos, A.; Sexton, P.M. Consequences of splice variation on Secretin family G protein-coupled receptor function. Br. J. Pharmacol. 2012, 166, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Pan, Z.; Zhang, Z.; Lin, L.; Xing, Y. The Expanding Landscape of Alternative Splicing Variation in Human Populations. Am. J. Hum. Genet. 2018, 102, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Dautzenberg, F.M.; Mevenkamp, G.; Wille, S.; Hauger, R.L. N-terminal splice variants of the type I PACAP receptor: Isolation, characterization and ligand binding/selectivity determinants. J. Neuroendocrinol. 1999, 11, 941–949. [Google Scholar] [CrossRef]
- Lutz, E.M.; Ronaldson, E.; Shaw, P.; Johnson, M.S.; Holland, P.J.; Mitchell, R. Characterization of novel splice variants of the PAC1 receptor in human neuroblastoma cells: Consequences for signaling by VIP and PACAP. Mol. Cell. Neurosci. 2006, 31, 193–209. [Google Scholar] [CrossRef]
- Journot, L.; Spengler, D.; Pantaloni, C.; Dumuis, A.; Sebben, M.; Bockaert, J. The PACAP receptor: Generation by alternative splicing of functional diversity among G protein-coupled receptors in nerve cells. Semin. Cell Biol. 1994, 5, 263–272. [Google Scholar] [CrossRef]
- Spengler, D.; Waeber, C.; Pantaloni, C.; Holsboer, F.; Bockaert, J.; Seeburgt, P.H.; Journot, L. Differential signal transduction by five splice variants of the PACAP receptor. Nature 1993, 365, 170–175. [Google Scholar] [CrossRef]
- Pantaloni, C.; Brabet, P.; Bilanges, B.; Dumuis, A.; Houssami, S.; Spengler, D.; Bockaert, J.; Journot, L. Alternative Splicing in the N-terminal Extracellular Domain of the Pituitary Adenylate Cyclase-activating Polypeptide (PACAP) Receptor Modulates Receptor Selectivity and Relative Potencies of PACAP-27 and PACAP-38 in Phospholipase C Activation. J. Biol. Chem. 1996, 271, 22146–22151. [Google Scholar] [CrossRef]
- Blechman, J.; Levkowitz, G. Alternative Splicing of the Pituitary Adenylate Cyclase-Activating Polypeptide Receptor PAC1: Mechanisms of Fine Tuning of Brain Activity. Front. Endocrinol. 2013, 4, 55. [Google Scholar] [CrossRef]
- Pisegna, J.R.; Moody, T.W.; Wank, S.A. Differential signaling and immediate-early gene activation by four splice variants of the human pituitary adenylate cyclase-activating polypeptide receptor (hPACAP-R). Ann. N. Y. Acad. Sci. 1996, 805, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Tasma, Z.; Siow, A.; Harris, P.W.R.; Brimble, M.A.; Hay, D.L.; Walker, C.S. Characterisation of agonist signalling profiles and agonist-dependent antagonism at PACAP-responsive receptors: Implications for drug discovery. Br. J. Pharmacol. 2022, 179, 435–453. [Google Scholar] [CrossRef]
- Ushiyama, M.; Ikeda, R.; Yoshida, M.; Mori, K.; Kangawa, K.; Sugawara, H.; Inoue, K.; Yamada, K.; Miyata, A. Alternative Splicing of the Pituitary Adenylate Cyclase-activating Polypetide (PACAP) Receptor Contributes to Function of PACAP-27. J. Mol. Neurosci. 2010, 42, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, T.; Grimaldi, M.; Eiden, L.E. The hop cassette of the PAC1 receptor confers coupling to Ca2+ elevation required for pituitary adenylate cyclase-activating polypeptide-evoked neurosecretion. J. Biol. Chem. 2007, 282, 8079–8091. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Remington, J.M.; Liao, C.; Parsons, R.L.; Schneebeli, S.; Braas, K.M.; May, V.; Brewer, M. GPCR Intracellular Loop Regulation of Beta-Arrestin-Mediated Endosomal Signaling Dynamics. J. Mol. Neurosci. 2022, 72, 1358–1373. [Google Scholar] [CrossRef]
- Pisegna, J.R.; Wank, S.A. Cloning and characterization of the signal transduction of four splice variants of the human pituitary adenylate cyclase activating polypeptide receptor. Evidence for dual coupling to adenylate cyclase and phospholipase C. J. Biol. Chem. 1996, 271, 17267–17274. [Google Scholar] [CrossRef] [PubMed]
- Shneider, Y.; Shtrauss, Y.; Yadid, G.; Pinhasov, A. Differential expression of PACAP receptors in postnatal rat brain. Neuropeptides 2010, 44, 509–514. [Google Scholar] [CrossRef]
- Lakk, M.; Szabó, B.; Völgyi, B.; Gábriel, R.; Dénes, V. Development-Related Splicing Regulates Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) Receptors in the Retina. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7825–7832. [Google Scholar] [CrossRef]
- Amir-Zilberstein, L.; Blechman, J.; Sztainberg, Y.; Norton, W.H.; Reuveny, A.; Borodovsky, N.; Tahor, M.; Bonkowsky, J.L.; Bally-Cuif, L.; Chen, A.; et al. Homeodomain protein otp and activity-dependent splicing modulate neuronal adaptation to stress. Neuron 2012, 73, 279–291. [Google Scholar] [CrossRef]
- Biran, J.; Gliksberg, M.; Shirat, I.; Swaminathan, A.; Levitas-Djerbi, T.; Appelbaum, L.; Levkowitz, G. Splice-specific deficiency of the PTSD-associated gene PAC1 leads to a paradoxical age-dependent stress behavior. Sci. Rep. 2020, 10, 9559. [Google Scholar] [CrossRef]
- Scaldaferri, M.L.; Modesti, A.; Palumbo, C.; Ulisse, S.; Fabbri, A.; Piccione, E.; Frajese, G.; Moretti, C. Pituitary adenylate cyclase-activating polypeptide (PACAP) and PACAP-receptor type 1 expression in rat and human placenta. Endocrinol. 2000, 141, 1158–1167. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sreedharan, S.P.; Patel, D.R.; Xia, M.H.; Ichikawa, S.; Goetzl, E.J. Human Vasoactive Intestinal Peptide1 Receptors Expressed by Stable Transfectants Couple to 2 Distinct Signaling Pathways. Biochem. Biophys. Res. Commun. 1994, 203, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Pugh, P.C.; Margiotta, J.F. PACAP support of neuronal survival requires MAPK- and activity-generated signals. Mol. Cell. Neurosci. 2006, 31, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.A.; Vaudry, D.; Basille, M.; Castel, H.; Fournier, A.; Vaudry, H.; Gonzalez, B.J. PACAP inhibits delayed rectifier potassium current via a cAMP/PKA transduction pathway: Evidence for the involvement of IK in the anti-apoptotic action of PACAP. Eur. J. Neurosci. 2004, 19, 1446–1458. [Google Scholar] [CrossRef]
- Falluel-Morel, A.; Vaudry, D.; Aubert, N.; Galas, L.; Benard, M.; Basille, M.; Fontaine, M.; Fournier, A.; Vaudry, H.; Gonzalez, B.J. PACAP and ceramides exert opposite effects on migration, neurite outgrowth, and cytoskeleton remodeling. Ann. N. Y. Acad. Sci. 2006, 1070, 265–270. [Google Scholar] [CrossRef]
- Yaka, R.; He, D.-Y.; Phamluong, K.; Ron, D. Pituitary Adenylate Cyclase-activating Polypeptide (PACAP(1–38)) Enhances N-Methyl-d-aspartate Receptor Function and Brain-derived Neurotrophic Factor Expression via RACK1. J. Biol. Chem. 2003, 278, 9630–9638. [Google Scholar] [CrossRef]
- Fila, T.; Trazzi, S.; Crochemore, C.; Bartesaghi, R.; Ciani, E. Lot1 is a key element of the pituitary adenylate cyclase-activating polypeptide (PACAP)/cyclic AMP pathway that negatively regulates neuronal precursor proliferation. J. Biol. Chem 2009, 284, 15325–15338. [Google Scholar] [CrossRef]
- Ster, J.; de Bock, F.; Guérineau, N.C.; Janossy, A.; Barrère-Lemaire, S.; Bos, J.L.; Bockaert, J.; Fagni, L. Exchange Protein Activated by cAMP (Epac) Mediates cAMP Activation of p38 MAPK and Modulation of Calcium-Dependent Potassium Channels in Cerebellar Neurons. Proc. Natl. Acad. Sci. USA 2007, 104, 2519–2524. [Google Scholar] [CrossRef]
- Shi, G.-X.; Rehmann, H.; Andres, D.A. A Novel Cyclic AMP-Dependent Epac-Rit Signaling Pathway Contributes to PACAP38-Mediated Neuronal Differentiation. Mol. Cell. Biol. 2006, 26, 9136–9147. [Google Scholar] [CrossRef]
- Emery, A.C.; Eiden, L.E. Signaling through the neuropeptide GPCR PAC1 induces neuritogenesis via a single linear cAMP- and ERK-dependent pathway using a novel cAMP sensor. FASEB J. 2012, 26, 3199–3211. [Google Scholar] [CrossRef]
- Ravni, A.; Vaudry, D.; Gerdin, M.J.; Eiden, M.V.; Falluel-Morel, A.; Gonzalez, B.J.; Vaudry, H.; Eiden, L.E. A cAMP-dependent, protein kinase A-independent signaling pathway mediating neuritogenesis through Egr1 in PC12 cells. Mol. Pharmacol. 2008, 73, 1688–1708. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.; Zak, D.E.; Sauter, T.; Schwaber, J.; Ogunnaike, B.A. Modeling the VPAC2-Activated cAMP/PKA Signaling Pathway: From Receptor to Circadian Clock Gene Induction. Biophys. J. 2006, 90, 1560–1571. [Google Scholar] [CrossRef] [PubMed]
- Cunha-Reis, D.; Ribeiro, J.A.; Sebastião, A.M. VIP enhances synaptic transmission to hippocampal CA1 pyramidal cells through activation of both VPAC1 and VPAC2 receptors. Brain Res. 2005, 1049, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Le Péchon-Vallée, C.; Magalon, K.; Rasolonjanahary, R.; Enjalbert, A.; Gérard, C. Vasoactive intestinal polypeptide and pituitary adenylate cyclase-activating polypeptides stimulate mitogen-activated protein kinase in the pituitary cell line GH4C1 by a 3’,5’-cyclic adenosine monophosphate pathway. Neuroendocrinology 2000, 72, 46–56. [Google Scholar] [CrossRef]
- Delgado, M.; Munoz-Elias, E.J.; Kan, Y.; Gozes, I.; Fridkin, M.; Brenneman, D.E.; Gomariz, R.P.; Ganea, D. Vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide inhibit tumor necrosis factor alpha transcriptional activation by regulating nuclear factor-kB and cAMP response element-binding protein/c-Jun. J. Biol. Chem. 1998, 273, 31427–31436. [Google Scholar] [CrossRef]
- El Zein, N.; Badran, B.; Sariban, E. VIP differentially activates β2 integrins, CR1, and matrix metalloproteinase-9 in human monocytes through cAMP/PKA, EPAC, and PI-3K signaling pathways via VIP receptor type 1 and FPRL1. J. Leukoc. Biol. 2008, 83, 972–981. [Google Scholar] [CrossRef]
- MacKenzie, C.J.; Lutz, E.M.; McCulloch, D.A.; Mitchell, R.; Harmar, A.J. Phospholipase C activation by VIP1 and VIP2 receptors expressed in COS 7 cells involves a pertussis toxin-sensitive mechanism. Ann. N. Y. Acad. Sci. 1996, 805, 579–584. [Google Scholar] [CrossRef]
- Holighaus, Y.; Mustafa, T.; Eiden, L.E. PAC1hop, null and hip receptors mediate differential signaling through cyclic AMP and calcium leading to splice variant-specific gene induction in neural cells. Peptides 2011, 32, 1647–1655. [Google Scholar] [CrossRef]
- Macdonald, D.S.; Weerapura, M.; Beazely, M.A.; Martin, L.; Czerwinski, W.; Roder, J.C.; Orser, B.A.; MacDonald, J.F. Modulation of NMDA receptors by pituitary adenylate cyclase activating peptide in CA1 neurons requires G alpha q, protein kinase C, and activation of Src. J. Neurosci. 2005, 25, 11374–11384. [Google Scholar] [CrossRef]
- May, V.; Buttolph, T.R.; Girard, B.M.; Clason, T.A.; Parsons, R.L. PACAP-induced ERK activation in HEK cells expressing PAC1 receptors involves both receptor internalization and PKC signaling. Am. J. Physiol. Cell Physiol. 2014, 306, C1068–C1079. [Google Scholar] [CrossRef]
- Bouschet, T.; Perez, V.; Fernandez, C.; Bockaert, J.; Eychene, A.; Journot, L. Stimulation of the ERK pathway by GTP-loaded Rap1 requires the concomitant activation of Ras, protein kinase C, and protein kinase A in neuronal cells. J. Biol. Chem. 2003, 278, 4778–4785. [Google Scholar] [CrossRef] [PubMed]
- Dejda, A.; Jozwiak-Bebenista, M.; Nowak, J.Z. PACAP, VIP, and PHI: Effects on AC-, PLC-, and PLD-driven signaling systems in the primary glial cell cultures. Ann. N. Y. Acad. Sci. 2006, 1070, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Van Rampelbergh, J.; Poloczek, P.; Françoys, I.; Christine, D.; Winand, J.; Robberecht, P.; Waelbroeck, M. The pituitary adenylate cyclase activating polypeptide (PACAP I) and VIP (PACAP II VIP1) receptors stimulate inositol phosphate synthesis in transfected CHO cells through interaction with different G proteins. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 1997, 1357, 249–255. [Google Scholar] [CrossRef][Green Version]
- Shreeve, S.M.; Sreedharan, S.P.; Hacker, M.P.; Gannon, D.E.; Morgan, M.J. VIP Activates Gs and Gi3 in Rat Alveolar Macrophages and Gs in HEK293 Cells Transfected with the Human VPAC1 Receptor. Biochem. Biophys. Res. Commun. 2000, 272, 922–928. [Google Scholar] [CrossRef]
- Murthy, K.S.; Makhlouf, G.M. Vasoactive intestinal peptide/pituitary adenylate cyclase-activating peptide-dependent activation of membrane-bound NO synthase in smooth muscle mediated by pertussis toxin-sensitive Gi1-2. J. Biol. Chem. 1994, 269, 15977–15980. [Google Scholar] [CrossRef]
- Luo, X.; Zeng, W.; Xu, X.; Popov, S.; Davignon, I.; Wilkie, T.M.; Mumby, S.M.; Muallem, S. Alternate Coupling of Receptors to Gs and Gi in Pancreatic and Submandibular Gland Cells*. J. Biol. Chem. 1999, 274, 17684–17690. [Google Scholar] [CrossRef] [PubMed]
- McCulloch, D.A.; Lutz, E.M.; Johnson, M.S.; Robertson, D.N.; MacKenzie, C.J.; Holland, P.J.; Mitchell, R. ADP-Ribosylation Factor-Dependent Phospholipase D Activation by VPAC Receptors and a PAC1 Receptor Splice Variant. Mol. Pharmacol. 2001, 59, 1523. [Google Scholar] [CrossRef]
- McCulloch, D.A.; Lutz, E.M.; Johnson, M.S.; MacKenzie, C.J.; Mitchell, R. Differential Activation of Phospholipase D by VPAC and PAC1 Receptors. Ann. N. Y. Acad. Sci. 2000, 921, 175–185. [Google Scholar] [CrossRef]
- Broca, C.; Quoyer, J.; Costes, S.; Linck, N.; Varrault, A.; Deffayet, P.M.; Bockaert, J.; Dalle, S.; Bertrand, G. b-Arrestin 1 is required for PAC1 receptor-mediated potentiation of long-lasting ERK1/2 activation by glucose in pancreatic beta-cells. J. Biol. Chem. 2009, 284, 4332–4342. [Google Scholar] [CrossRef]
- Selvy, P.E.; Lavieri, R.R.; Lindsley, C.W.; Brown, H.A. Phospholipase D: Enzymology, Functionality, and Chemical Modulation. Chem. Rev. 2011, 111, 6064–6119. [Google Scholar] [CrossRef]
- Ronaldson, E.; Robertson, D.N.; Johnson, M.S.; Holland, P.J.; Mitchell, R.; Lutz, E.M. Specific interaction between the hop1 intracellular loop 3 domain of the human PAC1 receptor and ARF. Regul. Pept. 2002, 109, 193–198. [Google Scholar] [CrossRef]
- Khoury, E.; Nikolajev, L.; Simaan, M.; Namkung, Y.; Laporte, S.A. Differential regulation of endosomal GPCR/β-arrestin complexes and trafficking by MAPK. J. Biol. Chem. 2014, 289, 23302–23317. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, J.D.; Hardwick, J.C.; Locknar, S.A.; Merriam, L.A.; Parsons, R.L. Ca2+ Influx, But Not Ca2+ Release From Internal Stores, Is Required for the PACAP-Induced Increase in Excitability in Guinea Pig Intracardiac Neurons. J. Neurophysiol. 2006, 95, 2134–2142. [Google Scholar] [CrossRef] [PubMed]
- May, V.; Parsons, R.L. G Protein-Coupled Receptor Endosomal Signaling and Regulation of Neuronal Excitability and Stress Responses: Signaling Options and Lessons From the PAC1 Receptor. J. Cell. Physiol. 2017, 232, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Clason, T.A.; Girard, B.M.; May, V.; Parsons, R.L. Activation of MEK/ERK Signaling by PACAP in Guinea Pig Cardiac Neurons. J. Mol. Neurosci. MN 2016, 59, 309–316. [Google Scholar] [CrossRef]
- Maunze, B.; Bruckner, K.W.; Desai, N.N.; Chen, C.; Chen, F.; Baker, D.; Choi, S. Pituitary adenylate cyclase-activating polypeptide receptor activation in the hypothalamus recruits unique signaling pathways involved in energy homeostasis. Am. J. Physiol.-Endocrinol. Metab. 2022, 322, E199–E210. [Google Scholar] [CrossRef]
- May, V.; Lutz, E.; MacKenzie, C.; Schutz, K.C.; Dozark, K.; Braas, K.M. Pituitary adenylate cyclase-activating polypeptide (PACAP)/PAC1HOP1 receptor activation coordinates multiple neurotrophic signaling pathways: Akt activation through phosphatidylinositol 3-kinase gamma and vesicle endocytosis for neuronal survival. J. Biol. Chem. 2010, 285, 9749–9761. [Google Scholar] [CrossRef]
- Langer, I.; Jeandriens, J.; Couvineau, A.; Sanmukh, S.; Latek, D. Signal Transduction by VIP and PACAP Receptors. Biomedicines 2022, 10, 406. [Google Scholar] [CrossRef]
- Tanaka, K.; Shibuya, I.; Harayama, N.; Nomura, M.; Kabashima, N.; Ueta, Y.; Yamashita, H. Pituitary Adenylate Cyclase-Activating Polypeptide Potentiation of Ca2+ Entry via Protein Kinase C and A Pathways in Melanotrophs of the Pituitary Pars Intermedia of Rats*. Endocrinology 1997, 138, 4086–4095. [Google Scholar] [CrossRef][Green Version]
- Chatterjee, T.K.; Sharma, R.V.; Fisher, R.A. Molecular Cloning of a Novel Variant of the Pituitary Adenylate Cyclase-activating Polypeptide (PACAP) Receptor That Stimulates Calcium Influx by Activation of L-type Calcium Channels. J. Biol. Chem. 1996, 271, 32226–32232. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Lakhman, S.S.; Singh, S. Modulation of L-type calcium channels in Drosophila via a pituitary adenylyl cyclase-activating polypeptide (PACAP)-mediated pathway. J. Biol. Chem. 2004, 279, 37291–37297. [Google Scholar] [CrossRef] [PubMed]
- Dziema, H.; Obrietan, K. PACAP Potentiates L-Type Calcium Channel Conductance in Suprachiasmatic Nucleus Neurons by Activating the MAPK Pathway. J. Neurophysiol. 2002, 88, 1374–1386. [Google Scholar] [CrossRef] [PubMed]
- Cunha-Reis, D.; Ribeiro, J.A.; de Almeida, R.F.M.; Sebastião, A.M. VPAC(1) and VPAC(2) receptor activation on GABA release from hippocampal nerve terminals involve several different signalling pathways. Br. J. Pharmacol. 2017, 174, 4725–4737. [Google Scholar] [CrossRef]
- Liu, M.; Yang, X.; Bai, T.; Liu, Z.; Liu, T.; Wang, Y.; Cui, L.; Liu, Y.; Zhang, Y. PACAP stimulates insulin secretion by PAC1 receptor and ion channels in β-cells. Cell. Signal. 2019, 61, 48–56. [Google Scholar] [CrossRef]
- Kurahashi, M.; Baker, S.A.; Kito, Y.; Bartlett, A.; Hara, M.; Takeyama, H.; Hashitani, H.; Sanders, K.M. PDGFRα+ Interstitial Cells are Effector Cells of PACAP Signaling in Mouse and Human Colon. Cell. Mol. Gastroenterol. Hepatol. 2022, 14, 357–373. [Google Scholar] [CrossRef] [PubMed]
- Delcourt, N.; Thouvenot, E.; Chanrion, B.; Galéotti, N.; Jouin, P.; Bockaert, J.; Marin, P. PACAP type I receptor transactivation is essential for IGF-1 receptor signalling and antiapoptotic activity in neurons. EMBO J. 2007, 26, 1542–1551. [Google Scholar] [CrossRef]
- Moody, T.W.; Osefo, N.; Nuche-Berenguer, B.; Ridnour, L.; Wink, D.; Jensen, R.T. Pituitary adenylate cyclase-activating polypeptide causes tyrosine phosphorylation of the epidermal growth factor receptor in lung cancer cells. J. Pharmacol. Exp. Ther. 2012, 341, 873–881. [Google Scholar] [CrossRef]
- Valdehita, A.; Bajo, A.M.; Schally, A.V.; Varga, J.L.; Carmena, M.J.; Prieto, J.C. Vasoactive intestinal peptide (VIP) induces transactivation of EGFR and HER2 in human breast cancer cells. Mol. Cell. Endocrinol. 2009, 302, 41–48. [Google Scholar] [CrossRef]
- Van Gastel, J.; Hendrickx, J.O.; Leysen, H.; Santos-Otte, P.; Luttrell, L.M.; Martin, B.; Maudsley, S. β-Arrestin Based Receptor Signaling Paradigms: Potential Therapeutic Targets for Complex Age-Related Disorders. Front. Pharmacol. 2018, 9, 1369. [Google Scholar] [CrossRef]
- Pelech, S.L.; Sanghera, J.S. MAP Kinases: Charting the Regulatory Pathways. Science 1992, 257, 1355–1356. [Google Scholar] [CrossRef]
- Gutiérrez-Cañas, I.; Juarranz, M.G.; Collado, B.; Rodríguez-Henche, N.; Chiloeches, A.; Prieto, J.C.; Carmena, M.J. Vasoactive intestinal peptide induces neuroendocrine differentiation in the LNCaP prostate cancer cell line through PKA, ERK, and PI3K. Prostate 2005, 63, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Moroo, I.; Tatsuno, I.; Uchida, D.; Tanaka, T.; Saito, J.; Saito, Y.; Hirai, A. Pituitary adenylate cyclase activating polypeptide (PACAP) stimulates mitogen-activated protein kinase (MAPK) in cultured rat astrocytes. Brain Res. 1998, 795, 191–196. [Google Scholar] [CrossRef]
- Lefkowitz Robert, J.; Shenoy Sudha, K. Transduction of Receptor Signals by ß-Arrestins. Science 2005, 308, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Frodin, M.; Sekine, N.; Roche, E.; Filloux, C.; Prentki, M.; Wollheim, C.B.; Van Obberghen, E. Glucose, other secretagogues, and nerve growth factor stimulate mitogen-activated protein kinase in the insulin-secreting beta-cell line, INS-1. J. Biol. Chem. 1995, 270, 7882–7889. [Google Scholar] [CrossRef]
- Sexton, P.M. Recent advances in our understanding of peptide hormone receptors and RAMPS. Curr. Opin. Drug Discov. Dev. 1999, 2, 440–448. [Google Scholar]
- Hay, D.L.; Pioszak, A.A. Receptor Activity-Modifying Proteins (RAMPs): New Insights and Roles. Annu. Rev Pharm. Toxicol 2016, 56, 469–487. [Google Scholar] [CrossRef]
- Poyner, D.R.; Sexton, P.M.; Marshall, I.; Smith, D.M.; Quirion, R.; Born, W.; Muff, R.; Fischer, J.A.; Foord, S.M. International Union of Pharmacology. XXXII. The mammalian calcitonin gene-related peptides, adrenomedullin, amylin, and calcitonin receptors. Pharmacol. Rev. 2002, 54, 233–246. [Google Scholar] [CrossRef]
- Wootten, D.L.; Simms, J.; Hay, D.L.; Christopoulos, A.; Sexton, P.M. Chapter 3—Receptor Activity Modifying Proteins and Their Potential as Drug Targets. In Progress in Molecular Biology and Translational Science; Lunn, C.A., Ed.; Academic Press: Cambridge, MA, USA, 2010; Volume 91, pp. 53–79. [Google Scholar]
- Lorenzen, E.; Dodig-Crnković, T.; Kotliar, I.B.; Pin, E.; Ceraudo, E.; Vaughan, R.D.; Uhlèn, M.; Huber, T.; Schwenk, J.M.; Sakmar, T.P. Multiplexed analysis of the secretin-like GPCR-RAMP interactome. Sci. Adv. 2019, 5, eaaw2778. [Google Scholar] [CrossRef]
- Wootten, D.; Lindmark, H.; Kadmiel, M.; Willcockson, H.; Caron, K.M.; Barwell, J.; Drmota, T.; Poyner, D.R. Receptor activity modifying proteins (RAMPs) interact with the VPAC2 receptor and CRF1 receptors and modulate their function. Br. J. Pharmacol. 2013, 168, 822–834. [Google Scholar] [CrossRef]
- Stachniak, T.; Krukoff, T.L. Receptor activity modifying protein 2 distribution in the rat central nervous system and regulation by changes in blood pressure. J. Neuroendocrinol. 2003, 15, 840–850. [Google Scholar] [CrossRef]
- Tilakaratne, N.; Christopoulos, G.; Zumpe, E.T.; Foord, S.M.; Sexton, P.M. Amylin receptor phenotypes derived from human calcitonin receptor/RAMP coexpression exhibit pharmacological differences dependent on receptor isoform and host cell environment. J. Pharmacol. Exp. Ther. 2000, 294, 61–72. [Google Scholar] [PubMed]
- Goodman, O.B.; Krupnick, J.G.; Santini, F.; Gurevich, V.V.; Penn, R.B.; Gagnon, A.W.; Keen, J.H.; Benovic, J.L. β-Arrestin acts as a clathrin adaptor in endocytosis of the β2-adrenergic receptor. Nature 1996, 383, 447–450. [Google Scholar] [CrossRef] [PubMed]
- Laporte, S.A.; Oakley, R.H.; Zhang, J.; Holt, J.A.; Ferguson, S.S.; Caron, M.G.; Barak, L.S. The beta2-adrenergic receptor/betaarrestin complex recruits the clathrin adaptor AP-2 during endocytosis. Proc. Natl. Acad. Sci. USA 1999, 96, 3712–3717. [Google Scholar] [CrossRef] [PubMed]
- Vilardaga, J.-P.; Jean-Alphonse, F.G.; Gardella, T.J. Endosomal generation of cAMP in GPCR signaling. Nat. Chem. Biol. 2014, 10, 700–706. [Google Scholar] [CrossRef]
- Lyu, R.-M.; Germano, P.M.; Choi, J.K.; Le, S.V.; Pisegna, J.R. Identification of an Essential Amino Acid Motif within the C Terminus of the Pituitary Adenylate Cyclase-activating Polypeptide Type I Receptor That Is Critical for Signal Transduction but Not for Receptor Internalization*. J. Biol. Chem. 2000, 275, 36134–36142. [Google Scholar] [CrossRef]
- Langlet, C.; Langer, I.; Vertongen, P.; Gaspard, N.; Vanderwinden, J.-M.; Robberecht, P. Contribution of the Carboxyl Terminus of the VPAC1 Receptor to Agonist-induced Receptor Phosphorylation, Internalization, and Recycling*. J. Biol. Chem. 2005, 280, 28034–28043. [Google Scholar] [CrossRef]
- Marie, J.C.; Rouyer-Fessard, C.; Couvineau, A.; Nicole, P.; Devaud, H.; El Benna, J.; Laburthe, M. Serine 447 in the carboxyl tail of human VPAC1 receptor is crucial for agonist-induced desensitization but not internalization of the receptor. Mol. Pharmacol. 2003, 64, 1565–1574. [Google Scholar] [CrossRef]
- Langer, I.; Langlet, C.; Robberecht, P. Effect of inactivating mutations on phosphorylation and internalization of the human VPAC2 receptor. J. Mol. Endocrinol. 2005, 34, 405–414. [Google Scholar] [CrossRef]
- Inglese, J.; Freedman, N.J.; Koch, W.J.; Lefkowitz, R.J. Structure and mechanism of the G protein-coupled receptor kinases. J. Biol. Chem. 1993, 268, 23735–23738. [Google Scholar] [CrossRef]
- Magalhaes, A.C.; Dunn, H.; Ferguson, S.S. Regulation of GPCR activity, trafficking and localization by GPCR-interacting proteins. Br. J. Pharmacol. 2012, 165, 1717–1736. [Google Scholar] [CrossRef]
- Dautzenberg, F.M.; Hauger, R.L. G-protein-coupled receptor kinase 3- and protein kinase C-mediated desensitization of the PACAP receptor type 1 in human Y-79 retinoblastoma cells. Neuropharmacology 2001, 40, 394–407. [Google Scholar] [CrossRef]
- Murthy, K.S.; Mahavadi, S.; Huang, J.; Zhou, H.; Sriwai, W. Phosphorylation of GRK2 by PKA augments GRK2-mediated phosphorylation, internalization, and desensitization of VPAC2 receptors in smooth muscle. Am. J. Physiology. Cell Physiol. 2008, 294, C477–C487. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Mahavadi, S.; Sriwai, W.; Grider, J.R.; Murthy, K.S. Cross-regulation of VPAC2 receptor desensitization by M3 receptors via PKC-mediated phosphorylation of RKIP and inhibition of GRK2. Am. J. Physiol.-Gastrointest. Liver Physiol. 2007, 292, G867–G874. [Google Scholar] [CrossRef] [PubMed]
- Shetzline, M.A.; Walker, J.K.; Valenzano, K.J.; Premont, R.T. Vasoactive intestinal polypeptide type-1 receptor regulation. Desensitization, phosphorylation, and sequestration. J. Biol. Chem. 2002, 277, 25519–25526. [Google Scholar] [CrossRef]
- Langer, I. Mechanisms involved in VPAC receptors activation and regulation: Lessons from pharmacological and mutagenesis studies. Front. Endocrinol. 2012, 3, 129. [Google Scholar] [CrossRef]
- Langlet, C.; Gaspard, N.; Nachtergael, I.; Robberecht, P.; Langer, I. Comparative efficacy of VIP and analogs on activation and internalization of the recombinant VPAC2 receptor expressed in CHO cells. Peptides 2004, 25, 2079–2086. [Google Scholar] [CrossRef]
- Shintani, Y.; Hayata-Takano, A.; Moriguchi, K.; Nakazawa, T.; Ago, Y.; Kasai, A.; Seiriki, K.; Shintani, N.; Hashimoto, H. β-Arrestin1 and 2 differentially regulate PACAP-induced PAC1 receptor signaling and trafficking. PLoS ONE 2018, 13, e0196946. [Google Scholar] [CrossRef]
- Gupte, R.P.; Kadunganattil, S.; Shepherd, A.J.; Merrill, R.; Planer, W.; Bruchas, M.R.; Strack, S.; Mohapatra, D.P. Convergent phosphomodulation of the major neuronal dendritic potassium channel Kv4.2 by pituitary adenylate cyclase-activating polypeptide. Neuropharmacology 2016, 101, 291–308. [Google Scholar] [CrossRef]
- Moldovan Loomis, C.; Dutzar, B.; Ojala, E.W.; Hendrix, L.; Karasek, C.; Scalley-Kim, M.; Mulligan, J.; Fan, P.; Billgren, J.; Rubin, V.; et al. Pharmacologic Characterization of ALD1910, a Potent Humanized Monoclonal Antibody against the Pituitary Adenylate Cyclase-Activating Peptide. J. Pharmacol. Exp. Ther. 2019, 369, 26–36. [Google Scholar] [CrossRef]
- Hoffmann, J.; Miller, S.; Martins-Oliveira, M.; Akerman, S.; Supronsinchai, W.; Sun, H.; Shi, L.; Wang, J.; Zhu, D.; Lehto, S.; et al. PAC1 receptor blockade reduces central nociceptive activity: New approach for primary headache? Pain 2020, 161, 1670–1681. [Google Scholar] [CrossRef]
- Rustichelli, C.; Lo Castro, F.; Baraldi, C.; Ferrari, A. Targeting pituitary adenylate cyclase-activating polypeptide (PACAP) with monoclonal antibodies in migraine prevention: A brief review. Expert Opin. Investig. Drugs 2020, 29, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Ashina, M.; Doležil, D.; Bonner, J.H.; Zhou, L.; Klatt, J.; Picard, H.; Mikol, D.D. A phase 2, randomized, double-blind, placebo-controlled trial of AMG 301, a pituitary adenylate cyclase-activating polypeptide PAC1 receptor monoclonal antibody for migraine prevention. Cephalalgia Int. J. Headache 2021, 41, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Cui, Z.; Li, M.; Yang, Y.; Zhong, J. Dimer-dependent intrinsic/basal activity of the class B G protein-coupled receptor PAC1 promotes cellular anti-apoptotic activity through Wnt/β-catenin pathways that are associated with dimer endocytosis. PLoS ONE 2014, 9, e113913. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Zhong, J.; Li, M.; Guo, X.; Zhang, H.; Chen, J. PACAP induces the dimerization of PAC1 on the nucleus associated with the cAMP increase in the nucleus. Neurosci. Lett. 2013, 549, 92–96. [Google Scholar] [CrossRef]
- Feher, M.; Gaszner, B.; Tamas, A.; Gil-Martinez, A.L.; Fernandez-Villalba, E.; Herrero, M.T.; Reglodi, D. Alteration of the PAC1 Receptor Expression in the Basal Ganglia of MPTP-Induced Parkinsonian Macaque Monkeys. Neurotox. Res. 2018, 33, 702–715. [Google Scholar] [CrossRef]
- Miura, A.; Kambe, Y.; Inoue, K.; Tatsukawa, H.; Kurihara, T.; Griffin, M.; Kojima, S.; Miyata, A. Pituitary adenylate cyclase-activating polypeptide type 1 receptor (PAC1) gene is suppressed by transglutaminase 2 activation. J. Biol. Chem. 2013, 288, 32720–32730. [Google Scholar] [CrossRef]
- Kenakin, T. Functional selectivity and biased receptor signaling. J. Pharmacol. Exp. Ther. 2011, 336, 296–302. [Google Scholar] [CrossRef]
- Kenakin, T.; Christopoulos, A. Signalling bias in new drug discovery: Detection, quantification and therapeutic impact. Nat. Rev. Drug Discov. 2013, 12, 205–216. [Google Scholar] [CrossRef]
- Wootten, D.; Miller, L.J.; Koole, C.; Christopoulos, A.; Sexton, P.M. Allostery and Biased Agonism at Class B G Protein-Coupled Receptors. Chem. Rev. 2017, 117, 111–138. [Google Scholar] [CrossRef]
- Koole, C.; Wootten, D.; Simms, J.; Valant, C.; Sridhar, R.; Woodman, O.L.; Miller, L.J.; Summers, R.J.; Christopoulos, A.; Sexton, P.M. Allosteric ligands of the glucagon-like peptide 1 receptor (GLP-1R) differentially modulate endogenous and exogenous peptide responses in a pathway-selective manner: Implications for drug screening. Mol. Pharmacol. 2010, 78, 456–465. [Google Scholar] [CrossRef]
- Liang, Y.-L.; Khoshouei, M.; Radjainia, M.; Zhang, Y.; Glukhova, A.; Tarrasch, J.; Thal, D.M.; Furness, S.G.B.; Christopoulos, G.; Coudrat, T.; et al. Phase-plate cryo-EM structure of a class B GPCR–G-protein complex. Nature 2017, 546, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.G.F.; DeVree, B.T.; Zou, Y.; Kruse, A.C.; Chung, K.Y.; Kobilka, T.S.; Thian, F.S.; Chae, P.S.; Pardon, E.; Calinski, D.; et al. Crystal structure of the β2 adrenergic receptor-Gs protein complex. Nature 2011, 477, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.-L.; Zhao, P.; Draper-Joyce, C.; Baltos, J.-A.; Glukhova, A.; Truong, T.T.; May, L.T.; Christopoulos, A.; Wootten, D.; Sexton, P.M.; et al. Dominant Negative G Proteins Enhance Formation and Purification of Agonist-GPCR-G Protein Complexes for Structure Determination. ACS Pharmacol. Transl. Sci. 2018, 1, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Koehl, A.; Matile, H.; Hu, H.; Hilger, D.; Schertler, G.F.X.; Manglik, A.; Skiniotis, G.; Dawson, R.J.P.; Kobilka, B.K. Development of an antibody fragment that stabilizes GPCR/G-protein complexes. Nat. Commun. 2018, 9, 3712. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.; Okashah, N.; Inoue, A.; Nehmé, R.; Carpenter, B.; Tate, C.G.; Lambert, N.A. Mini G protein probes for active G protein-coupled receptors (GPCRs) in live cells. J. Biol. Chem. 2018, 293, 7466–7473. [Google Scholar] [CrossRef] [PubMed]
- Danev, R.; Baumeister, W. Cryo-EM single particle analysis with the Volta phase plate. eLife 2016, 5, e13046. [Google Scholar] [CrossRef]
- Danev, R.; Belousoff, M.; Liang, Y.L.; Zhang, X.; Eisenstein, F.; Wootten, D.; Sexton, P.M. Routine sub-2.5 Å cryo-EM structure determination of GPCRs. Nat. Commun. 2021, 12, 4333. [Google Scholar] [CrossRef]
- Zhang, X.; Johnson, R.M.; Drulyte, I.; Yu, L.; Kotecha, A.; Danev, R.; Wootten, D.; Sexton, P.M.; Belousoff, M.J. Evolving cryo-EM structural approaches for GPCR drug discovery. Structure 2021, 29, 963–974. [Google Scholar] [CrossRef]
- Wu, M.; Lander, G.C. Present and Emerging Methodologies in Cryo-EM Single-Particle Analysis. Biophys. J. 2020, 119, 1281–1289. [Google Scholar] [CrossRef]
- Nygaard, R.; Kim, J.; Mancia, F. Cryo-electron microscopy analysis of small membrane proteins. Curr. Opin. Struct. Biol. 2020, 64, 26–33. [Google Scholar] [CrossRef]
- Nakane, T.; Kotecha, A.; Sente, A.; McMullan, G.; Masiulis, S.; Brown, P.M.G.E.; Grigoras, I.T.; Malinauskaite, L.; Malinauskas, T.; Miehling, J.; et al. Single-particle cryo-EM at atomic resolution. Nature 2020, 587, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Yip, K.M.; Fischer, N.; Paknia, E.; Chari, A.; Stark, H. Atomic-resolution protein structure determination by cryo-EM. Nature 2020, 587, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Parthier, C.; Kleinschmidt, M.; Neumann, P.; Rudolph, R.; Manhart, S.; Schlenzig, D.; Fanghänel, J.; Rahfeld, J.U.; Demuth, H.U.; Stubbs, M.T. Crystal structure of the incretin-bound extracellular domain of a G protein-coupled receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 13942–13947. [Google Scholar] [CrossRef] [PubMed]
- Runge, S.; Thøgersen, H.; Madsen, K.; Lau, J.; Rudolph, R. Crystal structure of the ligand-bound glucagon-like peptide-1 receptor extracellular domain. J. Biol. Chem. 2008, 283, 11340–11347. [Google Scholar] [CrossRef] [PubMed]
- Pike, A.C.W.; Barr, A.J.; Quigley, A.; Burgess Brown, N.; de Riso, A.; Bullock, A.; Berridge, G.; Muniz, J.R.C.; Chaikaud, A.; Vollmar, M.; et al. Crystal Structure of the Extracellular Domain of Human Vasoactive Intestinal Polypeptide Receptor 2. 2010. Available online: https://www.wwpdb.org/pdb?id=pdb_00002x57 (accessed on 22 June 2022).
- Kumar, S.; Pioszak, A.; Zhang, C.; Swaminathan, K.; Xu, H.E. Crystal structure of the PAC1R extracellular domain unifies a consensus fold for hormone recognition by class B G-protein coupled receptors. PLoS ONE 2011, 6, e19682. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Song, D.; Davis-Taber, R.A.; Barrett, L.W.; Scott, V.E.; Richardson, P.L.; Pereda-Lopez, A.; Uchic, M.E.; Solomon, L.R.; Lake, M.R.; et al. Solution Structure and Mutational Analysis of Pituitary Adenylate Cyclase-Activating Polypeptide Binding to the Extracellular Domain of PAC1-Rs. Proc. Natl. Acad. Sci. USA 2007, 104, 7875–7880. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Shen, Q.; Zhao, L.-H.; Mao, C.; Zhou, X.E.; Shen, D.-D.; de Waal, P.W.; Bi, P.; Li, C.; Jiang, Y.; et al. Molecular Basis for Hormone Recognition and Activation of Corticotropin-Releasing Factor Receptors. Mol. Cell 2020, 77, 669–680.e664. [Google Scholar] [CrossRef]
- Zhang, X.; Belousoff, M.J.; Zhao, P.; Kooistra, A.J.; Truong, T.T.; Ang, S.Y.; Underwood, C.R.; Egebjerg, T.; Šenel, P.; Stewart, G.D.; et al. Differential GLP-1R Binding and Activation by Peptide and Non-peptide Agonists. Mol. Cell 2020, 80, 485–500.e487. [Google Scholar] [CrossRef]
- Sun, W.; Chen, L.-N.; Zhou, Q.; Zhao, L.-H.; Yang, D.; Zhang, H.; Cong, Z.; Shen, D.-D.; Zhao, F.; Zhou, F.; et al. A unique hormonal recognition feature of the human glucagon-like peptide-2 receptor. Cell Res. 2020, 30, 1098–1108. [Google Scholar] [CrossRef]
- Zhao, F.; Zhang, C.; Zhou, Q.; Hang, K.; Zou, X.; Chen, Y.; Wu, F.; Rao, Q.; Dai, A.; Yin, W.; et al. Structural insights into hormone recognition by the human glucose-dependent insulinotropic polypeptide receptor. eLife 2021, 10, e68719. [Google Scholar] [CrossRef]
- Chang, R.; Zhang, X.; Qiao, A.; Dai, A.; Belousoff, M.J.; Tan, Q.; Shao, L.; Zhong, L.; Lin, G.; Liang, Y.L.; et al. Cryo-electron microscopy structure of the glucagon receptor with a dual-agonist peptide. J. Biol. Chem. 2020, 295, 9313–9325. [Google Scholar] [CrossRef]
- Dong, M.; Deganutti, G.; Piper, S.J.; Liang, Y.-L.; Khoshouei, M.; Belousoff, M.J.; Harikumar, K.G.; Reynolds, C.A.; Glukhova, A.; Furness, S.G.B.; et al. Structure and dynamics of the active Gs-coupled human secretin receptor. Nat. Commun. 2020, 11, 4137. [Google Scholar] [CrossRef]
- Liang, Y.-L.; Khoshouei, M.; Deganutti, G.; Glukhova, A.; Koole, C.; Peat, T.S.; Radjainia, M.; Plitzko, J.M.; Baumeister, W.; Miller, L.J.; et al. Cryo-EM structure of the active, Gs-protein complexed, human CGRP receptor. Nature 2018, 561, 492–497. [Google Scholar] [CrossRef]
- Cao, J.; Belousoff, M.J.; Liang, Y.-L.; Johnson, R.M.; Josephs, T.M.; Fletcher, M.M.; Christopoulos, A.; Hay, D.L.; Danev, R.; Wootten, D.; et al. A structural basis for amylin receptor phenotype. Science 2022, 375, eabm9609. [Google Scholar] [CrossRef]
- Liang, Y.-L.; Belousoff, M.J.; Fletcher, M.M.; Zhang, X.; Khoshouei, M.; Deganutti, G.; Koole, C.; Furness, S.G.B.; Miller, L.J.; Hay, D.L.; et al. Structure and Dynamics of Adrenomedullin Receptors AM1 and AM2 Reveal Key Mechanisms in the Control of Receptor Phenotype by Receptor Activity-Modifying Proteins. ACS Pharmacol. Transl. Sci. 2020, 3, 263–284. [Google Scholar] [CrossRef]
- Zhao, L.H.; Ma, S.; Sutkeviciute, I.; Shen, D.D.; Zhou, X.E.; de Waal, P.W.; Li, C.Y.; Kang, Y.; Clark, L.J.; Jean-Alphonse, F.G.; et al. Structure and dynamics of the active human parathyroid hormone receptor-1. Science 2019, 364, 148–153. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, X.; Zhao, L.; Wang, Y.; Ye, C.; Zou, X.; Dai, A.; Cong, Z.; Chen, J.; Zhou, Q.; et al. Molecular insights into differentiated ligand recognition of the human parathyroid hormone receptor 2. Proc. Natl. Acad. Sci. USA 2021, 118, e2101279118. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. A Publ. Protein Soc. 2021, 30, 70–82. [Google Scholar] [CrossRef]
- Goddard, T.D.; Huang, C.C.; Meng, E.C.; Pettersen, E.F.; Couch, G.S.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Sci. A Publ. Protein Soc. 2018, 27, 14–25. [Google Scholar] [CrossRef]
- Doan, N.-D.; Bourgault, S.; Dejda, A.; Létourneau, M.; Detheux, M.; Vaudry, D.; Vaudry, H.; Chatenet, D.; Fournier, A. Design and in vitro characterization of PAC1/VPAC1-selective agonists with potent neuroprotective effects. Biochem. Pharmacol. 2011, 81, 552–561. [Google Scholar] [CrossRef]
- Bourgault, S.; Vaudry, D.; Ségalas-Milazzo, I.; Guilhaudis, L.; Couvineau, A.; Laburthe, M.; Vaudry, H.; Fournier, A. Molecular and Conformational Determinants of Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) for Activation of the PAC1 Receptor. J. Med. Chem. 2009, 52, 3308–3316. [Google Scholar] [CrossRef] [PubMed]
- Wootten, D.; Simms, J.; Miller, L.J.; Christopoulos, A.; Sexton, P.M. Polar transmembrane interactions drive formation of ligand-specific and signal pathway-biased family B G protein-coupled receptor conformations. Proc. Natl. Acad. Sci. USA 2013, 110, 5211–5216. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, H.; Ito, T.; Pradhan, T.K.; Mantey, S.A.; Hou, W.; Coy, D.H.; Jensen, R.T. Elucidation of the Vasoactive Intestinal Peptide Pharmacophore for VPAC2 Receptors in Human and Rat and Comparison to the Pharmacophore for VPAC1 Receptors. J. Pharmacol. Exp. Ther. 2002, 303, 445–460. [Google Scholar] [CrossRef]
- Liao, C.; de Molliens, M.P.; Schneebeli, S.T.; Brewer, M.; Song, G.; Chatenet, D.; Braas, K.M.; May, V.; Li, J. Targeting the PAC1 Receptor for Neurological and Metabolic Disorders. Curr. Top. Med. Chem. 2019, 19, 1399–1417. [Google Scholar] [CrossRef] [PubMed]
- Bourgault, S.; Vaudry, D.; Dejda, A.; Doan, D.N.; Vaudry, H.; Fournier, A. Pituitary Adenylate Cyclase-Activating Polypeptide: Focus on Structure- Activity Relationships of a Neuroprotective Peptide. Curr. Med. Chem. 2009, 16, 4462–4480. [Google Scholar] [CrossRef] [PubMed]
- Bourgault, S.; Chatenet, D.; Wurtz, O.; Doan, N.D.; Leprince, J.; Vaudry, H.; Fournier, A.; Vaudry, D. Strategies to convert PACAP from a hypophysiotropic neurohormone into a neuroprotective drug. Curr. Pharm. Des. 2011, 17, 1002–1024. [Google Scholar] [CrossRef]
- Schäfer, H.; Zheng, J.; Morys-Wortmann, C.; Fölsch, U.R.; Schmidt, W.E. Structural motifs of pituitary adenylate cyclase-activating polypeptide (PACAP) defining PAC1-receptor selectivity. Regul. Pept. 1999, 79, 83–92. [Google Scholar] [CrossRef]
- Wei, M.; Fujiki, K.; Ando, E.; Zhang, S.; Ozaki, T.; Ishiguro, H.; Kondo, T.; Nokihara, K.; Wray, V.; Naruse, S. Identification of key residues that cause differential gallbladder response to PACAP and VIP in the guinea pig. Am. J. Physiol.-Gastrointest. Liver Physiol. 2007, 292, G76–G83. [Google Scholar] [CrossRef]
- Ando, E.; Nokihara, K.; Naruse, S.; Wray, V. Recognition of pituitary adenylate cyclase-activating polypeptide/vasoactive intestinal polypeptide (PACAP/VIP) hybrids and related peptides by rat brain membranes. Biomed. Pept. Proteins Nucleic Acids Struct. Synth. Biol. Act. 1996, 2, 41–46. [Google Scholar]
- Reddy, V.B.; Iuga, A.O.; Kounga, K.; Lerner, E.A. Functional analysis of recombinant mutants of maxadilan with a PAC1 receptor-expressing melanophore cell line. J. Biol. Chem. 2006, 281, 16197–16201. [Google Scholar] [CrossRef]
- Lerner, E.A.; Iuga, A.O.; Reddy, V.B. Maxadilan, a PAC1 receptor agonist from sand flies. Peptides 2007, 28, 1651–1654. [Google Scholar] [CrossRef]
- Moro, O.; Wakita, K.; Ohnuma, M.; Denda, S.; Lerner, E.A.; Tajima, M. Functional Characterization of Structural Alterations in the Sequence of the Vasodilatory Peptide Maxadilan Yields a Pituitary Adenylate Cyclase-activating Peptide Type 1 Receptor-specific Antagonist. J. Biol. Chem. 1999, 274, 23103–23110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, B.; Feng, D.; Hu, H.; Chu, M.; Qu, Q.; Tarrasch, J.T.; Li, S.; Sun Kobilka, T.; Kobilka, B.K.; et al. Cryo-EM structure of the activated GLP-1 receptor in complex with a G protein. Nature 2017, 546, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Wootten, D.; Reynolds, C.A.; Koole, C.; Smith, K.J.; Mobarec, J.C.; Simms, J.; Quon, T.; Coudrat, T.; Furness, S.G.B.; Miller, L.J.; et al. A Hydrogen-Bonded Polar Network in the Core of the Glucagon-Like Peptide-1 Receptor Is a Fulcrum for Biased Agonism: Lessons from Class B Crystal Structures. Mol. Pharmacol. 2016, 89, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Wootten, D.; Savage, E.E.; Willard, F.S.; Bueno, A.B.; Sloop, K.W.; Christopoulos, A.; Sexton, P.M. Differential Activation and Modulation of the Glucagon-Like Peptide-1 Receptor by Small Molecule Ligands. Mol. Pharmacol. 2013, 83, 822–834. [Google Scholar] [CrossRef]
- Hollenstein, K.; Kean, J.; Bortolato, A.; Cheng, R.K.; Doré, A.S.; Jazayeri, A.; Cooke, R.M.; Weir, M.; Marshall, F.H. Structure of class B GPCR corticotropin-releasing factor receptor 1. Nature 2013, 499, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Jazayeri, A.; Doré, A.S.; Lamb, D.; Krishnamurthy, H.; Southall, S.M.; Baig, A.H.; Bortolato, A.; Koglin, M.; Robertson, N.J.; Errey, J.C.; et al. Extra-helical binding site of a glucagon receptor antagonist. Nature 2016, 533, 274–277. [Google Scholar] [CrossRef]
- Siu, F.Y.; He, M.; de Graaf, C.; Han, G.W.; Yang, D.; Zhang, Z.; Zhou, C.; Xu, Q.; Wacker, D.; Joseph, J.S.; et al. Structure of the human glucagon class B G-protein-coupled receptor. Nature 2013, 499, 444–449. [Google Scholar] [CrossRef]
- Wu, F.; Yang, L.; Hang, K.; Laursen, M.; Wu, L.; Han, G.W.; Ren, Q.; Roed, N.K.; Lin, G.; Hanson, M.A.; et al. Full-length human GLP-1 receptor structure without orthosteric ligands. Nat. Commun. 2020, 11, 1272. [Google Scholar] [CrossRef]
- Deganutti, G.; Liang, Y.-L.; Zhang, X.; Khoshouei, M.; Clydesdale, L.; Belousoff, M.J.; Venugopal, H.; Truong, T.T.; Glukhova, A.; Keller, A.N.; et al. Dynamics of GLP-1R peptide agonist engagement are correlated with kinetics of G protein activation. Nat. Commun. 2022, 13, 92. [Google Scholar] [CrossRef]
- Krumm, B.; Roth, B.L. A Structural Understanding of Class B GPCR Selectivity and Activation Revealed. Structure 2020, 28, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Do, H.N.; Haldane, A.; Levy, R.M.; Miao, Y. Unique features of different classes of G-protein-coupled receptors revealed from sequence coevolutionary and structural analysis. Proteins Struct. Funct. Bioinform. 2022, 90, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Liang, Y.-L.; Belousoff, M.J.; Deganutti, G.; Fletcher, M.M.; Willard, F.S.; Bell, M.G.; Christe, M.E.; Sloop, K.W.; Inoue, A.; et al. Activation of the GLP-1 receptor by a non-peptidic agonist. Nature 2020, 577, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Venkatakrishnan, A.J.; Ma, A.K.; Fonseca, R.; Latorraca, N.R.; Kelly, B.; Betz, R.M.; Asawa, C.; Kobilka, B.K.; Dror, R.O. Diverse GPCRs exhibit conserved water networks for stabilization and activation. Proc. Natl. Acad. Sci. USA 2019, 116, 3288–3293. [Google Scholar] [CrossRef]
- Wootten, D.; Miller, L.J. Structural Basis for Allosteric Modulation of Class B G Protein-Coupled Receptors. Annu. Rev Pharm. Toxicol 2020, 60, 89–107. [Google Scholar] [CrossRef]
- Christopoulos, A.; Kenakin, T. G protein-coupled receptor allosterism and complexing. Pharmacol. Rev. 2002, 54, 323–374. [Google Scholar] [CrossRef]
- Langer, I.; Latek, D. Drug Repositioning For Allosteric Modulation of VIP and PACAP Receptors. Front. Endocrinol. 2021, 12, 711906. [Google Scholar] [CrossRef]
- Latek, D.; Langer, I.; Krzysko, K.; Charzewski, L. A Molecular Dynamics Study of Vasoactive Intestinal Peptide Receptor 1 and the Basis of Its Therapeutic Antagonism. Int. J. Mol. Sci. 2019, 20, 4348. [Google Scholar] [CrossRef]
- Song, G.; Yang, D.; Wang, Y.; de Graaf, C.; Zhou, Q.; Jiang, S.; Liu, K.; Cai, X.; Dai, A.; Lin, G.; et al. Human GLP-1 receptor transmembrane domain structure in complex with allosteric modulators. Nature 2017, 546, 312–315. [Google Scholar] [CrossRef]
- Zhang, H.; Qiao, A.; Yang, D.; Yang, L.; Dai, A.; de Graaf, C.; Reedtz-Runge, S.; Dharmarajan, V.; Zhang, H.; Han, G.W.; et al. Structure of the full-length glucagon class B G-protein-coupled receptor. Nature 2017, 546, 259–264. [Google Scholar] [CrossRef]
- Yu, R.; Zheng, L.; Cui, Y.; Zhang, H.; Ye, H. Doxycycline exerted neuroprotective activity by enhancing the activation of neuropeptide GPCR PAC1. Neuropharmacology 2016, 103, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Wang, L.; Li, J.; Huang, X.; Yu, R. The allosteric modulation effects of doxycycline, minocycline, and their derivatives on the neuropeptide receptor PAC1-R. Acta Biochim. Biophys. Sin. 2019, 51, 627. [Google Scholar] [CrossRef] [PubMed]
- Punjani, A.; Fleet, D.J. 3D variability analysis: Resolving continuous flexibility and discrete heterogeneity from single particle cryo-EM. J. Struct. Biol. 2021, 213, 107702. [Google Scholar] [CrossRef]
- Cary, B.P.; Deganutti, G.; Zhao, P.; Truong, T.T.; Piper, S.J.; Liu, X.; Belousoff, M.J.; Danev, R.; Sexton, P.M.; Wootten, D.; et al. Structural and functional diversity among agonist-bound states of the GLP-1 receptor. Nat. Chem. Biol. 2022, 18, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Deganutti, G.; Atanasio, S.; Rujan, R.-M.; Sexton, P.M.; Wootten, D.; Reynolds, C.A. Exploring Ligand Binding to Calcitonin Gene-Related Peptide Receptors. Front. Mol. Biosci. 2021, 8, 720561. [Google Scholar] [CrossRef]
- Josephs, T.M.; Belousoff, M.J.; Liang, Y.-L.; Piper, S.J.; Cao, J.; Garama, D.J.; Leach, K.; Gregory, K.J.; Christopoulos, A.; Hay, D.L.; et al. Structure and dynamics of the CGRP receptor in apo and peptide-bound forms. Science 2021, 372, eabf7258. [Google Scholar] [CrossRef]
- Asher, W.B.; Geggier, P.; Holsey, M.D.; Gilmore, G.T.; Pati, A.K.; Meszaros, J.; Terry, D.S.; Mathiasen, S.; Kaliszewski, M.J.; McCauley, M.D.; et al. Single-molecule FRET imaging of GPCR dimers in living cells. Nat. Methods 2021, 18, 397–405. [Google Scholar] [CrossRef]
- Dijkman, P.M.; Muñoz-García, J.C.; Lavington, S.R.; Kumagai, P.S.; Reis, R.I.d.; Yin, D.; Stansfeld, P.J.; Costa-Filho, A.J.; Watts, A. Conformational dynamics of a G protein–coupled receptor helix 8 in lipid membranes. Sci. Adv. 2020, 6, eaav8207. [Google Scholar] [CrossRef] [PubMed]
- Eps, N.V.; Altenbach, C.; Caro, L.N.; Latorraca, N.R.; Hollingsworth, S.A.; Dror, R.O.; Ernst, O.P.; Hubbell, W.L. Gi and Gs-coupled GPCRs show different modes of G-protein binding. Proc. Natl. Acad. Sci. USA 2018, 115, 2383–2388. [Google Scholar] [CrossRef]
- Yeliseev, A.A.; Zoretich, K.; Hooper, L.; Teague, W.; Zoubak, L.; Hines, K.G.; Gawrisch, K. Site-selective labeling and electron paramagnetic resonance studies of human cannabinoid receptor CB(2). Biochim. Biophys. Acta Biomembr. 2021, 1863, 183621. [Google Scholar] [CrossRef]
- Robertson, M.J.; Papasergi-Scott, M.; He, F.; Seven, A.B.; Meyerowitz, J.G.; Panova, O.; Peroto, M.C.; Che, T.; Skiniotis, G. Structure Determination of Inactive-State GPCRs with a Universal Nanobody. bioRxiv 2022, 2021, 466983. [Google Scholar] [CrossRef]
- Lei, S.; Clydesdale, L.; Dai, A.; Cai, X.; Feng, Y.; Yang, D.; Liang, Y.-L.; Koole, C.; Zhao, P.; Coudrat, T.; et al. Two distinct domains of the glucagon-like peptide-1 receptor control peptide-mediated biased agonism. J. Biol. Chem. 2018, 293, 9370–9387. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).