
I2-Imidazoline Ligand CR4056 Improves Memory, Increases ApoE Expression and Reduces BBB Leakage in 5xFAD Mice

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
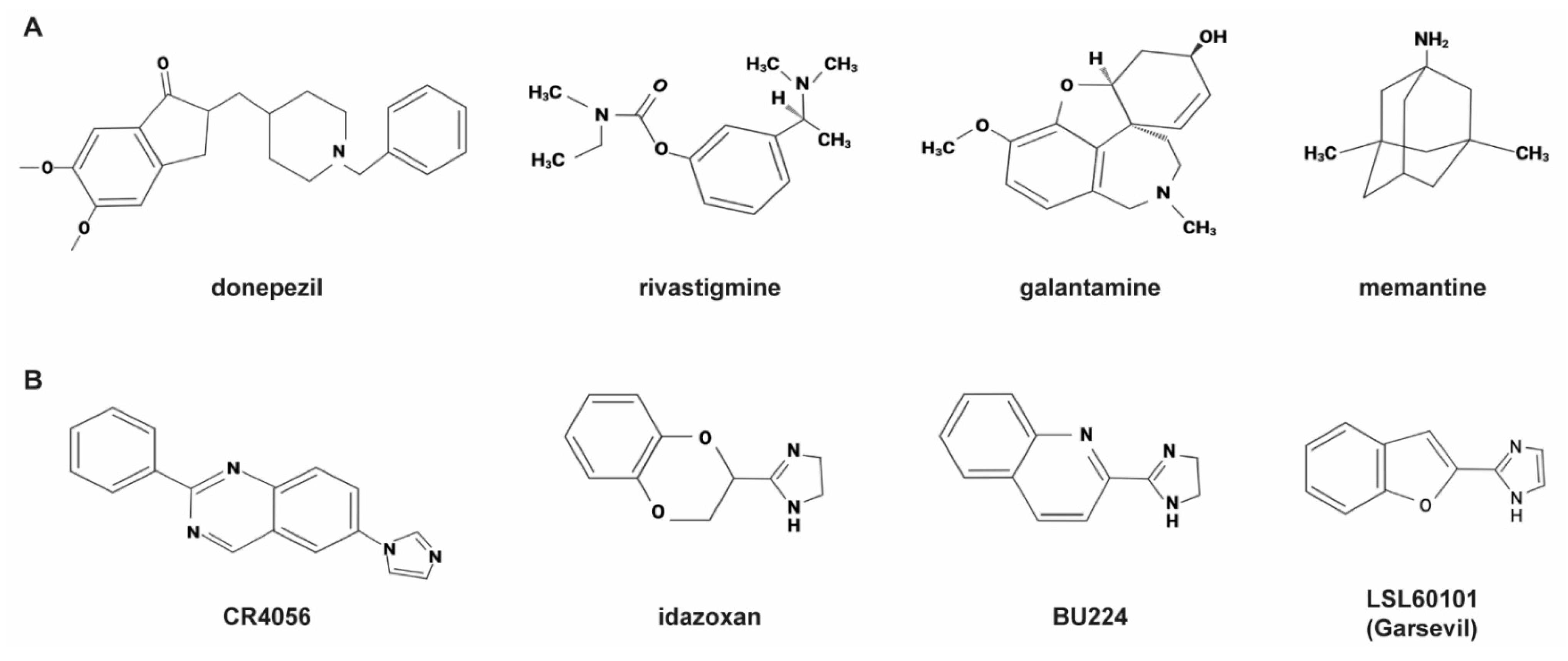
:1. Introduction
2. Results
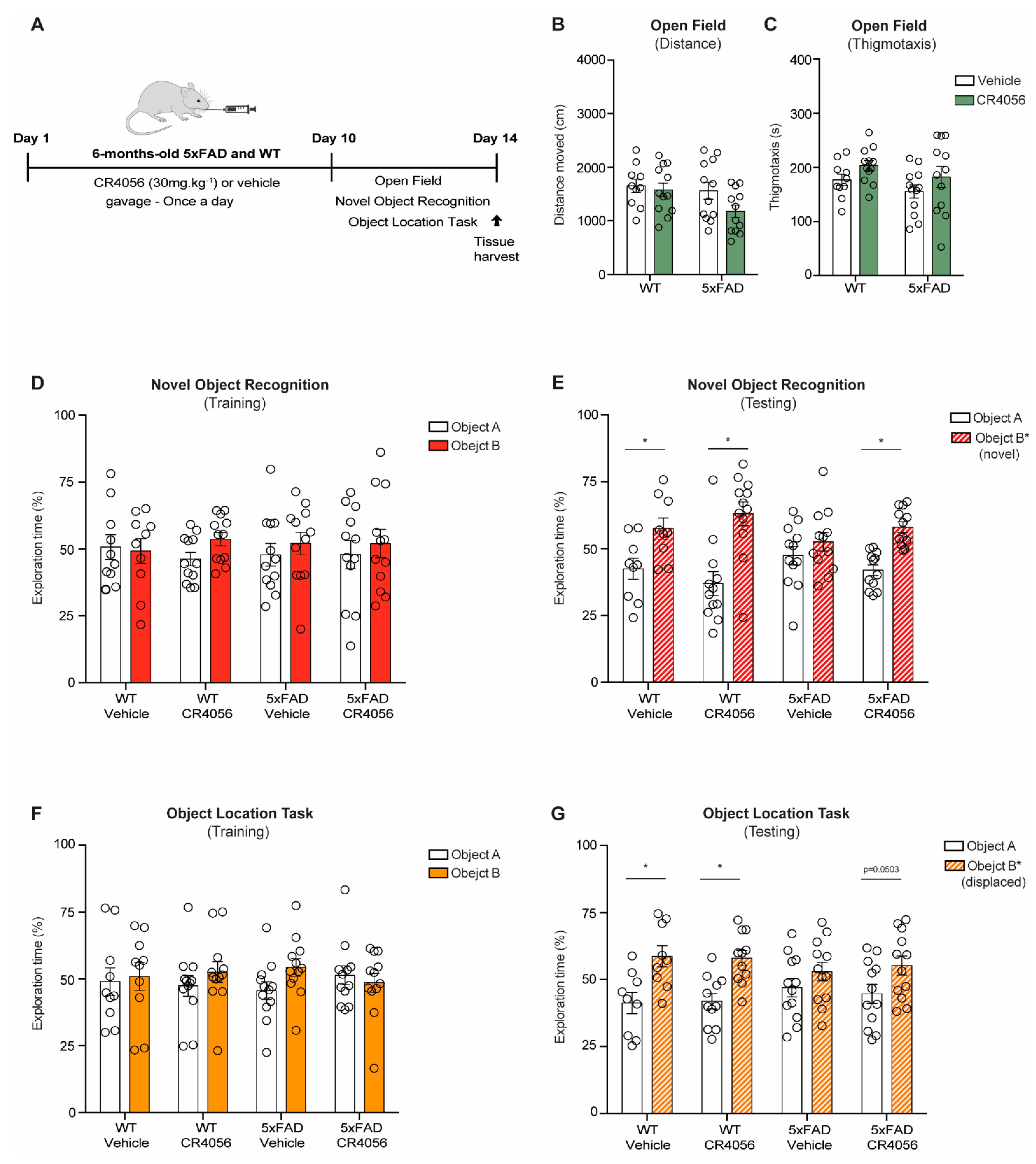
2.1. CR4056 Reverses Memory Deficits in the 5xFAD Model of AD
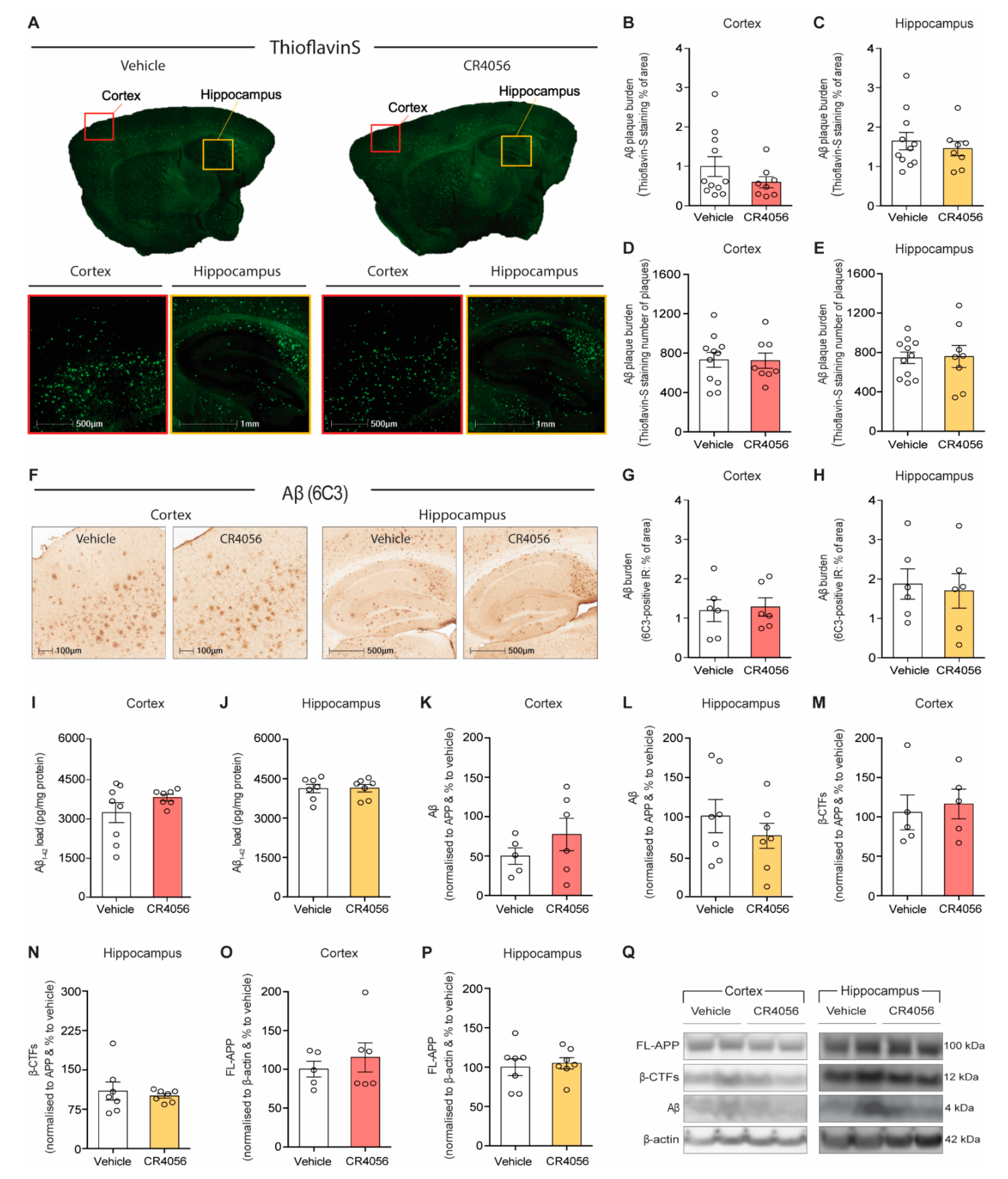
2.2. Sub-Chronic Administration of CR4056 Does Not Affect Aβ Deposition
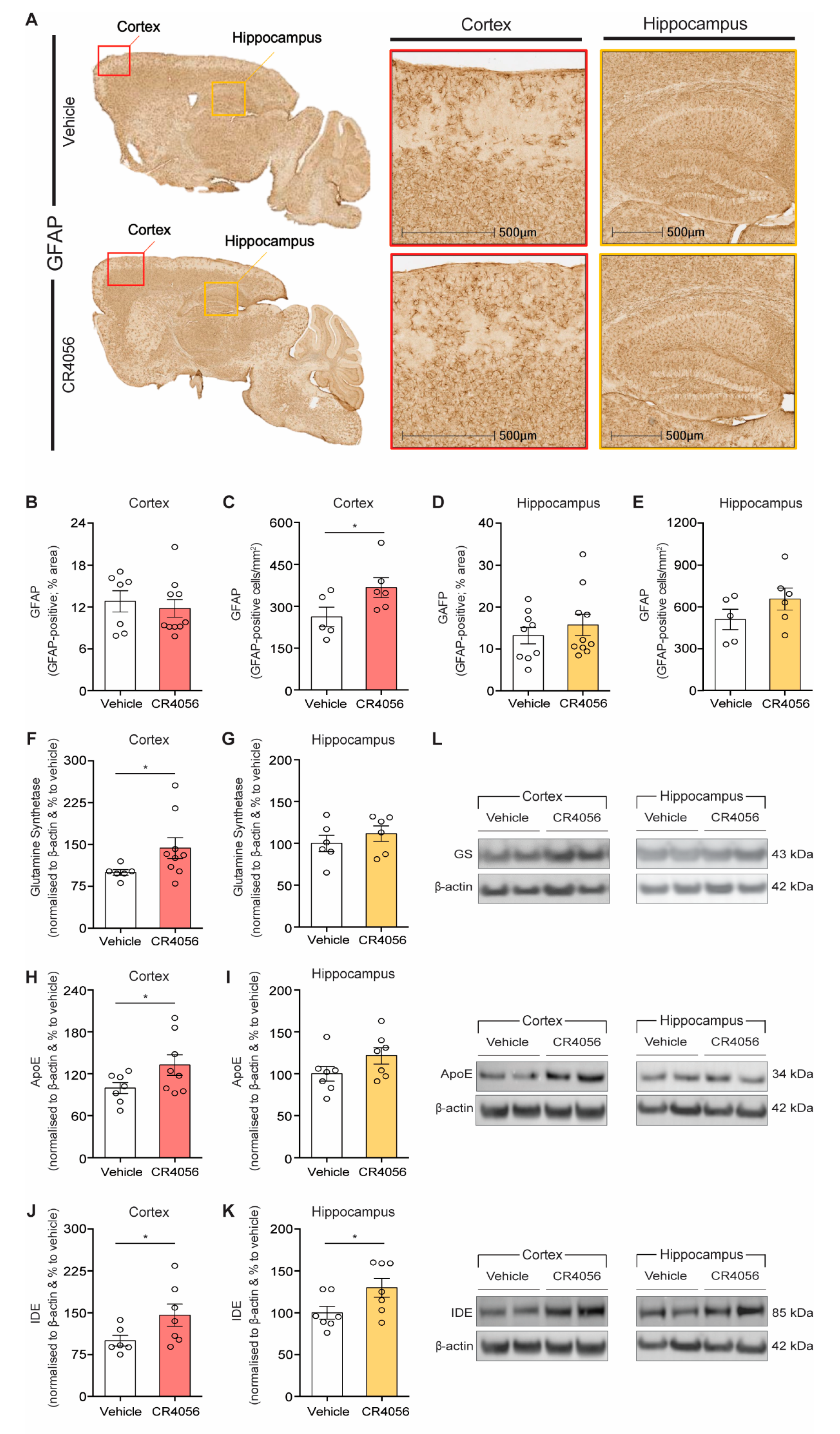
2.3. Effect of CR4056 on Astrocyte Density
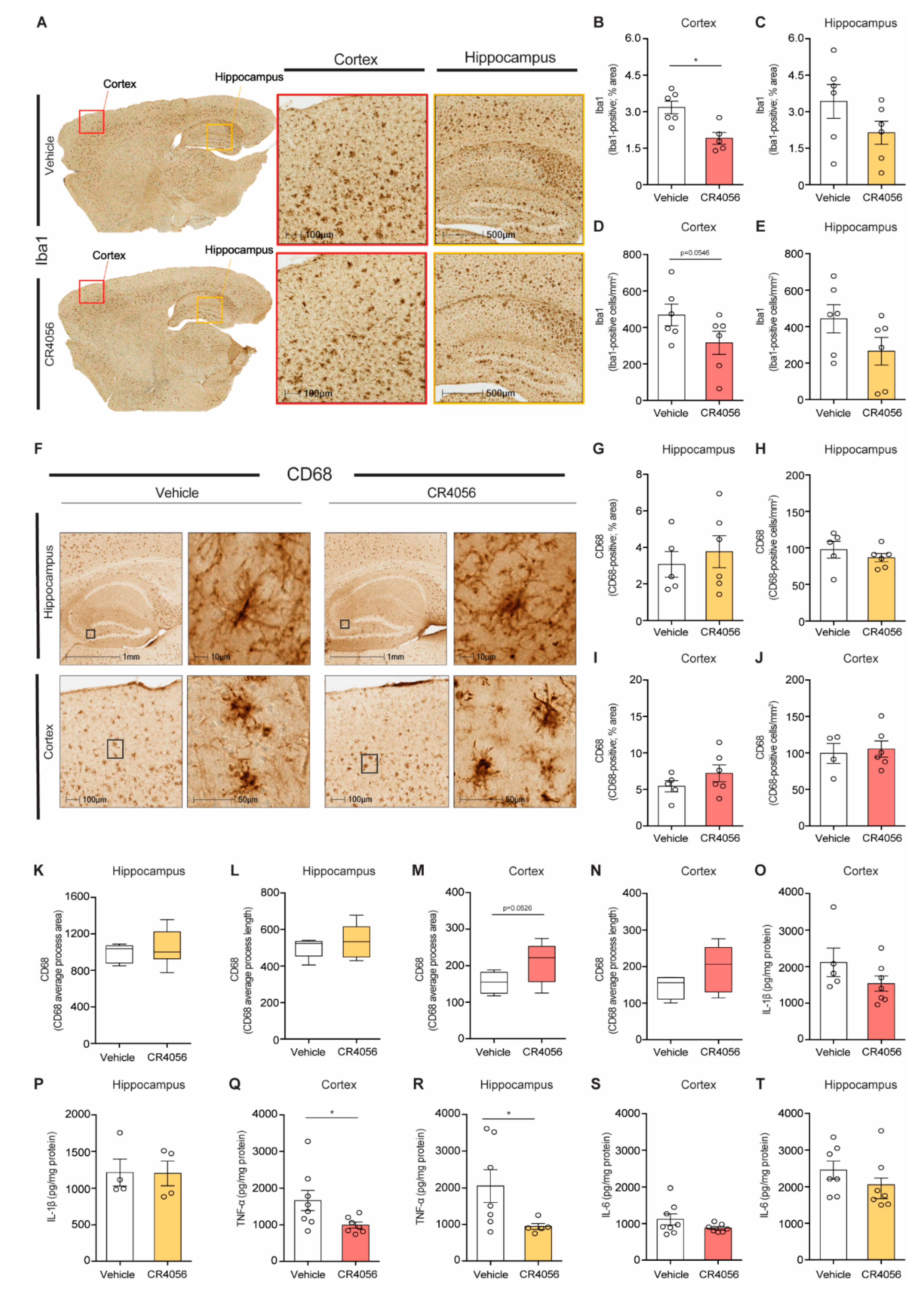
2.4. Anti-Inflammatory Effect of CR4056 in 5xFAD Mice
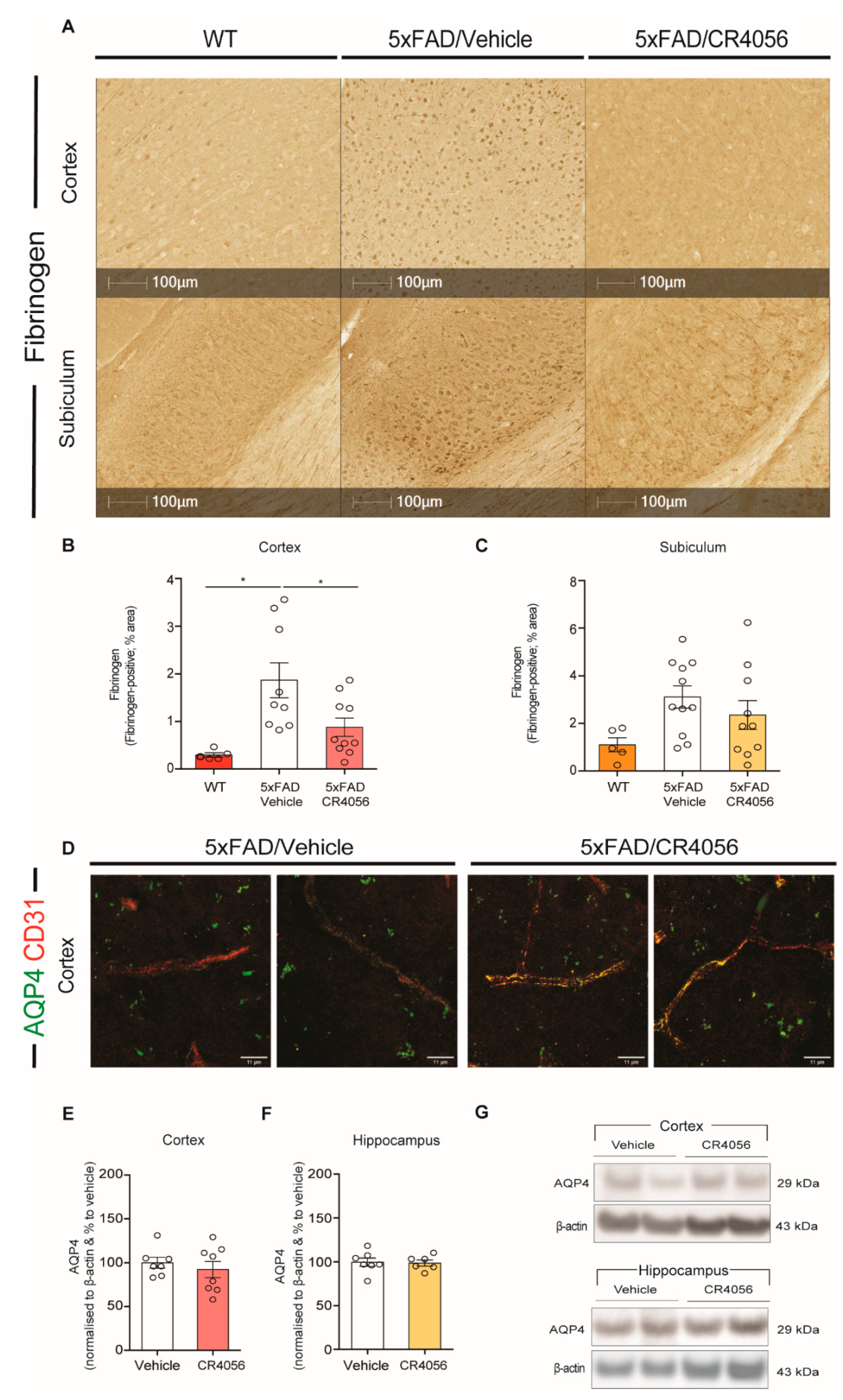
2.5. CR4056 Reverses BBB Leakage in 5xFAD Mice
3. Discussion
4. Materials and Methods
4.1. Materials and Antibodies
4.2. Animals and Treatment
4.3. Open Field (OF)
4.4. Object-Location Test (OLT)
4.5. Novel Object-Recognition Test (NOR)
4.6. Western Blotting
4.7. Immunohistochemistry
4.8. Thioflavin-S Staining
4.9. ELISA
4.10. Analysis of Images
4.11. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Weller, J.; Budson, A. Current understanding of Alzheimer’s disease diagnosis and treatment. F1000Res. Fac. Rev. 2018, 7, F1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, E.B.; Shadlen, M.F.; Wang, L.; McCormick, W.C.; Bowen, J.D.; Teri, L.; Kukull, W.A. Survival after initial diagnosis of Alzheimer disease. Ann. Intern Med. 2004, 140, 501–509. [Google Scholar] [CrossRef]
- Yiannopoulou, K.G.; Papageorgiou, S.G. Current and Future Treatments in Alzheimer Disease: An Update. J Cent. Nerv. Syst. Dis. 2020, 12, 1179573520907397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folch, J.; Busquets, O.; Ettcheto, M.; Sánchez-López, E. Memantine for the Treatment of Dementia: A Review on its Current and Future Applications. J. Alzheimers Dis. 2018, 62, 1223–1240. [Google Scholar] [CrossRef] [Green Version]
- Glinz, D.; Gloy, V.L.; Monsch, A.U.; Kressig, R.W. Acetylcholinesterase inhibitors combined with memantine for moderate to severe Alzheimer’s disease: A meta-analysis. Swiss Med. Wkly. 2019, 149, w20093. [Google Scholar] [CrossRef] [Green Version]
- Knopman, D.S.; Jones, D.T.; Greicius, M.D. Failure to demonstrate efficacy of aducanumab: An analysis of the EMERGE and ENGAGE trials as reported by Biogen, December 2019. Alzheimer Dement. 2021, 17, 696–701. [Google Scholar] [CrossRef]
- Lovestone, S.; Boada, M.; Dubois, B.; Hull, M.; Rinne, J.O.; Huppertz, H.J.; Calero, M.; Andres, M.V.; Gomez-Carrillo, B.; Leon, T.; et al. A phase II trial of tideglusib in Alzheimer’s disease. J. Alzheimers Dis. 2015, 45, 75–88. [Google Scholar] [CrossRef]
- Doody, R.S.; Raman, R.; Farlow, M.; Iwatsubo, T. Semagacestat Study Group. A phase 3 trial of semagacestat for treatment of Alzheimer’s disease. N. Engl. J. Med. 2013, 369, 341–350. [Google Scholar] [CrossRef]
- Kumar, D.; Ganeshpurkar, A.; Kumar, D.; Modi, G.; Gupta, S.K.; Singh, S.K. Secretase inhibitors for the treatment of Alzheimer’s disease: Long road ahead. Eur. J. Med. Chem. 2018, 148, 436–452. [Google Scholar] [CrossRef]
- Sastre, M.; Klockgether, T.; Heneka, M.T. Contribution of inflammatory processes to Alzheimer’s disease: Molecular mechanisms. Int. J. Dev. Neurosci. 2006, 24, 167–176. [Google Scholar] [CrossRef]
- Sastre, M.; Richardson, J.C.; Gentleman, S.M.; Brooks, D.J. Inflammatory risk factors and pathologies associated with Alzheimer’s disease. Curr. Alzheimer Res. 2011, 8, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Solito, E.; Sastre, M. Microglia function in Alzheimer’s disease. Front. Pharmacol. 2012, 3, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ries, M.; Watts, H.; Mota, B.C.; Lopez, M.Y.; Donat, C.K.; Baxan, N.; Pickering, J.A.; Chau, T.W.; Semmler, A. Annexin A1 restores cerebrovascular integrity concomitant with reduced amyloid-β and tau pathology. Brain 2021, 144, 1526–1541. [Google Scholar] [CrossRef]
- Baik, S.H.; Kang, S.; Son, S.M.; Mook-Jung, I. Microglia contributes to plaque growth by cell death due to uptake of amyloid in the brain of Alzheimer’s disease mouse model. Glia 2016, 64, 2274–2290. [Google Scholar] [CrossRef]
- Bolmont, T.; Haiss, F.; Eicke, D.; Radde, R.; Mathis, C.A. Dynamics of the microglial/amyloid interaction indicate a role in plaque maintenance. J. Neurosci. 2008, 28, 4283–4292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ries, M.; Sastre, M. Mechanisms of Aβ Clearance and Degradation by Glial Cells. Front. Aging Neurosci. 2016, 8, 160. [Google Scholar] [CrossRef] [Green Version]
- Hickman, S.; Allison, E.; Khoury, J. Microglial Dysfunction and Defective β-Amyloid Clearance Pathways in Aging Alzheimer’s Disease Mice. J. Neurosci. 2008, 28, 8354–8360. [Google Scholar] [CrossRef]
- Sastre, M.; Dewachter, I.; Landreth, G.E.; Willson, T.M.; Klockgether, T.; van Leuven, F.; Heneka, M.T. Nonsteroidal anti-inflammatory drugs and peroxisome proliferator activated receptor-gamma agonists modulate immunostimulated processing of amyloid precursor protein through regulation of beta-secretase. J. Neurosci. 2003, 23, 9796–9804. [Google Scholar] [CrossRef] [Green Version]
- Sastre, M.; Dewachter, I.; Rossner, S.; Bogdanovic, N.; Rosen, E.; Borghgraef, P. Nonsteroidal anti-inflammatory drugs repress beta-secretase gene promoter activity by the activation of PPARgamma. Proc. Natl. Acad. Sci. USA 2006, 103, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Grammas, P.; Ovase, R. Inflammatory factors are elevated in brain microvessels in Alzheimer’s disease. Neurobiol. Aging 2001, 22, 837–842. [Google Scholar] [CrossRef]
- Erickson, M.A.; Dohi, K.; Banks, W.A. Neuroinflammation: A common pathway in CNS diseases as mediated at the blood-brain barrier. Neuroimmunomodulation 2012, 19, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrano, A.; Hoozemans, J.J.M.; van der Vies, S.M.; van Horssen, J.; de Vries, H.E.; Rozemuller, A.J.M. Neuroinflammation and blood-brain barrier changes in capillary amyloid angiopathy. Neurodegener. Dis. 2012, 10, 329–331. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, P.; Hudson, A.; Garcia-Sevilla, J.A.; Li, J.-X. Imidazoline receptor system: The past, the present, and the future. Pharmacol. Rev. 2020, 72, 50–79. [Google Scholar] [CrossRef]
- Regunathan, S.; Feinstein, D.L.; Reis, D.J. Anti-proliferative and anti-inflammatory actions of imidazoline agents. Are imidazoline receptors involved? Ann. N. Y. Acad. Sci. 1999, 881, 410–419. [Google Scholar] [CrossRef]
- Ruiz, J.; Martin, I.; Callado, L.F.; Meana, J.J.; Barturen, F.; Garcia-Sevilla, J.A. Nonadrenoceptor [3H]idazoxan binding sites (12-imidazoline sites) are increased in post-mortem brain from patients with Alzheimer’s disease. Neurosci. Lett. 1993, 160, 109–112. [Google Scholar] [CrossRef]
- Calsolaro, V.; Matthews, P.M.; Donat, C.K.; Livingston, N.R.; Femminella, G.D. Astrocyte reactivity with late-onset cognitive impairment assessed in vivo using 11 C-BU99008 PET and its relationship with amyloid load. Mol. Psychiatr. 2021, 26, 5848–5855. [Google Scholar] [CrossRef] [PubMed]
- Regunathan, S.; Feinstein, D.L.; Reis, D.J. Expression of non-adrenergic imidazoline sites in rat cerebral cortical astrocytes. J. Neurosci Res. 1993, 34, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Regunathan, S.; Meeley, M.P.; Reis, D.J. Expression of non-adrenergic imidazoline sites in chromaffin cells and mitochondrial membranes of bovine adrenal medulla. Biochem. Pharmacol. 1993, 45, 1667–1675. [Google Scholar] [CrossRef]
- Tesson, F.; Prip-Buus, C.; Lemoine, A.; Pegorier, J.P.; Parini, A. Subcellular distribution of imidazoline-guanidinium-receptive sites in human and rabbit liver. Major localization to the mitochondrial outer membrane. J. Biol. Chem. 1991, 266, 155–160. [Google Scholar] [CrossRef]
- Olmos, G.; Alemany, R.; Escribá, P.V.; García-Sevilla, J.A. The effects of chronic imidazoline drug treatment on glial fibrillary acidic protein concentrations in rat brain. Br. J. Pharmacol. 1994, 111, 997–1002. [Google Scholar] [CrossRef] [Green Version]
- Lanza, M.; Ferrari, F.; Menghetti, I.; Tremolada, D.; Caselli, G. Modulation of imidazoline I2 binding sites by CR4056 relieves postoperative hyperalgesia in male and female rats. Br. J. Pharmacol. 2014, 171, 3693–3701. [Google Scholar] [CrossRef] [PubMed]
- Hudson, A.L.; Gough, R.; Tyacke, R.; Lione, L.; Lalies, M.; Lewis, J.; Husbands, S.; Knight, P.; Murray, F.; Hutson, P.; et al. Novel selective compounds for the investigation of imidazoline receptors. Ann. N. Y. Acad. Sci. 1999, 881, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, F.; Fiorentino, S.; Mennuni, L. Analgesic efficacy of CR4056, a novel imidazoline-2 receptor ligand, in rat models of inflammatory and neuropathic pain. J. Pain Res. 2011, 4, 111–125. [Google Scholar] [PubMed] [Green Version]
- Vucicevic, J.; Nikolic, K.; Dobricic, V.; Agbaba, D. Prediction of blood-brain barrier permeation of a-adrenergic and imidazoline receptor ligands using PAMPA technique and quantitative-structure permeability relationship analysis. Eur. J. Pharm. Sci. 2015, 68, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Vellani, V.; Sabatini, C.; Milia, C.; Caselli, G.; Lanza, M.; Letari, O.; Rovati, L.C.; Giacomoni, C. CR4056, a powerful analgesic imidazoline-2 receptor ligand, inhibits the inflammation-induced PKCε phosphorylation and membrane translocation in sensory neurons. Br. J. Pharmacol. 2020, 177, 48–64. [Google Scholar] [CrossRef]
- Rovati, L.C.; Brambilla, N.; Blicharski, T.; Connell, J.; Vitalini, C.; Bonazzi, A.; Giacovelli, G.; Girolami, F.; D’Amato, M. Efficacy and safety of the first-in-class imidazoline-2 receptor ligand CR4056 in pain from knee osteoarthritis and disease phenotypes: A randomized, double-blind, placebo-controlled phase 2 trial. Osteoarthr. Cartil. 2020, 28, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Mirzaei, N.; Mota, B.C.; Birch, A.M.; Davis, N.; Romero-Molina, C.; Katsouri, L.; Palmer, E.O.C.; Golbano, A.; Riggall, L.I.; Nagy, I.; et al. lmidazoline ligand BU224 reverses cognitive deficits, reduces microgliosis and enhances synaptic connectivity in a mouse model of Alzheimer’s disease. Br. J. Pharmacol. 2021, 178, 654–671. [Google Scholar] [CrossRef]
- Abás, S.; Rodríguez-Arévalo, S.; Bagán, A.; Griñán-Ferré, C.; Vasilopoulou, F. Bicyclic α-Iminophosphonates as High Affinity Imidazoline I2 Receptor Ligands for Alzheimer’s Disease. J. Med. Chem. 2020, 63, 3610–3633. [Google Scholar] [CrossRef]
- Vasilopoulou, F.; Rodríguez-Arévalo, S.; Bagán, A.; Escolano, C.; Griñán-Ferré, C.; Pallàs, M. Disease-modifying treatment with I2 imidazoline receptor ligand LSL60101 in an Alzheimer’s disease mouse model: A comparative study with donepezil. Br. J. Pharmacol. 2021, 178, 3017–3033. [Google Scholar] [CrossRef]
- Sala, E.; Ferrari, F.; Lanza, M.; Milia, C.; Sabatini, C.; Bonazzi, A.; Comi, E.; Borsi Franchini, M.; Caselli, G.; Rovati, L.C. Improved efficacy, tolerance, safety, and abuse liability profile of the combination of CR4056 and morphine over morphine alone in rodent models. Br. J. Pharmacol. 2020, 177, 3291–3308. [Google Scholar] [CrossRef]
- Alemany, R.; Olmos, G.; Escriba, P.V.; Menargues, A.; Obach, R.; Garcia-Sevilla, J.A. LSL 60101,a selective ligand for imidazoline 12 receptors, on glia l fibrillary acidic protein concentration. Eur. J. Pharmacol. 1995, 280, 205–210. [Google Scholar] [CrossRef]
- Igarashi, H.; Suzuki, Y.; Kwee, I.L.; Nakada, T. Water influx into cerebrospinal fluid is significantly reduced in senile plaque bearing transgenic mice, supporting beta-amyloid clearance hypothesis of Alzheimer’s disease. Neurol. Res. 2014, 36, 1094–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixit, M.; Rahmatkar, S.; Raut, P. Evidences for agmatine alterations in Aβ-42 induced memory impairment in mice. Neurosci Lett. 2021, 740, 135447. [Google Scholar] [CrossRef] [PubMed]
- Griñán-Ferré, C.; Vasilopoulou, F.; Abás, S.; Rodríguez-Arévalo, S.; Bagán, A. Behavioral and Cognitive Improvement Induced by Novel Imidazoline I2 Receptor Ligands in Female SAMP8 Mice. Neurotherapeutics 2019, 16, 416–431. [Google Scholar] [CrossRef] [Green Version]
- Tundo, G.R.; Sbardella, D.; Ciaccio, C.; Grasso, G.; Gioia, M.; Coletta, A.; Polticelli, F. Multiple functions of insulin-degrading enzyme: A metabolic crosslight? Crit. Rev. Biochem. Mol. Biol. 2017, 52, 554–582. [Google Scholar] [CrossRef]
- Vasilopoulou, F.; Bagan, A.; Rodriguez-Arevalo, S.; Escolano, C.; Griñán-Ferré, C.; Pallàs, M. Amelioration of BPSD-Like Phenotype and Cognitive Decline in SAMP8 Mice Model Accompanied by Molecular Changes after Treatment with I 2-Imidazoline Receptor Ligand MCR5. Pharmaceutics 2020, 12, 475. [Google Scholar] [CrossRef]
- Vasilopoulou, F.; Escolano, C.; Pallàs, M.; Griñán-Ferré, C. Microarray Analysis Revealed Inflammatory Transcriptomic Changes after LSL60101 Treatment in 5xFAD Mice Model. Genes 2021, 12, 1315. [Google Scholar] [CrossRef]
- Katsouri, L.; Birch, A.M.; Renziehausen, A.W.J.; Zach, C.; Aman, Y.; Steeds, H.; Bonsu, A.; Palmer, E.O.C.; Mirzaei, N.; Ries, M.; et al. Ablation of reactive astrocytes exacerbates disease pathology in a model of Alzheimer’s disease. Glia 2020, 68, 1017–1030. [Google Scholar] [CrossRef] [Green Version]
- Davis, N.; Mota, B.C.; Stead, L.; Palmer, E.O.C.; Lombardero, L.; Rodríguez-Puertas, R.; de Paola, V.; Barnes, S.J.; Sastre, M. Pharmacological ablation of astrocytes reduces A degradation and synaptic connectivity in an ex vivo model of Alzheimer’s disease. J. Neuroinf. 2021, 18, 73. [Google Scholar] [CrossRef]
- Long, J.M.; Holtzman, D.M. Alzheimer Disease: An Update on Pathobiology and Treatment Strategies. Cell 2019, 179, 312–339. [Google Scholar] [CrossRef]
- Bousquet, P.; Greney, H.; Bruban, V.; Schann, S.; Ehrhardt, J.D.; Monassier, L.; Feldman, J. 11-imidazoline receptors involved in cardiovascular regulation: Where are we and where are we going? Ann. N. Y. Acad. Sci. 2003, 1009, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Cheng, Z.H.; Liu, S.; Yang, J.L.; Xiao, M.J.; Zheng, R.Y.; Hou, S.T. Neurovascular protection conferred by 2-BFI treatment during rat cerebral ischemia. Biochem. Biophys. Res. Com. 2012, 424, 544–548. [Google Scholar] [CrossRef] [PubMed]
- Regunathan, S.; Youngson, C.; Wang, H.; Reis, D.J. Imidazoline receptors in vascular smooth muscle and endothelial cells. Ann. N. Y. Acad. Sci. 1995, 763, 580–590. [Google Scholar] [CrossRef]
- Oakley, H.; Cole, S.L.; Logan, S.; Maus, E.; Shao, P.; Craft, J. Intraneuronal beta-amyloid aggregates, neurodegeneration, and neuron loss in transgenic mice with five familial Alzheimer’s disease mutations: Potential factors in amyloid plaque formation. J. Neurosci. 2006, 26, 10129–10140. [Google Scholar] [CrossRef]
- Meregalli, C.; Ceresa, C.; Canta, A.; Carozzi, V.A.; Chiorazzi, A.; Sala, B.; Oggioni, N.; Lanza, M.; Letari, O.; Ferrari, F.; et al. CR4056, a new analgesic I2 ligand, is highly effective against bortezomib-induced painful neuropathy in rats. J. Pain. Res. 2012, 5, 151–167. [Google Scholar] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mota, B.C.; Ashburner, N.; Abelleira-Hervas, L.; Liu, L.; Aleksynas, R.; Rovati, L.C.; Caselli, G.; Sastre, M. I2-Imidazoline Ligand CR4056 Improves Memory, Increases ApoE Expression and Reduces BBB Leakage in 5xFAD Mice. Int. J. Mol. Sci. 2022, 23, 7320. https://doi.org/10.3390/ijms23137320
Mota BC, Ashburner N, Abelleira-Hervas L, Liu L, Aleksynas R, Rovati LC, Caselli G, Sastre M. I2-Imidazoline Ligand CR4056 Improves Memory, Increases ApoE Expression and Reduces BBB Leakage in 5xFAD Mice. International Journal of Molecular Sciences. 2022; 23(13):7320. https://doi.org/10.3390/ijms23137320
Chicago/Turabian StyleMota, Bibiana C., Nathan Ashburner, Laura Abelleira-Hervas, Liyueyue Liu, Robertas Aleksynas, Lucio Claudio Rovati, Gianfranco Caselli, and Magdalena Sastre. 2022. "I2-Imidazoline Ligand CR4056 Improves Memory, Increases ApoE Expression and Reduces BBB Leakage in 5xFAD Mice" International Journal of Molecular Sciences 23, no. 13: 7320. https://doi.org/10.3390/ijms23137320
APA StyleMota, B. C., Ashburner, N., Abelleira-Hervas, L., Liu, L., Aleksynas, R., Rovati, L. C., Caselli, G., & Sastre, M. (2022). I2-Imidazoline Ligand CR4056 Improves Memory, Increases ApoE Expression and Reduces BBB Leakage in 5xFAD Mice. International Journal of Molecular Sciences, 23(13), 7320. https://doi.org/10.3390/ijms23137320