
Ligation of TLR Homologue CD180 of B Cells Activates the PI3K/Akt/mTOR Pathway in Systemic Sclerosis and Induces a Pathological Shift in the Expression of BAFF Receptors

 ,
,
Abstract
:1. Introduction
2. Results
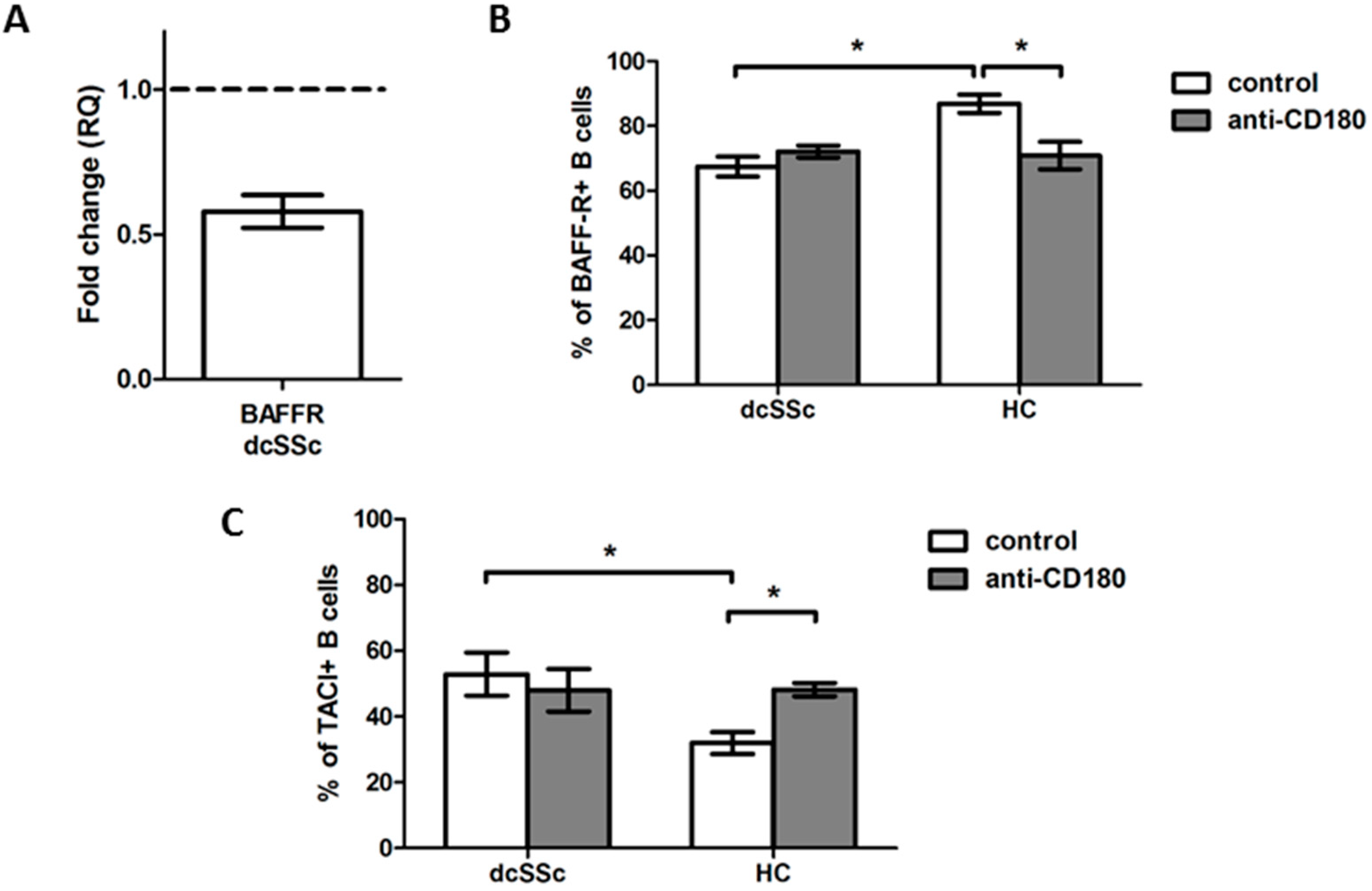
2.1. Stimulation via CD180 Induces the Phosphorylation of Akt to a Lower Extent in dcSSc B Cells
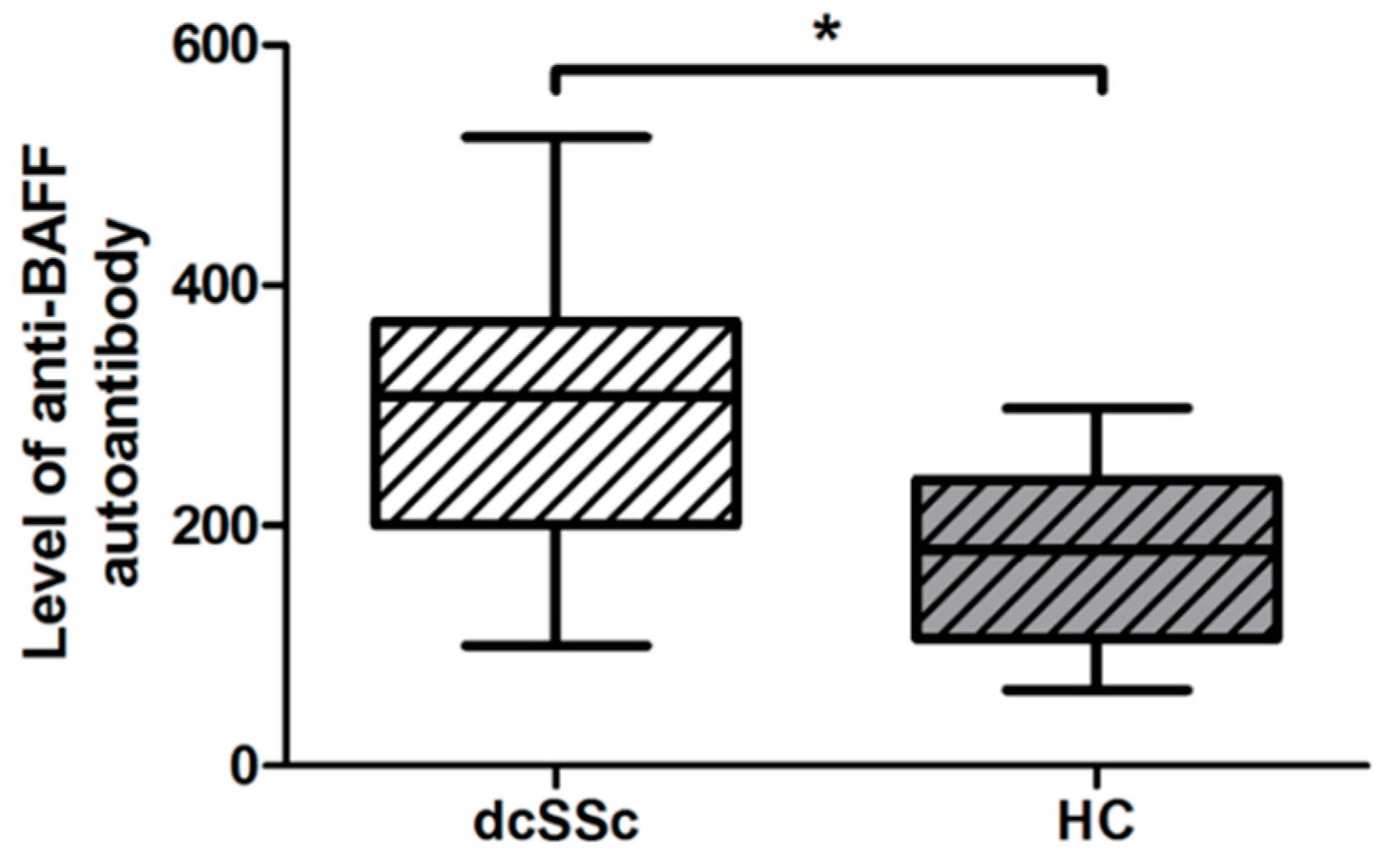
2.2. Anti-BAFF Autoantibody Serum Level Is Higher in dcSSc
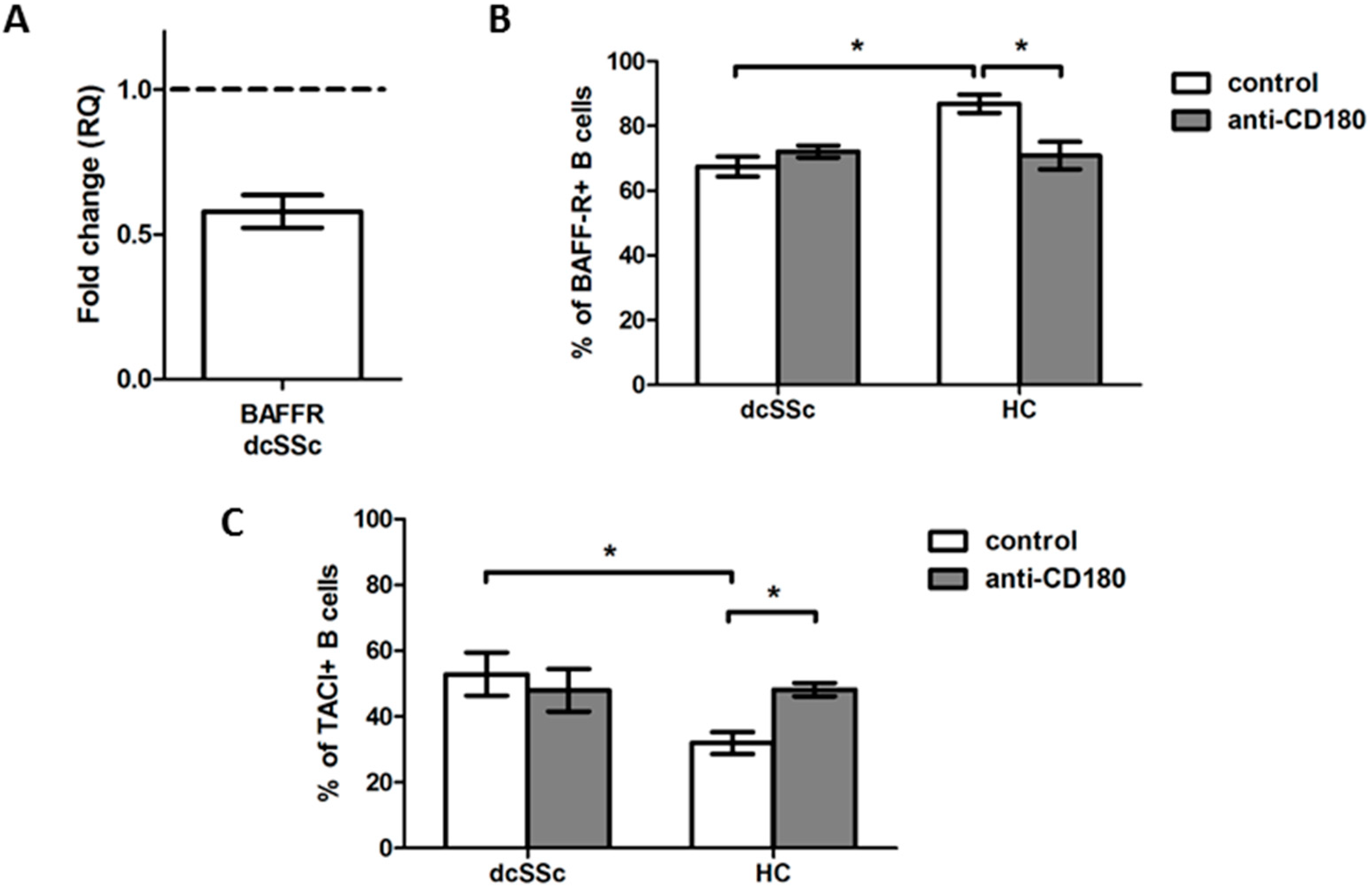
2.3. Decreased Basal Expression of BAFF-R Is Accompanied by Elevated Expression of TACI in dcSSc B Cells
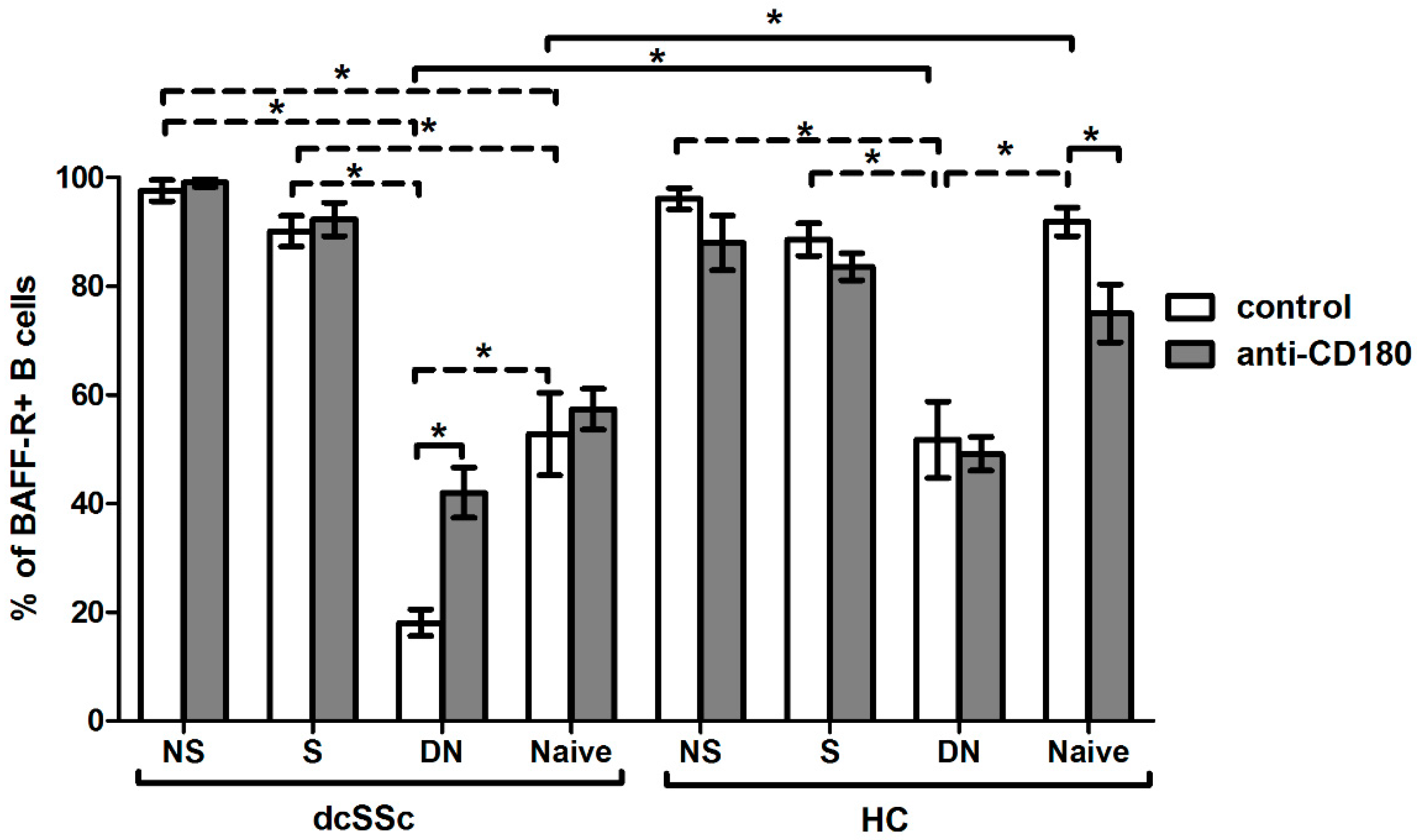
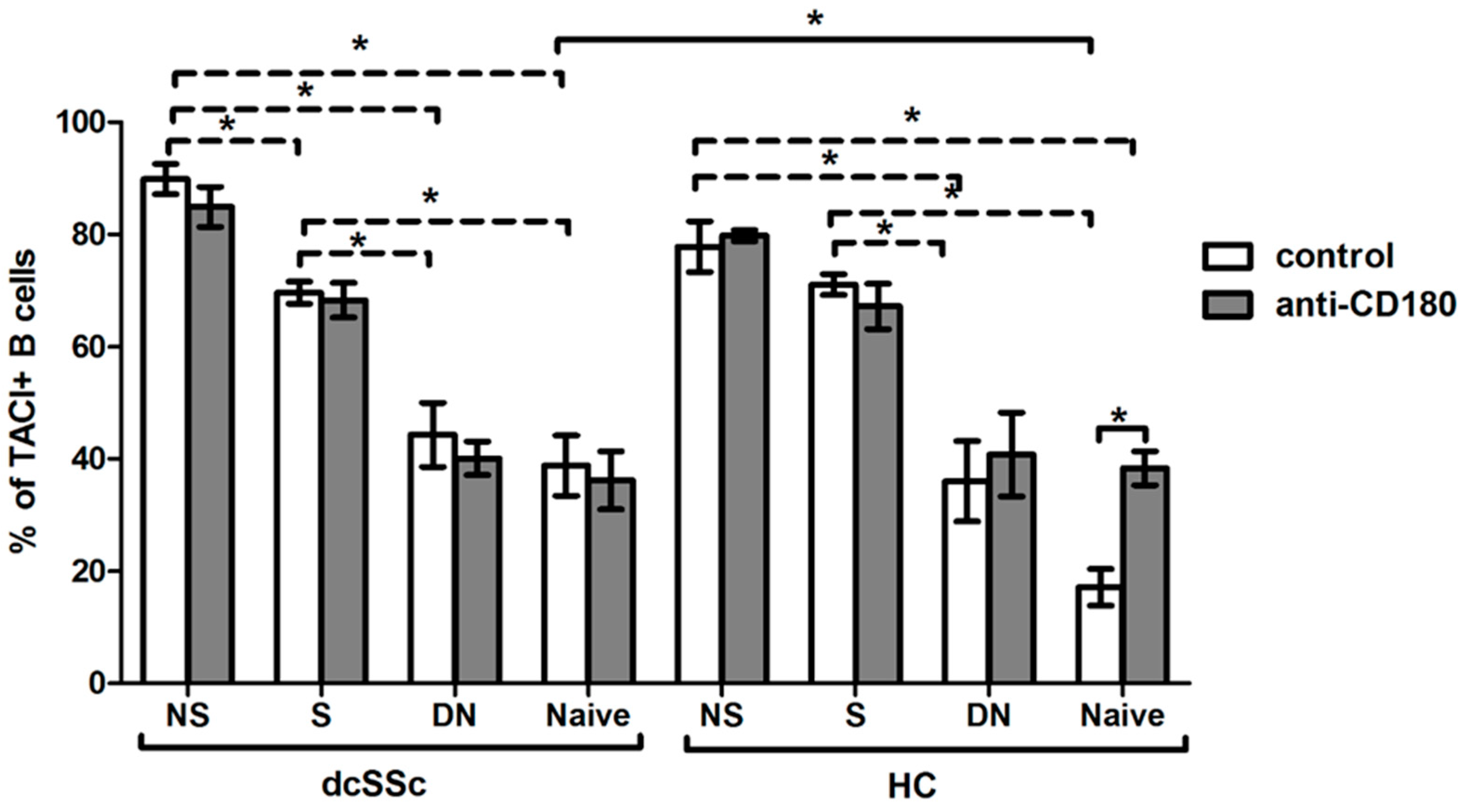
2.4. Basal Expression of BAFF-R Is Higher in HC Naive B Cells and Is Reduced by Anti-CD180 Antibody Treatment to the Level of dcSSc Naive B Cells
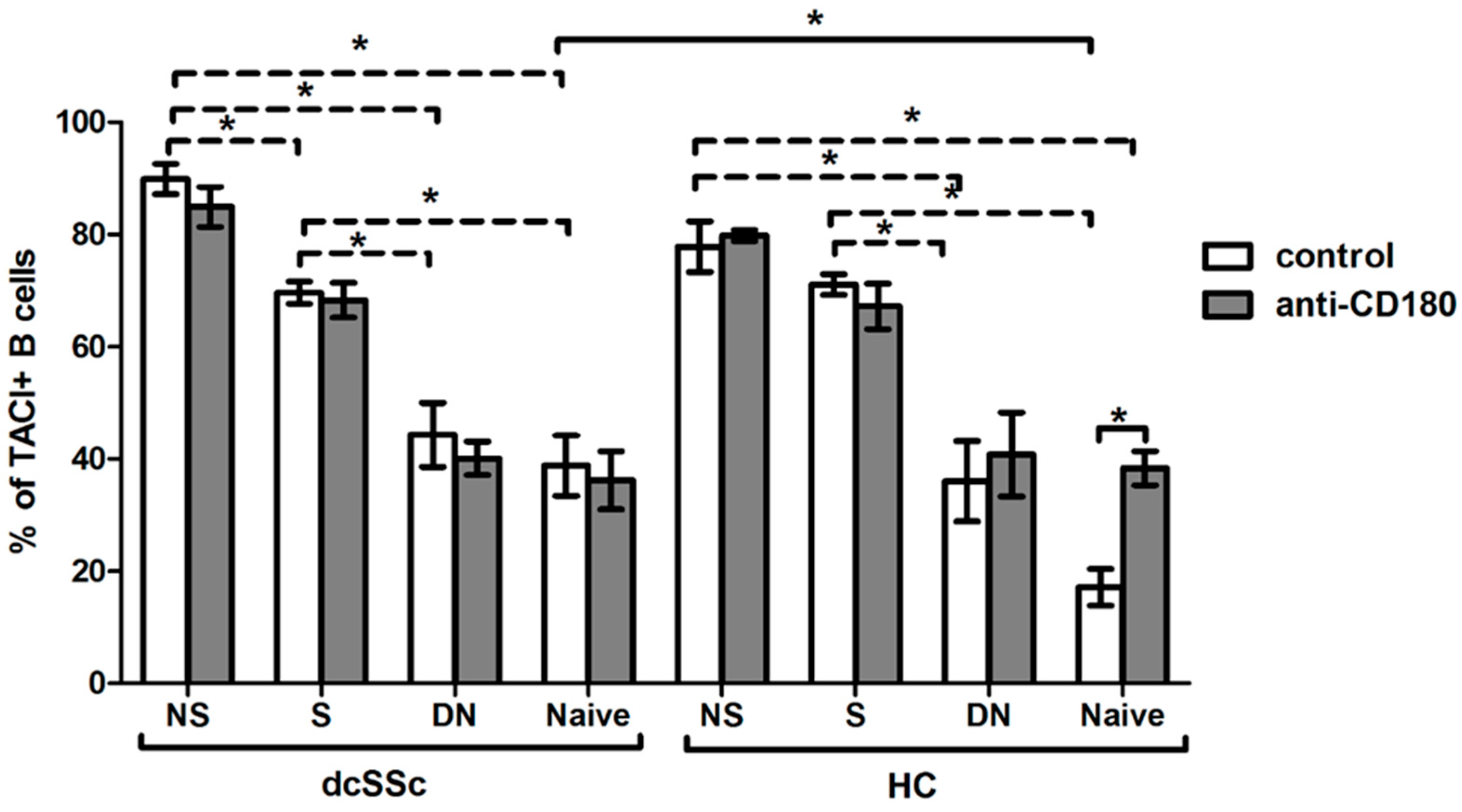
2.5. Basal Expression of TACI Is Lower in HC Naive B Cells, and Is Increased with Ligation of CD180 to the Level of dcSSc Naive B Cells
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Peripheral Blood Mononuclear Cell Isolation and B Cell Separation
4.3. Anti-CD180 Antibody Stimulation
4.4. RNA Isolation, cDNA Synthesis and qPCR for the Evaluation of BAFF-R Expression
4.5. Flow Cytometric Analysis of BAFF-R and TACI Expression
4.6. Assessment of the Phosphorylation of Akt and S6
4.7. Anti-BAFF Autoantibody Measurement
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liang, M.; Lv, J.; Chu, H.; Wang, J.; Chen, X.; Zhu, X.; Xue, Y.; Guan, M.; Zou, H. Vertical inhibition of PI3K/Akt/mTOR signaling demonstrates in vitro and in vivo anti-fibrotic activity. J. Dermatol. Sci. 2014, 76, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Limon, J.J.; Fruman, D.A. Akt and mTOR in B cell activation and differentiation. Front. Immunol. 2012, 3, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varga, J.; Trojanowska, M.; Kuwana, M. Pathogenesis of systemic sclerosis: Recent insights of molecular and cellular mechanisms and therapeutic opportunities. J. Scleroderma Relat. Disord. 2017, 2, 137–152. [Google Scholar] [CrossRef]
- Porta, C.; Paglino, C.; Mosca, A. Targeting PI3K/Akt/mTOR signaling in cancer. Front. Oncol. 2014, 4, 64. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Serada, S.; Fujimoto, M.; Terao, M.; Kotobuki, Y.; Kitaba, S.; Matsui, S.; Kudo, A.; Naka, T.; Murota, H.; et al. Periostin facilitates skin sclerosis via PI3K/Akt dependent mechanism in a mouse model of scleroderma. PLoS ONE 2012, 7, e41994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perl, A. Activation of mTOR (mechanistic target of rapamycin) in rheumatic diseases. Nat. Rev. Rheumatol. 2016, 12, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Pauls, S.D.; Lafarge, S.T.; Landego, I.; Zhang, T.; Marshall, A.J. The phosphoinositide 3-kinase signaling pathway in normal and malignant B cells: Activation mechanisms, regulation, and impact on cellular functions. Front. Immunol. 2012, 3, 224. [Google Scholar] [CrossRef] [Green Version]
- Ge, F.; Wang, F.; Yan, X.; Li, Z.; Wang, X. Association of BAFF with PI3K/Akt/mTOR signaling in lupus nephritis. Mol. Med. Rep. 2017, 16, 5793. [Google Scholar] [CrossRef] [Green Version]
- MacKay, F.; Schneider, P. Cracking the BAFF code. Nat. Rev. Immunol. 2009, 9, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, T.; Hasegawa, M.; Yanaba, K.; Kodera, M.; Takehara, K.; Sato, S. Elevated serum BAFF levels in patients with systemic sclerosis: Enhanced BAFF signaling in systemic sclerosis B lymphocytes. Arthritis Rheum. 2006, 54, 192–201. [Google Scholar] [CrossRef]
- Vincent, T.; Plawecki, M.; Goulabchand, R.; Guilpain, P.; Eliaou, J.F. Emerging clinical phenotypes associated with anti-cytokine autoantibodies. Autoimmun. Rev. 2015, 14, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Price, J.V.; Haddon, D.J.; Kemmer, D.; Delepine, G.; Mandelbaum, G.; Jarrell, J.A.; Gupta, R.; Balboni, I.; Chakravarty, E.F.; Sokolove, J.; et al. Protein microarray analysis reveals BAFF-binding autoantibodies in systemic lupus erythematosus. J. Clin. Investig. 2013, 123, 5135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieper, K.; Grimbacher, B.; Eibel, H. B-cell biology and development. J. Allergy Clin. Immunol. 2013, 131, 959–971. [Google Scholar] [CrossRef]
- Mackay, F.; Schneider, P. TACI, an enigmatic BAFF/APRIL receptor, with new unappreciated biochemical and biological properties. Cytokine Growth Factor Rev. 2008, 19, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.; Zhang, Y.M.; Zhang, X.M.; Tao, J. Effect of TACI signaling on humoral immunity and autoimmune diseases. J. Immunol. Res. 2015, 2015, 247426. [Google Scholar] [CrossRef]
- Abu-Rish, E.Y.; Amrani, Y.; Browning, M.J. Toll-like receptor 9 activation induces expression of membrane-bound B-cell activating factor (BAFF) on human B cells and leads to increased proliferation in response to both soluble and membrane-bound BAFF. Rheumatology 2013, 52, 1190–1201. [Google Scholar] [CrossRef] [Green Version]
- Simon, D.; Erdő-Bonyár, S.; Rapp, J.; Balogh, P.; Minier, T.; Nagy, G.; Czirják, L.; Berki, T. Analysis of PI3K Pathway Associated Molecules Reveals Dysregulated Innate and Adaptive Functions of B Cells in Early Diffuse Cutaneous Systemic Sclerosis. Int. J. Mol. Sci. 2021, 22, 2877. [Google Scholar] [CrossRef]
- Chaplin, J.W.; Kasahara, S.; Clark, E.A.; Ledbetter, J.A. Anti-CD180 (RP105) Activates B Cells To Rapidly Produce Polyclonal Ig via a T Cell and MyD88-Independent Pathway. J. Immunol. 2011, 187, 4199–4209. [Google Scholar] [CrossRef] [Green Version]
- Erdő-Bonyár, S.; Rapp, J.; Minier, T.; Ráth, G.; Najbauer, J.; Czirják, L.; Németh, P.; Berki, T.; Simon, D. Toll-like receptor mediated activation of natural autoantibody producing b cell subpopulations in an autoimmune disease model. Int. J. Mol. Sci. 2019, 20, 6152. [Google Scholar] [CrossRef] [Green Version]
- Porakishvili, N.; Vispute, K.; Steele, A.J.; Rajakaruna, N.; Kulikova, N.; Tsertsvadze, T.; Nathwani, A.; Damle, R.N.; Clark, E.A.; Rai, K.R.; et al. Rewiring of sigm-mediated intracellular signaling through the CD180 toll-like receptor. Mol. Med. 2015, 21, 46–57. [Google Scholar] [CrossRef]
- Teng, Y.; Fan, Y.; Ma, J.; Lu, W.; Liu, N.; Chen, Y.; Pan, W.; Tao, X. The pi3k/akt pathway: Emerging roles in skin homeostasis and a group of non-malignant skin disorders. Cells 2021, 10, 1219. [Google Scholar] [CrossRef]
- Hay, N.; Sonenberg, N. Upstream and downstream of mTOR. Genes Dev. 2004, 18, 1926–1945. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Chu, H.; Jiang, S.; Liu, Q.; Liu, L.; Xue, Y.; Zheng, S.; Wan, W.; Qiu, J.; Wang, J.; et al. Sirt1 ameliorates systemic sclerosis by targeting the mTOR pathway. J. Dermatol. Sci. 2017, 87, 149–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, T.I.K.; Khanna, D.; Furst, D.E.; Danovitch, G.; Burger, C.; Maranian, P.; Clements, P.J. Rapamycin versus methotrexate in early diffuse systemic sclerosis: Results from a randomized, single-blind pilot study. Arthritis Rheum. 2009, 60, 3821–3830. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, S.; Tapping, R.I. Toll-like receptor signaling in cell proliferation and survival. Cytokine 2010, 49, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Porakishvili, N.; Memon, A.; Vispute, K.; Kulikova, N.; Clark, E.A.; Rai, K.R.; Nathwani, A.; Damle, R.N.; Chiorazzi, N.; Lydyard, P.M. CD180 functions in activation, survival and cycling of B chronic lymphocytic leukaemia cells. Br. J. Haematol. 2011, 153, 486–498. [Google Scholar] [CrossRef]
- Forestier, A.; Guerrier, T.; Jouvray, M.; Giovannelli, J.; Lefèvre, G.; Sobanski, V.; Hauspie, C.; Hachulla, E.; Hatron, P.Y.; Zéphir, H.; et al. Altered B lymphocyte homeostasis and functions in systemic sclerosis. Autoimmun. Rev. 2018, 17, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Li, P.P.; Liu, D.D.; Liu, Y.J.; Song, S.S.; Wang, Q.T.; Chang, Y.; Wu, Y.J.; Chen, J.Y.; Zhao, W.D.; Zhang, L.L.; et al. BAFF/BAFF-R involved in antibodies production of rats with collagen-induced arthritis via PI3K-Akt-mTOR signaling and the regulation of paeoniflorin. J. Ethnopharmacol. 2012, 141, 290–300. [Google Scholar] [CrossRef]
- Thien, M.; Phan, T.G.; Gardam, S.; Amesbury, M.; Basten, A.; MacKay, F.; Brink, R. Excess BAFF Rescues Self-Reactive B Cells from Peripheral Deletion and Allows Them to Enter Forbidden Follicular and Marginal Zone Niches. Immunity 2004, 20, 785–798. [Google Scholar] [CrossRef] [Green Version]
- Haddon, D.J.; Diep, V.K.; Price, J.V.; Limb, C.; Utz, P.J.; Balboni, I. Autoantigen microarrays reveal autoantibodies associated with proliferative nephritis and active disease in pediatric systemic lupus erythematosus. Arthritis Res. Ther. 2015, 17, 162. [Google Scholar] [CrossRef] [Green Version]
- Schiemann, B.; Gommerman, J.L.; Vora, K.; Cachero, T.G.; Shutga-Morskaya, S.; Dobles, M.; Frew, E.; Scott, M.L. An essential role for BAFF in the normal development of B cells through a BCMA-independent pathway. Science 2001, 293, 2111–2114. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Camarena, D.C.; Ortiz-Lazareno, P.C.; Cruz, A.; Oregon-Romero, E.; Machado-Contreras, J.R.; Muñoz-Valle, J.F.; Orozco-López, M.; Marín-Rosales, M.; Palafox-Sánchez, C.A. Association of BAFF, APRIL serum levels, BAFF-R, TACI and BCMA expression on peripheral B-cell subsets with clinical manifestations in systemic lupus erythematosus. Lupus 2016, 25, 582–592. [Google Scholar] [CrossRef]
- Zhao, L.D.; Li, Y.; Smith, M.F.; Wang, J.S.; Zhang, W.; Tang, F.L.; Tian, X.P.; Wang, H.Y.; Zhang, F.C.; Ba, D.N.; et al. Expressions of BAFF/BAFF receptors and their correlation with disease activity in Chinese SLE patients. Lupus 2010, 19, 1534–1549. [Google Scholar] [CrossRef]
- Darce, J.R.; Arendt, B.K.; Wu, X.; Jelinek, D.F. Regulated Expression of BAFF-Binding Receptors during Human B Cell Differentiation. J. Immunol. 2007, 179, 7276–7286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senécal, J.-L.; Hoa, S.; Yang, R.; Koenig, M. Pathogenic roles of autoantibodies in systemic sclerosis: Current understandings in pathogenesis. J. Scleroderma Relat. Disord. 2019, 5, 103–129. [Google Scholar] [CrossRef]
- Sato, S.; Fujimoto, M.; Hasegawa, M.; Takehara, K. Altered blood B lymphocyte homeostasis in systemic sclerosis: Expanded naive B cells and diminished but activated memory B cells. Arthritis Rheum. 2004, 50, 1918–1927. [Google Scholar] [CrossRef]
- Simon, D.; Balogh, P.; Bognár, A.; Kellermayer, Z.; Engelmann, P.; Németh, P.; Farkas, N.; Minier, T.; Lóránd, V.; Czirják, L.; et al. Reduced non-switched memory B cell subsets cause imbalance in B cell repertoire in systemic sclerosis. Clin. Exp. Rheumatol. 2016, 34, 30–36. [Google Scholar]
- Katsenelson, N.; Kanswal, S.; Puig, M.; Mostowski, H.; Verthelyi, D.; Akkoyunlu, M. Synthetic CpG oligodeoxynucleotides augment BAFF- and APRIL-mediated immunoglobulin secretion. Eur. J. Immunol. 2007, 37, 1785–1795. [Google Scholar] [CrossRef]
- Acosta-Rodríguez, E.E.; Craxton, A.; Hendricks, D.W.; Merino, M.C.; Montes, C.L.; Clark, E.A.; Gruppi, A. BAFF and LPS cooperate to induce B cells to become susceptible to CD95/Fas-mediated cell death. Eur. J. Immunol. 2007, 37, 990–1000. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | dcSSc Patients (n = 26) |
|---|---|
| Age (years), mean (SD) | 49.88 (15.6) |
| Gender (female), n (%) | 22 (84.6%) |
| Disease duration 1 (years), mean (SD) | 1.73 (0.9) |
| Organ involvement | |
| MRSS mean (SD) | 16.42 (11.9) |
| Lung fibrosis 2, n (%) | 19/26 (73.1%) |
| Pulmonary arterial hypertension 3, n (%) | 0/26 (0%) |
| Renal involvement 4, n (%) | 1/26 (3.9%) |
| Gastrointestinal involvement 5, n (%) | 18/26 (69.2%) |
| Cardiac involvement 6, n (%) | 14/26 (53.9%) |
| Antibodies | |
| Anti-Scl-70+, n (%) | 12/26 (46.2%) |
| Anti-RNA-polymerase III+, n (%) | 4/26 (15.4%) |
| Immunosuppressive therapy 7, n (%) | 17/26 (65.4%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erdő-Bonyár, S.; Rapp, J.; Szinger, D.; Minier, T.; Kumánovics, G.; Czirják, L.; Berki, T.; Simon, D. Ligation of TLR Homologue CD180 of B Cells Activates the PI3K/Akt/mTOR Pathway in Systemic Sclerosis and Induces a Pathological Shift in the Expression of BAFF Receptors. Int. J. Mol. Sci. 2022, 23, 6777. https://doi.org/10.3390/ijms23126777
Erdő-Bonyár S, Rapp J, Szinger D, Minier T, Kumánovics G, Czirják L, Berki T, Simon D. Ligation of TLR Homologue CD180 of B Cells Activates the PI3K/Akt/mTOR Pathway in Systemic Sclerosis and Induces a Pathological Shift in the Expression of BAFF Receptors. International Journal of Molecular Sciences. 2022; 23(12):6777. https://doi.org/10.3390/ijms23126777
Chicago/Turabian StyleErdő-Bonyár, Szabina, Judit Rapp, Dávid Szinger, Tünde Minier, Gábor Kumánovics, László Czirják, Timea Berki, and Diána Simon. 2022. "Ligation of TLR Homologue CD180 of B Cells Activates the PI3K/Akt/mTOR Pathway in Systemic Sclerosis and Induces a Pathological Shift in the Expression of BAFF Receptors" International Journal of Molecular Sciences 23, no. 12: 6777. https://doi.org/10.3390/ijms23126777
APA StyleErdő-Bonyár, S., Rapp, J., Szinger, D., Minier, T., Kumánovics, G., Czirják, L., Berki, T., & Simon, D. (2022). Ligation of TLR Homologue CD180 of B Cells Activates the PI3K/Akt/mTOR Pathway in Systemic Sclerosis and Induces a Pathological Shift in the Expression of BAFF Receptors. International Journal of Molecular Sciences, 23(12), 6777. https://doi.org/10.3390/ijms23126777