
Perivascular Mesenchymal Stem/Stromal Cells, an Immune Privileged Niche for Viruses?

 , , , , and
, , , , and
Abstract
1. Introduction
2. Cell Biology and Immune Functions of Mesenchymal Stem Cell (MSC)
2.1. Definition of MSC
2.2. Origins of Pericytes/Perivascular MSC-Fibroblasts Derived from the Neural Crest and/or Mesoderm Embryonic Tissues
2.3. Role of MSC in Health (Immunoregulatory Activities and Tissue Repair) and Diseases (Fibrosis)
2.4. MSCs’ Pathological Contributions (Fibrosis and Vessel Calcification)
3. MSC Viral Infection and Host Responses
3.1. MSCs a Gatekeper and/or Reservoir of Viruses
3.1.1. Bone Marrow-Derived MSC (BM-MSC)
3.1.2. Lung Resident-MSC and Viruses
3.1.3. Adipose Stem Cells and Viruses
3.1.4. MSC of the Liver: Hepatic Stellate Cell/Ito Cell and Viruses
3.1.5. MSC of the Kidney: Mesangial Cell and Viruses
3.1.6. Brain MSC: Brain Pericytes and Viruses
3.2. Osteoblasts and Viral Infections
3.3. Schwann Cell of Peripheral Nerves and Viral Infection
4. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AIDS | Acquired Immunodeficiency Syndrome |
| ALP | Alkaline Phosphatase |
| BBB | Blood Brain Barrier |
| BM-MSC | Bone Marrow MSC |
| BP | Brain Pericyte |
| CHIKV | Chikungunya Virus |
| CNS | Central Nervous System |
| DENV | Dengue Virus |
| EV | Extracellular Vesicle |
| GBS | Guillain-Barré Syndrome |
| GFAP | Glial Fibrillary Acidic Protein |
| HCMV | Human Cytomegalovirus |
| HCV | Hepatitis C Virus |
| HHV | Human Herpes Virus |
| HIV | Human Immunodeficiency Virus |
| HepSC | Hepatic Stellate Cell |
| HSC | Hematopoietic stem cells |
| HSV | Herpes Simplex Virus |
| IDO | Indoleamine 2 3-dioxygenase |
| IFN | Interferon |
| IFNAR | Interferon alpha receptor |
| IRF | Interferon Regulated Factor |
| ISG | Interferon Stimulated Gene |
| JEV | Japanese Encephalitis Virus |
| MC | Mesangial Cell |
| MeV | Measle morbillivirus |
| MSC | Multipotent Mesenchymal Stem/stromal Cell |
| OB | Osteoblast |
| OPG | Osteoprotegerin |
| PGE2 | Prostaglandin E2 |
| Poly I:C | Polyinosinic:polycytidylic acid |
| PRR | Pattern Recognition Receptor |
| RANKL | Receptor Activator of Nuclear factor-KappaB Ligand |
| RIG-I | Retinoic acid-Inducible Gene-I |
| RRV | Ross River Virus |
| SC | Schwann Cell |
| TGF-β | Transforming Growth Factor β |
| TLR | Toll-Like Receptor |
| TNF | Tumor Necrosis Factor |
| VEGF | Vascular Endothelial Growth Factor |
| VZV | Varicella-Zoster Virus |
| ZIKV | Zika Virus |
References
- Caplan, A. Mesenchymal Stem-Cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I.; Dennis, J.E. Mesenchymal Stem Cells as Trophic Mediators. J. Cell. Biochem. 2006, 98, 1076–1084. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Salazar, M.; Gonzalez-Galofre, Z.N.; Casamitjana, J.; Crisan, M.; James, A.W.; Peault, B. Five Decades Later, Are Mesenchymal Stem Cells Still Relevant? Front. Bioeng. Biotechnol. 2020, 8, 148. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Meirelles, L.; Caplan, A.I.; Nardi, N.B. In Search of the In Vivo Identity of Mesenchymal Stem Cells. Stem Cells 2008, 26, 2287–2299. [Google Scholar] [CrossRef] [PubMed]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.-W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A Perivascular Origin for Mesenchymal Stem Cells in Multiple Human Organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef]
- Da Silva Meirelles, L.; Chagastelles, P.C.; Nardi, N.B. Mesenchymal Stem Cells Reside in Virtually All Post-Natal Organs and Tissues. J. Cell Sci. 2006, 119, 2204–2213. [Google Scholar] [CrossRef]
- Bedoui, Y.; Lebeau, G.; Guillot, X.; Dargai, F.; Guiraud, P.; Neal, J.W.; Ralandison, S.; Gasque, P. Emerging Roles of Perivascular Mesenchymal Stem Cells in Synovial Joint Inflammation. J. Neuroimmune Pharmacol. 2020, 15, 838–851. [Google Scholar] [CrossRef]
- Deans, R.J.; Moseley, A.B. Mesenchymal Stem Cells: Biology and Potential Clinical Uses. Exp. Hematol. 2000, 28, 875–884. [Google Scholar] [CrossRef]
- El Agha, E.; Kramann, R.; Schneider, R.K.; Li, X.; Seeger, W.; Humphreys, B.D.; Bellusci, S. Mesenchymal Stem Cells in Fibrotic Disease. Cell Stem Cell 2017, 21, 166–177. [Google Scholar] [CrossRef]
- Le Blanc, K.; Mougiakakos, D. Multipotent Mesenchymal Stromal Cells and the Innate Immune System. Nat. Rev. Immunol. 2012, 12, 383–396. [Google Scholar] [CrossRef]
- Armulik, A.; Genove, G.; Betsholtz, C. Pericytes: Developmental, Physiological, and Pathological Perspectives, Problems, and Promises. Dev. Cell 2011, 21, 193–215. [Google Scholar] [CrossRef] [PubMed]
- Asada, N.; Takase, M.; Nakamura, J.; Oguchi, A.; Asada, M.; Suzuki, N.; Yamarnura, K.; Nagoshi, N.; Shibata, S.; Rao, T.N.; et al. Dysfunction of Fibroblasts of Extrarenal Origin Underlies Renal Fibrosis and Renal Anemia in Mice. J. Clin. Investig. 2011, 121, 3981–3990. [Google Scholar] [CrossRef] [PubMed]
- Cassiman, D.; Libbrecht, L.; Desmet, V.; Denef, C.; Roskams, T. Hepatic Stellate Cell/Myofibroblast Subpopulations in Fibrotic Human and Rat Livers. J. Hepatol. 2002, 36, 200–209. [Google Scholar] [CrossRef]
- Duffield, J.S. Cellular and Molecular Mechanisms in Kidney Fibrosis. J. Clin. Investig. 2014, 124, 2299–2306. [Google Scholar] [CrossRef]
- Dupin, E.; Sommer, L. Neural Crest Progenitors and Stem Cells: From Early Development to Adulthood. Dev. Biol. 2012, 366, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Furlan, A.; Adameyko, I. Schwann Cell Precursor: A Neural Crest Cell in Disguise? Dev. Biol. 2018, 444, S25–S35. [Google Scholar] [CrossRef]
- Isern, J.; Garcia-Garcia, A.; Martin, A.M.; Arranz, L.; Martin-Perez, D.; Torroja, C.; Sanchez-Csabo, F.; Mendez-Ferrer, S. The Neural Crest Is a Source of Mesenchymal Stem Cells with Specialized Hematopoietic Stem-Cell-Niche Function. eLife 2014, 3, e03696. [Google Scholar] [CrossRef]
- Neubauer, K.; Knittel, T.; Aurisch, S.; Fellmer, P.; Ramadori, G. Glial Fibrillary Acidic Protein—A Cell Type Specific Marker for Ito Cells in Vivo and in Vitro. J. Hepatol. 1996, 24, 719–730. [Google Scholar] [CrossRef]
- Parfejevs, V.; Antunes, A.T.; Sommer, L. Injury and Stress Responses of Adult Neural Crest-Derived Cells. Dev. Biol. 2018, 444, S356–S365. [Google Scholar] [CrossRef]
- Caplan, A.I. New MSC: MSCs as Pericytes Are Sentinels and Gatekeepers. J. Orthop. Res. 2017, 35, 1151–1159. [Google Scholar] [CrossRef]
- Naji, A.; Eitoku, M.; Favier, B.; Deschaseaux, F.; Rouas-Freiss, N.; Suganuma, N. Biological Functions of Mesenchymal Stem Cells and Clinical Implications. Cell. Mol. Life Sci. 2019, 76, 3323–3348. [Google Scholar] [CrossRef] [PubMed]
- Duffield, J.S.; Lupher, M.; Thannickal, V.J.; Wynn, T.A. Host Responses in Tissue Repair and Fibrosis. Annu. Rev. Pathol. Mech. Dis. 2013, 8, 241–276. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L.; Sheppard, D.; Duffield, J.S.; Violette, S. Therapy for Fibrotic Diseases: Nearing the Starting Line. Sci. Transl. Med. 2013, 5, 167sr1. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A. Cellular and Molecular Mechanisms of Fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Kheirollahi, V.; Wasnick, R.M.; Biasin, V.; Vazquez-Armendariz, A.I.; Chu, X.; Moiseenko, A.; Weiss, A.; Wilhelm, J.; Zhang, J.-S.; Kwapiszewska, G.; et al. Metformin Induces Lipogenic Differentiation in Myofibroblasts to Reverse Lung Fibrosis. Nat. Commun. 2019, 10, 2987. [Google Scholar] [CrossRef] [PubMed]
- Rangarajan, S.; Bone, N.B.; Zmijewskal, A.A.; Jiang, S.; Park, D.W.; Bernard, K.; Locy, M.L.; Ravi, S.; Deshane, J.; Mannon, R.B.; et al. Metformin Reverses Established Lung Fibrosis in a Bleomycin Model. Nat. Med. 2018, 24, 1121–1127. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhao, S.; Wang, X.; Teng, Z.; Li, D.; Zeng, Y. HIV-1 P55-Gag Protein Induces Senescence of Human Bone Marrow Mesenchymal Stem Cells and Reduces Their Capacity to Support Expansion of Hematopoietic Stem Cells in Vitro. Cell Biol. Int. 2017, 41, 969–981. [Google Scholar] [CrossRef]
- Mattana, J.; Abramovici, M.; Singhal, P.C. Effects of Human Immunodeficiency Virus Sera and Macrophage Supernatants on Mesangial Cell Proliferation and Matrix Synthesis. Am. J. Pathol. 1993, 143, 814–822. [Google Scholar]
- Kallmeyer, K.; Ryder, M.A.; Pepper, M.S. Mesenchymal Stromal Cells: A Possible Reservoir for HIV-1? Stem Cell Rev. Rep. 2022, 18, 1253–1280. [Google Scholar] [CrossRef]
- Soland, M.A.; Keyes, L.R.; Bayne, R.; Moon, J.; Porada, C.D.; Jeor, S.S.; Almeida-Porada, G. Perivascular Stromal Cells as a Potential Reservoir of Human Cytomegalovirus. Am. J. Transplant. 2014, 14, 820–830. [Google Scholar] [CrossRef]
- Horwitz, E.M.; Le Blanc, K.; Dominici, M.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Deans, R.J.; Krause, D.S.; Keating, A. Clarification of the Nomenclature for MSC: The International Society for Cellular Therapy Position Statement. Cytotherapy 2005, 7, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.; Sheridan, J.; Veiga-Fernandes, H.; Roderick, K.; Pachnis, V.; Adams, R.; Blackburn, C.; Kioussis, D.; Coles, M. Contribution of Neural Crest-Derived Cells in the Embryonic and Adult Thymus. J. Immunol. 2008, 180, 3183–3189. [Google Scholar] [CrossRef] [PubMed]
- Le Douarin, N.; Kalcheim, C. The Neural Crest (Developmental and Cell Biology Series), 2nd ed.; Cambridge University Press: Cambridge, UK, 1999; ISBN 978-0-521-62010-9. [Google Scholar]
- Takashima, Y.; Era, T.; Nakao, K.; Kondo, S.; Kasuga, M.; Smith, A.G.; Nishikawa, S.-I. Neuroepithelial Cells Supply an Initial Transient Wave of MSC Differentiation. Cell 2007, 129, 1377–1388. [Google Scholar] [CrossRef] [PubMed]
- Corselli, M.; Chen, C.-W.; Crisan, M.; Lazzari, L.; Péault, B. Perivascular Ancestors of Adult Multipotent Stem Cells. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1104–1109. [Google Scholar] [CrossRef]
- Singer, N.G.; Caplan, A.I. Mesenchymal Stem Cells: Mechanisms of Inflammation. Annu. Rev. Pathol. Mech. Dis. 2011, 6, 457–478. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage Potential of Adult Human Mesenchymal Stem Cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Gazit, Z.; Pelled, G.; Sheyn, D.; Kimelman, N.; Gazit, D. Mesenchymal Stem Cells. In Principles of Regenerative Medicine; Elsevier: Amsterdam, The Netherlands, 2011; pp. 285–304. ISBN 978-0-12-381422-7. [Google Scholar]
- Chamberlain, G.; Fox, J.; Ashton, B.; Middleton, J. Concise Review: Mesenchymal Stem Cells: Their Phenotype, Differentiation Capacity, Immunological Features, and Potential for Homing. Stem Cells 2007, 25, 2739–2749. [Google Scholar] [CrossRef]
- Phinney, D.G.; Prockop, D.J. Concise Review: Mesenchymal Stem/Multipotent Stromal Cells: The State of Transdifferentiation and Modes of Tissue Repair—Current Views. Stem Cells 2007, 25, 2896–2902. [Google Scholar] [CrossRef]
- Quirici, N.; Soligo, D.; Bossolasco, P.; Servida, F.; Lumini, C.; Deliliers, G.L. Isolation of Bone Marrow Mesenchymal Stem Cells by Anti-Nerve Growth Factor Receptor Antibodies. Exp. Hematol. 2002, 30, 783–791. [Google Scholar] [CrossRef]
- Álvarez-Viejo, M.; Menéndez-Menéndez, Y.; Otero-Hernández, J. CD271 as a Marker to Identify Mesenchymal Stem Cells from Diverse Sources before Culture. World J. Stem Cells 2015, 7, 470–476. [Google Scholar] [CrossRef]
- Da Silva Meirelles, L.; Fontes, A.M.; Covas, D.T.; Caplan, A.I. Mechanisms Involved in the Therapeutic Properties of Mesenchymal Stem Cells. Cytokine Growth Factor Rev. 2009, 20, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Yap, C.; Mieremet, A.; de Vries, C.J.M.; Micha, D.; de Waard, V. Six Shades of Vascular Smooth Muscle Cells Illuminated by KLF4 (Kruppel-Like Factor 4). Arterioscler. Thromb. Vasc. Biol. 2021, 41, 2693–2707. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.R.; Sinha, S.; Owens, G.K. Vascular Smooth Muscle Cells in Atherosclerosis. Circ. Res. 2016, 118, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Cordes, K.R.; Sheehy, N.T.; White, M.P.; Berry, E.C.; Morton, S.U.; Muth, A.N.; Lee, T.-H.; Miano, J.M.; Ivey, K.N.; Srivastava, D. MiR-145 and MiR-143 Regulate Smooth Muscle Cell Fate and Plasticity. Nature 2009, 460, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal Stem Cell Perspective: Cell Biology to Clinical Progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef]
- Friedenstein, A.J.; Chailakhyan, R.K.; Latsinik, N.V.; Panasyuk, A.F.; Keiliss-Borok, I.V. Stromal Cells Responsible for Transferring the Microenvironment of the Hemopoietic Tissues. Cloning in Vitro and Retransplantation in Vivo. Transplantation 1974, 17, 331–340. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human Adipose Tissue Is a Source of Multipotent Stem Cells. MBoC 2002, 13, 4279–4295. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef]
- Sabatini, F.; Petecchia, L.; Tavian, M.; de Villeroché, V.J.; Rossi, G.A.; Brouty-Boyé, D. Human Bronchial Fibroblasts Exhibit a Mesenchymal Stem Cell Phenotype and Multilineage Differentiating Potentialities. Lab. Investig. 2005, 85, 962–971. [Google Scholar] [CrossRef]
- Bari, C.D.; Dell’Accio, F.; Tylzanowski, P.; Luyten, F.P. Multipotent Mesenchymal Stem Cells from Adult Human Synovial Membrane. Arthritis Rheum. 2001, 44, 1928–1942. [Google Scholar] [CrossRef]
- Kordes, C.; Sawitza, I.; Götze, S.; Herebian, D.; Häussinger, D. Stellate Cells Are Mesenchymal Stem Cells. Eur. J. Med. Res. 2014, 19, S6. [Google Scholar] [CrossRef]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal Human Dental Pulp Stem Cells (DPSCs) in Vitro and in Vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. SHED: Stem Cells from Human Exfoliated Deciduous Teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef] [PubMed]
- Seo, B.-M.; Miura, M.; Gronthos, S.; Mark Bartold, P.; Batouli, S.; Brahim, J.; Young, M.; Gehron Robey, P.; Wang, C.Y.; Shi, S. Investigation of Multipotent Postnatal Stem Cells from Human Periodontal Ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef]
- Sonoyama, W.; Liu, Y.; Fang, D.; Yamaza, T.; Seo, B.-M.; Zhang, C.; Liu, H.; Gronthos, S.; Wang, C.-Y.; Shi, S.; et al. Mesenchymal Stem Cell-Mediated Functional Tooth Regeneration in Swine. PLoS ONE 2006, 1, e79. [Google Scholar] [CrossRef]
- Morsczeck, C.; Moehl, C.; Götz, W.; Heredia, A.; Schäffer, T.E.; Eckstein, N.; Sippel, C.; Hoffmann, K.H. In Vitro Differentiation of Human Dental Follicle Cells with Dexamethasone and Insulin. Cell Biol. Int. 2005, 29, 567–575. [Google Scholar] [CrossRef]
- Hutson, E.L.; Boyer, S.; Genever, P.G. Rapid Isolation, Expansion, and Differentiation of Osteoprogenitors from Full-Term Umbilical Cord Blood. Tissue Eng. 2005, 11, 1407–1420. [Google Scholar] [CrossRef]
- Fong, C.; Richards, M.; Manasi, N.; Biswas, A.; Bongso, A. Comparative Growth Behaviour and Characterization of Stem Cells from Human Wharton’s Jelly. Reprod. Biomed. Online 2007, 15, 708–718. [Google Scholar] [CrossRef]
- De Coppi, P.; Bartsch, G.; Siddiqui, M.M.; Xu, T.; Santos, C.C.; Perin, L.; Mostoslavsky, G.; Serre, A.C.; Snyder, E.Y.; Yoo, J.J.; et al. Isolation of Amniotic Stem Cell Lines with Potential for Therapy. Nat. Biotechnol. 2007, 25, 100–106. [Google Scholar] [CrossRef]
- Funk, P.E.; Stephan, R.P.; Witte, P.L. Vascular Cell Adhesion Molecule 1-Positive Reticular Cells Express Interleukin-7 and Stem Cell Factor in the Bone Marrow. Blood 1995, 86, 2661–2671. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; MacArthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and Haematopoietic Stem Cells Form a Unique Bone Marrow Niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Schmitt-Gräff, A.; Skalli, O.; Gabbiani, G. α-Smooth Muscle Actin Is Expressed in a Subset of Bone Marrow Stromal Cells in Normal and Pathological Conditions. Virchows Arch. B Cell Pathol. 1989, 57, 291. [Google Scholar] [CrossRef] [PubMed]
- Risbud, M.; Guttapalli, A.; Tsai, T.-T.; Lee, J.; Danielson, K.; Vaccaro, A.; Albert, T.; Gazit, Z.; Gazit, D.; Shapiro, I. Evidence for Skeletal Progenitor Cells in the Degenerate Human Intervertebral Disc. Spine 2007, 32, 2537–2544. [Google Scholar] [CrossRef] [PubMed]
- Nygaard, G.; Firestein, G.S. Restoring Synovial Homeostasis in Rheumatoid Arthritis by Targeting Fibroblast-like Synoviocytes. Nat. Rev. Rheumatol. 2020, 16, 316–333. [Google Scholar] [CrossRef]
- Appaix, F. Brain Mesenchymal Stem Cells: The Other Stem Cells of the Brain? World J. Stem Cells 2014, 6, 134–143. [Google Scholar] [CrossRef]
- Garcia, F.J.; Sun, N.; Lee, H.; Godlewski, B.; Mathys, H.; Galani, K.; Zhou, B.; Jiang, X.; Ng, A.P.; Mantero, J.; et al. Single-Cell Dissection of the Human Brain Vasculature. Nature 2022, 603, 893–899. [Google Scholar] [CrossRef]
- Geerts, A. History, Heterogeneity, Developmental Biology, and Functions of Quiescent Hepatic Stellate Cells. Semin. Liver Dis. 2001, 21, 311–336. [Google Scholar] [CrossRef]
- Bergers, G.; Song, S. The Role of Pericytes in Blood-Vessel Formation and Maintenance. Neuro-Oncology 2005, 7, 452–464. [Google Scholar] [CrossRef]
- Shaw, I.; Rider, S.; Mullins, J.; Hughes, J.; Peault, B. Pericytes in the Renal Vasculature: Roles in Health and Disease. Nat. Rev. Nephrol. 2018, 14, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Hungerford, J.E.; Little, C.D. Developmental Biology of the Vascular Smooth Muscle Cell: Building a Multilayered Vessel Wall. J. Vasc. Res. 1999, 36, 2–27. [Google Scholar] [CrossRef] [PubMed]
- Farrington-Rock, C.; Crofts, N.J.; Doherty, M.J.; Ashton, B.A.; Griffin-Jones, C.; Canfield, A.E. Chondrogenic and Adipogenic Potential of Microvascular Pericytes. Circulation 2004, 110, 2226–2232. [Google Scholar] [CrossRef]
- Vanlandewijck, M.; He, L.; Mae, M.A.A.; Andrae, J.; Ando, K.; Del Gaudio, F.; Nahar, K.; Lebouvier, T.; Lavina, B.; Gouveia, L.; et al. A Molecular Atlas of Cell Types and Zonation in the Brain Vasculature. Nature 2018, 554, 475–480. [Google Scholar] [CrossRef]
- Rajan, A.M.; Ma, R.C.; Kocha, K.M.; Zhang, D.J.; Huang, P. Dual Function of Perivascular Fibroblasts in Vascular Stabilization in Zebrafish. PLoS Genet. 2020, 16, e1008800. [Google Scholar] [CrossRef] [PubMed]
- Majesky, M.W. Developmental Basis of Vascular Smooth Muscle Diversity. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1248–1258. [Google Scholar] [CrossRef] [PubMed]
- Etchevers, H.C.; Vincent, C.; Le Douarin, M.; Couly, G.F. The Cephalic Neural Crest Provides Pericytes and Smooth Muscle Cells to All Blood Vessels of the Face and Forebrain. Development 2001, 128, 1059–1068. [Google Scholar] [CrossRef]
- Joseph, N.M.; Mukouyama, Y.S.; Mosher, J.T.; Jaegle, M.; Crone, S.A.; Dormand, E.L.; Lee, K.F.; Meijer, D.; Anderson, D.J.; Morrison, S.J. Neural Crest Stem Cells Undergo Multilineage Differentiation in Developing Peripheral Nerves to Generate Endoneurial Fibroblasts in Addition to Schwann Cells. Development 2004, 131, 5599–5612. [Google Scholar] [CrossRef]
- Yamazaki, T.; Nalbandian, A.; Uchida, Y.; Li, W.; Arnold, T.D.; Kubota, Y.; Yamamoto, S.; Ema, M.; Mukouyama, Y. Tissue Myeloid Progenitors Differentiate into Pericytes through TGF-Beta Signaling in Developing Skin Vasculature. Cell Rep. 2017, 18, 2991–3004. [Google Scholar] [CrossRef]
- Yamamoto, S.; Muramatsu, M.; Azuma, E.; Ikutani, M.; Nagai, Y.; Sagara, H.; Koo, B.-N.; Kita, S.; O’Donnell, E.; Osawa, T.; et al. A Subset of Cerebrovascular Pericytes Originates from Mature Macrophages in the Very Early Phase of Vascular Development in CNS. Sci. Rep. 2017, 7, 3855. [Google Scholar] [CrossRef]
- Jiang, X.B.; Rowitch, D.H.; Soriano, P.; McMahon, A.P.; Sucov, H.M. Fate of the Mammalian Cardiac Neural Crest. Development 2000, 127, 1607–1616. [Google Scholar] [CrossRef]
- Zachariah, M.A.; Cyster, J.G. Neural Crest-Derived Pericytes Promote Egress of Mature Thymocytes at the Corticomedullary Junction. Science 2010, 328, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Wilm, B.; Ipenberg, A.; Hastie, N.D.; Burch, J.B.E.; Bader, D.M. The Serosal Mesothelium Is a Major Source of Smooth Muscle Cells of the Gut Vasculature. Development 2005, 132, 5317–5328. [Google Scholar] [CrossRef] [PubMed]
- Que, J.; Wilm, B.; Hasegawa, H.; Wang, F.; Bader, D.; Hogan, B.L.M. Mesothelium Contributes to Vascular Smooth Muscle and Mesenchyme during Lung Development. Proc. Natl. Acad. Sci. USA 2008, 105, 16626–16630. [Google Scholar] [CrossRef]
- Asahina, K.; Zhou, B.; Pu, W.T.; Tsukamoto, H. Septum Transversum-Derived Mesothelium Gives Rise to Hepatic Stellate Cells and Perivascular Mesenchymal Cells in Developing Mouse Liver. Hepatology 2011, 53, 983–995. [Google Scholar] [CrossRef]
- Kruger, G.M.; Mosher, J.T.; Bixby, S.; Joseph, N.; Iwashita, T.; Morrison, S.J. Neural Crest Stem Cells Persist in the Adult Gut but Undergo Changes in Self-Renewal, Neuronal Subtype Potential, and Factor Responsiveness. Neuron 2002, 35, 657–669. [Google Scholar] [CrossRef]
- Kobayashi, A.; Mugford, J.W.; Krautzberger, A.M.; Naiman, N.; Liao, J.; McMahon, A.P. Identification of a Multipotent Self-Renewing Stromal Progenitor Population during Mammalian Kidney Organogenesis. Stem Cell Rep. 2014, 3, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Dettman, R.W.; Denetclaw, W.; Ordahl, C.P.; Bristow, J. Common Epicardial Origin of Coronary Vascular Smooth Muscle, Perivascular Fibroblasts, and Intermyocardial Fibroblasts in the Avian Heart. Dev. Biol. 1998, 193, 169–181. [Google Scholar] [CrossRef]
- Hellstrom, M.; Kalen, M.; Lindahl, P.; Abramsson, A.; Betsholtz, C. Role of PDGF-B and PDGFR-Beta in Recruitment of Vascular Smooth Muscle Cells and Pericytes during Embryonic Blood Vessel Formation in the Mouse. Development 1999, 126, 3047–3055. [Google Scholar] [CrossRef]
- Orange, D.E.; Yao, V.; Sawicka, K.; Fak, J.; Frank, M.O.; Parveen, S.; Blachere, N.E.; Hale, C.; Zhang, F.; Raychaudhuri, S.; et al. RNA Identification of PRIME Cells Predicting Rheumatoid Arthritis Flares. N. Engl. J. Med. 2020, 383, 218–228. [Google Scholar] [CrossRef]
- Sato, S.; Tang, Y.J.; Wei, Q.; Hirata, M.; Weng, A.; Han, I.; Okawa, A.; Takeda, S.; Whetstone, H.; Nadesan, P.; et al. Mesenchymal Tumors Can Derive from Ng2/Cspg4-Expressing Pericytes with Beta-Catenin Modulating the Neoplastic Phenotype. Cell Rep. 2016, 16, 917–927. [Google Scholar] [CrossRef]
- Adameyko, I.; Lallemend, F.; Aquino, J.B.; Pereira, J.A.; Topilko, P.; Müller, T.; Fritz, N.; Beljajeva, A.; Mochii, M.; Liste, I.; et al. Schwann Cell Precursors from Nerve Innervation Are a Cellular Origin of Melanocytes in Skin. Cell 2009, 139, 366–379. [Google Scholar] [CrossRef]
- Di Carlo, S.E.; Peduto, L. The Perivascular Origin of Pathological Fibroblasts. J. Clin. Investig. 2018, 128, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Ehninger, A.; Trumpp, A. The Bone Marrow Stem Cell Niche Grows up: Mesenchymal Stem Cells and Macrophages Move In. J. Exp. Med. 2011, 208, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Ergün, S.; Tilki, D.; Klein, D. Vascular Wall as a Reservoir for Different Types of Stem and Progenitor Cells. Antioxid. Redox Signal. 2010, 15, 981–995. [Google Scholar] [CrossRef]
- Aggarwal, S.; Pittenger, M.F. Human Mesenchymal Stem Cells Modulate Allogeneic Immune Cell Responses. Blood 2005, 105, 1815–1822. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, Y.; Li, Q.; Liu, K.; Hou, J.; Shao, C.; Wang, Y. Immunoregulatory Mechanisms of Mesenchymal Stem and Stromal Cells in Inflammatory Diseases. Nat. Rev. Nephrol. 2018, 14, 493–507. [Google Scholar] [CrossRef]
- Le Blanc, K.; Davies, L.C. Mesenchymal Stromal Cells and the Innate Immune Response. Immunol. Lett. 2015, 168, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Spees, J.L.; Lee, R.H.; Gregory, C.A. Mechanisms of Mesenchymal Stem/Stromal Cell Function. Stem Cell Res. Ther. 2016, 7, 125. [Google Scholar] [CrossRef] [PubMed]
- Uccelli, A.; Moretta, L.; Pistoia, V. Mesenchymal Stem Cells in Health and Disease. Nat. Rev. Immunol. 2008, 8, 726–736. [Google Scholar] [CrossRef]
- Nauta, A.J.; Kruisselbrink, A.B.; Lurvink, E.; Willemze, R.; Fibbe, W.E. Mesenchymal Stem Cells Inhibit Generation and Function of Both CD34+-Derived and Monocyte-Derived Dendritic Cells. J. Immunol. 2006, 177, 2080–2087. [Google Scholar] [CrossRef]
- Raffaghello, L.; Bianchi, G.; Bertolotto, M.; Montecucco, F.; Busca, A.; Dallegri, F.; Ottonello, L.; Pistoia, V. Human Mesenchymal Stem Cells Inhibit Neutrophil Apoptosis: A Model for Neutrophil Preservation in the Bone Marrow Niche. Stem Cells 2008, 26, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Spaggiari, G.M.; Capobianco, A.; Becchetti, S.; Mingari, M.C.; Moretta, L. Mesenchymal Stem Cell-Natural Killer Cell Interactions: Evidence That Activated NK Cells Are Capable of Killing MSCs, Whereas MSCs Can Inhibit IL-2-Induced NK-Cell Proliferation. Blood 2006, 107, 1484–1490. [Google Scholar] [CrossRef] [PubMed]
- Galland, S.; Vuille, J.; Martin, P.; Letovanec, I.; Caignard, A.; Fregni, G.; Stamenkovic, I. Tumor-Derived Mesenchymal Stem Cells Use Distinct Mechanisms to Block the Activity of Natural Killer Cell Subsets. Cell Rep. 2017, 20, 2891–2905. [Google Scholar] [CrossRef] [PubMed]
- Krampera, M.; Glennie, S.; Dyson, J.; Scott, D.; Laylor, R.; Simpson, E.; Dazzi, F. Bone Marrow Mesenchymal Stem Cells Inhibit the Response of Naive and Memory Antigen-Specific T Cells to Their Cognate Peptide. Blood 2003, 101, 3722–3729. [Google Scholar] [CrossRef]
- Sotiropoulou, P.A.; Perez, S.A.; Gritzapis, A.D.; Baxevanis, C.N.; Papamichail, M. Interactions Between Human Mesenchymal Stem Cells and Natural Killer Cells. Stem Cells 2006, 24, 74–85. [Google Scholar] [CrossRef]
- Nemeth, K.; Leelahavanichkul, A.; Yuen, P.S.T.; Mayer, B.; Parmelee, A.; Doi, K.; Robey, P.G.; Leelahavanichkul, K.; Koller, B.H.; Brown, J.M.; et al. Bone Marrow Stromal Cells Attenuate Sepsis via Prostaglandin E-2-Dependent Reprogramming of Host Macrophages to Increase Their Interleukin-10 Production. Nat. Med. 2009, 15, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Tsyb, A.F.; Petrov, V.N.; Konoplyannikov, A.G.; Saypina, E.V.; Lepechina, L.A.; Kalsina, S.S.H.; Semenkova, I.V.; Agaeva, E.V. In Vitro Inhibitory Effect of Mesenchymal Stem Cells on Zymosan-Induced Production of Reactive Oxygen Species. Bull. Exp. Biol. Med. 2008, 146, 158–164. [Google Scholar] [CrossRef]
- Vasandan, A.B.; Jahnavi, S.; Shashank, C.; Prasad, P.; Kumar, A.; Prasanna, S.J. Human Mesenchymal Stem Cells Program Macrophage Plasticity by Altering Their Metabolic Status via a PGE 2 -Dependent Mechanism. Sci. Rep. 2016, 6, 38308. [Google Scholar] [CrossRef]
- Augello, A.; Tasso, R.; Negrini, S.M.; Amateis, A.; Indiveri, F.; Cancedda, R.; Pennesi, G. Bone Marrow Mesenchymal Progenitor Cells Inhibit Lymphocyte Proliferation by Activation of the Programmed Death 1 Pathway. Eur. J. Immunol. 2005, 35, 1482–1490. [Google Scholar] [CrossRef]
- Engela, A.U.; Baan, C.C.; Litjens, N.H.R.; Franquesa, M.; Betjes, M.G.H.; Weimar, W.; Hoogduijn, M.J. Mesenchymal Stem Cells Control Alloreactive CD8+CD28− T Cells. Clin. Exp. Immunol. 2013, 174, 449–458. [Google Scholar] [CrossRef]
- De Mare-Bredemeijer, E.L.D.; Mancham, S.; Verstegen, M.M.A.; de Ruiter, P.E.; van Gent, R.; O’Neill, D.; Tilanus, H.W.; Metselaar, H.J.; de Jonge, J.; Kwekkeboom, J.; et al. Human Graft-Derived Mesenchymal Stromal Cells Potently Suppress Alloreactive T-Cell Responses. Stem Cells Dev. 2015, 24, 1436–1447. [Google Scholar] [CrossRef] [PubMed]
- Corcione, A.; Benvenuto, F.; Ferretti, E.; Giunti, D.; Cappiello, V.; Cazzanti, F.; Risso, M.; Gualandi, F.; Mancardi, G.L.; Pistoia, V.; et al. Human Mesenchymal Stem Cells Modulate B-Cell Functions. Blood 2006, 107, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Gerdoni, E.; Gallo, B.; Casazza, S.; Musio, S.; Bonanni, I.; Pedemonte, E.; Mantegazza, R.; Frassoni, F.; Mancardi, G.; Pedotti, R.; et al. Mesenchymal Stem Cells Effectively Modulate Pathogenic Immune Response in Experimental Autoimmune Encephalomyelitis. Ann. Neurol. 2007, 61, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Ma, T.; Zhou, X.; Jiang, Y.; Han, Y.; Li, H. B Lymphocytes Are the Target of Mesenchymal Stem Cells Immunoregulatory Effect in a Murine Graft-versus-Host Disease Model. Cell Transplant. 2019, 28, 1279–1288. [Google Scholar] [CrossRef]
- Di Ianni, M.; Del Papa, B.; De Ioanni, M.; Moretti, L.; Bonifacio, E.; Cecchini, D.; Sportoletti, P.; Falzetti, F.; Tabilio, A. Mesenchymal Cells Recruit and Regulate T Regulatory Cells. Exp. Hematol. 2008, 36, 309–318. [Google Scholar] [CrossRef]
- Selmani, Z.; Naji, A.; Zidi, I.; Favier, B.; Gaiffe, E.; Obert, L.; Borg, C.; Saas, P.; Tiberghien, P.; Rouas-Freiss, N.; et al. Human Leukocyte Antigen-G5 Secretion by Human Mesenchymal Stem Cells Is Required to Suppress T Lymphocyte and Natural Killer Function and to Induce CD4+CD25highFOXP3+ Regulatory T Cells. Stem Cells 2008, 26, 212–222. [Google Scholar] [CrossRef]
- Zhang, Q.; Fu, L.; Liang, Y.; Guo, Z.; Wang, L.; Ma, C.; Wang, H. Exosomes Originating from MSCs Stimulated with TGF-β and IFN-γ Promote Treg Differentiation. J. Cell. Physiol. 2018, 233, 6832–6840. [Google Scholar] [CrossRef]
- Rashedi, I.; Gómez-Aristizábal, A.; Wang, X.-H.; Viswanathan, S.; Keating, A. TLR3 or TLR4 Activation Enhances Mesenchymal Stromal Cell-Mediated Treg Induction via Notch Signaling. Stem Cells 2017, 35, 265–275. [Google Scholar] [CrossRef]
- Akiyama, K.; Chen, C.; Wang, D.; Xu, X.; Qu, C.; Yamaza, T.; Cai, T.; Chen, W.; Sun, L.; Shi, S. Mesenchymal-Stem-Cell-Induced Immunoregulation Involves FAS-Ligand-/FAS-Mediated T Cell Apoptosis. Cell Stem Cell 2012, 10, 544–555. [Google Scholar] [CrossRef]
- Delorme, B.; Ringe, J.; Gallay, N.; Le Vern, Y.; Kerboeuf, D.; Jorgensen, C.; Rosset, P.; Sensebe, L.; Layrolle, P.; Haeupl, T.; et al. Specific Plasma Membrane Protein Phenotype of Culture-Amplified and Native Human Bone Marrow Mesenchymal Stem Cells. Blood 2008, 111, 2631–2635. [Google Scholar] [CrossRef]
- Giunti, D.; Parodi, B.; Usai, C.; Vergani, L.; Casazza, S.; Bruzzone, S.; Mancardi, G.; Uccelli, A. Mesenchymal Stem Cells Shape Microglia Effector Functions Through the Release of CX3CL1. Stem Cells 2012, 30, 2044–2053. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, F.; Zhou, Y.; Jin, B.; Sun, Q.; Guo, S. Immunosuppressive Property of MSCs Mediated by Cell Surface Receptors. Front. Immunol. 2020, 11, 1076. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Su, G.; Wang, Q.; Wang, R.; Zhang, M. The CD200/CD200R Mechanism in Mesenchymal Stem Cells’ Regulation of Dendritic Cells. Am. J. Transl. Res. 2021, 13, 9607–9613. [Google Scholar] [PubMed]
- Tatara, R.; Ozaki, K.; Kikuchi, Y.; Hatanaka, K.; Oh, I.; Meguro, A.; Matsu, H.; Sato, K.; Ozawa, K. Mesenchymal Stromal Cells Inhibit Th17 but Not Regulatory T-Cell Differentiation. Cytotherapy 2011, 13, 686–694. [Google Scholar] [CrossRef]
- Adamo, A.; Brandi, J.; Caligola, S.; Delfino, P.; Bazzoni, R.; Carusone, R.; Cecconi, D.; Giugno, R.; Manfredi, M.; Robotti, E.; et al. Extracellular Vesicles Mediate Mesenchymal Stromal Cell-Dependent Regulation of B Cell PI3K-AKT Signaling Pathway and Actin Cytoskeleton. Front. Immunol. 2019, 10, 446. [Google Scholar] [CrossRef]
- Collino, F.; Deregibus, M.C.; Bruno, S.; Sterpone, L.; Aghemo, G.; Viltono, L.; Tetta, C.; Camussi, G. Microvesicles Derived from Adult Human Bone Marrow and Tissue Specific Mesenchymal Stem Cells Shuttle Selected Pattern of MiRNAs. PLoS ONE 2010, 5, e11803. [Google Scholar] [CrossRef]
- Iso, Y.; Spees, J.L.; Serrano, C.; Bakondi, B.; Pochampally, R.; Song, Y.-H.; Sobel, B.E.; Delafontaine, P.; Prockop, D.J. Multipotent Human Stromal Cells Improve Cardiac Function after Myocardial Infarction in Mice without Long-Term Engraftment. Biochem. Biophys. Res. Commun. 2007, 354, 700–706. [Google Scholar] [CrossRef]
- Ferguson, S.W.; Wang, J.; Lee, C.J.; Liu, M.; Neelamegham, S.; Canty, J.M.; Nguyen, J. The MicroRNA Regulatory Landscape of MSC-Derived Exosomes: A Systems View. Sci. Rep. 2018, 8, 1419. [Google Scholar] [CrossRef]
- Katsuda, T.; Ochiya, T. Molecular Signatures of Mesenchymal Stem Cell-Derived Extracellular Vesicle-Mediated Tissue Repair. Stem Cell Res. Ther. 2015, 6, 212. [Google Scholar] [CrossRef]
- Nakamura, Y.; Miyaki, S.; Ishitobi, H.; Matsuyama, S.; Nakasa, T.; Kamei, N.; Akimoto, T.; Higashi, Y.; Ochi, M. Mesenchymal-Stem-Cell-Derived Exosomes Accelerate Skeletal Muscle Regeneration. FEBS Lett. 2015, 589, 1257–1265. [Google Scholar] [CrossRef]
- Kishore, R.; Khan, M. More than Tiny Sacks: Stem Cell Exosomes as Cell-Free Modality for Cardiac Repair. Circ. Res. 2016, 118, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Collino, F.; Bruno, S.; Incarnato, D.; Dettori, D.; Neri, F.; Provero, P.; Pomatto, M.; Oliviero, S.; Tetta, C.; Quesenberry, P.J.; et al. AKI Recovery Induced by Mesenchymal Stromal Cell-Derived Extracellular Vesicles Carrying MicroRNAs. J. Am. Soc. Nephrol. 2015, 26, 2349–2360. [Google Scholar] [CrossRef] [PubMed]
- Barile, L.; Lionetti, V.; Cervio, E.; Matteucci, M.; Gherghiceanu, M.; Popescu, L.M.; Torre, T.; Siclari, F.; Moccetti, T.; Vassalli, G. Extracellular Vesicles from Human Cardiac Progenitor Cells Inhibit Cardiomyocyte Apoptosis and Improve Cardiac Function after Myocardial Infarction. Cardiovasc. Res. 2014, 103, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Karnoub, A.E.; Dash, A.B.; Vo, A.P.; Sullivan, A.; Brooks, M.W.; Bell, G.W.; Richardson, A.L.; Polyak, K.; Tubo, R.; Weinberg, R.A. Mesenchymal Stem Cells within Tumour Stroma Promote Breast Cancer Metastasis. Nature 2007, 449, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Quante, M.; Tu, S.P.; Tomita, H.; Gonda, T.; Wang, S.S.W.; Takashi, S.; Baik, G.H.; Shibata, W.; DiPrete, B.; Betz, K.S.; et al. Bone Marrow-Derived Myofibroblasts Contribute to the Mesenchymal Stem Cell Niche and Promote Tumor Growth. Cancer Cell 2011, 19, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Spaeth, E.L.; Dembinski, J.L.; Sasser, A.K.; Watson, K.; Klopp, A.; Hall, B.; Andreeff, M.; Marini, F. Mesenchymal Stem Cell Transition to Tumor-Associated Fibroblasts Contributes to Fibrovascular Network Expansion and Tumor Progression. PLoS ONE 2009, 4, e4992. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B.; Phan, S.H.; Thannickal, V.J.; Galli, A.; Bochaton-Piallat, M.-L.; Gabbiani, G. The Myofibroblast: One Function, Multiple Origins. Am. J. Pathol. 2007, 170, 1807–1816. [Google Scholar] [CrossRef]
- Kalluri, R.; Weinberg, R.A. The Basics of Epithelial-Mesenchymal Transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef]
- Zeisberg, M.; Kalluri, R. Cellular Mechanisms of Tissue Fibrosis. 1. Common and Organ-Specific Mechanisms Associated with Tissue Fibrosis. Am. J. Physiol. Cell Physiol. 2012, 304, C216–C225. [Google Scholar] [CrossRef]
- Göritz, C.; Dias, D.O.; Tomilin, N.; Barbacid, M.; Shupliakov, O.; Frisén, J. A Pericyte Origin of Spinal Cord Scar Tissue. Science 2011, 333, 238–242. [Google Scholar] [CrossRef]
- Humphreys, B.D.; Lin, S.-L.; Kobayashi, A.; Hudson, T.E.; Nowlin, B.T.; Bonventre, J.V.; Valerius, M.T.; McMahon, A.P.; Duffield, J.S. Fate Tracing Reveals the Pericyte and Not Epithelial Origin of Myofibroblasts in Kidney Fibrosis. Am. J. Pathol. 2010, 176, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-L.; Kisseleva, T.; Brenner, D.A.; Duffield, J.S. Pericytes and Perivascular Fibroblasts Are the Primary Source of Collagen-Producing Cells in Obstructive Fibrosis of the Kidney. Am. J. Pathol. 2008, 173, 1617–1627. [Google Scholar] [CrossRef] [PubMed]
- Schrimpf, C.; Duffield, J.S. Mechanisms of Fibrosis: The Role of the Pericyte. Curr. Opin. Nephrol. Hypertens. 2011, 20, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.W.; Eardley, K.S.; Croft, A.P.; Nwosu, J.; Howie, A.J.; Cockwell, P.; Isacke, C.M.; Buckley, C.D.; Savage, C.O.S. CD248+ Stromal Cells Are Associated with Progressive Chronic Kidney Disease. Kidney Int. 2011, 80, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Kramann, R.; Schneider, R.K.; DiRocco, D.P.; Machado, F.; Fleig, S.; Bondzie, P.A.; Henderson, J.M.; Ebert, B.L.; Humphreys, B.D. Perivascular Gli1+ Progenitors Are Key Contributors to Injury-Induced Organ Fibrosis. Cell Stem Cell 2015, 16, 51–66. [Google Scholar] [CrossRef]
- Mederacke, I.; Hsu, C.C.; Troeger, J.S.; Huebener, P.; Mu, X.; Dapito, D.H.; Pradere, J.-P.; Schwabe, R.F. Fate Tracing Reveals Hepatic Stellate Cells as Dominant Contributors to Liver Fibrosis Independent of Its Aetiology. Nat. Commun. 2013, 4, 2823. [Google Scholar] [CrossRef]
- Xie, T.; Liang, J.; Liu, N.; Huan, C.; Zhang, Y.; Liu, W.; Kumar, M.; Xiao, R.; D’Armiento, J.; Metzger, D.; et al. Transcription Factor TBX4 Regulates Myofibroblast Accumulation and Lung Fibrosis. J. Clin. Investig. 2016, 126, 3063–3079. [Google Scholar] [CrossRef]
- Carlson, S.; Trial, J.; Soeller, C.; Entman, M.L. Cardiac Mesenchymal Stem Cells Contribute to Scar Formation after Myocardial Infarction. Cardiovasc. Res. 2011, 91, 99–107. [Google Scholar] [CrossRef]
- Schneider, R.K.; Mullally, A.; Dugourd, A.; Peisker, F.; Hoogenboezem, R.; Van Strien, P.M.H.; Bindels, E.M.; Heckl, D.; Büsche, G.; Fleck, D.; et al. Gli1+ Mesenchymal Stromal Cells Are a Key Driver of Bone Marrow Fibrosis and an Important Cellular Therapeutic Target. Cell Stem Cell 2017, 20, 785–800. [Google Scholar] [CrossRef]
- Lemos, D.R.; Duffield, J.S. Tissue-Resident Mesenchymal Stromal Cells: Implications for Tissue-Specific Antifibrotic Therapies. Sci. Transl. Med. 2018, 10, eaan5174. [Google Scholar] [CrossRef]
- Dulauroy, S.; Di Carlo, S.E.; Langa, F.; Eberl, G.; Peduto, L. Lineage Tracing and Genetic Ablation of ADAM12(+) Perivascular Cells Identify a Major Source of Profibrotic Cells during Acute Tissue Injury. Nat. Med. 2012, 18, 1262–1270. [Google Scholar] [CrossRef]
- Driskell, R.R.; Lichtenberger, B.M.; Hoste, E.; Kretzschmar, K.; Simons, B.D.; Charalambous, M.; Ferron, S.R.; Herault, Y.; Pavlovic, G.; Ferguson-Smith, A.C.; et al. Distinct Fibroblast Lineages Determine Dermal Architecture in Skin Development and Repair. Nature 2013, 504, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Rinkevich, Y.; Walmsley, G.G.; Hu, M.S.; Maan, Z.N.; Newman, A.M.; Drukker, M.; Januszyk, M.; Krampitz, G.W.; Gurtner, G.C.; Lorenz, H.P.; et al. Skin Fibrosis. Identification and Isolation of a Dermal Lineage with Intrinsic Fibrogenic Potential. Science 2015, 348, aaa2151. [Google Scholar] [CrossRef] [PubMed]
- Marangoni, R.G.; Korman, B.D.; Wei, J.; Wood, T.A.; Graham, L.V.; Whitfield, M.L.; Scherer, P.E.; Tourtellotte, W.G.; Varga, J. Myofibroblasts in Murine Cutaneous Fibrosis Originate From Adiponectin-Positive Intradermal Progenitors. Arthritis Rheumatol. 2015, 67, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- Shook, B.A.; Wasko, R.R.; Mano, O.; Rutenberg-Schoenberg, M.; Rudolph, M.C.; Zirak, B.; Rivera-Gonzalez, G.C.; Lopez-Giraldez, F.; Zarini, S.; Rezza, A.; et al. Dermal Adipocyte Lipolysis and Myofibroblast Conversion Are Required for Efficient Skin Repair. Cell Stem Cell 2020, 26, 880–895. [Google Scholar] [CrossRef] [PubMed]
- Joe, A.W.B.; Yi, L.; Natarajan, A.; Le Grand, F.; So, L.; Wang, J.; Rudnicki, M.A.; Rossi, F.M.V. Muscle Injury Activates Resident Fibro/Adipogenic Progenitors That Facilitate Myogenesis. Nat. Cell Biol. 2010, 12, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Uezumi, A.; Fukada, S.; Yamamoto, N.; Takeda, S.; Tsuchida, K. Mesenchymal Progenitors Distinct from Satellite Cells Contribute to Ectopic Fat Cell Formation in Skeletal Muscle. Nat. Cell Biol. 2010, 12, 143–152. [Google Scholar] [CrossRef]
- Leinroth, A.P.; Mirando, A.J.; Rouse, D.; Kobayahsi, Y.; Tata, P.R.; Rueckert, H.E.; Liao, Y.; Long, J.T.; Chakkalakal, J.; Hilton, M.J. Identification of Distinct Non-Myogenic Skeletal-Muscle-Resident Mesenchymal Cell Populations. Cell Rep. 2022, 39, 110785. [Google Scholar] [CrossRef]
- Kanisicak, O.; Khalil, H.; Ivey, M.J.; Karch, J.; Maliken, B.D.; Correll, R.N.; Brody, M.J.; Lin, S.-C.J.; Aronow, B.J.; Tallquist, M.D.; et al. Genetic Lineage Tracing Defines Myofibroblast Origin and Function in the Injured Heart. Nat. Commun. 2016, 7, 12260. [Google Scholar] [CrossRef] [PubMed]
- LeBleu, V.S.; Taduri, G.; O’Connell, J.; Teng, Y.; Cooke, V.G.; Woda, C.; Sugimoto, H.; Kalluri, R. Origin and Function of Myofibroblasts in Kidney Fibrosis. Nat. Med. 2013, 19, 1047–1054. [Google Scholar] [CrossRef]
- Kuppe, C.; Ibrahim, M.M.; Kranz, J.; Zhang, X.; Ziegler, S.; Perales-Paton, J.; Jansen, J.; Reimer, K.C.; Smith, J.R.; Dobie, R.; et al. Decoding Myofibroblast Origins in Human Kidney Fibrosis. Nature 2021, 589, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Iwaisako, K.; Jiang, C.; Zhang, M.; Cong, M.; Moore-Morris, T.J.; Park, T.J.; Liu, X.; Xu, J.; Wang, P.; Paik, Y.-H.; et al. Origin of Myofibroblasts in the Fibrotic Liver in Mice. Proc. Natl. Acad. Sci. USA 2014, 111, E3297–E3305. [Google Scholar] [CrossRef] [PubMed]
- Baron, M.; Veres, A.; Wolock, S.L.; Faust, A.L.; Gaujoux, R.; Vetere, A.; Ryu, J.H.; Wagner, B.K.; Shen-Orr, S.S.; Klein, A.M.; et al. A Single-Cell Transcriptomic Map of the Human and Mouse Pancreas Reveals Inter- and Intra-Cell Population Structure. Cell Syst. 2016, 3, 346–360. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Kim, J.; Yang, S.; Wang, H.; Wu, C.-J.; Sugimoto, H.; LeBleu, V.S.; Kalluri, R. Type I Collagen Deletion in Alpha SMA(+) Myofibroblasts Augments Immune Suppression and Accelerates Progression of Pancreatic Cancer. Cancer Cell 2021, 39, 548–565. [Google Scholar] [CrossRef]
- Rock, J.R.; Barkauskas, C.E.; Cronce, M.J.; Xue, Y.; Harris, J.R.; Liang, J.; Noble, P.W.; Hogan, B.L.M. Multiple Stromal Populations Contribute to Pulmonary Fibrosis without Evidence for Epithelial to Mesenchymal Transition. Proc. Natl. Acad. Sci. USA 2011, 108, E1475–E1483. [Google Scholar] [CrossRef]
- Hung, C.; Linn, G.; Chow, Y.-H.; Kobayashi, A.; Mittelsteadt, K.; Altemeier, W.A.; Gharib, S.A.; Schnapp, L.M.; Duffield, J.S. Role of Lung Pericytes and Resident Fibroblasts in the Pathogenesis of Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med. 2013, 188, 820–830. [Google Scholar] [CrossRef]
- Zepp, J.A.; Zacharias, W.J.; Frank, D.B.; Cavanaugh, C.A.; Zhou, S.; Morley, M.P.; Morrisey, E.E. Distinct Mesenchymal Lineages and Niches Promote Epithelial Self-Renewal and Myofibrogenesis in the Lung. Cell 2017, 170, 1134–1148. [Google Scholar] [CrossRef]
- El Agha, E.; Moiseenko, A.; Kheirollahi, V.; De langhe, S.; Crnkovic, S.; Kwapiszewska, G.; Kosanovic, D.; Schwind, F.; Schermuly, R.T.; Henneke, I.; et al. Two-Way Conversion between Lipogenic and Myogenic Fibroblastic Phenotypes Marks the Progression and Resolution of Lung Fibrosis. Cell Stem Cell 2017, 20, 261–273. [Google Scholar] [CrossRef]
- Tsukui, T.; Sun, K.-H.; Wetter, J.B.; Wilson-Kanamori, J.R.; Hazelwood, L.A.; Henderson, N.C.; Adams, T.S.; Schupp, J.C.; Poli, S.D.; Rosas, I.O.; et al. Collagen-Producing Lung Cell Atlas Identifies Multiple Subsets with Distinct Localization and Relevance to Fibrosis. Nat. Commun. 2020, 11, 1920. [Google Scholar] [CrossRef]
- Dias, D.O.; Kalkitsas, J.; Kelahmetoglu, Y.; Estrada, C.P.; Tatarishvili, J.; Holl, D.; Jansson, L.; Banitalebi, S.; Amiry-Moghaddam, M.; Ernst, A.; et al. Pericyte-Derived Fibrotic Scarring Is Conserved across Diverse Central Nervous System Lesions. Nat. Commun. 2021, 12, 5501. [Google Scholar] [CrossRef]
- Dias, D.O.; Kim, H.; Holl, D.; Solnestam, B.W.; Lundeberg, J.; Carlen, M.; Goritz, C.; Frisen, J. Reducing Pericyte-Derived Scarring Promotes Recovery after Spinal Cord Injury. Cell 2018, 173, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Dorrier, C.E.; Aran, D.; Haenelt, E.A.; Sheehy, R.N.; Hoi, K.K.; Pintaric, L.; Chen, Y.; Lizama, C.O.; Cautivo, K.M.; Weiner, G.A.; et al. CNS Fibroblasts Form a Fibrotic Scar in Response to Immune Cell Infiltration. Nat. Neurosci. 2021, 24, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Decker, M.; Martinez-Morentin, L.; Wang, G.; Lee, Y.; Liu, Q.; Leslie, J.; Ding, L. Leptin-Receptor-Expressing Bone Marrow Stromal Cells Are Myofibroblasts in Primary Myelofibrosis. Nat. Cell Biol. 2017, 19, 677–688. [Google Scholar] [CrossRef]
- Zhou, B.O.; Yue, R.; Murphy, M.M.; Peyer, J.G.; Morrison, S.J. Leptin-Receptor-Expressing Mesenchymal Stromal Cells Represent the Main Source of Bone Formed by Adult Bone Marrow. Cell Stem Cell 2014, 15, 154–168. [Google Scholar] [CrossRef]
- Ding, L.; Saunders, T.L.; Enikolopov, G.; Morrison, S.J. Endothelial and Perivascular Cells Maintain Haematopoietic Stem Cells. Nature 2012, 481, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Croft, A.P.; Naylor, A.J.; Marshall, J.L.; Hardie, D.L.; Zimmermann, B.; Turner, J.; Desanti, G.; Adams, H.; Yemm, A.I.; Müller-Ladner, U.; et al. Rheumatoid Synovial Fibroblasts Differentiate into Distinct Subsets in the Presence of Cytokines and Cartilage. Arthritis Res. Ther. 2016, 18, 270. [Google Scholar] [CrossRef]
- Croft, A.P.; Campos, J.; Jansen, K.; Turner, J.D.; Marshall, J.; Attar, M.; Savary, L.; Wehmeyer, C.; Naylor, A.J.; Kemble, S.; et al. Distinct Fibroblast Subsets Drive Inflammation and Damage in Arthritis. Nature 2019, 570, 246–251. [Google Scholar] [CrossRef]
- Stephenson, W.; Donlin, L.T.; Butler, A.; Rozo, C.; Bracken, B.; Rashidfarrokhi, A.; Goodman, S.M.; Ivashkiv, L.B.; Bykerk, V.P.; Orange, D.E.; et al. Single-Cell RNA-Seq of Rheumatoid Arthritis Synovial Tissue Using Low-Cost Microfluidic Instrumentation. Nat. Commun. 2018, 9, 791. [Google Scholar] [CrossRef]
- Mizoguchi, F.; Slowikowski, K.; Wei, K.; Marshall, J.L.; Rao, D.A.; Chang, S.K.; Nguyen, H.N.; Noss, E.H.; Turner, J.D.; Earp, B.E.; et al. Functionally Distinct Disease-Associated Fibroblast Subsets in Rheumatoid Arthritis. Nat. Commun. 2018, 9, 789. [Google Scholar] [CrossRef]
- Zhang, F.; Wei, K.; Slowikowski, K.; Fonseka, C.Y.; Rao, D.A.; Kelly, S.; Goodman, S.M.; Tabechian, D.; Hughes, L.B.; Salomon-Escoto, K.; et al. Defining Inflammatory Cell States in Rheumatoid Arthritis Joint Synovial Tissues by Integrating Single-Cell Transcriptomics and Mass Cytometry. Nat. Immunol. 2019, 20, 928–942. [Google Scholar] [CrossRef]
- Baker, A.H.; Peault, B. A Gli(1)Ttering Role for Perivascular Stem Cells in Blood Vessel Remodeling. Cell Stem Cell 2016, 19, 563–565. [Google Scholar] [CrossRef]
- Johnson, R.C.; Leopold, J.A.; Loscalzo, J. Vascular Calcification: Pathobiological Mechanisms and Clinical Implications. Circ. Res. 2006, 99, 1044–1059. [Google Scholar] [CrossRef] [PubMed]
- Sartore, S.; Chiavegato, A.; Faggin, E.; Franch, R.; Puato, M.; Ausoni, S.; Pauletto, P. Contribution of Adventitial Fibroblasts to Neointima Formation and Vascular Remodeling: From Innocent Bystander to Active Participant. Circ. Res. 2001, 89, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Kramann, R.; Goettsch, C.; Wongboonsin, J.; Iwata, H.; Schneider, R.K.; Kuppe, C.; Kaesler, N.; Chang-Panesso, M.; Machado, F.G.; Gratwohl, S.; et al. Adventitial MSC-like Cells Are Progenitors of Vascular Smooth Muscle Cells and Drive Vascular Calcification in Chronic Kidney Disease. Cell Stem Cell 2016, 19, 628–642. [Google Scholar] [CrossRef] [PubMed]
- Chandra, P.K.; Gerlach, S.L.; Wu, C.; Khurana, N.; Swientoniewski, L.T.; Abdel-Mageed, A.B.; Li, J.; Braun, S.E.; Mondal, D. Mesenchymal Stem Cells Are Attracted to Latent HIV-1-Infected Cells and Enable Virus Reactivation via a Non-Canonical PI3K-NFκB Signaling Pathway. Sci. Rep. 2018, 8, 14702. [Google Scholar] [CrossRef] [PubMed]
- Cotter, E.J.; Chew, N.; Powderly, W.G.; Doran, P.P. HIV Type 1 Alters Mesenchymal Stem Cell Differentiation Potential and Cell Phenotype Ex Vivo. AIDS Res. Hum. Retrovir. 2010, 27, 187–199. [Google Scholar] [CrossRef]
- Beaupere, C.; Garcia, M.; Larghero, J.; Fève, B.; Capeau, J.; Lagathu, C. The HIV Proteins Tat and Nef Promote Human Bone Marrow Mesenchymal Stem Cell Senescence and Alter Osteoblastic Differentiation. Aging Cell 2015, 14, 534–546. [Google Scholar] [CrossRef] [PubMed]
- Sundin, M.; Örvell, C.; Rasmusson, I.; Sundberg, B.; Ringdén, O.; Le Blanc, K. Mesenchymal Stem Cells Are Susceptible to Human Herpesviruses, but Viral DNA Cannot Be Detected in the Healthy Seropositive Individual. Bone Marrow Transplant. 2006, 37, 1051–1059. [Google Scholar] [CrossRef]
- Smirnov, S.V.; Harbacheuski, R.; Lewis-Antes, A.; Zhu, H.; Rameshwar, P.; Kotenko, S.V. Bone-Marrow-Derived Mesenchymal Stem Cells as a Target for Cytomegalovirus Infection: Implications for Hematopoiesis, Self-Renewal and Differentiation Potential. Virology 2007, 360, 6–16. [Google Scholar] [CrossRef]
- Lee, M.-S.; Yuan, H.; Jeon, H.; Zhu, Y.; Yoo, S.; Shi, S.; Krueger, B.; Renne, R.; Lu, C.; Jung, J.U.; et al. Human Mesenchymal Stem Cells of Diverse Origins Support Persistent Infection with Kaposi’s Sarcoma-Associated Herpesvirus and Manifest Distinct Angiogenic, Invasive, and Transforming Phenotypes. mBio 2016, 7, e02109. [Google Scholar] [CrossRef]
- Pessina, A.; Bonomi, A.; Coccè, V.; Bernardo, M.E.; Cometa, A.M.; Ferrari, M.; Sisto, F.; Cavicchini, L.; Locatelli, F. Assessment of Human Herpesvirus-6 Infection in Mesenchymal Stromal Cells Ex Vivo Expanded for Clinical Use. Transpl. Infect. Dis. Off. J. Transplant. Soc. 2009, 11, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Khatri, M.; Saif, Y.M. Influenza Virus Infects Bone Marrow Mesenchymal Stromal Cells in Vitro: Implications for Bone Marrow Transplantation. Cell Transplant. 2013, 22, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Thanunchai, M.; Kanrai, P.; Wiboon-Ut, S.; Puthavathana, P.; Hongeng, S.; Thitithanyanont, A. Tropism of Avian Influenza A (H5N1) Virus to Mesenchymal Stem Cells and CD34+ Hematopoietic Stem Cells. PLoS ONE 2013, 8, e81805. [Google Scholar] [CrossRef]
- Cheung, M.B.; Sampayo-Escobar, V.; Green, R.; Moore, M.L.; Mohapatra, S.; Mohapatra, S.S. Respiratory Syncytial Virus-Infected Mesenchymal Stem Cells Regulate Immunity via Interferon Beta and Indoleamine-2,3-Dioxygenase. PLoS ONE 2016, 11, e0163709. [Google Scholar] [CrossRef]
- Mumtaz, N.; Koedam, M.; van den Doel, P.B.; van Leeuwen, J.P.T.M.; Koopmans, M.P.G.; van der Eerden, B.C.J.; Rockx, B. Zika Virus Infection Perturbs Osteoblast Function. Sci. Rep. 2018, 8, 16975. [Google Scholar] [CrossRef] [PubMed]
- Rong, Q.; Zhang, L.; Su, E.; Li, J.; Li, J.; Liu, Z.; Huang, Z.; Ma, W.; Cao, K.; Huang, J. Bone Marrow-Derived Mesenchymal Stem Cells Are Capable of Mediating Hepatitis B Virus Infection in Injured Tissues. J. Viral Hepat. 2008, 15, 607–614. [Google Scholar] [CrossRef]
- Cardot-Leccia, N.; Hubiche, T.; Dellamonica, J.; Burel-Vandenbos, F.; Passeron, T. Pericyte Alteration Sheds Light on Micro-Vasculopathy in COVID-19 Infection. Intensive Care Med. 2020, 46, 1777–1778. [Google Scholar] [CrossRef]
- Rendeiro, A.F.; Ravichandran, H.; Bram, Y.; Chandar, V.; Kim, J.; Meydan, C.; Park, J.; Foox, J.; Hether, T.; Warren, S.; et al. The Spatial Landscape of Lung Pathology during COVID-19 Progression. Nature 2021, 593, 564–569. [Google Scholar] [CrossRef]
- Stephenson, S.E.; Wilson, C.L.; Bond, N.G.; Kaur, A.; Alvarez, X.; Midkiff, C.C.; Schnapp, L.M. Pericytes as Novel Targets for HIV/SIV Infection in the Lung. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2020, 319, L848–L853. [Google Scholar] [CrossRef]
- Zhang, L.; Bansal, M.B. Role of Kupffer Cells in Driving Hepatic Inflammation and Fibrosis in HIV Infection. Front. Immunol. 2020, 11, 1086. [Google Scholar] [CrossRef]
- Tuyama, A.C.; Hong, F.; Saiman, Y.; Wang, C.; Ozkok, D.; Mosoian, A.; Chen, P.; Chen, B.K.; Klotman, M.E.; Bansal, M.B. Human Immunodeficiency Virus (HIV)-1 Infects Human Hepatic Stellate Cells and Promotes Collagen I and Monocyte Chemoattractant Protein-1 Expression: Implications for the Pathogenesis of HIV/Hepatitis C Virus-Induced Liver Fibrosis. Hepatology 2010, 52, 612–622. [Google Scholar] [CrossRef]
- Schulze-Krebs, A.; Preimel, D.; Popov, Y.; Bartenschlager, R.; Lohmann, V.; Pinzani, M.; Schuppan, D. Hepatitis C Virus-Replicating Hepatocytes Induce Fibrogenic Activation of Hepatic Stellate Cells. Gastroenterology 2005, 129, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.-C.; Tseng, C.-P.; Liao, M.-H.; Peng, C.-Y.; Yu, J.-S.; Chuang, P.-H.; Huang, J.-T.; Chen, J.J.W. Activation of Hepatic Stellate Cells by the Ubiquitin C-Terminal Hydrolase 1 Protein Secreted from Hepatitis C Virus-Infected Hepatocytes. Sci. Rep. 2017, 7, 4448. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Yuan, H.; Liang, J.; Hong, J.; Qu, C. Expression of Hepatic Stellate Cell Activation-Related Genes in HBV-, HCV-, and Nonalcoholic Fatty Liver Disease-Associated Fibrosis. PLoS ONE 2020, 15, e0233702. [Google Scholar] [CrossRef] [PubMed]
- Salloum, S.; Holmes, J.A.; Jindal, R.; Bale, S.S.; Brisac, C.; Alatrakchi, N.; Lidofsky, A.; Kruger, A.J.; Fusco, D.N.; Luther, J.; et al. Exposure to Human Immunodeficiency Virus/Hepatitis C Virus in Hepatic and Stellate Cell Lines Reveals Cooperative Profibrotic Transcriptional Activation between Viruses and Cell Types: Salloum et Al. Hepatology 2016, 64, 1951–1968. [Google Scholar] [CrossRef]
- Mazzocca, A.; Sciammetta, S.C.; Carloni, V.; Cosmi, L.; Annunziato, F.; Harada, T.; Abrignani, S.; Pinzani, M. Binding of Hepatitis C Virus Envelope Protein E2 to CD81 Up-Regulates Matrix Metalloproteinase-2 in Human Hepatic Stellate Cells. J. Biol. Chem. 2005, 280, 11329–11339. [Google Scholar] [CrossRef]
- Bataller, R.; Paik, Y.; Lindquist, J.N.; Lemasters, J.J.; Brenner, D.A. Hepatitis C Virus Core and Nonstructural Proteins Induce Fibrogenic Effects in Hepatic Stellate Cells. Gastroenterology 2004, 126, 529–540. [Google Scholar] [CrossRef]
- Wang, Q.; Zhou, J.; Zhang, B.; Tian, Z.; Tang, J.; Zheng, Y.; Huang, Z.; Tian, Y.; Jia, Z.; Tang, Y.; et al. Hepatitis B Virus Induces IL-23 Production in Antigen Presenting Cells and Causes Liver Damage via the IL-23/IL-17 Axis. PLoS Pathog. 2013, 9, e1003410. [Google Scholar] [CrossRef]
- Tan, Z.; Qian, X.; Jiang, R.; Liu, Q.; Wang, Y.; Chen, C.; Wang, X.; Ryffel, B.; Sun, B. IL-17A Plays a Critical Role in the Pathogenesis of Liver Fibrosis through Hepatic Stellate Cell Activation. J. Immunol. 2013, 191, 1835–1844. [Google Scholar] [CrossRef]
- Green, D.F.; Resnick, L.; Bourgoignie, J.J. HIV Infects Glomerular Endothelial and Mesangial but Not Epithelial Cells in Vitro. Kidney Int. 1992, 41, 956–960. [Google Scholar] [CrossRef]
- Tokizawa, S.; Shimizu, N.; Hui-Yu, L.; Deyu, F.; Haraguchi, Y.; Oite, T.; Hoshino, H. Infection of Mesangial Cells with HIV and SIV: Identification of GPR1 as a Coreceptor. Kidney Int. 2000, 58, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Alcendor, D.J. Human Vascular Pericytes and Cytomegalovirus Pathobiology. Int. J. Mol. Sci. 2019, 20, 1456. [Google Scholar] [CrossRef] [PubMed]
- Popik, W.; Correa, H.; Khatua, A.; Aronoff, D.M.; Alcendor, D.J. Mesangial Cells, Specialized Renal Pericytes and Cytomegalovirus Infectivity: Implications for HCMV Pathology in the Glomerular Vascular Unit and Post-Transplant Renal Disease. J. Transl. Sci. 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Ustinov, J.A.; Loginov, R.J.; Mattila, P.M.; Nieminen, V.K.; Suni, J.I.; Häyry, P.J.; Lautenschlager, I.T. Cytomegalovirus Infection of Human Kidney Cells in Vitro. Kidney Int. 1991, 40, 954–960. [Google Scholar] [CrossRef]
- Heieren, M.H.; van der Woude, F.J.; Balfour, H.H. Cytomegalovirus Replicates Efficiently in Human Kidney Mesangial Cells. Proc. Natl. Acad. Sci. USA 1988, 85, 1642–1646. [Google Scholar] [CrossRef]
- Sansonno, D.; Gesualdo, L.; Manno, C.; Schena, F.P.; Dammacco, F. Hepatitis C Virus-Related Proteins in Kidney Tissue from Hepatitis C Virus-Infected Patients with Cryoglobulinemic Membranoproliferative Glomerulonephritis. Hepatology 1997, 25, 1237–1244. [Google Scholar] [CrossRef]
- Alcendor, D.J. Zika Virus Infection of the Human Glomerular Cells: Implications for Viral Reservoirs and Renal Pathogenesis. J. Infect. Dis. 2017, 216, 162–171. [Google Scholar] [CrossRef]
- Jansen, J.; Reimer, K.C.; Nagai, J.S.; Varghese, F.S.; Overheul, G.J.; de Beer, M.; Roverts, R.; Daviran, D.; Fermin, L.A.S.; Willemsen, B.; et al. SARS-CoV-2 Infects the Human Kidney and Drives Fibrosis in Kidney Organoids. Cell Stem Cell 2022, 29, 217–231. [Google Scholar] [CrossRef]
- Niu, F.; Yao, H.; Zhang, W.; Sutliff, R.L.; Buch, S. Tat 101-Mediated Enhancement of Brain Pericyte Migration Involves Platelet-Derived Growth Factor Subunit B Homodimer: Implications for Human Immunodeficiency Virus-Associated Neurocognitive Disorders. J. Neurosci. 2014, 34, 11812–11825. [Google Scholar] [CrossRef]
- Nakagawa, S.; Castro, V.; Toborek, M. Infection of Human Pericytes by HIV-1 Disrupts the Integrity of the Blood–Brain Barrier. J. Cell. Mol. Med. 2012, 16, 2950–2957. [Google Scholar] [CrossRef]
- Persidsky, Y.; Hill, J.; Zhang, M.; Dykstra, H.; Winfield, M.; Reichenbach, N.L.; Potula, R.; Mukherjee, A.; Ramirez, S.H.; Rom, S. Dysfunction of Brain Pericytes in Chronic Neuroinflammation. J. Cereb. Blood Flow Metab. 2016, 36, 794–807. [Google Scholar] [CrossRef]
- Cheeran, M.C.-J.; Lokensgard, J.R.; Schleiss, M.R. Neuropathogenesis of Congenital Cytomegalovirus Infection: Disease Mechanisms and Prospects for Intervention. Clin. Microbiol. Rev. 2009, 22, 99–126, Table of Contents. [Google Scholar] [CrossRef] [PubMed]
- Alcendor, D.J.; Charest, A.M.; Zhu, W.Q.; Vigil, H.E.; Knobel, S.M. Infection and Upregulation of Proinflammatory Cytokines in Human Brain Vascular Pericytes by Human Cytomegalovirus. J. Neuroinflamm. 2012, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-J.; Ou, Y.-C.; Li, J.-R.; Chang, C.-Y.; Pan, H.-C.; Lai, C.-Y.; Liao, S.-L.; Raung, S.-L.; Chang, C.-J. Infection of Pericytes In Vitro by Japanese Encephalitis Virus Disrupts the Integrity of the Endothelial Barrier. J. Virol. 2014, 88, 1150–1161. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-Y.; Li, J.-R.; Ou, Y.-C.; Lin, S.-Y.; Wang, Y.-Y.; Chen, W.-Y.; Hu, Y.-H.; Lai, C.-Y.; Chang, C.-J.; Chen, C.-J. Interplay of Inflammatory Gene Expression in Pericytes Following Japanese Encephalitis Virus Infection. Brain Behav. Immun. 2017, 66, 230–243. [Google Scholar] [CrossRef]
- Liu, H.; Qiu, K.; He, Q.; Lei, Q.; Lu, W. Mechanisms of Blood-Brain Barrier Disruption in Herpes Simplex Encephalitis. J. Neuroimmune Pharm. 2019, 14, 157–172. [Google Scholar] [CrossRef]
- Kim, J.; Alejandro, B.; Hetman, M.; Hattab, E.M.; Joiner, J.; Schroten, H.; Ishikawa, H.; Chung, D.-H. Zika Virus Infects Pericytes in the Choroid Plexus and Enters the Central Nervous System through the Blood-Cerebrospinal Fluid Barrier. PLoS Pathog. 2020, 16, e1008204. [Google Scholar] [CrossRef]
- Krauer, F.; Riesen, M.; Reveiz, L.; Oladapo, O.T.; Martínez-Vega, R.; Porgo, T.V.; Haefliger, A.; Broutet, N.J.; Low, N.; Group, W.Z.C.W. Zika Virus Infection as a Cause of Congenital Brain Abnormalities and Guillain–Barré Syndrome: Systematic Review. PLoS Med. 2017, 14, e1002203. [Google Scholar] [CrossRef]
- Butler, J.S.; Dunning, E.C.; Murray, D.W.; Doran, P.P.; O’Byrne, J.M. HIV-1 Protein Induced Modulation of Primary Human Osteoblast Differentiation and Function via a Wnt/β-Catenin-Dependent Mechanism. J. Orthop. Res. 2013, 31, 218–226. [Google Scholar] [CrossRef]
- Chen, W.; Foo, S.-S.; Li, R.W.; Smith, P.N.; Mahalingam, S. Osteoblasts from Osteoarthritis Patients Show Enhanced Susceptibility to Ross River Virus Infection Associated with Delayed Type I Interferon Responses. Virol. J. 2014, 11, 189. [Google Scholar] [CrossRef]
- Chen, W.; Foo, S.-S.; Rulli, N.E.; Taylor, A.; Sheng, K.-C.; Herrero, L.J.; Herring, B.L.; Lidbury, B.A.; Li, R.W.; Walsh, N.C.; et al. Arthritogenic Alphaviral Infection Perturbs Osteoblast Function and Triggers Pathologic Bone Loss. Proc. Natl. Acad. Sci. USA 2014, 111, 6040–6045. [Google Scholar] [CrossRef] [PubMed]
- Noret, M.; Herrero, L.; Rulli, N.; Rolph, M.; Smith, P.N.; Li, R.W.; Roques, P.; Gras, G.; Mahalingam, S. Interleukin 6, RANKL, and Osteoprotegerin Expression by Chikungunya Virus–Infected Human Osteoblasts. J. Infect. Dis. 2012, 206, 455–457. [Google Scholar] [CrossRef] [PubMed]
- Manganelli, P.; Giuliani, N.; Fietta, P.; Mancini, C.; Lazzaretti, M.; Pollini, A.; Quaini, F.; Pedrazzoni, M. OPG/RANKL System Imbalance in a Case of Hepatitis C-Associated Osteosclerosis: The Pathogenetic Key? Clin. Rheumatol. 2005, 24, 296–300. [Google Scholar] [CrossRef]
- Kluger, R.; Mühlberger, H.; Hoffmann, O.; Berger, C.E.; Engel, A.; Pavlova, B.G. Osteoprogenitor Cells and Osteoblasts Are Targets for Hepatitis C Virus. Clin. Orthop. Relat. Res. 2005, 433, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.-B.E.; Omland, L.H.; Krarup, H.; Obel, N. Fracture Risk in Hepatitis C Virus Infected Persons: Results from the DANVIR Cohort Study. J. Hepatol. 2014, 61, 15–21. [Google Scholar] [CrossRef]
- Ayala-Peña, V.; Santillán, G.; Scolaro, L. Experimental in Vitro Infection of Rat Osteoblasts with Measles Virus Stimulates Osteogenic Differentiation. Biochem. Biophys. Res. Commun. 2014, 451, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Potocka-Bakłażec, M.; Sakowicz-Burkiewicz, M.; Kuczkowski, J.; Pawełczyk, T.; Stankiewicz, C.; Sierszeń, W.; Jankowski, Z.; Buczny, J. Expression of TNF-α, OPG, IL-1β and the Presence of the Measles Virus RNA in the Stapes of the Patients with Otosclerosis. Eur. Arch. Oto-Rhino-Laryngol. 2015, 272, 1907–1912. [Google Scholar] [CrossRef]
- Teramachi, J.; Nagata, Y.; Mohammad, K.; Inagaki, Y.; Ohata, Y.; Guise, T.; Michou, L.; Brown, J.P.; Windle, J.J.; Kurihara, N.; et al. Measles Virus Nucleocapsid Protein Increases Osteoblast Differentiation in Paget’s Disease. J. Clin. Investig. 2016, 126, 1012–1022. [Google Scholar] [CrossRef]
- Keswani, S.C.; Polley, M.; Pardo, C.A.; Griffin, J.W.; McArthur, J.C.; Hoke, A. Schwann Cell Chemokine Receptors Mediate HIV-1 Gp120 Toxicity to Sensory Neurons. Ann. Neurol. 2003, 54, 287–296. [Google Scholar] [CrossRef]
- Dilena, R.; Strazzer, S.; Esposito, S.; Paglialonga, F.; Tadini, L.; Barbieri, S.; Giannini, A. Locked-in–like Fulminant Infantile Guillain–Barré Syndrome Associated with Herpes Simplex Virus 1 Infection. Muscle Nerve 2016, 53, 140–143. [Google Scholar] [CrossRef]
- Behar, R.; Wiley, C.; McCutchan, J.A. Cytomegalovirus Polyradiculoneuropathy in Acquired Immune Deficiency Syndrome. Neurology 1987, 37, 557. [Google Scholar] [CrossRef] [PubMed]
- Orlikowski, D.; Porcher, R.; Sivadon-Tardy, V.; Quincampoix, J.-C.; Raphael, J.-C.; Durand, M.-C.; Sharshar, T.; Roussi, J.; Caudie, C.; Annane, D.; et al. Guillain-Barre Syndrome Following Primary Cytomegalovirus Infection: A Prospective Cohort Study. Clin. Infect. Dis. 2011, 52, 837–844. [Google Scholar] [CrossRef]
- Mohan, A.; Smith-Rohrberg, D.; Sethu, M.; Sharma, S.K. Cytomegalovirus Polyradiculopathy: A Rare Neurological Manifestation of Acquired Immunodeficiency Syndrome. Neurol. India 2008, 56, 493–494. [Google Scholar] [CrossRef] [PubMed]
- Sawai, S.; Satoh, M.; Mori, M.; Misawa, S.; Sogawa, K.; Kazami, T.; Ishibashi, M.; Beppu, M.; Shibuya, K.; Ishige, T.; et al. Moesin Is a Possible Target Molecule for Cytomegalovirus-Related Guillain-Barre Syndrome. Neurology 2014, 83, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Miyaji, K.; Devaux, J.; Yuki, N.; Sawai, S.; Mori, M.; Kuwabara, S.; Miyaji, K.; Yuki, N.; Sawai, S.; Mori, M.; et al. Moesin Is a Possible Target Molecule for Cytomegalovirus-Related Guillain-Barre Syndrome. Neurology 2014, 83, 2314–2315. [Google Scholar] [CrossRef] [PubMed]
- Cumberworth, S.L.; Barrie, J.A.; Cunningham, M.E.; de Figueiredo, D.P.G.; Schultz, V.; Wilder-Smith, A.J.; Brennan, B.; Pena, L.J.; Freitas de Oliveira França, R.; Linington, C.; et al. Zika Virus Tropism and Interactions in Myelinating Neural Cell Cultures: CNS Cells and Myelin Are Preferentially Affected. Acta Neuropathol. Commun. 2017, 5, 50. [Google Scholar] [CrossRef]
- Bernardo, M.E.; Fibbe, W.E. Mesenchymal Stromal Cells: Sensors and Switchers of Inflammation. Cell Stem Cell 2013, 13, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Waterman, R.S.; Tomchuck, S.L.; Henkle, S.L.; Betancourt, A.M. A New Mesenchymal Stem Cell (MSC) Paradigm: Polarization into a Pro-Inflammatory MSC1 or an Immunosuppressive MSC2 Phenotype. PLoS ONE 2010, 5, e10088. [Google Scholar] [CrossRef]
- Tomchuck, S.L.; Zwezdaryk, K.J.; Coffelt, S.B.; Waterman, R.S.; Danka, E.S.; Scandurro, A.B. Toll-like Receptors on Human Mesenchymal Stem Cells Drive Their Migration and Immunomodulating Responses. Stem Cells 2008, 26, 99–107. [Google Scholar] [CrossRef]
- Sacchetti, B.; Funari, A.; Michienzi, S.; Di Cesare, S.; Piersanti, S.; Saggio, I.; Tagliafico, E.; Ferrari, S.; Robey, P.G.; Riminucci, M.; et al. Self-Renewing Osteoprogenitors in Bone Marrow Sinusoids Can Organize a Hematopoietic Microenvironment. Cell 2007, 131, 324–336. [Google Scholar] [CrossRef]
- Hmadcha, A.; Martin-Montalvo, A.; Gauthier, B.R.; Soria, B.; Capilla-Gonzalez, V. Therapeutic Potential of Mesenchymal Stem Cells for Cancer Therapy. Front. Bioeng. Biotechnol. 2020, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Rollín, R.; Álvarez-Lafuente, R.; Marco, F.; Jover, J.A.; Hernández-García, C.; Rodríguez-Navas, C.; López-Durán, L.; Fernández-Gutiérrez, B. Human Parvovirus B19, Varicella Zoster Virus, and Human Herpesvirus-6 in Mesenchymal Stem Cells of Patients with Osteoarthritis: Analysis with Quantitative Real-Time Polymerase Chain Reaction. Osteoarthr. Cartil. 2007, 15, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Raicevic, G.; Najar, M.; Busser, H.; Crompot, E.; Bron, D.; Toungouz, M.; Lagneaux, L. Comparison and Immunobiological Characterization of Retinoic Acid Inducible Gene-I-like Receptor Expression in Mesenchymal Stromal Cells. Sci. Rep. 2017, 7, 2896. [Google Scholar] [CrossRef] [PubMed]
- Mastri, M.; Shah, Z.; McLaughlin, T.; Greene, C.J.; Baum, L.; Suzuki, G.; Lee, T. Activation of Toll-like Receptor 3 Amplifies Mesenchymal Stem Cell Trophic Factors and Enhances Therapeutic Potency. Am. J. Physiol.-Cell Physiol. 2012, 303, C1021–C1033. [Google Scholar] [CrossRef] [PubMed]
- Raicevic, G.; Rouas, R.; Najar, M.; Stordeur, P.; Id Boufker, H.; Bron, D.; Martiat, P.; Goldman, M.; Nevessignsky, M.T.; Lagneaux, L. Inflammation Modifies the Pattern and the Function of Toll-like Receptors Expressed by Human Mesenchymal Stromal Cells. Hum. Immunol. 2010, 71, 235–244. [Google Scholar] [CrossRef]
- Meisel, R.; Brockers, S.; Heseler, K.; Degistirici, Ö.; Bülle, H.; Woite, C.; Stuhlsatz, S.; Schwippert, W.; Jäger, M.; Sorg, R.; et al. Human but Not Murine Multipotent Mesenchymal Stromal Cells Exhibit Broad-Spectrum Antimicrobial Effector Function Mediated by Indoleamine 2,3-Dioxygenase. Leukemia 2011, 25, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, V.; Sengupta, S.; Lazo, A.; Woods, P.; Nolan, A.; Bremer, N. Exosomes Derived from Bone Marrow Mesenchymal Stem Cells as Treatment for Severe COVID-19. Stem Cells Dev. 2020, 29, 747–754. [Google Scholar] [CrossRef]
- Sinclair, K.; Yerkovich, S.T.; Chambers, D.C. Mesenchymal Stem Cells and the Lung. Respirology 2013, 18, 397–411. [Google Scholar] [CrossRef]
- Avanzini, M.A.; Mura, M.; Percivalle, E.; Bastaroli, F.; Croce, S.; Valsecchi, C.; Lenta, E.; Nykjaer, G.; Cassaniti, I.; Bagnarino, J.; et al. Human Mesenchymal Stromal Cells Do Not Express ACE2 and TMPRSS2 and Are Not Permissive to SARS-CoV-2 Infection. Stem Cells Transl. Med. 2021, 10, 636–642. [Google Scholar] [CrossRef]
- Uzbas, F.; May, I.D.; Parisi, A.M.; Thompson, S.K.; Kaya, A.; Perkins, A.D.; Memili, E. Molecular Physiognomies and Applications of Adipose-Derived Stem Cells. Stem Cell Rev. Rep. 2015, 11, 298–308. [Google Scholar] [CrossRef]
- Bajek, A.; Gurtowska, N.; Olkowska, J.; Kazmierski, L.; Maj, M.; Drewa, T. Adipose-Derived Stem Cells as a Tool in Cell-Based Therapies. Arch. Immunol. Ther. Exp. 2016, 64, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zhang, Y.; Zhou, J.; Cao, K. Adipose-Derived Mesenchymal Stem Cell Exosomes: A Novel Pathway for Tissues Repair. Cell Tissue Bank 2019, 20, 153–161. [Google Scholar] [CrossRef]
- Kingham, P.J.; Kalbermatten, D.F.; Mahay, D.; Armstrong, S.J.; Wiberg, M.; Terenghi, G. Adipose-Derived Stem Cells Differentiate into a Schwann Cell Phenotype and Promote Neurite Outgrowth in Vitro. Exp. Neurol. 2007, 207, 267–274. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in Inflammation and Metabolic Disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Vernochet, C.; Azoulay, S.; Duval, D.; Guedj, R.; Cottrez, F.; Vidal, H.; Ailhaud, G.; Dani, C. Human Immunodeficiency Virus Protease Inhibitors Accumulate into Cultured Human Adipocytes and Alter Expression of Adipocytokines. J. Biol. Chem. 2005, 280, 2238–2243. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, F.; Zhao, H.; Zhang, X.; Chen, H.; Zhang, K. Human Adipose-Derived Mesenchymal Stem Cells Are Resistant to HBV Infection during Differentiation into Hepatocytes in Vitro. Int. J. Mol. Sci. 2014, 15, 6096–6110. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.E.; Hur, W.; Kim, J.-H.; Li, T.Z.; Lee, E.B.; Lee, S.W.; Kang, W.; Shin, E.-C.; Wakita, T.; Yoon, S.K. MicroRNA-27a Modulates HCV Infection in Differentiated Hepatocyte-Like Cells from Adipose Tissue-Derived Mesenchymal Stem Cells. PLoS ONE 2014, 9, e91958. [Google Scholar] [CrossRef] [PubMed]
- Kupffer, C. Ueber Sternzellen der Leber: Briefliche Mittheilung an Prof. Waldeyer. Arch. Mikrosk. Anat. 1876, 12, 353–358. [Google Scholar] [CrossRef]
- Ito, T.; Nemoto, M. Über Die Kupfferschen Sternzellen Und Die “Fettspeicherungszellen” (“Fat Storing Cells”) in Der Blutkapillarenwand Der Menschlichen Leber. Okajimas Folia Anat. Jpn. 1952, 24, 243–258. [Google Scholar] [CrossRef]
- Miyata, E.; Masuya, M.; Yoshida, S.; Nakamura, S.; Kato, K.; Sugimoto, Y.; Shibasaki, T.; Yamamura, K.; Ohishi, K.; Nishii, K.; et al. Hematopoietic Origin of Hepatic Stellate Cells in the Adult Liver. Blood 2008, 111, 2427–2435. [Google Scholar] [CrossRef]
- Friedman, S.L. Hepatic Stellate Cells: Protean, Multifunctional, and Enigmatic Cells of the Liver. Physiol. Rev. 2008, 88, 125–172. [Google Scholar] [CrossRef] [PubMed]
- Wake, K. Perisinusoidal Stellate Cells (Fat-Storing Cells, Interstitial Cells, Lipocytes), Their Related Structure in and around the Liver Sinusoids, and Vitamin A-Storing Cells in Extrahepatic Organs. In International Review of Cytology; Elsevier: Amsterdam, The Netherlands, 1980; Volume 66, pp. 303–353. ISBN 978-0-12-364466-4. [Google Scholar]
- Zhang, C.-Y.; Yuan, W.-G.; He, P.; Lei, J.-H.; Wang, C.-X. Liver Fibrosis and Hepatic Stellate Cells: Etiology, Pathological Hallmarks and Therapeutic Targets. World J. Gastroenterol. 2016, 22, 10512. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Zhou, L.; Han, Y.-P. Contribution of Hepatic Stellate Cells and Matrix Metalloproteinase 9 in Acute Liver Failure: HSCs and MMP 9 in ALF. Liver Int. 2008, 28, 959–971. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.; Khadem, F.; Uzonna, J.E. Role of Hepatic Stellate Cell (HSC)-Derived Cytokines in Hepatic Inflammation and Immunity. Cytokine 2019, 124, 154542. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Trippler, M.; Pei, R.; Lu, M.; Broering, R.; Gerken, G.; Schlaak, J.F. Toll-like Receptor Activated Human and Murine Hepatic Stellate Cells Are Potent Regulators of Hepatitis C Virus Replication. J. Hepatol. 2009, 51, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Blüm, P.; Pircher, J.; Merkle, M.; Czermak, T.; Ribeiro, A.; Mannell, H.; Krötz, F.; Hennrich, A.; Spannagl, M.; Köppel, S.; et al. Arterial Thrombosis in the Context of HCV-Associated Vascular Disease Can Be Prevented by Protein C. Cell. Mol. Immunol. 2017, 14, 986–996. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kobayashi, N.; Bagheri, N.; Nedrud, J.G.; Strieter, R.M.; Tomino, Y.; Lamm, M.E.; Emancipator, S.N. Differential Effects of Sendai Virus Infection on Mediator Synthesis by Mesangial Cells from Two Mouse Strains. Kidney Int. 2003, 64, 1675–1684. [Google Scholar] [CrossRef]
- Pasch, A.; Frey, F.J. Coxsackie B Viruses and the Kidney—A Neglected Topic. Nephrol. Dial. Transplant. 2006, 21, 1184–1187. [Google Scholar] [CrossRef]
- Popik, W.; Khatua, A.K.; Fabre, N.F.; Hildreth, J.E.K.; Alcendor, D.J. BK Virus Replication in the Glomerular Vascular Unit: Implications for BK Virus Associated Nephropathy. Viruses 2019, 11, 583. [Google Scholar] [CrossRef]
- Zhai, S.; Hu, L.; Zhong, L.; Guo, Y.; Dong, L.; Jia, R.; Wang, Z. Respiratory Syncytial Virus Aggravates Renal Injury through Cytokines and Direct Renal Injury. Front. Cell. Infect. Microbiol. 2016, 6, 112. [Google Scholar] [CrossRef]
- Kotton, C.N.; Fishman, J.A. Viral Infection in the Renal Transplant Recipient. J. Am. Soc. Nephrol. 2005, 16, 1758–1774. [Google Scholar] [CrossRef] [PubMed]
- Baumert, T.F.; Berg, T.; Lim, J.K.; Nelson, D.R. Status of Direct-Acting Antiviral Therapy for Hepatitis C Virus Infection and Remaining Challenges. Gastroenterology 2019, 156, 431–445. [Google Scholar] [CrossRef] [PubMed]
- Flür, K.; Allam, R.; Zecher, D.; Kulkarni, O.P.; Lichtnekert, J.; Schwarz, M.; Beutler, B.; Vielhauer, V.; Anders, H.-J. Viral RNA Induces Type I Interferon-Dependent Cytokine Release and Cell Death in Mesangial Cells via Melanoma-Differentiation-Associated Gene-5: Implications for Viral Infection-Associated Glomerulonephritis. Am. J. Pathol. 2009, 175, 2014–2022. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, T.; Tanaka, H.; Matsumiya, T.; Yoshida, H.; Tanji, K.; Tsuruga, K.; Oki, E.; Aizawa-Yashiro, T.; Ito, E.; Satoh, K. Retinoic Acid-Inducible Gene-I Is Induced by Double-Stranded RNA and Regulates the Expression of CC Chemokine Ligand (CCL) 5 in Human Mesangial Cells. Nephrol. Dial. Transplant. 2010, 25, 3534–3539. [Google Scholar] [CrossRef] [PubMed]
- Allam, R.; Lichtnekert, J.; Moll, A.G.; Taubitz, A.; Vielhauer, V.; Anders, H.-J. Viral RNA and DNA Trigger Common Antiviral Responses in Mesangial Cells. J. Am. Soc. Nephrol. 2009, 20, 1986–1996. [Google Scholar] [CrossRef]
- Patole, P.S.; Gröne, H.-J.; Segerer, S.; Ciubar, R.; Belemezova, E.; Henger, A.; Kretzler, M.; Schlöndorff, D.; Anders, H.-J. Viral Double-Stranded RNA Aggravates Lupus Nephritis through Toll-Like Receptor 3 on Glomerular Mesangial Cells and Antigen-Presenting Cells. J. Am. Soc. Nephrol. 2005, 16, 1326–1338. [Google Scholar] [CrossRef]
- Nyúl-Tóth, Á.; Kozma, M.; Nagyőszi, P.; Nagy, K.; Fazakas, C.; Haskó, J.; Molnár, K.; Farkas, A.E.; Végh, A.G.; Váró, G.; et al. Expression of Pattern Recognition Receptors and Activation of the Non-Canonical Inflammasome Pathway in Brain Pericytes. Brain Behav. Immun. 2017, 64, 220–231. [Google Scholar] [CrossRef]
- Kovac, A.; Erickson, M.A.; Banks, W.A. Brain Microvascular Pericytes Are Immunoactive in Culture: Cytokine, Chemokine, Nitric Oxide, and LRP-1 Expression in Response to Lipopolysaccharide. J. Neuroinflamm. 2011, 8, 139. [Google Scholar] [CrossRef]
- Duan, L.; Zhang, X.-D.; Miao, W.-Y.; Sun, Y.-J.; Xiong, G.; Wu, Q.; Li, G.; Yang, P.; Yu, H.; Li, H.; et al. PDGFRβ Cells Rapidly Relay Inflammatory Signal from the Circulatory System to Neurons via Chemokine CCL2. Neuron 2018, 100, 183–200.e8. [Google Scholar] [CrossRef]
- Cho, H.J.; Kuo, A.M.-S.; Bertrand, L.; Toborek, M. HIV Alters Gap Junction-Mediated Intercellular Communication in Human Brain Pericytes. Front. Mol. Neurosci. 2017, 10, 410. [Google Scholar] [CrossRef]
- Bertrand, L.; Cho, H.J.; Toborek, M. Blood–Brain Barrier Pericytes as a Target for HIV-1 Infection. Brain 2019, 142, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Piekna-Przybylska, D.; Nagumotu, K.; Reid, D.M.; Maggirwar, S.B. HIV-1 Infection Renders Brain Vascular Pericytes Susceptible to the Extracellular Glutamate. J. Neurovirol. 2019, 25, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Gaceb, A.; Özen, I.; Padel, T.; Barbariga, M.; Paul, G. Pericytes Secrete Pro-Regenerative Molecules in Response to Platelet-Derived Growth Factor-BB. J. Cereb. Blood Flow Metab. 2018, 38, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Blecharz-Lang, K.G.; Wagner, J.; Fries, A.; Nieminen-Kelhä, M.; Rösner, J.; Schneider, U.C.; Vajkoczy, P. Interleukin 6-Mediated Endothelial Barrier Disturbances Can Be Attenuated by Blockade of the IL6 Receptor Expressed in Brain Microvascular Endothelial Cells. Transl. Stroke Res. 2018, 9, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Saint-Pastou Terrier, C.; Gasque, P. Bone Responses in Health and Infectious Diseases: A Focus on Osteoblasts. J. Infect. 2017, 75, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Foo, S.-S.; Taylor, A.; Lulla, A.; Merits, A.; Hueston, L.; Forwood, M.R.; Walsh, N.C.; Sims, N.A.; Herrero, L.J.; et al. Bindarit, an Inhibitor of Monocyte Chemotactic Protein Synthesis, Protects against Bone Loss Induced by Chikungunya Virus Infection. J. Virol. 2015, 89, 581–593. [Google Scholar] [CrossRef]
- Chen, C.-H.; Lin, C.-L.; Kao, C.-H. Relation Between Hepatitis C Virus Exposure and Risk of Osteoporosis: A Nationwide Population-Based Study. Medicine 2015, 94, e2086. [Google Scholar] [CrossRef]
- Gibellini, D.; Crignis, E.D.; Ponti, C.; Cimatti, L.; Borderi, M.; Tschon, M.; Giardino, R.; Re, M.C. HIV-1 Triggers Apoptosis in Primary Osteoblasts and HOBIT Cells through TNFα Activation. J. Med. Virol. 2008, 80, 1507–1514. [Google Scholar] [CrossRef]
- Bar-Shavit, Z. Taking a Toll on the Bones: Regulation of Bone Metabolism by Innate Immune Regulators. Autoimmunity 2008, 41, 195–203. [Google Scholar] [CrossRef]
- Nakamura, K.; Deyama, Y.; Yoshimura, Y.; Suzuki, K.; Morita, M. Toll-like Receptor 3 Ligand-Induced Antiviral Response in Mouse Osteoblastic Cells. Int. J. Mol. Med. 2007, 19, 771–775. [Google Scholar] [CrossRef]
- Nakamura, K.; Deyama, Y.; Yoshimura, Y.; Suzuki, K.; Morita, M. Synthetic Double-Stranded RNA Induces Retinoic Acid-Inducible Gene-I in Mouse Osteoblastic Cells. Mol. Med. Rep. 2008, 1, 833–836. [Google Scholar] [CrossRef] [PubMed]
- Larousserie, F.; Bsiri, L.; Dumaine, V.; Dietrich, C.; Audebourg, A.; Radenen-Bussière, B.; Anract, P.; Vacher-Lavenu, M.-C.; Devergne, O. Frontline Science: Human Bone Cells as a Source of IL-27 under Inflammatory Conditions: Role of TLRs and Cytokines. J. Leukoc. Biol. 2017, 101, 1289–1300. [Google Scholar] [CrossRef] [PubMed]
- Matsuse, D.; Kitada, M.; Kohama, M.; Nishikawa, K.; Makinoshima, H.; Wakao, S.; Fujiyoshi, Y.; Heike, T.; Nakahata, T.; Akutsu, H.; et al. Human Umbilical Cord-Derived Mesenchymal Stromal Cells Differentiate Into Functional Schwann Cells That Sustain Peripheral Nerve Regeneration. J. Neuropathol. Exp. Neurol. 2010, 69, 973–985. [Google Scholar] [CrossRef]
- Sun, X.; Zhu, Y.; Yin, H.; Guo, Z.; Xu, F.; Xiao, B.; Jiang, W.; Guo, W.; Meng, H.; Lu, S.; et al. Differentiation of Adipose-Derived Stem Cells into Schwann Cell-like Cells through Intermittent Induction: Potential Advantage of Cellular Transient Memory Function. Stem Cell Res. Ther. 2018, 9, 133. [Google Scholar] [CrossRef]
- Assouline, J.G.; Levin, M.J.; Straus, E.; Ostrovet, J.M. Varicella-Zoster Virus Infection of Human Astrocytes, Schwann Ceils, and Neurons. Virology 1990, 179, 834–844. [Google Scholar] [CrossRef]
- Shimeld, C.; Efstathiou, S.; Hill, T. Tracking the Spread of a LacZ-Tagged Herpes Simplex Virus Type 1 between the Eye and the Nervous System of the Mouse: Comparison of Primary and Recurrent Infection. J. Virol. 2001, 75, 5252–5262. [Google Scholar] [CrossRef]
- Volpi, V.G.; Pagani, I.; Ghezzi, S.; Iannacone, M.; D’Antonio, M.; Vicenzi, E. Zika Virus Replication in Dorsal Root Ganglia Explants from Interferon Receptor1 Knockout Mice Causes Myelin Degeneration. Sci. Rep. 2018, 8, 10166. [Google Scholar] [CrossRef]
- Dhiman, G.; Abraham, R.; Griffin, D.E. Human Schwann Cells Are Susceptible to Infection with Zika and Yellow Fever Viruses, but Not Dengue Virus. Sci. Rep. 2019, 9, 9951. [Google Scholar] [CrossRef]
- Muñoz, L.; Barreras, P.; Pardo, C. Zika Virus–Associated Neurological Disease in the Adult: Guillain–Barré Syndrome, Encephalitis, and Myelitis. Semin. Reprod. Med. 2016, 34, 273–279. [Google Scholar] [CrossRef]
- Lebeau, G.; Frumence, E.; Turpin, J.; Begue, F.; Hoarau, J.-J.; Gadea, G.; Krejbich-Trotot, P.; Desprès, P.; Viranaicken, W. Zika E Glycan Loop Region and Guillain–Barré Syndrome-Related Proteins: A Possible Molecular Mimicry to Be Taken in Account for Vaccine Development. Vaccines 2021, 9, 283. [Google Scholar] [CrossRef]
- Goethals, S.; Ydens, E.; Timmerman, V.; Janssens, S. Toll-like Receptor Expression in the Peripheral Nerve. Glia 2010, 58, 1701–1709. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.H.; Nardi, N.B. From Leading Role to the Backstage: Mesenchymal Stem Cells as Packaging Cell Lines for in Situ Production of Viral Vectors. Med. Hypotheses 2006, 67, 922–925. [Google Scholar] [CrossRef] [PubMed]




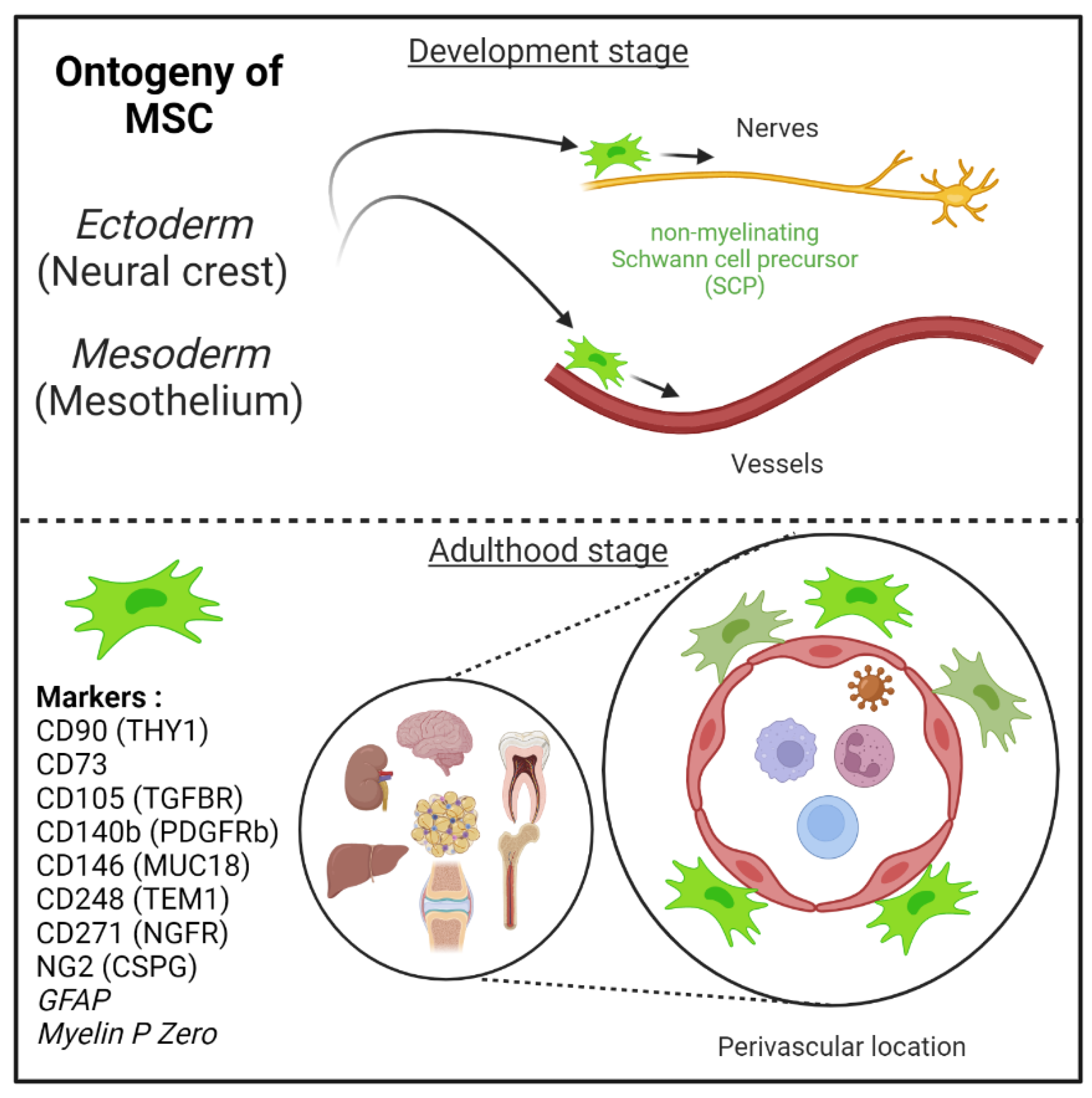{kind=link}
{kind=link}
{kind=link}
| Tissue | Human | Reference |
|---|---|---|
| Bone Marrow | Bone Marrow-Mesenchymal Stem Cell or Adventitial Reticular Cell or Myoid Cell | [17,63,64,65] |
| Adipose tissue | Adipose Stem Cell | [50,51] |
| Intervertebral disc | Skeletal Progenitor Cell | [66] |
| Synovial membrane | Synovial Membrane-derived MSCs Fibroblast-like synoviocytes (FLS) | [53,67] |
| Amniotic fluid | Amniotic Stem Cell | [62] |
| Cord blood Placenta | Umbilical Cord Blood Stem Cell or Wharton’s Jelly derived MSC | [60,61] |
| Dental tissue | Dental Pulp Stem Cell or Stem cells from Human Exfoliated Deciduous teeth or Periodontal Ligament MSC or Stem Cell from Apical Papilla or Dental Follicle Precursor Cell | [55,56,57,58,59] |
| Kidney | Mesangial Cell | [4] |
| Brain | Pericytes, perivascular fibroblasts | [68,69] |
| Liver | Hepatic Stellate Cell (HSC) or Perisinusoidal Cell or Ito Cell | [54,70] |
| Lung | Human Bronchial Fibroblasts or Lung-resident MSC | [52] |
| Tissue | Cell Markers | Lineage Tracing Marker | Location in Tissue | Model of Injury | Pathological Role | Reference |
|---|---|---|---|---|---|---|
| Skin | ADAM12+ PDGFRα+ CD140a + Sca1+ | Adam12 | Perivascular | Injury (Adjuvant, CFA) | Myofib. | [153] |
| PDGFRα+ Dlk1+ Sca1/Ly6a+ | PDGFRα, Dlk1 | Low dermis | Wound | Myofib/Adipo. | [154] | |
| CD26/DPP4+ | Engrailed-1 | Tissue-resident | Wound | Myofib. | [155] | |
| Adiponectin+ | Adiponectin | Intradermal adipocyte | Bleomycin-induced skin fibrosis | Myofib. | [156,157] | |
| Gli1+, PDGFRβ+, PDGFRα+ Nestin+ | Gli1 | Perivascular | - | Myofib. | [147] | |
| Skeletal muscle | PDGFRα+, Sca1+, CD34+ | - | Perivascular | Injury (notexin, Glyc) | Myofib/adipo. | [158,159] |
| ADAM12+, PDGFRα+, Sca1+ | Adam12 | Perivascular | Injury (CX) | Myofib. | [153] | |
| Gli1+, PDGFRβ+, PDGFRα+ | Gli1 | Perivascular | I/R injury | Myofib. | [147] | |
| PDGFRα+, Sca1+ Clusterin+ Osr1+ progenitors | Prrx1 | - | Barium Chl., Glycerol, Sciatic nerve injuries | Adipo/Osteob. | [160] | |
| Heart | Gli1+, PDGFRβ+, PDGFRα+ | Gli1 | Perivascular | Angiotensin 2-fibrosis | Myofib. | [147] |
| Tcf21 + Vimentin+, PDGFRα+, α-SMA+ | Periostin | Tissue-resident | I/R myocardial fibrosis | Myofib. | [161] | |
| Kidney | PDGFRβ+, CD73+ | FoxD1 | Perivascular, peritubular | UUO, I/R injury | Myofib. | [143] |
| PDGFRβ+, CD73+ CD271(P75) + EPO+ | Myelin P Zero | Perivascular | UUO/Folic acid injury | Myofib. | [12] | |
| Alpha SMA | Interstitial Fib | UUO | Myofib. | [162] | ||
| Gli1+, PDGFRβ+, PDGFRα+ | Gli1 | Perivascular | UUO, I/R injury | Myofib. | [147] | |
| POSTN+ PDGFRβ+ | PDGFRβ | Mo/Human | Myofib | [163] | ||
| Liver | PDGFRβ+, desmin+ | LRAt Col1α1 | Perivascular | CCl4-, TAA- injuries | Myofib. | [148] |
| Vit A+ GFAP+, CD90- Desmin+ Calcitonin a | Collα1 | Perivascular HSC and portal Fib | CCl4-, bile duct ligation injuries | Myofib. | [164] | |
| Gli1+, PDGFRβ+, PDGFRα+ | Gli1 | Perivascular | CCl4 | Myofib. | [147] | |
| Pancreas | 2 subsets: PDGFRβ+ PLP1, and PMP22 (Schwann) | - | Stellate cells | Carcinoma | Collagen+ Stellate cells Neural crest Schwann-like, Cancer | [165,166] |
| Lung | PDGFRβ+, NG2+ | NG2 | Perivascular | Bleomycin injury | Myofib. | [167] |
| PDGFRβ+, NG2+ | FoxD1 | Perivascular tissue-resident | Bleomycin injury | Myofib. | [168] | |
| Gli1+, PDGFRβ+, PDGFRα+ | Gli1 | Perivascular | Bleomycin injury | Myofib. | [147] | |
| Axin2+, PDGFRα– | Axin2 PDGFRα Wnt2 | Perivascular Peribronchial | Bleomycin, naphthalene injuries | Myofib. | [169] | |
| Acta2- | Acta2 Adrp | Perivascular | Bleomycin injury | Myofib. and reversion to lipofib. | [170] | |
| Collagen+ | Collα1 | Perivascular and interstitial | Bleomycin injury | Fib. Clusters (11) of collagen-producing subpopulations including highly pathogenic cluster 8: cthrc1+ | [171] | |
| Spinal cord | PDGFR a/β+, CD13+ desmin– | Glast | Type-A pericytes | Spinal cord injury | Myofib Scar tissue | [142] |
| Brain cortex | PDGFR β+ | Glast | Type-A pericytes | Cortico-striatal stab wound, EAE autoimmunity Mid. cer. Art.occlusion (MCAO) | Myofib Scar tissue | [172,173] |
| - | Collα1 | Type-A pericytes | EAE autoimmunity | Myofib. and Immune stroma | [174] | |
| - | perivascular | Healthy CNS | Three cluster of perivascular fibroblasts including type I fibulin+ (ECMhigh) | [69] | ||
| Bone Marrow | CXCL12+ | Nestin | Pericyte-like | - | Stromal cells of HSC | [64] |
| PDGFR a+ CXCL12/SDF1a+ | SCF LepR | Pericyte-like | - | Myofibroblast, osteocyte, chondrocyte, adipocyte | [175,176,177] | |
| Gli1+, PDGFRβ+, PDGFRα+ Nestin+ | Gli1 | Perivascular | - | Myofib. | [147] | |
| CD105+ (TGFbR) Nestin+ CD73+ CD44+, CD29+, Sca1+, and CD51+ | Gli1 | Perivascular endosteal niche | Thrombo- poietin-induced fibrosis | Myofibroblast | [151] | |
| Synovial tissue | CD90/THY1+ FAP+ CD248+ | - | Sublining Fib. | Rheumatoid Arthritis (RA) | Immune effector profile: high expression of cytokines and chemokines | [163,178,179,180,181,182] |
| CD55/DAF+ CD90- FAP+ PPDN+ | - | Lining Fib. | RA | Bone effector profile: mediate bone and cartilage damage | [163,178,179,180,181,182] |
| Cells | Viruses | Related Outcomes |
|---|---|---|
| Bone marrow-MSC | HIV | Inability to support Hematopoietic Stem Cells expansion and implication in HIV-related cytopenia [27]; HIV-related reactivation [187,188,189] |
| HCMV | Transmit to neighboring cells after reactivation [190,191] | |
| Modifies the physiological interaction between BM-MSCs and HSC | ||
| Impairment of osteoblast regeneration, cartilage regeneration, hematopoiesis and properties of immune progenitor cells [191] | ||
| HHV | Lower proliferation rates and altered phenotypes related to malignant transformation [190,192,193] | |
| Influenza A H5N1 | Risk of transmission during bone marrow transplantation [194,195] | |
| RSV | Alteration of immunoregulatory functions [196] | |
| ZIKV | Impaired osteoblast differentiation and possible implication in development of bone pathologies [197] | |
| HBV | HBV-associated myocarditis and other HBV-related extrahepatic diseases [198] | |
| Lung resident MSC/pericytes | SARS-CoV-2 | Pericyte apoptosis and loss in COVID-19 patients [199,200] |
| SIV/HIV | Development of HIV-related pulmonary complications [197,201] | |
| Hepatic Stellate Cell (HepSC, Ito cells) | HIV | HepSC activation and chemotaxis through HIV gp120 envelope protein [202,203] |
| HCV | HCV proteins as well as RNA released by hepatocytes are activating HepSC [204,205]. Constant activation leading to liver fibrosis [206,207,208,209] | |
| HBV | Release of IL-17 by infected cells which stimulate liver fibrosis by activation of HepSC [210,211]. | |
| Mesangial Cell (Kidney) | HIV | HIV-associated glomerulosclerosis due to increased proliferation and matrix synthesis [212,213] |
| HCMV | Glomerulosclerosis [214,215,216,217] | |
| HCV | Glomerulonephritis [218] | |
| ZIKV | Viral reservoir (persistent viruria) [219] | |
| SARS-CoV-2 | Stromal (MSC-like cells) are infected and may contribute to kidney fibrosis in a model of spheroid cultures and to be correlated with kidney fibrosis in COVID19 patients [220]. | |
| Brain Pericytes (BP) | HIV | Decreased BPs coverage of blood brain barrier (BBB) associated with higher permeability [221,222] |
| IL-6 and PDGF-BB secretions concur in HIV-induced CNS damage and BBB disruption [221,223] | ||
| HCMV | Contribute to HCMV dissemination [224] | |
| CXCL8, CXCL11, CCL5, TNF-α, IL-1β and IL-6 secretion causing neuroinflammation [225] | ||
| JEV | IL-6 secretion leads to ZO-1 degradation and BBB impairment [226] | |
| PGE2 and RANTES secretion by BPs recruit leukocytes to the site of infection. Associated with BBB impairment, this provoke leukocyte infiltration and major neuroinflammation [227] | ||
| HSV | BBB impairment associated with leukocytes recruitment leading to major neuroinflammation [228] | |
| ZIKV | Brain abnormalities and BBB defect [229,230] | |
| Osteoblasts | HIV (gp120) | TNF-α and impaired Wnt/β-Catenin signaling promote bone demineralization and reduced bone mass leading to osteopenia and osteoprosis [231] |
| RRV | Imbalance in RANKL/OPG ratio in favor of osteoclastogenic activities and bone loss [232,233] | |
| CHIKV | Proinflammatory (IL-6) and pro-osteoclastic (RANKL) effects in infected cells [234] | |
| HCV | Associated with bone density hardening and osteosclerosis [235] | |
| Increased risk of osteoporosis [236] | ||
| Increased risk of fracture [237] | ||
| Impairment of RANKL/OPG ratio [235] | ||
| ZIKV | Impaired osteoblasts function triggering an imbalance in bone homeostasis and inducing bone-related disorders [197] | |
| MeV | Higher expression of several osteogenic markers and osteogenic differentiation [238] | |
| Otosclerosis [239] | ||
| Paget’s disease [240] | ||
| Schwann Cell (SC) | HIV | Dorsal root ganglion neurotoxicity, including axon and myelin injury [241] |
| HSV/VZV | The principal mechanism evoked for HSV-induced GBS is a molecular mimicry of viral proteins [242] | |
| HCMV | Probable molecular mimicry generating autoantibodies against moesin expressed by SCs [243,244,245,246,247] | |
| ZIKV | Possible direct viral pathogenic effect or a cell-mediated inflammation in pathogenesis of ZIKV-associated GBS [248] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebeau, G.; Ah-Pine, F.; Daniel, M.; Bedoui, Y.; Vagner, D.; Frumence, E.; Gasque, P. Perivascular Mesenchymal Stem/Stromal Cells, an Immune Privileged Niche for Viruses? Int. J. Mol. Sci. 2022, 23, 8038. https://doi.org/10.3390/ijms23148038
Lebeau G, Ah-Pine F, Daniel M, Bedoui Y, Vagner D, Frumence E, Gasque P. Perivascular Mesenchymal Stem/Stromal Cells, an Immune Privileged Niche for Viruses? International Journal of Molecular Sciences. 2022; 23(14):8038. https://doi.org/10.3390/ijms23148038
Chicago/Turabian StyleLebeau, Grégorie, Franck Ah-Pine, Matthieu Daniel, Yosra Bedoui, Damien Vagner, Etienne Frumence, and Philippe Gasque. 2022. "Perivascular Mesenchymal Stem/Stromal Cells, an Immune Privileged Niche for Viruses?" International Journal of Molecular Sciences 23, no. 14: 8038. https://doi.org/10.3390/ijms23148038
APA StyleLebeau, G., Ah-Pine, F., Daniel, M., Bedoui, Y., Vagner, D., Frumence, E., & Gasque, P. (2022). Perivascular Mesenchymal Stem/Stromal Cells, an Immune Privileged Niche for Viruses? International Journal of Molecular Sciences, 23(14), 8038. https://doi.org/10.3390/ijms23148038