
Stem Cell Therapy Enhances Motor Activity of Triceps Surae Muscle in Mice with Hereditary Peripheral Neuropathy

 , , ,
, , ,  ,
, 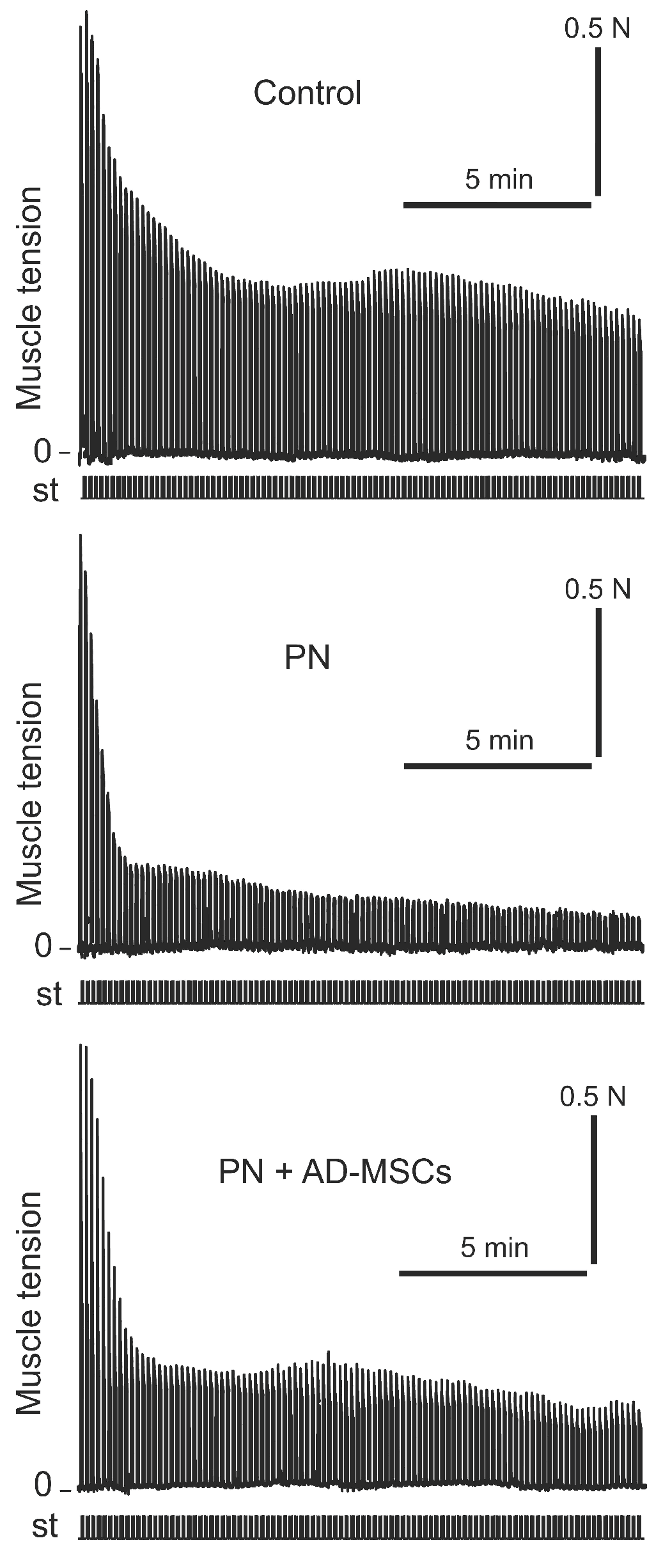{kind=link}
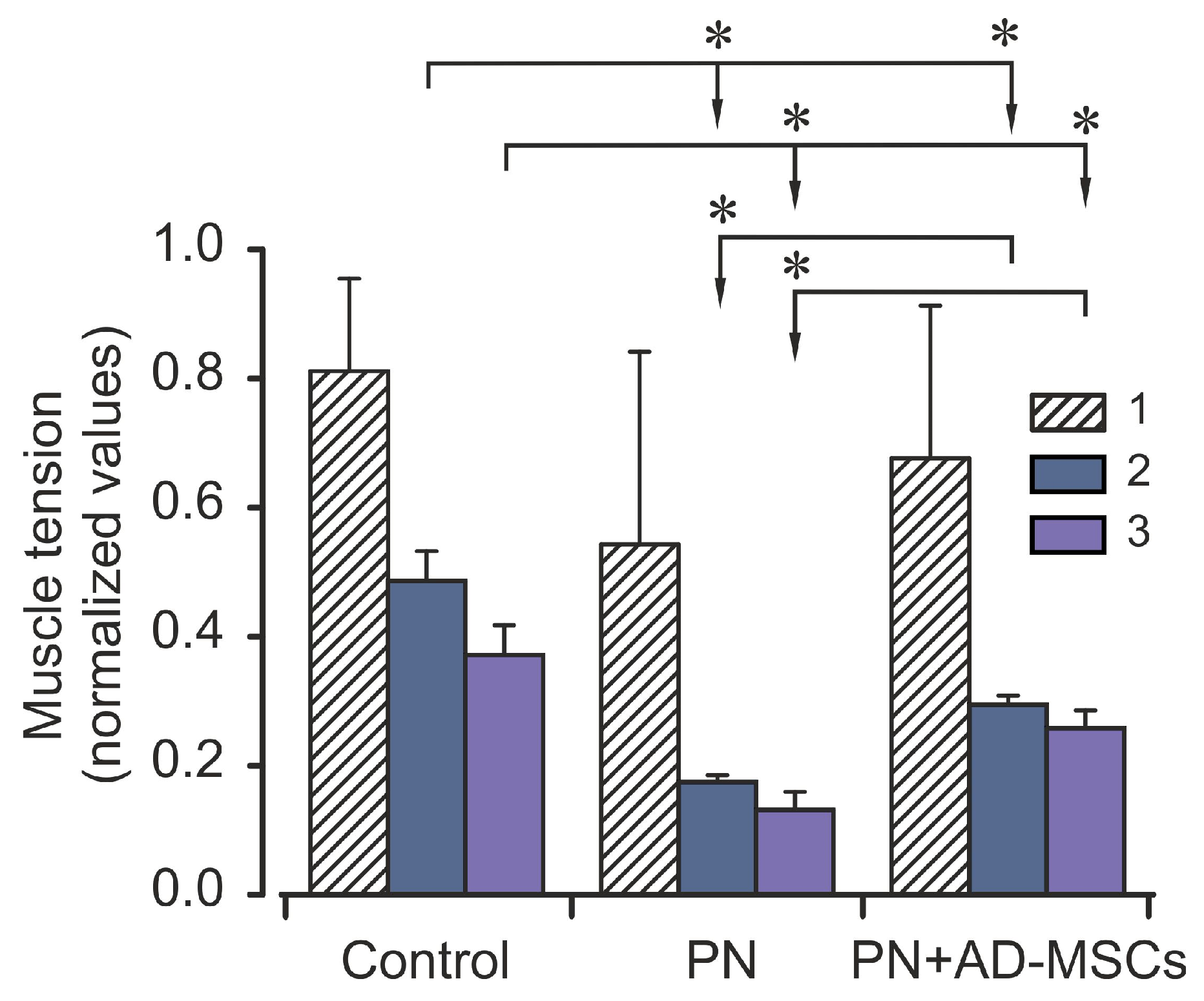{kind=link}
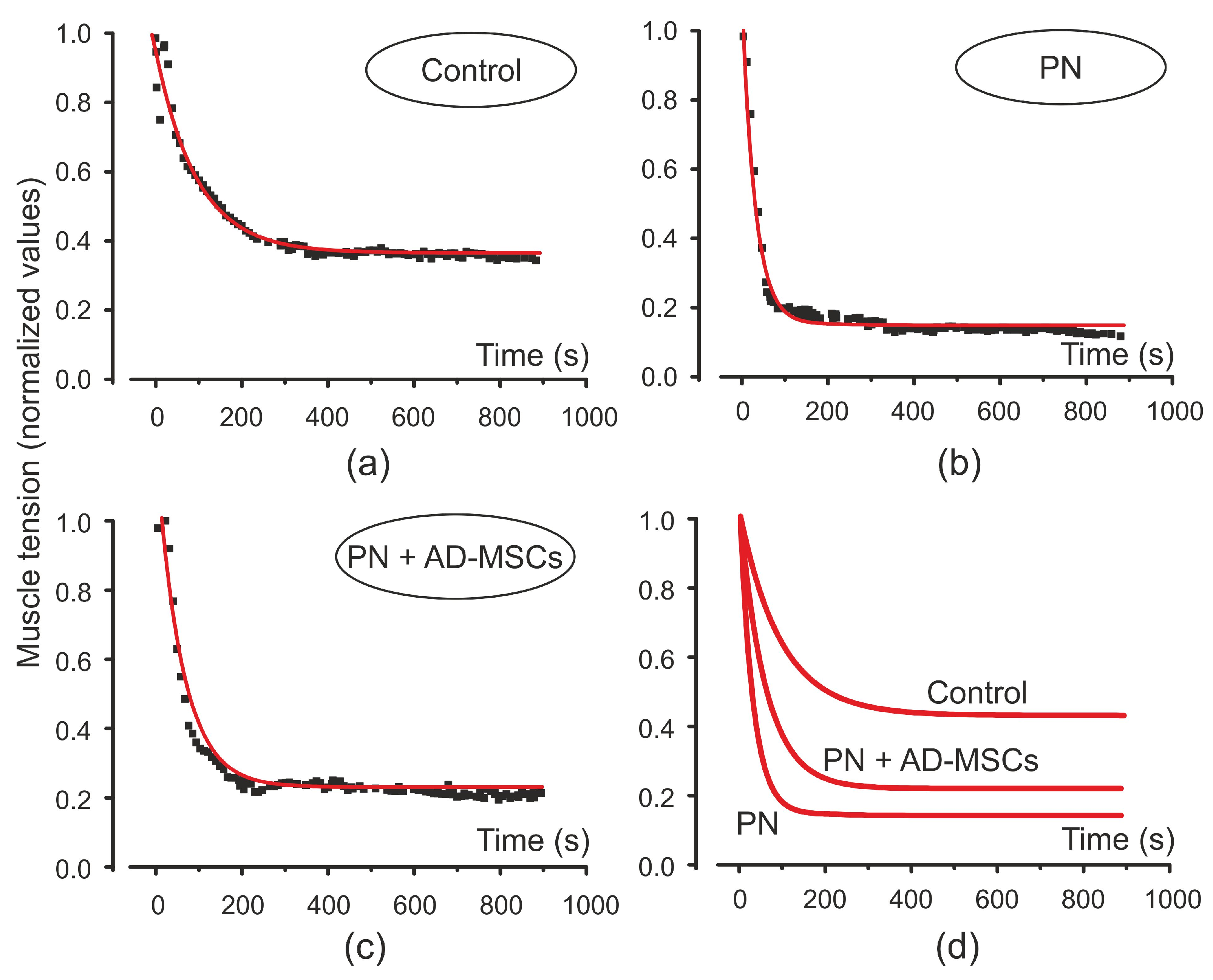{kind=link}
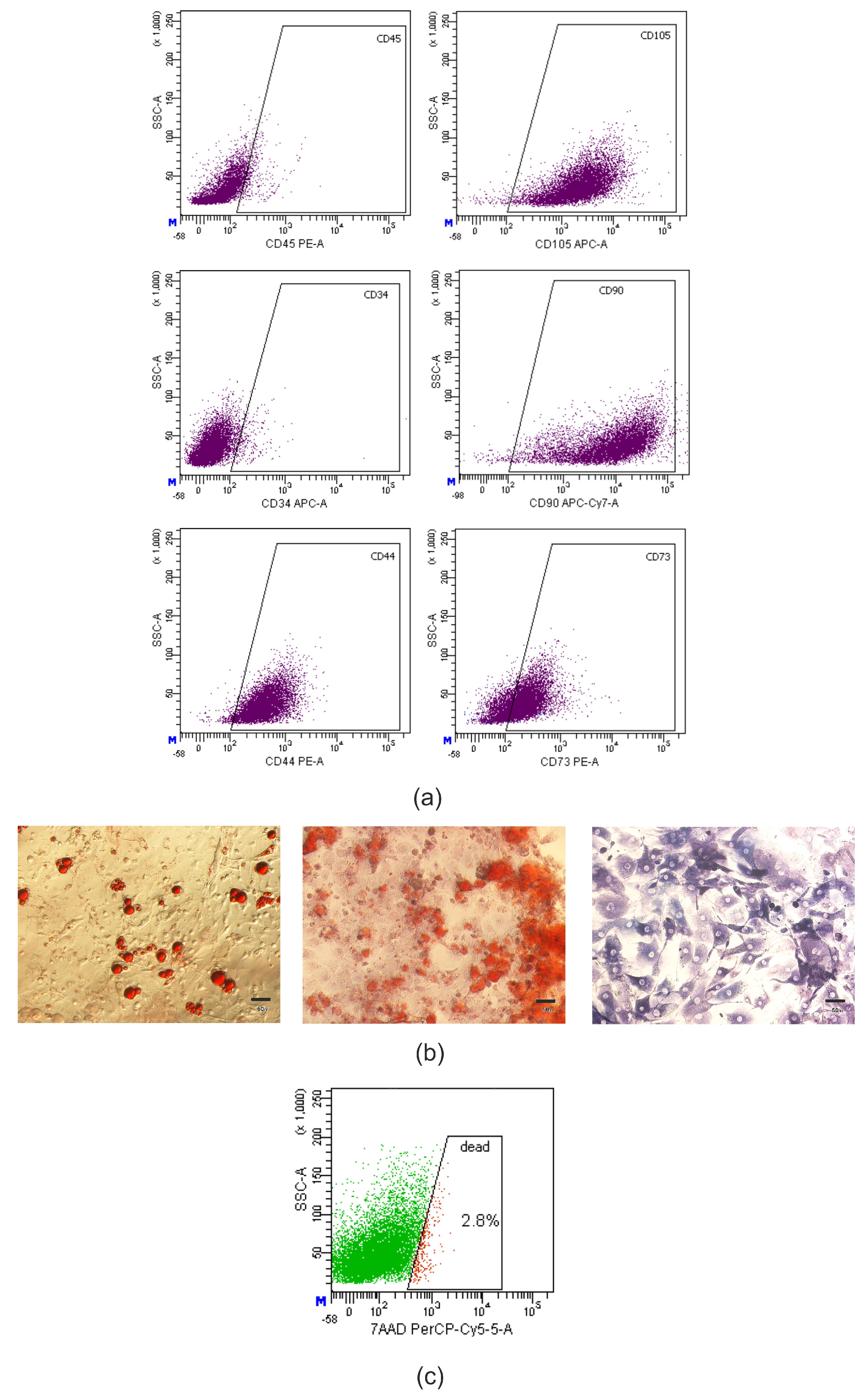{kind=link}
Abstract
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Stem Cells Isolation and Characterization
4.2.1. Adipose-Derived Mesenchymal Stromal Cells Isolation and Culture
4.2.2. Transplantation of AD-MSCs
4.3. Electrophysiological Experiment
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Caruso, D.; Scurati, S.; Roglio, I.; Nobbio, L.; Schenone, A.; Melcangi, R.C. Neuroactive Steroid Levels in a Transgenic Rat Model of CMT1A Neuropathy. J. Mol. Neurosci. 2008, 34, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Morena, J.; Gupta, A.; Hoyle, J.C. Charcot-Marie-Tooth: From Molecules to Therapy. Int. J. Mol. Sci. 2019, 20, 3419. [Google Scholar] [CrossRef]
- Kenis-Coskun, O.; Matthews, D.J. Rehabilitation issues in Charcot-Marie-Tooth disease. J. Pediatr. Rehabil. Med. 2016, 9, 31–34. [Google Scholar] [CrossRef]
- Wrabetz, L.; Feltri, M.L.; Quattrini, A.; Imperiale, D.; Previtali, S.C.; D’Antonio, M.; Martini, R.; Yin, X.; Trapp, B.D.; Zhou, L.; et al. P0 Glycoprotein Overexpression Causes Congenital Hypomyelination of Peripheral Nerves. J. Cell Biol. 2000, 148, 1021–1034. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, G.A. Penetrance of the hereditary motor and sensory neuropathy la mutation: Assessment by nerve conduction studies. Neurology 1991, 41, 547. [Google Scholar] [CrossRef] [PubMed]
- Govbakh, I.O.; Zavodovskiy, D.O.; Bulgakova, N.V.; Sokołowska, I.V.; Maznychenko, A.V.; Vasylenko, D.A. Nerve Conduction and Neuromuscular Transmission in C57Bl/6 Mice with Genetically Determined Peripheral Neuropathy. Neurophysiology 2019, 51, 248–252. [Google Scholar] [CrossRef]
- Govbakh, I.O.; Zavodovskiy, D.O.; Bulgakova, N.V.; Tsupykov, O.M.; Vasylenko, D.A.; Maznychenko, A.V. Coordination of Locomotor Activity in Transgenic C57Bl/6 Mice with Hereditary Neuropathy. Neurophysiology 2019, 51, 353–357. [Google Scholar] [CrossRef]
- Sivera, R.; Lupo, V.; Frasquet, M.; Argente-Escrig, H.; Alonso-Pérez, J.; Díaz-Manera, J.; Querol, L.; García-Romero, M.M.; Pascual, S.I.; García-Sobrino, T.; et al. Charcot–Marie–Tooth disease due to MORC2 mutations in Spain. Eur. J. Neurol. 2021, 28, 3001–3011. [Google Scholar] [CrossRef]
- Rubtsov, V.; Govbach, I.; Ustymenko, A.; Kyryk, V.; Tsupykov, O. The effects of adipose-derived multipotent mesenchymal stromal cells transplantation on motor activity and function of the sciatic nerve in mice with peripheral neuropathy. Cell Organ Transplantol. 2020, 8, 159–165. [Google Scholar] [CrossRef]
- Ma, K.H.; Hung, H.A.; Srinivasan, R.; Xie, H.; Orkin, S.H.; Svaren, J. Regulation of Peripheral Nerve Myelin Maintenance by Gene Repression through Polycomb Repressive Complex 2. J. Neurosci. 2015, 35, 8640–8652. [Google Scholar] [CrossRef] [PubMed]
- Boutary, S.; Echaniz-Laguna, A.; Adams, D.; Loisel-Duwattez, J.; Schumacher, M.; Massaad, C.; Massaad-Massade, L. Treating PMP22 gene duplication-related Charcot-Marie-Tooth disease: The past, the present and the future. Transl. Res. 2021, 227, 100–111. [Google Scholar] [CrossRef]
- Wallace, A.; Pietrusz, A.; Dewar, E.; Dudziec, M.; Jones, K.; Hennis, P.; Sterr, A.; Baio, G.; Machado, P.; Laurá, M.; et al. Community exercise is feasible for neuromuscular diseases and can improve aerobic capacity. Neurology 2019, 92, e1773–e1785. [Google Scholar] [CrossRef]
- Prada, V.; Schizzi, S.; Poggi, I.; Mori, L.; Gemelli, C.; Hamedani, M.; Accogli, S.; Maggi, G.; Grandis, M.; Schenone, A.; et al. Hand Rehabilitation Treatment for Charcot-Marie-Tooth Disease: An Open Label Pilot Study. J. Neurol. Neurophysiol. 2018, 9, 465. [Google Scholar] [CrossRef]
- Leal, A.; Ichim, T.E.; Marleau, A.M.; Lara, F.; Kaushal, S.; Riordan, N.H. Immune effects of mesenchymal stem cells: Implications for Charcot–Marie–Tooth disease. Cell. Immunol. 2008, 253, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Sottile, V.; Jackson, L.; Jones, D.R.; Scotting, P. Adult mesenchymal stem cells: Differentiation potential and therapeutic applications. J. Postgrad. Med. 2007, 53, 121–127. [Google Scholar] [CrossRef]
- Wang, M.; Crisostomo, P.R.; Herring, C.; Meldrum, K.K.; Meldrum, D.R. Human progenitor cells from bone marrow or adipose tissue produce VEGF, HGF, and IGF-I in response to TNF by a p38 MAPK-dependent mechanism. Am. J. Physiol. Integr. Comp. Physiol. 2006, 291, R880–R884. [Google Scholar] [CrossRef]
- Wankhade, U.D.; Shen, M.; Kolhe, R.; Fulzele, S. Advances in Adipose-Derived Stem Cells Isolation, Characterization, and Application in Regenerative Tissue Engineering. Stem Cells Int. 2016, 2016, 1–6. [Google Scholar] [CrossRef]
- Govbakh, I.; Smozhanik, K.; Patseva, M.; Rubtsov, V.; Ustymenko, A.; Kyryk, V.; Tsupykov, O. Ultrastructural analysis of sciatic nerve in mice with peripheral neuropathy after transplantation of adipose-derived multipotent mesenchymal stromal cells. Fiziol. Zh. 2021, 67, 17–26. [Google Scholar] [CrossRef]
- Andrzejewska, A.; Dabrowska, S.; Lukomska, B.; Janowski, M. Mesenchymal Stem Cells for Neurological Disorders. Adv. Sci. 2021, 8, 2002944. [Google Scholar] [CrossRef] [PubMed]
- Mukai, T.; Tojo, A.; Nagamura-Inoue, T. Mesenchymal stromal cells as a potential therapeutic for neurological disorders. Regen. Ther. 2018, 9, 32–37. [Google Scholar] [CrossRef]
- Yao, P.; Zhou, L.; Zhu, L.; Zhou, B.; Yu, Q. Mesenchymal Stem Cells: A Potential Therapeutic Strategy for Neurodegenerative Diseases. Eur. Neurol. 2020, 83, 235–241. [Google Scholar] [CrossRef]
- Paradisi, M.; Alviano, F.; Pirondi, S.; Lanzoni, G.; Fernandez, M.; Lizzo, G.; Giardino, L.; Giuliani, A.; Costa, R.; Marchionni, C.; et al. Human Mesenchymal Stem Cells Produce Bioactive Neurotrophic Factors: Source, Individual Variability and Differentiation Issues. Int. J. Immunopathol. Pharmacol. 2014, 27, 391–402. [Google Scholar] [CrossRef]
- Kyurkchiev, D. Secretion of immunoregulatory cytokines by mesenchymal stem cells. World J. Stem Cells 2014, 6, 552–570. [Google Scholar] [CrossRef] [PubMed]
- Martini, R. Neuregulin-1 alleviates Charcot-Marie-Tooth disease in rats. Nat. Med. 2014, 20, 984–985. [Google Scholar] [CrossRef]
- Park, S.; Jung, N.; Myung, S.; Choi, Y.; Chung, K.W.; Choi, B.-O.; Jung, S.-C. Differentiation of Human Tonsil-Derived Mesenchymal Stem Cells into Schwann-Like Cells Improves Neuromuscular Function in a Mouse Model of Charcot-Marie-Tooth Disease Type 1A. Int. J. Mol. Sci. 2018, 19, 2393. [Google Scholar] [CrossRef]
- Pan, H.-C.; Cheng, F.-C.; Chen, C.-J.; Lai, S.-Z.; Lee, C.-W.; Yang, D.-Y.; Chang, M.-H.; Ho, S.-P. Post-injury regeneration in rat sciatic nerve facilitated by neurotrophic factors secreted by amniotic fluid mesenchymal stem cells. J. Clin. Neurosci. 2007, 14, 1089–1098. [Google Scholar] [CrossRef]
- Juárez, P.; Palau, F. Neural and Molecular Features on Charcot-Marie-Tooth Disease Plasticity and Therapy. Neural Plast. 2012, 2012, 1–11. [Google Scholar] [CrossRef]
- Keilhoff, G.; Goihl, A.; Langnäse, K.; Fansa, H.; Wolf, G. Transdifferentiation of mesenchymal stem cells into Schwann cell-like myelinating cells. Eur. J. Cell Biol. 2006, 85, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Marconi, S.; Castiglione, G.; Turano, E.; Bissolotti, G.; Angiari, S.; Farinazzo, A.; Constantin, G.; Bedogni, G.; Bedogni, A.; Bonetti, B. Human Adipose-Derived Mesenchymal Stem Cells Systemically Injected Promote Peripheral Nerve Regeneration in the Mouse Model of Sciatic Crush. Tissue Eng. Part A 2012, 18, 1264–1272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.-Q.; Wang, Y.; Li, Z.-W.; Luo, M.; Li, Y.-J. Biological conduits combining bone marrow mesenchymal stem cells and extracellular matrix to treat long-segment sciatic nerve defects. Neural Regen. Res. 2015, 10, 965–971. [Google Scholar] [CrossRef]
- Zarbakhsh, S.; Goudarzi, N.; Shirmohammadi, M.; Safari, M. Histological Study of Bone Marrow and Umbilical Cord Stromal Cell Transplantation in Regenerating Rat Peripheral Nerve. Cell J. 2017, 17, 668–677. [Google Scholar]
- Fraser, J.K.; Wulur, I.; Alfonso, Z.; Hedrick, M.H. Fat tissue: An underappreciated source of stem cells for biotechnology. Trends Biotechnol. 2006, 24, 150–154. [Google Scholar] [CrossRef]
- Parker, A.M.; Katz, A.J. Adipose-derived stem cells for the regeneration of damaged tissues. Expert Opin. Biol. Ther. 2006, 6, 567–578. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Chen, J.; Cui, Y.; Lu, M.; Elias, S.B.; Mitchell, J.B.; Hammill, L.; Vanguri, P.; Chopp, M. Human bone marrow stromal cell treatment improves neurological functional recovery in EAE mice. Exp. Neurol. 2005, 195, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Na Zhou, L.; Wang, J.C.; Zilundu, P.L.M.; Wang, Y.Q.; Guo, W.P.; Zhang, S.X.; Luo, H.; Zhou, J.H.; Deng, R.D.; Chen, D.F. A comparison of the use of adipose-derived and bone marrow-derived stem cells for peripheral nerve regeneration in vitro and in vivo. Stem Cell Res. Ther. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Yang, J.-J.; Yang, X.; Liu, Z.-Q.; Hu, S.-Y.; Du, Z.-Y.; Feng, L.-L.; Liu, J.-F.; Chen, Y.-D. Transplantation of Adipose Tissue-Derived Stem Cells Overexpressing Heme Oxygenase-1 Improves Functions and Remodeling of Infarcted Myocardium in Rabbits. Tohoku J. Exp. Med. 2012, 226, 231–241. [Google Scholar] [CrossRef]
- Chung, J.Y.; Kim, W.; Im, W.; Yoo, D.Y.; Choi, J.H.; Hwang, I.K.; Won, M.-H.; Chang, I.B.; Cho, B.M.; Hwang, H.S.; et al. Neuroprotective effects of adipose-derived stem cells against ischemic neuronal damage in the rabbit spinal cord. J. Neurol. Sci. 2012, 317, 40–46. [Google Scholar] [CrossRef]
- Ustymenko, A.; Kyryk, V.; Lutsenko, T.; Tsupykov, O.; Butenko, G. Morphofunctional properties of adipose-derived multipotent mesenchymal stromal cells in vitro in ovariectomized mice of different ages. Cell Organ Transplantol. 2019, 7, 158–167. [Google Scholar] [CrossRef][Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Viswanathan, S.; Shi, Y.; Galipeau, J.; Krampera, M.; Leblanc, K.; Martin, I.; Nolta, J.; Phinney, D.G.; Sensebe, L. Mesenchymal stem versus stromal cells: International Society for Cell & Gene Therapy (ISCT(R)) Mesenchymal Stromal Cell committee position statement on nomenclature. Cytotherapy 2019, 21, 1019–1024. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Govbakh, I.; Kyryk, V.; Ustymenko, A.; Rubtsov, V.; Tsupykov, O.; Bulgakova, N.V.; Zavodovskiy, D.O.; Sokolowska, I.; Maznychenko, A. Stem Cell Therapy Enhances Motor Activity of Triceps Surae Muscle in Mice with Hereditary Peripheral Neuropathy. Int. J. Mol. Sci. 2021, 22, 12026. https://doi.org/10.3390/ijms222112026
Govbakh I, Kyryk V, Ustymenko A, Rubtsov V, Tsupykov O, Bulgakova NV, Zavodovskiy DO, Sokolowska I, Maznychenko A. Stem Cell Therapy Enhances Motor Activity of Triceps Surae Muscle in Mice with Hereditary Peripheral Neuropathy. International Journal of Molecular Sciences. 2021; 22(21):12026. https://doi.org/10.3390/ijms222112026
Chicago/Turabian StyleGovbakh, Iryna, Vitalii Kyryk, Alina Ustymenko, Volodymyr Rubtsov, Oleg Tsupykov, Nataliya V. Bulgakova, Danylo O. Zavodovskiy, Inna Sokolowska, and Andriy Maznychenko. 2021. "Stem Cell Therapy Enhances Motor Activity of Triceps Surae Muscle in Mice with Hereditary Peripheral Neuropathy" International Journal of Molecular Sciences 22, no. 21: 12026. https://doi.org/10.3390/ijms222112026
APA StyleGovbakh, I., Kyryk, V., Ustymenko, A., Rubtsov, V., Tsupykov, O., Bulgakova, N. V., Zavodovskiy, D. O., Sokolowska, I., & Maznychenko, A. (2021). Stem Cell Therapy Enhances Motor Activity of Triceps Surae Muscle in Mice with Hereditary Peripheral Neuropathy. International Journal of Molecular Sciences, 22(21), 12026. https://doi.org/10.3390/ijms222112026