
Effects of Hypoxia on RNA Cargo in Extracellular Vesicles from Human Adipose-Derived Stromal/Stem Cells

 , ,
, ,  ,
,
Abstract
:1. Introduction
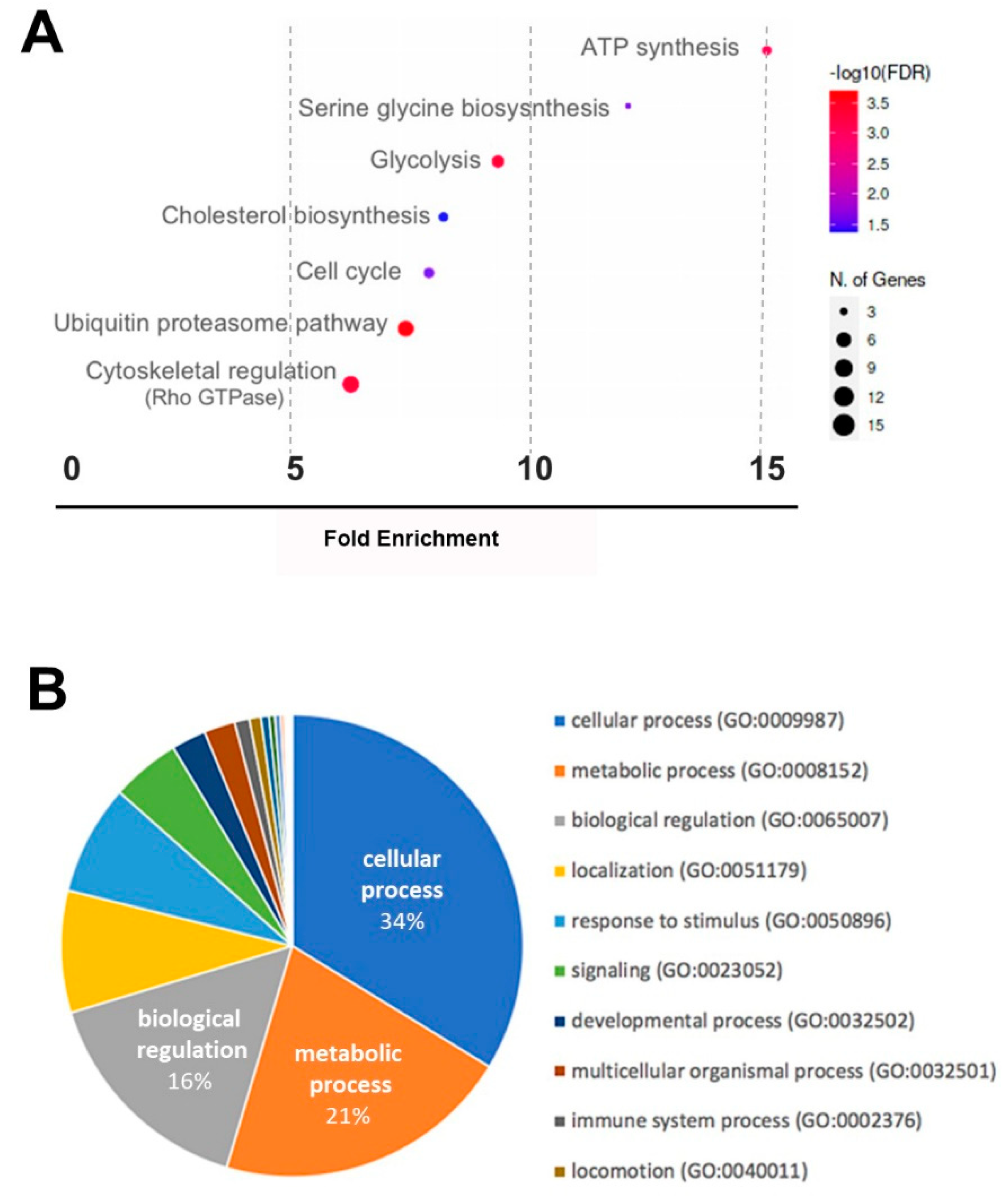
2. Results
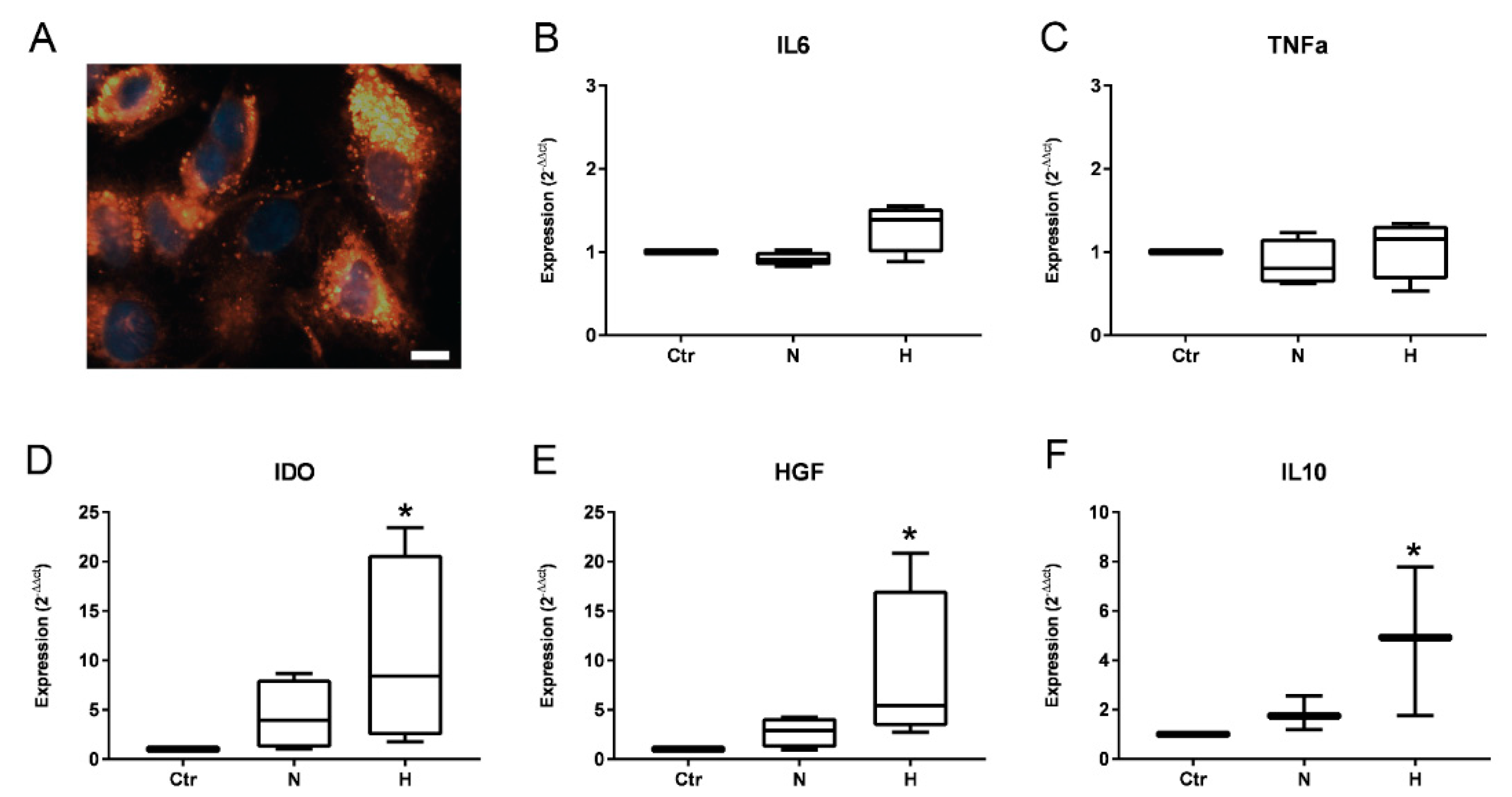
2.1. Isolation and Characterization of EVs
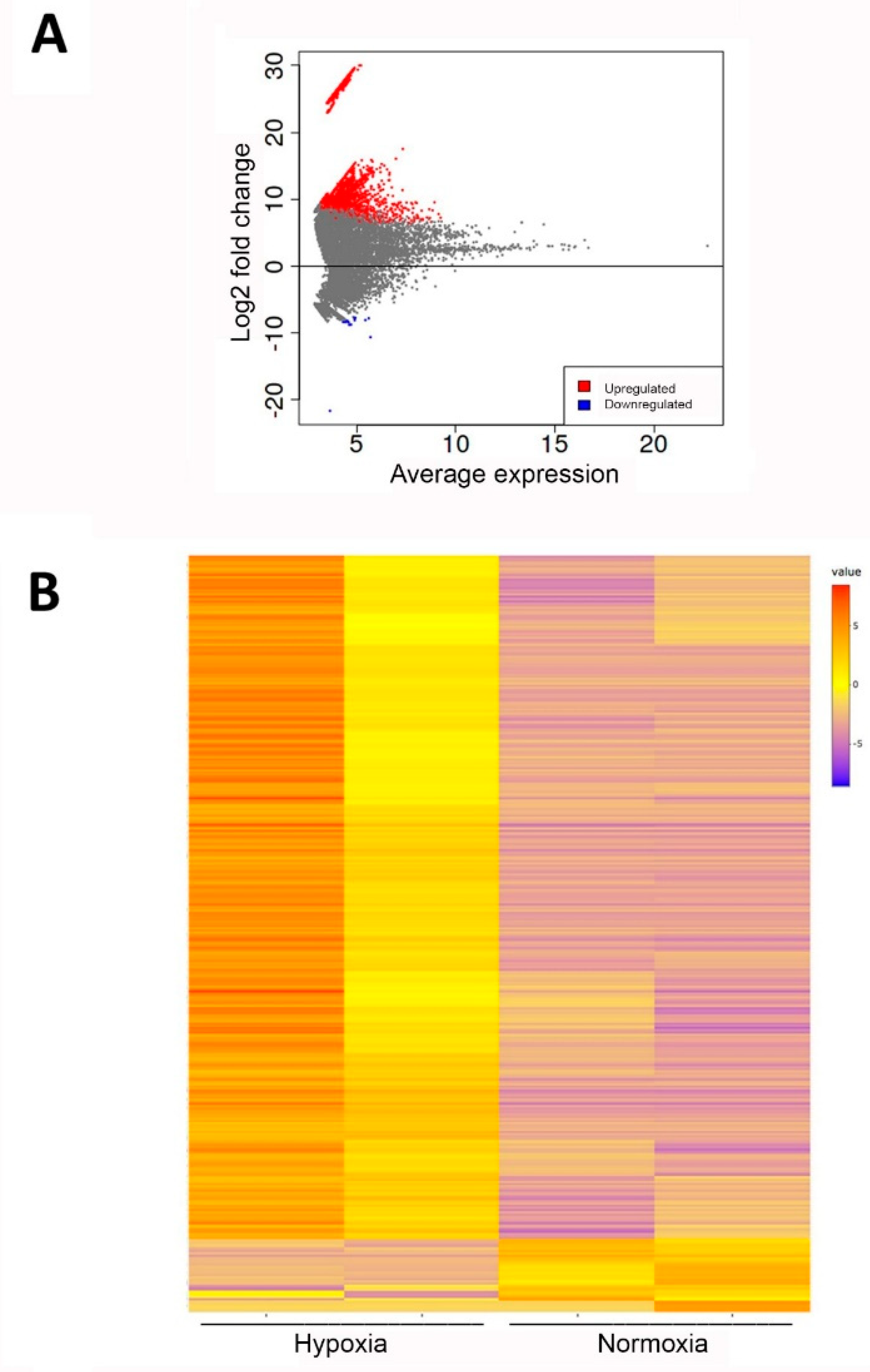
2.2. Whole RNA Transcriptomics
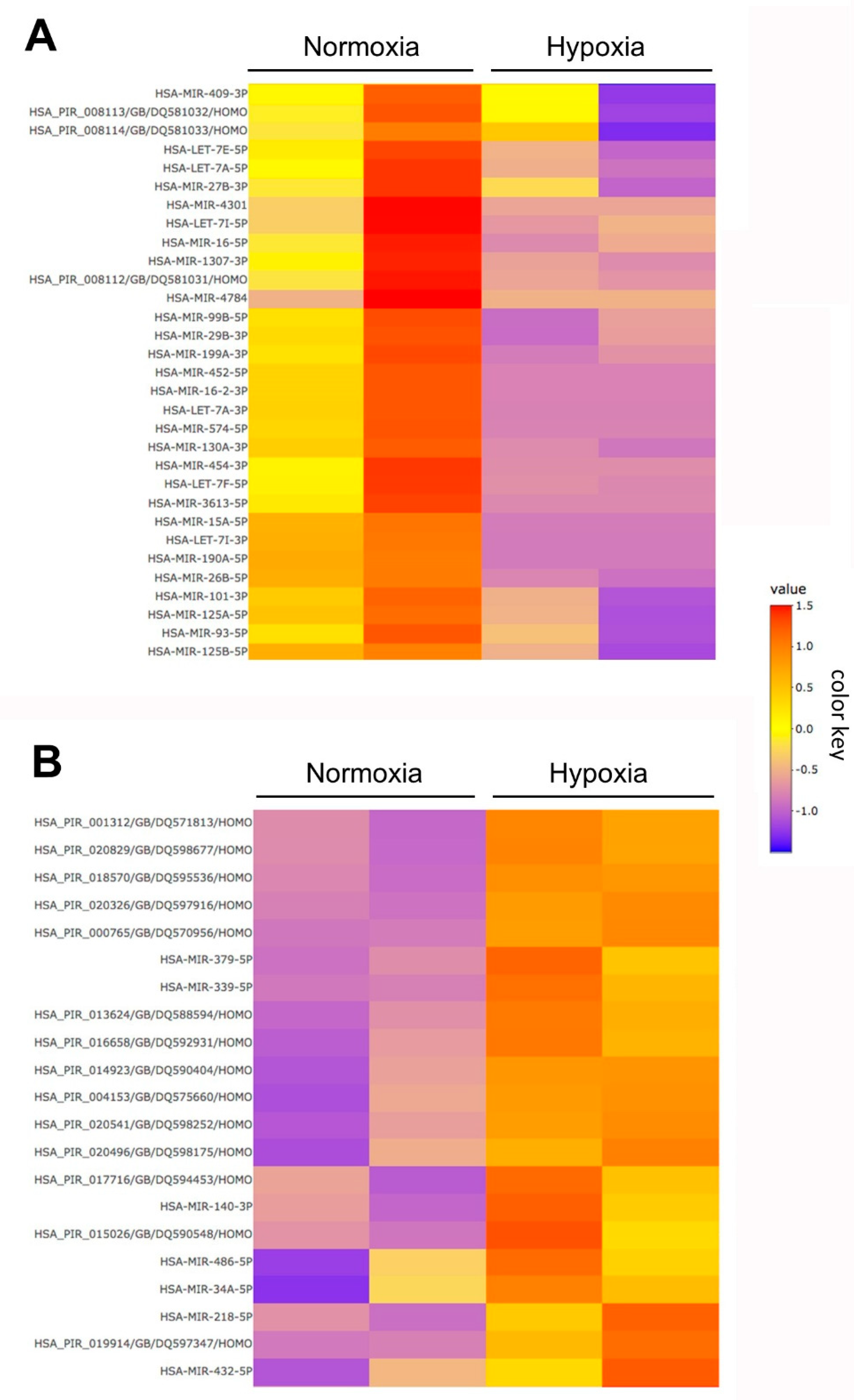
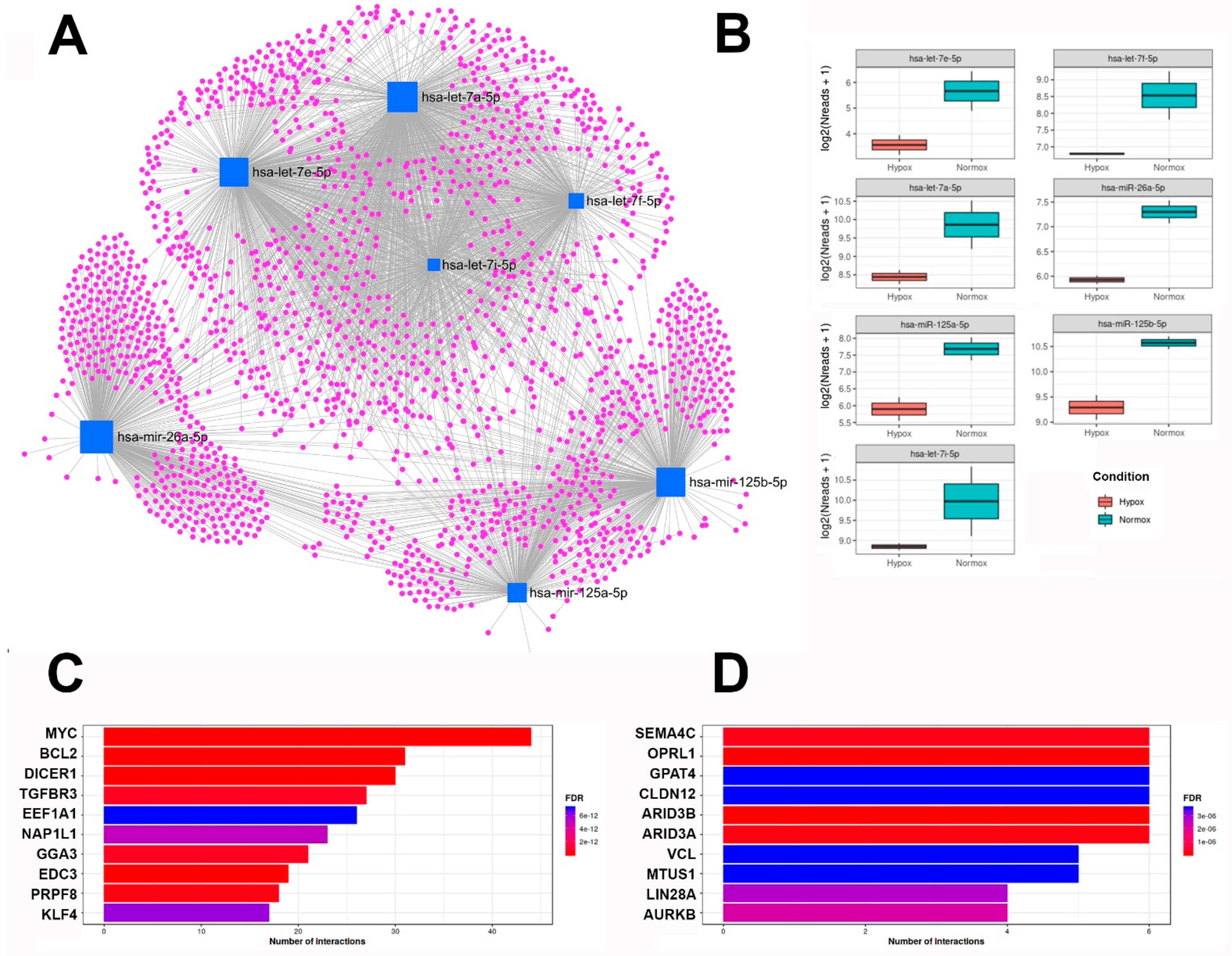
2.3. Small RNA Transcriptomics
2.4. Effect of EVs on Renal Epithelial Cells
3. Discussion
4. Materials and Methods
4.1. Cell Isolation, Culture, and Characterization
4.2. Preconditioning of ASCs
4.3. Isolation of Extracellular Vesicles
4.4. Characterization of Extracellular Vesicles
4.5. Isolation of Whole RNA from EVs for Sequencing
4.6. Isolation of miR from ASC-EVs for Sequencing
4.7. Effect of EVs on Renal Tubular Epithelial Cells
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASCs | Adipose-derived stromal/stem cells |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| EVs | Extracellular vesicles |
| FBS | Fetal bovine serum |
| hEVs | Extracellular vesicles after hypoxic pretreatment |
| HGF | Hepatocyte growth factor |
| IDO | Indoleamine 2,3-dioxygenase 1 |
| IL | Interleukin |
| miR | microRNA |
| MSCs | Mesenchymal stromal/stem cells |
| nEVs | Extracellular vesicles after normoxic culture |
| PCM | Preconditioned medium |
| PCR | Polymerase chain reaction |
| piR | Piwi-interacting RNA |
| RT | Room temperature |
| SEC TEC | Size exclusion chromatography Tubular epithelial cells |
| TNFα | Tumor necrosis factor alpha |
References
- Mol, E.A.; Goumans, M.-J.; Doevendans, P.A.; Sluijter, J.P.G.; Vader, P. Higher functionality of extracellular vesicles isolated using size-exclusion chromatography compared to ultracentrifugation. Nanomedicine 2017, 13, 2061–2065. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hade, M.D.; Suire, C.N.; Suo, Z. Mesenchymal Stem Cell-Derived Exosomes: Applications in Regenerative Medicine. Cells 2021, 10, 1959. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, L.; Huo, Y.; Yang, Y.; Wang, Y. Hypoxia-pretreated human MSCs attenuate acute kidney injury through enhanced angiogenic and antioxidative capacities. Biomed Res. Int. 2014, 2014, 462472. [Google Scholar] [CrossRef]
- Ekström, K.; Valadi, H.; Sjöstrand, M.; Malmhäll, C.; Bossios, A.; Eldh, M.; Lötvall, J. Characterization of mRNA and microRNA in human mast cell-derived exosomes and their transfer to other mast cells and blood CD34 progenitor cells. J. Extracell. Vesicles 2012, 1, 18389. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Rani, S.; Ryan, A.E.; Griffin, M.D.; Ritter, T. Mesenchymal Stem Cell-derived Extracellular Vesicles: Toward Cell-free Therapeutic Applications. Mol. Ther. 2015, 23, 812–823. [Google Scholar] [CrossRef] [Green Version]
- Barreca, M.M.; Cancemi, P.; Geraci, F. Mesenchymal and Induced Pluripotent Stem Cells-Derived Extracellular Vesicles: The New Frontier for Regenerative Medicine? Cells 2020, 9, 1163. [Google Scholar] [CrossRef]
- Schäfer, R.; Spohn, G.; Baer, P.C. Mesenchymal Stem/Stromal Cells in Regenerative Medicine: Can Preconditioning Strategies Improve Therapeutic Efficacy? Transfus. Med. Hemother. 2016, 43, 256–267. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Rong, Y.; Wang, J.; Zhou, Z.; Ge, X.; Ji, C.; Jiang, D.; Gong, F.; Li, L.; Chen, J.; et al. Exosome-shuttled miR-216a-5p from hypoxic preconditioned mesenchymal stem cells repair traumatic spinal cord injury by shifting microglial M1/M2 polarization. J. Neuroinflamm. 2020, 17, 47. [Google Scholar] [CrossRef] [Green Version]
- Baer, P.C.; Overath, J.M.; Urbschat, A.; Schubert, R.; Koch, B.; Bohn, A.A.; Geiger, H. Effect of Different Preconditioning Regimens on the Expression Profile of Murine Adipose-Derived Stromal/Stem Cells. Int. J. Mol. Sci. 2018, 19, 1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Ruffier, M.; Kähäri, A.; Komorowska, M.; Keenan, S.; Laird, M.; Longden, I.; Proctor, G.; Searle, S.; Staines, D.; Taylor, K.; et al. Ensembl core software resources: Storage and programmatic access for DNA sequence and genome annotation. Database 2017, 2017, bax020. [Google Scholar] [CrossRef] [PubMed]
- Ge, X. iDEP Web Application for RNA-Seq Data Analysis. Methods Mol. Biol. 2021, 2284, 417–443. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.-P.; Mushayamaha, T.; Thomas, P.D. PANTHER version 16: A revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- Chang, L.; Zhou, G.; Soufan, O.; Xia, J. miRNet 2.0: Network-based visual analytics for miRNA functional analysis and systems biology. Nucleic Acids Res. 2020, 48, W244–W251. [Google Scholar] [CrossRef]
- Hsu, S.-D.; Lin, F.-M.; Wu, W.-Y.; Liang, C.; Huang, W.-C.; Chan, W.-L.; Tsai, W.-T.; Chen, G.-Z.; Lee, C.-J.; Chiu, C.-M.; et al. miRTarBase: A database curates experimentally validated microRNA-target interactions. Nucleic Acids Res. 2011, 39, D163–D169. [Google Scholar] [CrossRef] [Green Version]
- Luck, J.; Weil, B.D.; Lowdell, M.; Mosahebi, A. Adipose-Derived Stem Cells for Regenerative Wound Healing Applications: Understanding the Clinical and Regulatory Environment. Aesthet. Surg. J. 2020, 40, 784–799. [Google Scholar] [CrossRef]
- Toh, W.S.; Zhang, B.; Lai, R.C.; Lim, S.K. Immune regulatory targets of mesenchymal stromal cell exosomes/small extracellular vesicles in tissue regeneration. Cytotherapy 2018, 20, 1419–1426. [Google Scholar] [CrossRef]
- Almeria, C.; Weiss, R.; Roy, M.; Tripisciano, C.; Kasper, C.; Weber, V.; Egger, D. Hypoxia Conditioned Mesenchymal Stem Cell-Derived Extracellular Vesicles Induce Increased Vascular Tube Formation in vitro. Front. Bioeng. Biotechnol. 2019, 7, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.S.; Lai, R.C.; Lee, M.M.; Choo, A.B.H.; Lee, C.N.; Lim, S.K. Mesenchymal stem cell secretes microparticles enriched in pre-microRNAs. Nucleic Acids Res. 2010, 38, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Yuan, T.; Tschannen, M.; Sun, Z.; Jacob, H.; Du, M.; Liang, M.; Dittmar, R.L.; Liu, Y.; Liang, M.; et al. Characterization of human plasma-derived exosomal RNAs by deep sequencing. BMC Genom. 2013, 14, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böing, A.N.; van der Pol, E.; Grootemaat, A.E.; Coumans, F.A.W.; Sturk, A.; Nieuwland, R. Single-step isolation of extracellular vesicles by size-exclusion chromatography. J. Extracell. Vesicles 2014, 3, 23430. [Google Scholar] [CrossRef]
- Monguió-Tortajada, M.; Gálvez-Montón, C.; Bayes-Genis, A.; Roura, S.; Borràs, F.E. Extracellular vesicle isolation methods: Rising impact of size-exclusion chromatography. Cell. Mol. Life Sci. 2019, 76, 2369–2382. [Google Scholar] [CrossRef]
- Geßner, A.; Koch, B.; Klann, K.; Fuhrmann, D.C.; Farmand, S.; Schubert, R.; Münch, C.; Geiger, H.; Baer, P.C. Characterization of Extracellular Vesicles from Preconditioned Human Adipose-Derived Stromal/Stem Cells. Int. J. Mol. Sci. 2021, 22, 2873. [Google Scholar] [CrossRef]
- Kang, S.; Kim, S.-M.; Sung, J.-H. Cellular and molecular stimulation of adipose-derived stem cells under hypoxia. Cell Biol. Int. 2014, 38, 553–562. [Google Scholar] [CrossRef]
- Lando, D.; Peet, D.J.; Gorman, J.J.; Whelan, D.A.; Whitelaw, M.L.; Bruick, R.K. FIH-1 is an asparaginyl hydroxylase enzyme that regulates the transcriptional activity of hypoxia-inducible factor. Genes Dev. 2002, 16, 1466–1471. [Google Scholar] [CrossRef] [Green Version]
- Jha, S.K.; Rauniyar, K.; Chronowska, E.; Mattonet, K.; Maina, E.W.; Koistinen, H.; Stenman, U.-H.; Alitalo, K.; Jeltsch, M. KLK3/PSA and cathepsin D activate VEGF-C and VEGF-D. Elife 2019, 8, e44478. [Google Scholar] [CrossRef]
- Son, J.-W.; Park, J.; Kim, Y.E.; Ha, J.; Park, D.W.; Chang, M.-S.; Koh, S.-H. Glia-Like Cells from Late-Passage Human MSCs Protect Against Ischemic Stroke Through IGFBP-4. Mol. Neurobiol. 2019, 56, 7617–7630. [Google Scholar] [CrossRef]
- Kojima, Y.; Tsuchiya, A.; Ogawa, M.; Nojiri, S.; Takeuchi, S.; Watanabe, T.; Nakajima, K.; Hara, Y.; Yamashita, J.; Kikuta, J.; et al. Mesenchymal stem cells cultured under hypoxic conditions had a greater therapeutic effect on mice with liver cirrhosis compared to those cultured under normal oxygen conditions. Regen. Ther. 2019, 11, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, E.; Izawa, T.; Juniantito, V.; Kuwamura, M.; Sugiura, K.; Takeuchi, T.; Yamate, J. Involvement of endogenous prostaglandin E2 in tubular epithelial regeneration through inhibition of apoptosis and epithelial-mesenchymal transition in cisplatin-induced rat renal lesions. Histol. Histopathol. 2010, 25, 995–1007. [Google Scholar] [CrossRef] [PubMed]
- Bruno, S.; Grange, C.; Collino, F.; Deregibus, M.C.; Cantaluppi, V.; Biancone, L.; Tetta, C.; Camussi, G. Microvesicles derived from mesenchymal stem cells enhance survival in a lethal model of acute kidney injury. PLoS ONE 2012, 7, e33115. [Google Scholar] [CrossRef]
- Gao, F.; Hu, X.; Xie, X.; Xu, Q.; Wang, Y.; Liu, X.; Xiang, M.; Sun, Y.; Wang, J. Heat shock protein 90 protects rat mesenchymal stem cells against hypoxia and serum deprivation-induced apoptosis via the PI3K/Akt and ERK1/2 pathways. J. Zhejiang Univ. Sci. B 2010, 11, 608–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Hozumi, Y.; Martelli, A.M.; Iino, M.; Goto, K. Nucleosome assembly proteins NAP1L1 and NAP1L4 modulate p53 acetylation to regulate cell fate. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 118560. [Google Scholar] [CrossRef] [PubMed]
- Trionfini, P.; Benigni, A.; Remuzzi, G. MicroRNAs in kidney physiology and disease. Nat. Rev. Nephrol. 2015, 11, 23–33. [Google Scholar] [CrossRef]
- Lee, H.; Han, S.; Kwon, C.S.; Lee, D. Biogenesis and regulation of the let-7 miRNAs and their functional implications. Protein Cell 2016, 7, 100–113. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.-P.; Tian, T.; Wang, J.-Y.; He, J.-N.; Chen, T.; Pan, M.; Xu, L.; Zhang, H.-X.; Qiu, X.-T.; Li, C.-C.; et al. Hypoxia-elicited mesenchymal stem cell-derived exosomes facilitates cardiac repair through miR-125b-mediated prevention of cell death in myocardial infarction. Theranostics 2018, 8, 6163–6177. [Google Scholar] [CrossRef]
- Cao, J.-Y.; Wang, B.; Tang, T.-T.; Wen, Y.; Li, Z.-L.; Feng, S.-T.; Wu, M.; Liu, D.; Yin, D.; Ma, K.-L.; et al. Exosomal miR-125b-5p deriving from mesenchymal stem cells promotes tubular repair by suppression of p53 in ischemic acute kidney injury. Theranostics 2021, 11, 5248–5266. [Google Scholar] [CrossRef]
- Polesskaya, A.; Cuvellier, S.; Naguibneva, I.; Duquet, A.; Moss, E.G.; Harel-Bellan, A. Lin-28 binds IGF-2 mRNA and participates in skeletal myogenesis by increasing translation efficiency. Genes Dev. 2007, 21, 1125–1138. [Google Scholar] [CrossRef] [Green Version]
- Baer, P.C.; Bereiter-Hahn, J. Epithelial cells in culture: Injured or differentiated cells? Cell Biol. Int. 2012, 36, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Gröne, H.J.; Weber, K.; Gröne, E.; Helmchen, U.; Osborn, M. Coexpression of keratin and vimentin in damaged and regenerating tubular epithelia of the kidney. Am. J. Pathol. 1987, 129, 1–8. [Google Scholar] [PubMed]
- Collino, F.; Lopes, J.A.; Corrêa, S.; Abdelhay, E.; Takiya, C.M.; Wendt, C.H.C.; de Miranda, K.R.; Vieyra, A.; Lindoso, R.S. Adipose-Derived Mesenchymal Stromal Cells Under Hypoxia: Changes in Extracellular Vesicles Secretion and Improvement of Renal Recovery after Ischemic Injury. Cell. Physiol. Biochem. 2019, 52, 1463–1483. [Google Scholar] [CrossRef] [PubMed]
- Lindoso, R.S.; Collino, F.; Bruno, S.; Araujo, D.S.; Sant’Anna, J.F.; Tetta, C.; Provero, P.; Quesenberry, P.J.; Vieyra, A.; Einicker-Lamas, M.; et al. Extracellular vesicles released from mesenchymal stromal cells modulate miRNA in renal tubular cells and inhibit ATP depletion injury. Stem Cells Dev. 2014, 23, 1809–1819. [Google Scholar] [CrossRef] [Green Version]
- Brzoska, M.; Geiger, H.; Gauer, S.; Baer, P. Epithelial differentiation of human adipose tissue-derived adult stem cells. Biochem. Biophys. Res. Commun. 2005, 330, 142–150. [Google Scholar] [CrossRef]
- Baer, P.C.; Kuçi, S.; Krause, M.; Kuçi, Z.; Zielen, S.; Geiger, H.; Bader, P.; Schubert, R. Comprehensive phenotypic characterization of human adipose-derived stromal/stem cells and their subsets by a high throughput technology. Stem Cells Dev. 2013, 22, 330–339. [Google Scholar] [CrossRef]
- Baer, P.C.; Koch, B.; Hickmann, E.; Schubert, R.; Cinatl, J.; Hauser, I.A.; Geiger, H. Isolation, Characterization, Differentiation and Immunomodulatory Capacity of Mesenchymal Stromal/Stem Cells from Human Perirenal Adipose Tissue. Cells 2019, 8, 1346. [Google Scholar] [CrossRef] [Green Version]
- Bachurski, D.; Schuldner, M.; Nguyen, P.-H.; Malz, A.; Reiners, K.S.; Grenzi, P.C.; Babatz, F.; Schauss, A.C.; Hansen, H.P.; Hallek, M.; et al. Extracellular vesicle measurements with nanoparticle tracking analysis—An accuracy and repeatability comparison between NanoSight NS300 and ZetaView. J. Extracell. Vesicles 2019, 8, 1596016. [Google Scholar] [CrossRef]
- Wingett, S.W.; Andrews, S. FastQ Screen: A tool for multi-genome mapping and quality control. F1000Research 2018, 7, 1338. [Google Scholar] [CrossRef]
- Yousif, A.; Drou, N.; Rowe, J.; Khalfan, M.; Gunsalus, K.C. NASQAR: A web-based platform for high-throughput sequencing data analysis and visualization. BMC Bioinform. 2020, 21, 267. [Google Scholar] [CrossRef]
- Licursi, V.; Conte, F.; Fiscon, G.; Paci, P. MIENTURNET: An interactive web tool for microRNA-target enrichment and network-based analysis. BMC Bioinform. 2019, 20, 545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Ensembl No. (Transcript) | log2-Fold (hEVs/nEVs) | p Value (FDR) |
|---|---|---|---|
| CTSD | ENST00000236671.7 | 29.37 | <0.0001 |
| HSP90B1 | ENST00000299767.10 | 28.45 | <0.0001 |
| IGFBP6 | ENST00000650247 | 27.55 | <0.0001 |
| MMP2 | ENST00000543485 | 27.53 | <0.0001 |
| BNIP2 | ENST00000415213 | 27.15 | <0.0001 |
| BCL2L12 | ENST00000246784 | 26.53 | <0.0001 |
| CFL1 | ENST00000526975 | 26.51 | <0.0001 |
| TMBIM4 | ENST00000398033 | 25.89 | <0.0001 |
| CCL18 | ENST00000616054.2 | 25.17 | <0.0001 |
| PTGES | ENST00000340607 | 14.07 | 0.0001 |
| VEGFB | ENST00000543462.1 | 13.11 | 0.0006 |
| IGFBP4 | ENST00000269593.5 | 10.62 | 0.0004 |
| CCL2 | ENST00000225831 | −8.07 | 0.0495 |
| NAP1L4 | ENST00000632437.1 | −8.35 | 0.0473 |
| IDO1 | ENST00000518237 | −8.73 | 0.0367 |
| Gene Symbol | Accession | log2-Fold (hEVs/nEVs) | p Value (FDR) |
|---|---|---|---|
| hsa-piR-019914 | DQ597347 | 4.27 | <0.0001 |
| hsa-piR-13624 | DQ588594 | 4.15 | <0.0001 |
| hsa-piR-020829 | DQ598677 | 3.99 | <0.0001 |
| hsa-pir-018570 | DQ595536 | 3.83 | <0.0001 |
| hsa-pir-020541 | DQ598252 | 3.61 | 0.0032 |
| hsa-pir-001312 | DQ571813 | 3.58 | <0.0001 |
| hsa-pir-014923 | DQ590404 | 2.77 | 0.0095 |
| hsa-pir-004153 | DQ575660 | 2.57 | 0.0095 |
| hsa-piR-016658 | DQ592931 | 1.83 | 0.016 |
| hsa-piR-020326 | DQ597916 | 1.79 | 0.054 |
| hsa-piR-000765 | DQ570956 | 1.34 | 0.018 |
| hsa-miR-125b-5p | MIMAT0000423 | −1.27 | 0.040 |
| hsa-let-7i-5p | MIMAT0000415 | −1.37 | 0.098 |
| hsa-miR-26a-5p | MIMAT0000082 | −1.40 | 0.065 |
| hsa-let-7a-5p | MIMAT0000062 | −1.55 | 0.031 |
| hsa-miR-125a-5p | MIMAT0000443 | −1.80 | 0.015 |
| hsa-let-7f-5p | MIMAT0000067 | −1.92 | 0.010 |
| hsa-let-7e-5p | MIMAT0000066 | −2.34 | 0.047 |
| hsa-miR-125b-5p | hsa-let-7a-5p | hsa-miR-125a-5p | hsa-miR-26a-5p | hsa-let-7e-5p | hsa-let-7f-5p | hsa-let-7i-5p | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BMPR1B | PRDM1 | IL6R | ICAM2 | SFRP5 | MYC | LIN28A | AURKB | CDKN1A | ESRRA | HMGA2 | TET2 | HMGA2 | KLK6 | SOCS1 |
| EIF4EBP1 | IRF4 | ABTB1 | SET | LIFR | NKIRAS2 | CASP9 | RAB40C | LIN28A | VPS51 | HMGA1 | PTPN13 | EIF3J | PRDM1 | IL13 |
| HMGA2 | TP53INP1 | HK2 | CCNJ | Fas | NF2 | IL6 | ARG2 | NTRK3 | SIRT7 | CDK8 | PIK3C2A | SMC1A | IL13 | COPS8 |
| HMGA1 | STAT3 | E2F2 | ENPEP | IGF1R | NRAS | E2F2 | TNFRSF10B | CD34 | CLEC5A | EZH2 | TDG | WNT1 | MPL | COPS6 |
| GLI1 | IGF2 | MMP13 | CSNK2A1 | NEU1 | PRDM1 | IGF2BP1 | TGFBR3 | ERBB3 | GALNT14 | SMAD4 | PHB | CCND1 | CYP19A1 | GPS1 |
| NKIRAS2 | LIN28B | MAPK14 | MEGF9 | ALOX5 | TRIM71 | MPL | LIN28B | ERBB2 | STAT3 | MYC | NRAS | MPL | COPS8 | BMP4 |
| SMO | PPP1CA | EPO | MAN1B1 | EGFR | NR1I2 | CCR7 | PARP1 | BAK1 | MCL1 | GDAP1 | E2F2 | AGO1 | GPS1 | IGF1 |
| VDR | PRKRA | MUC1 | AHRR | ANGPT2 | VDR | CDKN1A | WNT1 | ARID3B | MAPK14 | CCNE1 | PIK3CG | IGF1R | CCND1 | AGO1 |
| BAK1 | BCL2 | NES | SCNN1A | FES | HMGA2 | EGFR | STAT3 | TNFAIP3 | EDN1 | NOS2 | IGF1 | IGF1 | COPS6 | NEUROG1 |
| ERBB3 | TNFAIP3 | CEBPA | VPS51 | CDKN2D | HMGA1 | RRM2 | MAP4K4 | SMAD4 | PIK3CG | IL6 | PLOD2 | LIN28A | DYRK2 | IL2 |
| ERBB2 | PIGF | ARID3A | SIRT7 | JAK2 | UHRF2 | AGO1 | TNFAIP3 | MMP11 | AKT1 | MCL1 | FUT8 | AURKB | CCL7 | AURKB |
| BMF | TBC1D1 | MXD1 | TET2 | PTH1R | IGF2 | EZH2 | HK2 | MEG3 | AMACR | ST3GAL6 | EZH2 | AGO1 | ||
| NTRK3 | FGFR2 | PIAS3 | SEMA4C | HOTTIP | EIF4EBP1 | HDAC5 | HGF | TRPC6 | TNFRSF10B | IL6 | ||||
| LIN28A | ARID3B | PCTP | SPHK1 | DRAM2 | LIFR | RAF1 | LIN28B | LARP1 | ARID3A | POSTN | ||||
| AKT1 | SMAD4 | GSS | MMP26 | EGFR | JAK2 | ZCCHC11 | TNFAIP3 | |||||||
| RAF1 | MCL1 | IKZF3 | MAP3K11 | BCL2 | ARID3A | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koch, B.; Geßner, A.; Farmand, S.; Fuhrmann, D.C.; Chiocchetti, A.G.; Schubert, R.; Baer, P.C. Effects of Hypoxia on RNA Cargo in Extracellular Vesicles from Human Adipose-Derived Stromal/Stem Cells. Int. J. Mol. Sci. 2022, 23, 7384. https://doi.org/10.3390/ijms23137384
Koch B, Geßner A, Farmand S, Fuhrmann DC, Chiocchetti AG, Schubert R, Baer PC. Effects of Hypoxia on RNA Cargo in Extracellular Vesicles from Human Adipose-Derived Stromal/Stem Cells. International Journal of Molecular Sciences. 2022; 23(13):7384. https://doi.org/10.3390/ijms23137384
Chicago/Turabian StyleKoch, Benjamin, Alec Geßner, Samira Farmand, Dominik C. Fuhrmann, Andreas G. Chiocchetti, Ralf Schubert, and Patrick C. Baer. 2022. "Effects of Hypoxia on RNA Cargo in Extracellular Vesicles from Human Adipose-Derived Stromal/Stem Cells" International Journal of Molecular Sciences 23, no. 13: 7384. https://doi.org/10.3390/ijms23137384
APA StyleKoch, B., Geßner, A., Farmand, S., Fuhrmann, D. C., Chiocchetti, A. G., Schubert, R., & Baer, P. C. (2022). Effects of Hypoxia on RNA Cargo in Extracellular Vesicles from Human Adipose-Derived Stromal/Stem Cells. International Journal of Molecular Sciences, 23(13), 7384. https://doi.org/10.3390/ijms23137384