
The Role of Kisspeptin in the Pathogenesis of Pregnancy Complications: A Narrative Review

 , ,
, ,
Abstract
1. Introduction
2. Scope and Methodology
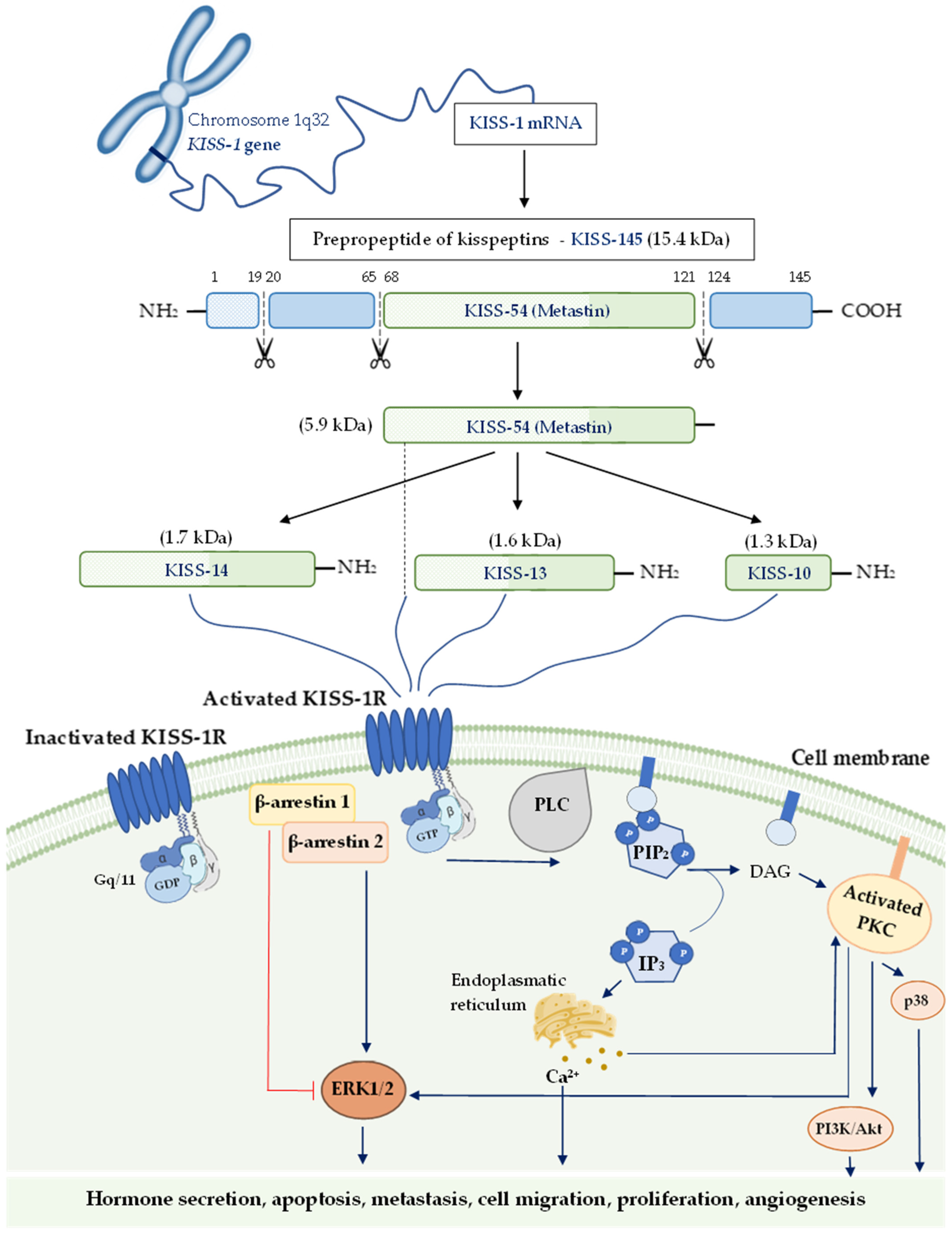
3. The KISS-1 Gene, Kisspeptin and Its Signaling
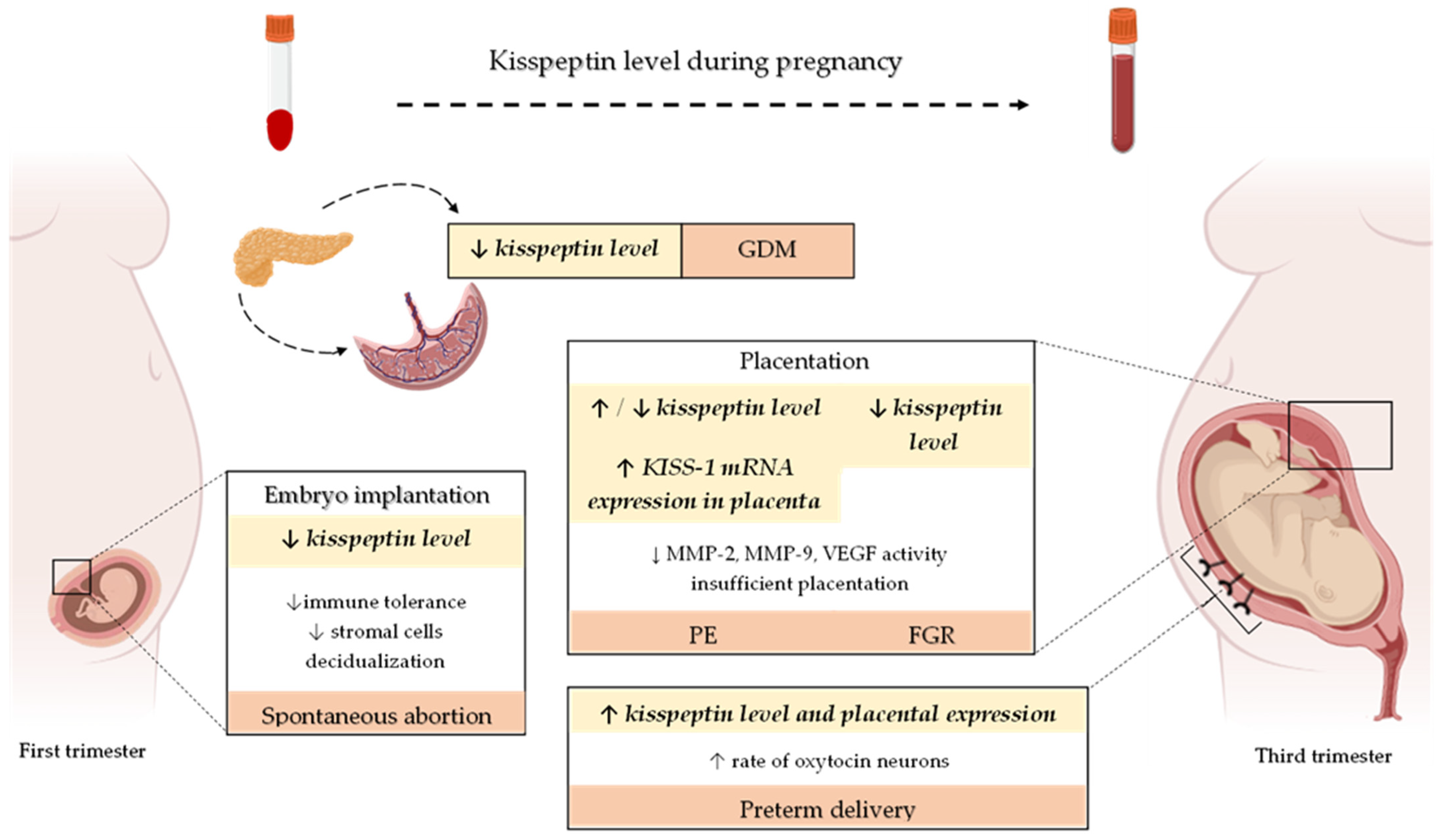
4. Kisspeptin Level throughout Pregnancy
5. Role of Kisspeptin in Gestational Diabetes Mellitus—A Placenta-Pancreas Crosstalk
6. Role of Kisspeptin in the Pathogenesis of Disorders from the Spectrum of Impaired Implantation and Placentation
6.1. Implantation and Kisspeptin-Induced Miscarriages
6.2. Kisspeptin in Placentation and Pathophysiology of Placental Diseases
6.2.1. Kisspeptin and Preeclampsia
6.2.2. Kisspeptin and Fetal Growth Restriction
7. Kisspeptin and Neonatal Outcomes
8. Conclusions and Future Perspectives

{kind=link}
{kind=link}
| Disorder | KISS Derivatives | Ethnicity (Country) | Sample Type | Trimester of Pregnancy | Model (Number) of the Study Groups | Description of Findings | Authors |
|---|---|---|---|---|---|---|---|
| KISS | Mixed (White, Black, Asian, Other) | plasma | All trimester | PE (20): mild (7), severe (13) PIH (12) CG (265) | ↑ KISS level in HDP women vs. CG The rate of rise in KISS with gestation ↑ in pregnancies affected by HDP than in CG OR of HDP increased by 30% (95% CI, 16–47%; p < 0.0001) for every 1 nmol/L increase in plasma kisspeptin | Abbara et al., 2022 [11] | |
| KISS-1 | Asian (Turkey) | plasma | <34. hbd ≥34. hbd | early-onset PE (20) vs. CG (20) late-onset PE (45) vs. CG (40) | No difference ↑ level of KISS-1 in late-onset PE vs. CG | Ibanoglu et al., 2022 [121] | |
| KISS-1 | Asian (China) | serum | Second Third | PE women (25) CG (25) | ↑ levels of KISS-1 and TGF-β1 in PE women vs. CG | Fang et al., 2022 [48] | |
| KISS-1 | Asian (China) | human trophoblast cell line placenta serum | Second Third | PE women (17) CG (16) | EGF downregulated KISS-1 by activating EGFR-mediated PI3K/AKT signaling pathway ↓ EGF and ↑ KISS-1 level in PE women vs. CG | Fang et al., 2021 [111] | |
| KISS-10 | Asian (Jordan) | plasma | Second Third | PE women (60) CG (40) | ↓ KISS-10 level in PE women vs. CG KISS-10 levels correlate positively with β-hCG and negatively with LH, and FSH in PE women in the third trimester | Al-Kaabi et al., 2020 [112] | |
| KISS-10 | Asian (Iraq) | plasma | Second Third | PE women (60) Mild form (39) Severe form (21) CG (40) | ↓ KISS-10 level in PE vs. CG ↓ KISS-10 level in severe PE vs. mild one ↓ KISS-10 level in severe PE during the second trimester vs. the third one | Ziyaraa et al., 2016 [114] | |
| KISS-10 GPR-54 | European (UK) | placenta serum (maternal, umbilical cord blood | Third | PE women (19) CG (30) | ↑ KISS expression in PE placenta vs. CG No differences in KISS-1 and GPR-54 mRNA expressions in placentas ↓ KISS-10 level in serum in SG vs. CG No significant distinctions in KISS-10 levels in cord blood between SG and CG | Matjila et al., 2016 [116] | |
| KISS-1KISS-1R | European (UK) | placenta | First Third | First trimester (10) PE women at delivery (10) CG (10) | ↓ KISS-1 and KISS-1R expression between early and term pregnancy PE vs. normal pregnancy placental samples: ↓ KISS-1 expression and ↑ expression of KISS-1R | Cartwright et al., 2012 [55] | |
| KISS | European (UK) | plasma | 16. hbd 28. hbd 36. hbd | Obese women with PE (11) Uncomplicated pregnancy in obese women (158) Lean pregnant women—CG (48) | Maternal KISS levels ↑ during pregnancy ↓ KISS level in obese women with PE at 16. hbd compared to obese pregnant women without PE and CG Optimal cut-off concentration of KISS at 16. hbd to predict PE—596 pmol/L (sensitivity: 85.7%; specificity: 71.4%) | Logie et al., 2012 [127] | |
| KISS-1 GPR-54 | Asian (China) | placenta | 23+0–33+6. hbd 34+0–39+0. hbd | early-onset PE (36) vs. CG (40) late-onset PE (40) vs. CG (40) | ↑ KISS-1 mRNA expression in early-onset SG vs. CG No differences in KISS-1 mRNA expression in late-onset PE No differences in GPR-54 expression | Qiao et al., 2012 [118] | |
| KISS-54 | Asian (Turkey) | serum | First | Women who developed PE (31) CG (30) | ↓ KISS-54 level in SG vs. CG (AUC: 0.797 to predict PE) | Madazli et al., 2012 [115] | |
| KISS-54 | Asian (Turkey) | plasma | Third | Mild PE (15) Severe PE (24) CG (50) | ↓ KISS-54 level in PE vs. CG No differences between mild and severe PE | Adali et al., 2012 [113] | |
| KISS-54 KISS-14 KISS-10 | European (Serbia) | plasma | Second Third | PE (28) GH (18) CG (25) | ↓ KISS-54 level in PE vs. CG No differences in KISS level between GH and CG | Cetković et al., 2012 [68] | |
| KISS-1 | Asian (China) | placenta human trophoblast cell line | Third | PE women (47) CG (30) Human trophoblast cell line transfected (SG) and non-transfected one (CG) with KISS-1 vector | ↑ KISS-1 mRNA and protein expression in PE women vs. CG ↓ MMP-9, MMP-2 mRNA, and protein expression in PE vs. CG No differences in cell proliferation between SG and CG ↓ invasion ability in SG compared to CG | Zhang et al., 2011 [119] | |
| KISS-1 | Amerindian (Mexico) | placenta | Third | PE women (27) CG (27) | ↑ KISS-1 expression in SG vs. CG | Vazquez-Alaniz et al., 2011 [120] | |
| KISS-54 | European (UK) | plasma | Third | PE women (8) PIH (19) CG (78) | No differences | Nijher et al., 2010 [136] | |
| KISS-54 | European (UK) | serum | Second | PE women (57) CG (317) | ↓ KISS-54 level in PE individuals vs. CG | Armstrong et al., 2009 [117] | |
| KISS-1 | European (Italy) | whole blood | Third | PE (6) CG (30) | ↓ KISS-1 mRNA expression in PE vs. CG | Farina et al., 2006 [137] | |
| Fetal Growth Restriction | KISS | Mixed (White, Black, Asian, Other) | plasma | All trimester | FGR or SGA (17) CG (265) | ↓ KISS level in FGR vs. CG (especially in the late first and third trimester) Lower increase in KISS level during gestation in FGR vs. CG KISS in diagnosis FGR: OR 0.72 (0.54–0.96), p = 0.025 | Abbara et al., 2022 [11] |
| KISS-10 | Asian (Iraq) | plasma | Second Third | PE women (60) Mild form (39) Severe form (21) CG (40) | Positive correlation between KISS level in the third trimester and EFW (r = 0.395, p = 0.012) in CG, and in the second (r = 0.760, p = 0.001), third (r = 0.920, p = 0.0001) trimesters in severe PE Negative correlation between KISS level in the third trimester and FBW in CG (r = −0.410, p = 0.009) | Ziyaraa et al., 2016 [114] | |
| KISS-1 | Mixed (Australia) | Whole blood (maternal) | 26–30 hbd | Late-onset FGR (40) CG (80) | ↑ KISS-1 expression in FGR vs. CG AUC for KISS01 as a discriminative marker for FGR: 0.64, p = 0.01 (FGR: 0.19 (0.08–0.81) vs. CG: 0.96 (0.13–2.85) | Whitehead et al., 2016 [138] | |
| KISS-1 KISS-1R | Animal model (rat) | Placenta | 16. and 22. day of gestation | Dexamethasone-induced FGR (6) CG (6) | ↑ KISS-1 mRNA expression in FGR vs. CG ↑ KISS-1R mRNA expression in 16. day of gestation and ↓ in 22. day of gestation in FGR vs. CG | Mark et al., 2013 [139] | |
| KISS-54 KISS-14 KISS-10 | European (Serbia) | plasma | Second Third | PE (28) GH (18) CG (25) | No correlation between KISS-54, placental, and birth weight at delivery in all groups | Cetković et al., 2012 [68] | |
| KISS | European (UK) | plasma | 16. hbd 28. hbd 36. hbd | Obese women with PE (11) Uncomplicated pregnancy in obese women (158) Lean pregnant women—CG (48) | Lower KISS levels at 16. hbd were associated with lower birth weight (r = 0.16, p = 0.06) | Logie et al., 2012 [127] | |
| KISS-1 | Mixed (Australia) | Whole blood Placenta | Third (<34. hbd) | FGR (20) Preterm Birth (15—blood samples, 8—placenta) Term Birth (8—placenta) | ↑ KISS-1 RNA expression in maternal blood in FGR vs. women with preterm delivery ↑ KISS-1 RNA expression in the placenta in FGR vs. women with preterm and term delivery | Whitehead et al., 2012 [140] | |
| KISS-10 | Asian (Turkey) | Plasma | First | Women with early pregnancy bleeding (20) CG (20) | ↓ KISS-10 level in the first trimester in SG and was associated with an increased risk of IUGR (10% vs. 0%) | Kavvasoglu et al., 2012 [83] | |
| KISS-54 | European (UK) | Serum | Second | FGR (118) CG (317) | ↓ KISS-54 level in FGR vs. CG | Armstrong et al., 2009 [117] | |
| KISS-54 | European (Netherlands) | Plasma | 8–14 hbd. | SGA (31) CG (31) | ↓ KISS-54 level in SGA vs. CG | Smets et al., 2008 [128] |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Napso, T.; Yong, H.E.J.; Lopez-Tello, J.; Sferruzzi-Perri, A.N. The Role of Placental Hormones in Mediating Maternal Adaptations to Support Pregnancy and Lactation. Front. Physiol. 2018, 9, 1091. [Google Scholar] [CrossRef]
- Urato, A.C.; Norwitz, E.R. A guide towards pre-pregnancy management of defective implantation and placentation. Best. Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 367–387. [Google Scholar] [CrossRef] [PubMed]
- Fowden, A.L.; Forhead, A.J.; Coan, P.M.; Burton, G.J. The placenta and intrauterine programming. J. Neuroendocr. 2008, 20, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Jena, M.K.; Sharma, N.R.; Petitt, M.; Maulik, D.; Nayak, N.R. Pathogenesis of Preeclampsia and Therapeutic Approaches Targeting the Placenta. Biomolecules 2020, 10, 953. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Groom, K.M.; Oyston, C.; Chamley, L.W.; Clark, A.R.; James, J.L. The placenta in fetal growth restriction: What is going wrong? Placenta 2020, 96, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Wong, I.; Moller, A.; Giachini, F.R.; Lima, V.V.; Toledo, F.; Stojanova, J.; Sobrevia, L.; San Martín, S. Placental structure in gestational diabetes mellitus. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165535. [Google Scholar] [CrossRef]
- Morgan, T.K. Role of the Placenta in Preterm Birth: A Review. Am. J. Perinatol. 2016, 33, 258–266. [Google Scholar] [CrossRef]
- Kebede, Z.T.; Yigezaw, G.S.; Yilma, T.M.; Delele, T.G. Prevalence of pregnancy-related complications and associated factors among reproductive-aged women in northwest Ethiopia: A community-based retrospective cross-sectional study. Int. J. Gynaecol. Obstet. 2021, 154, 62–71. [Google Scholar] [CrossRef]
- Sullivan-Pyke, C.; Haisenleder, D.J.; Senapati, S.; Nicolais, O.; Eisenberg, E.; Sammel, M.D.; Barnhart, K.T. Kisspeptin as a new serum biomarker to discriminate miscarriage from viable intrauterine pregnancy. Fertil. Steril. 2018, 109, 137–141.e2. [Google Scholar] [CrossRef]
- Kapustin, R.V.; Drobintseva, A.O.; Alekseenkova, E.N.; Onopriychuk, A.R.; Arzhanova, O.N.; Polyakova, V.O.; Kvetnoy, I.M. Placental protein expression of kisspeptin-1 (KISS1) and the kisspeptin-1 receptor (KISS1R) in pregnancy complicated by diabetes mellitus or preeclampsia. Arch. Gynecol. Obstet. 2020, 301, 437–445. [Google Scholar] [CrossRef]
- Abbara, A.; Al-Memar, M.; Phylactou, M.; Daniels, E.; Patel, B.; Eng, P.C.; Nadir, R.; Izzi-Engbeaya, C.; Clarke, S.A.; Mills, E.G.; et al. Changes in Circulating Kisspeptin Levels During Each Trimester in Women with Antenatal Complications. J. Clin. Endocrinol. Metab. 2022, 107, e71–e83. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Miele, M.E.; Hicks, D.J.; Phillips, K.K.; Trent, J.M.; Weissman, B.E.; Welch, D.R. KiSS-1, a novel human malignant melanoma metastasis-suppressor gene. J. Natl. Cancer Inst. 1996, 88, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- De Roux, N.; Genin, E.; Carel, J.-C.; Matsuda, F.; Chaussain, J.-L.; Milgrom, E. Hypogonadotropic hypogonadism due to loss of function of the KiSS1-derived peptide receptor GPR54. Proc. Natl. Acad. Sci. USA 2003, 100, 10972–10976. [Google Scholar] [CrossRef]
- Ke, R.; Ma, X.; Lee, L.T.O. Understanding the functions of kisspeptin and kisspeptin receptor (Kiss1R) from clinical case studies. Peptides 2019, 120, 170019. [Google Scholar] [CrossRef] [PubMed]
- Mead, E.J.; Maguire, J.J.; Kuc, R.E.; Davenport, A.P. Kisspeptins are novel potent vasoconstrictors in humans, with a discrete localization of their receptor, G protein-coupled receptor 54, to atherosclerosis-prone vessels. Endocrinology 2007, 148, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Gorbunova, O.L.; Shirshev, S.V. Role of Kisspeptin in Regulation of Reproductive and Immune Reactions. Biochemistry 2020, 85, 839–853. [Google Scholar] [CrossRef] [PubMed]
- Messager, S.; Chatzidaki, E.E.; Ma, D.; Hendrick, A.G.; Zahn, D.; Dixon, J.; Thresher, R.R.; Malinge, I.; Lomet, D.; Carlton, M.B.L.; et al. Kisspeptin directly stimulates gonadotropin-releasing hormone release via G protein-coupled receptor 54. Proc. Natl. Acad. Sci. USA 2005, 102, 1761–1766. [Google Scholar] [CrossRef]
- Öztin, H.; Çağıltay, E.; Çağlayan, S.; Kaplan, M.; Akpak, Y.K.; Karaca, N.; Tığlıoğlu, M. Kisspeptin levels in idiopathic hypogonadotropic hypogonadism diagnosed male patients and its relation with glucose-insulin dynamic. Gynecol. Endocrinol. 2016, 32, 991–994. [Google Scholar] [CrossRef]
- Hauge-Evans, A.C.; Richardson, C.C.; Milne, H.M.; Christie, M.R.; Persaud, S.J.; Jones, P.M. A role for kisspeptin in islet function. Diabetologia 2006, 49, 2131–2135. [Google Scholar] [CrossRef]
- Schwetz, T.A.; Reissaus, C.A.; Piston, D.W. Differential stimulation of insulin secretion by GLP-1 and Kisspeptin-10. PLoS ONE 2014, 9, e113020. [Google Scholar] [CrossRef]
- Bowe, J.E.; Foot, V.L.; Amiel, S.A.; Huang, G.C.; Lamb, M.W.; Lakey, J.; Jones, P.M.; Persaud, S.J. GPR54 peptide agonists stimulate insulin secretion from murine, porcine and human islets. Islets 2012, 4, 20–23. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Izzi-Engbeaya, C.; Comninos, A.N.; Clarke, S.A.; Jomard, A.; Yang, L.; Jones, S.; Abbara, A.; Narayanaswamy, S.; Eng, P.C.; Papadopoulou, D.; et al. The effects of kisspeptin on β-cell function, serum metabolites and appetite in humans. Diabetes Obes. Metab. 2018, 20, 2800–2810. [Google Scholar] [CrossRef] [PubMed]
- Tolson, K.P.; Garcia, C.; Yen, S.; Simonds, S.; Stefanidis, A.; Lawrence, A.; Smith, J.T.; Kauffman, A.S. Impaired kisspeptin signaling decreases metabolism and promotes glucose intolerance and obesity. J. Clin. Investig. 2014, 124, 3075–3079. [Google Scholar] [CrossRef] [PubMed]
- Backholer, K.; Smith, J.T.; Rao, A.; Pereira, A.; Iqbal, J.; Ogawa, S.; Li, Q.; Clarke, I.J. Kisspeptin cells in the ewe brain respond to leptin and communicate with neuropeptide Y and proopiomelanocortin cells. Endocrinology 2010, 151, 2233–2243. [Google Scholar] [CrossRef]
- Orlando, G.; Leone, S.; Ferrante, C.; Chiavaroli, A.; Mollica, A.; Stefanucci, A.; Macedonio, G.; Dimmito, M.P.; Leporini, L.; Menghini, L.; et al. Effects of Kisspeptin-10 on Hypothalamic Neuropeptides and Neurotransmitters Involved in Appetite Control. Molecules 2018, 23, 3071. [Google Scholar] [CrossRef]
- Kołodziejski, P.A.; Pruszyńska-Oszmałek, E.; Korek, E.; Sassek, M.; Szczepankiewicz, D.; Kaczmarek, P.; Nogowski, L.; Maćkowiak, P.; Nowak, K.W.; Krauss, H.; et al. Serum levels of spexin and kisspeptin negatively correlate with obesity and insulin resistance in women. Physiol. Res. 2018, 67, 45–56. [Google Scholar] [CrossRef]
- Fu, L.-Y.; van den Pol, A.N. Kisspeptin directly excites anorexigenic proopiomelanocortin neurons but inhibits orexigenic neuropeptide Y cells by an indirect synaptic mechanism. J. Neurosci. 2010, 30, 10205–10219. [Google Scholar] [CrossRef]
- Bilban, M.; Ghaffari-Tabrizi, N.; Hintermann, E.; Bauer, S.; Molzer, S.; Zoratti, C.; Malli, R.; Sharabi, A.; Hiden, U.; Graier, W.; et al. Kisspeptin-10, a KiSS-1/metastin-derived decapeptide, is a physiological invasion inhibitor of primary human trophoblasts. J. Cell Sci. 2004, 117, 1319–1328. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, H.; Tian, J.; Liu, L.; Dong, Y.; Mao, T. Expression of kisspeptin/GPR54 and PIBF/PR in the first trimester trophoblast and decidua of women with recurrent spontaneous abortion. Pathol. Res. Pract. 2014, 210, 47–54. [Google Scholar] [CrossRef]
- Howell, K.R.; Powell, T.L. Effects of maternal obesity on placental function and fetal development. Reproduction 2017, 153, R97–R108. [Google Scholar] [CrossRef]
- West, A.; Vojta, P.J.; Welch, D.R.; Weissman, B.E. Chromosome localization and genomic structure of the KiSS-1 metastasis suppressor gene (KISS1). Genomics 1998, 54, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Ohtaki, T.; Shintani, Y.; Honda, S.; Matsumoto, H.; Hori, A.; Kanehashi, K.; Terao, Y.; Kumano, S.; Takatsu, Y.; Masuda, Y.; et al. Metastasis suppressor gene KiSS-1 encodes peptide ligand of a G-protein-coupled receptor. Nature 2001, 411, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Kotani, M.; Detheux, M.; Vandenbogaerde, A.; Communi, D.; Vanderwinden, J.M.; Le Poul, E.; Brézillon, S.; Tyldesley, R.; Suarez-Huerta, N.; Vandeput, F.; et al. The metastasis suppressor gene KiSS-1 encodes kisspeptins, the natural ligands of the orphan G protein-coupled receptor GPR54. J. Biol. Chem. 2001, 276, 34631–34636. [Google Scholar] [CrossRef] [PubMed]
- Pinilla, L.; Aguilar, E.; Dieguez, C.; Millar, R.P.; Tena-Sempere, M. Kisspeptins and reproduction: Physiological roles and regulatory mechanisms. Physiol. Rev. 2012, 92, 1235–1316. [Google Scholar] [CrossRef]
- Lee, D.K.; Nguyen, T.; O’Neill, G.P.; Cheng, R.; Liu, Y.; Howard, A.D.; Coulombe, N.; Tan, C.P.; Tang-Nguyen, A.T.; George, S.R.; et al. Discovery of a receptor related to the galanin receptors. FEBS Lett. 1999, 446, 103–107. [Google Scholar] [CrossRef]
- Muir, A.I.; Chamberlain, L.; Elshourbagy, N.A.; Michalovich, D.; Moore, D.J.; Calamari, A.; Szekeres, P.G.; Sarau, H.M.; Chambers, J.K.; Murdock, P.; et al. AXOR12, a novel human G protein-coupled receptor, activated by the peptide KiSS-1. J. Biol. Chem. 2001, 276, 28969–28975. [Google Scholar] [CrossRef]
- Gottsch, M.L.; Clifton, D.K.; Steiner, R.A. From KISS1 to kisspeptins: An historical perspective and suggested nomenclature. Peptides 2009, 30, 4–9. [Google Scholar] [CrossRef][Green Version]
- Fredriksson, R.; Lagerström, M.C.; Lundin, L.-G.; Schiöth, H.B. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol. Pharmacol. 2003, 63, 1256–1272. [Google Scholar] [CrossRef]
- Castaño, J.P.; Martínez-Fuentes, A.J.; Gutiérrez-Pascual, E.; Vaudry, H.; Tena-Sempere, M.; Malagón, M.M. Intracellular signaling pathways activated by kisspeptins through GPR54: Do multiple signals underlie function diversity? Peptides 2009, 30, 10–15. [Google Scholar] [CrossRef]
- Liu, X.; Lee, K.; Herbison, A.E. Kisspeptin excites gonadotropin-releasing hormone neurons through a phospholipase C/calcium-dependent pathway regulating multiple ion channels. Endocrinology 2008, 149, 4605–4614. [Google Scholar] [CrossRef]
- Peng, J.; Tang, M.; Zhang, B.-P.; Zhang, P.; Zhong, T.; Zong, T.; Yang, B.; Kuang, H.-B. Kisspeptin stimulates progesterone secretion via the Erk1/2 mitogen-activated protein kinase signaling pathway in rat luteal cells. Fertil. Steril. 2013, 99, 1436–1443.e1. [Google Scholar] [CrossRef] [PubMed]
- Szereszewski, J.M.; Pampillo, M.; Ahow, M.R.; Offermanns, S.; Bhattacharya, M.; Babwah, A.V. GPR54 regulates ERK1/2 activity and hypothalamic gene expression in a Gα(q/11) and β-arrestin-dependent manner. PLoS ONE 2010, 5, e12964. [Google Scholar] [CrossRef] [PubMed]
- Pampillo, M.; Camuso, N.; Taylor, J.E.; Szereszewski, J.M.; Ahow, M.R.; Zajac, M.; Millar, R.P.; Bhattacharya, M.; Babwah, A.V. Regulation of GPR54 signaling by GRK2 and {beta}-arrestin. Mol. Endocrinol. 2009, 23, 2060–2074. [Google Scholar] [CrossRef] [PubMed]
- Ramaesh, T.; Logie, J.J.; Roseweir, A.K.; Millar, R.P.; Walker, B.R.; Hadoke, P.W.F.; Reynolds, R.M. Kisspeptin-10 inhibits angiogenesis in human placental vessels ex vivo and endothelial cells in vitro. Endocrinology 2010, 151, 5927–5934. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-G.; Yi, Z.; Pang, X.; Yi, T.; Wang, Y.; Luo, J.; Wu, Z.; Li, D.; Liu, M. Kisspeptin-10, a KISS1-derived decapeptide, inhibits tumor angiogenesis by suppressing Sp1-mediated VEGF expression and FAK/Rho GTPase activation. Cancer Res. 2009, 69, 7062–7070. [Google Scholar] [CrossRef]
- Francis, V.A.; Abera, A.B.; Matjila, M.; Millar, R.P.; Katz, A.A. Kisspeptin regulation of genes involved in cell invasion and angiogenesis in first trimester human trophoblast cells. PLoS ONE 2014, 9, e99680. [Google Scholar] [CrossRef]
- Lechan, R.M.; Fekete, C. Feedback regulation of thyrotropin-releasing hormone (TRH): Mechanisms for the non-thyroidal illness syndrome. J. Endocrinol. Investig. 2004, 27, 105–119. [Google Scholar]
- Fang, L.; Yan, Y.; Gao, Y.; Wu, Z.; Wang, Z.; Yang, S.; Cheng, J.-C.; Sun, Y.-P. TGF-β1 inhibits human trophoblast cell invasion by upregulating kisspeptin expression through ERK1/2 but not SMAD signaling pathway. Reprod. Biol. Endocrinol. 2022, 20, 22. [Google Scholar] [CrossRef]
- Horikoshi, Y.; Matsumoto, H.; Takatsu, Y.; Ohtaki, T.; Kitada, C.; Usuki, S.; Fujino, M. Dramatic elevation of plasma metastin concentrations in human pregnancy: Metastin as a novel placenta-derived hormone in humans. J. Clin. Endocrinol. Metab. 2003, 88, 914–919. [Google Scholar] [CrossRef]
- Babwah, A.V. Uterine and placental KISS1 regulate pregnancy: What we know and the challenges that lie ahead. Reproduction 2015, 150, R121–R128. [Google Scholar] [CrossRef]
- Mondal, M.; Baruah, K.K.; Prakash, B.S. Determination of plasma kisspeptin concentrations during reproductive cycle and different phases of pregnancy in crossbred cows using bovine specific enzyme immunoassay. Gen. Comp. Endocrinol. 2015, 224, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Jayasena, C.N.; Comninos, A.N.; Narayanaswamy, S.; Abbara, A.; Nijher, G.M.K.; Cheema, M.; Malik, Z.; Ghatei, M.A.; Bloom, S.R.; Dhillo, W.S. The identification of elevated urinary kisspeptin-immunoreactivity during pregnancy. Ann. Clin. Biochem. 2015, 52, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Jayasena, C.N.; Abbara, A.; Izzi-Engbeaya, C.; Comninos, A.N.; Harvey, R.A.; Gonzalez Maffe, J.; Sarang, Z.; Ganiyu-Dada, Z.; Padilha, A.I.; Dhanjal, M.; et al. Reduced levels of plasma kisspeptin during the antenatal booking visit are associated with increased risk of miscarriage. J. Clin. Endocrinol. Metab. 2014, 99, E2652–E2660. [Google Scholar] [CrossRef] [PubMed]
- Janneau, J.-L.; Maldonado-Estrada, J.; Tachdjian, G.; Miran, I.; Motté, N.; Saulnier, P.; Sabourin, J.-C.; Coté, J.-F.; Simon, B.; Frydman, R.; et al. Transcriptional expression of genes involved in cell invasion and migration by normal and tumoral trophoblast cells. J. Clin. Endocrinol. Metab. 2002, 87, 5336–5339. [Google Scholar] [CrossRef]
- Cartwright, J.E.; Williams, P.J. Altered placental expression of kisspeptin and its receptor in pre-eclampsia. J. Endocrinol. 2012, 214, 79–85. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, C. Prevalence of Gestational Diabetes and Risk of Progression to Type 2 Diabetes: A Global Perspective. Curr. Diabetes Rep. 2016, 16, 7. [Google Scholar] [CrossRef]
- ACOG Practice Bulletin No. 190: Gestational Diabetes Mellitus. Obstet. Gynecol. 2018, 131, e49–e64. [CrossRef]
- Ramezani Tehrani, F.; Naz, M.S.G.; Yarandi, R.B.; Behboudi-Gandevani, S. The Impact of Diagnostic Criteria for Gestational Diabetes Mellitus on Adverse Maternal Outcomes: A Systematic Review and Meta-Analysis. J. Clin. Med. 2021, 10, 666. [Google Scholar] [CrossRef]
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2020. Diabetes Care 2020, 43, S14–S31. [Google Scholar] [CrossRef]
- Yen, I.-W.; Lee, C.-N.; Lin, M.-W.; Fan, K.-C.; Wei, J.-N.; Chen, K.-Y.; Chen, S.-C.; Tai, Y.-Y.; Kuo, C.-H.; Lin, C.-H.; et al. Overweight and obesity are associated with clustering of metabolic risk factors in early pregnancy and the risk of GDM. PLoS ONE 2019, 14, e0225978. [Google Scholar] [CrossRef]
- Alejandro, E.U.; Mamerto, T.P.; Chung, G.; Villavieja, A.; Gaus, N.L.; Morgan, E.; Pineda-Cortel, M.R.B. Gestational Diabetes Mellitus: A Harbinger of the Vicious Cycle of Diabetes. Int. J. Mol. Sci. 2020, 21, 5003. [Google Scholar] [CrossRef] [PubMed]
- Baeyens, L.; Hindi, S.; Sorenson, R.L.; German, M.S. β-Cell adaptation in pregnancy. Diabetes Obes. Metab. 2016, 18 (Suppl. S1), 63–70. [Google Scholar] [CrossRef] [PubMed]
- Baz, B.; Riveline, J.-P.; Gautier, J.-F. Endocrinology of Pregnancy: Gestational diabetes mellitus: Definition, aetiological and clinical aspects. Eur. J. Endocrinol. 2016, 174, R43–R51. [Google Scholar] [CrossRef] [PubMed]
- Bowe, J.E.; Hill, T.G.; Hunt, K.F.; Smith, L.I.; Simpson, S.J.; Amiel, S.A.; Jones, P.M. A role for placental kisspeptin in β cell adaptation to pregnancy. JCI Insight 2019, 4, e124540. [Google Scholar] [CrossRef]
- Wang, T.; Han, S.; Tian, W.; Zhao, M.; Zhang, H. Effects of kisspeptin on pathogenesis and energy metabolism in polycystic ovarian syndrome (PCOS). Gynecol. Endocrinol. 2019, 35, 807–810. [Google Scholar] [CrossRef]
- Panidis, D.; Rousso, D.; Koliakos, G.; Kourtis, A.; Katsikis, I.; Farmakiotis, D.; Votsi, E.; Diamanti-Kandarakis, E. Plasma metastin levels are negatively correlated with insulin resistance and free androgens in women with polycystic ovary syndrome. Fertil. Steril. 2006, 85, 1778–1783. [Google Scholar] [CrossRef]
- Arslan, E.; Gorkem, U.; Togrul, C. Is There an Association Between Kisspeptin Levels and Gestational Diabetes Mellitus? Gynecol. Obstet. Reprod. Med. 2020, 26, 179–183. [Google Scholar] [CrossRef]
- Cetković, A.; Miljic, D.; Ljubić, A.; Patterson, M.; Ghatei, M.; Stamenković, J.; Nikolic-Djurovic, M.; Pekic, S.; Doknic, M.; Glišić, A.; et al. Plasma kisspeptin levels in pregnancies with diabetes and hypertensive disease as a potential marker of placental dysfunction and adverse perinatal outcome. Endocr. Res. 2012, 37, 78–88. [Google Scholar] [CrossRef]
- Loegl, J.; Nussbaumer, E.; Cvitic, S.; Huppertz, B.; Desoye, G.; Hiden, U. GDM alters paracrine regulation of feto-placental angiogenesis via the trophoblast. Lab. Investig. 2017, 97, 409–418. [Google Scholar] [CrossRef]
- Norwitz, E.R.; Schust, D.J.; Fisher, S.J. Implantation and the survival of early pregnancy. N. Engl. J. Med. 2001, 345, 1400–1408. [Google Scholar] [CrossRef]
- National Institute for Health and Care Excellence National Collaborating Centre for Ws. Children’s H. National Institute for Health and Clinical excellence: Guidance. Ectopic Pregnancy and Miscarriage: Diagnosis and Initial Management; National Collaborating Centre for Women’s and Children’s Health, Rcog: London, UK, 2012. [Google Scholar]
- Fayazi, M.; Calder, M.; Bhattacharya, M.; Vilos, G.A.; Power, S.; Babwah, A.V. The pregnant mouse uterus exhibits a functional kisspeptin/KISS1R signaling system on the day of embryo implantation. Reprod. Biol. Endocrinol. 2015, 13, 105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Tang, M.; Zhong, T.; Lin, Y.; Zong, T.; Zhong, C.; Zhang, B.; Ren, M.; Kuang, H. Expression and function of kisspeptin during mouse decidualization. PLoS ONE 2014, 9, e97647. [Google Scholar] [CrossRef] [PubMed]
- McEwan, M.; Lins, R.J.; Munro, S.K.; Vincent, Z.L.; Ponnampalam, A.P.; Mitchell, M.D. Cytokine regulation during the formation of the fetal-maternal interface: Focus on cell-cell adhesion and remodelling of the extra-cellular matrix. Cytokine Growth Factor Rev. 2009, 20, 241–249. [Google Scholar] [CrossRef]
- Taylor, J.; Pampillo, M.; Bhattacharya, M.; Babwah, A.V. Kisspeptin/KISS1R signaling potentiates extravillous trophoblast adhesion to type-I collagen in a PKC- and ERK1/2-dependent manner. Mol. Reprod. Dev. 2014, 81, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Calder, M.; Chan, Y.-M.; Raj, R.; Pampillo, M.; Elbert, A.; Noonan, M.; Gillio-Meina, C.; Caligioni, C.; Bérubé, N.G.; Bhattacharya, M.; et al. Implantation failure in female Kiss1-/- mice is independent of their hypogonadic state and can be partially rescued by leukemia inhibitory factor. Endocrinology 2014, 155, 3065–3078. [Google Scholar] [CrossRef]
- Wu, H.-M.; Huang, H.-Y.; Soong, Y.-K.; Leung, P.C.K.; Wang, H.-S. Kisspeptin regulation of human decidual stromal cells motility via FAK-Src intracellular tyrosine kinases. Hum. Reprod. 2019, 34, 1291–1301. [Google Scholar] [CrossRef]
- Park, D.-W.; Lee, S.-K.; Hong, S.R.; Han, A.-R.; Kwak-Kim, J.; Yang, K.M. Expression of Kisspeptin and its receptor GPR54 in the first trimester trophoblast of women with recurrent pregnancy loss. Am. J. Reprod. Immunol. 2012, 67, 132–139. [Google Scholar] [CrossRef]
- Shirshev, S.V.; Nekrasova, I.V.; Gorbunova, O.L.; Orlova, E.G.; Maslennikova, I.L. The effect of kisspeptin on the functional characteristics of isolated NK cells. Dokl. Biol. Sci. 2015, 464, 267–269. [Google Scholar] [CrossRef]
- Ephrem, A.; Epstein, A.L.; Stephens, G.L.; Thornton, A.M.; Glass, D.; Shevach, E.M. Modulation of Treg cells/T effector function by GITR signaling is context-dependent. Eur. J. Immunol. 2013, 43, 2421–2429. [Google Scholar] [CrossRef]
- Mellor, A.L.; Munn, D.H. Tryptophan catabolism prevents maternal T cells from activating lethal anti-fetal immune responses. J. Reprod. Immunol. 2001, 52, 5–13. [Google Scholar] [CrossRef]
- Ichiyama, K.; Yoshida, H.; Wakabayashi, Y.; Chinen, T.; Saeki, K.; Nakaya, M.; Takaesu, G.; Hori, S.; Yoshimura, A.; Kobayashi, T. Foxp3 inhibits RORgammat-mediated IL-17A mRNA transcription through direct interaction with RORgammat. J. Biol. Chem. 2008, 283, 17003–17008. [Google Scholar] [CrossRef] [PubMed]
- Kavvasoglu, S.; Ozkan, Z.S.; Kumbak, B.; Sımsek, M.; Ilhan, N. Association of kisspeptin-10 levels with abortus imminens: A preliminary study. Arch. Gynecol. Obstet. 2012, 285, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Abbara, A.; Al-Memar, M.; Phylactou, M.; Kyriacou, C.; Eng, P.C.; Nadir, R.; Izzi-Engbeaya, C.; Clarke, S.A.; Mills, E.G.; Daniels, E.; et al. Performance of plasma kisspeptin as a biomarker for miscarriage improves with gestational age during the first trimester. Fertil. Steril. 2021, 116, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Gorkem, U.; Kan, O.; Bostanci, M.O.; Taskiran, D.; Inal, H.A. Kisspeptin and Hematologic Parameters as Predictive Biomarkers for First-Trimester Abortions. Medeni Med. J. 2021, 36, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Colak, E.; Ozcimen, E.E.; Erinanç, O.H.; Tohma, Y.A.; Ceran, M.U. Is placental KISS-1 expression associated with first trimester abortion spontaneous? Obstet. Gynecol. Sci. 2020, 63, 490–496. [Google Scholar] [CrossRef]
- Li, L.; Tian, J.; Zhou, L.; Wu, S.; Zhang, S.; Qi, L.; Zhang, H. Role of kisspeptin/GPR54 in the first trimester trophoblast of women with a history of recurrent spontaneous abortion. Int. J. Clin. Exp. Pathol. 2017, 10, 8161–8173. [Google Scholar]
- Mumtaz, A.; Khalid, A.; Jamil, Z.; Fatima, S.S.; Arif, S.; Rehman, R. Kisspeptin: A Potential Factor for Unexplained Infertility and Impaired Embryo Implantation. Int. J. Fertil. Steril. 2017, 11, 99–104. [Google Scholar] [CrossRef]
- Yu, H.; Liu, J.; Guo, H.; Chen, C.; Han, Y.; Cui, Y. Prognostic value of repeated serum kisspeptin measurements in early first trimester pregnancy: A preliminary study. Reprod. Biom. Online 2019, 38, 465–471. [Google Scholar] [CrossRef]
- Hu, K.-L.; Zhang, Y.; Yang, Z.; Zhao, H.; Xu, H.; Yu, Y.; Li, R. Predictive value of serum kisspeptin concentration at 14 and 21 days after frozen-thawed embryo transfer. Reprod. Biom. Online 2019, 39, 161–167. [Google Scholar] [CrossRef]
- Costanzo, V.; Bardelli, A.; Siena, S.; Abrignani, S. Exploring the links between cancer and placenta development. Open Biol. 2018, 8, 180081. [Google Scholar] [CrossRef]
- Yoshioka, K.; Ohno, Y.; Horiguchi, Y.; Ozu, C.; Namiki, K.; Tachibana, M. Effects of a KiSS-1 peptide, a metastasis suppressor gene, on the invasive ability of renal cell carcinoma cells through a modulation of a matrix metalloproteinase 2 expression. Life Sci. 2008, 83, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Fellows, E.J.; Hazzard, T.M.; Kutzler, M.A. Gene expression in pre-term, pre-labour and parturient canine placenta. Reprod. Domest. Anim. 2012, 47 (Suppl. S6), 182–185. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.H.; Oh, S.C.; Lee, H.J.; Kang, H.N.; Kim, J.L.; Kim, J.S.; Yoo, Y.A. Metastatic function of BMP-2 in gastric cancer cells: The role of PI3K/AKT, MAPK, the NF-κB pathway, and MMP-9 expression. Exp. Cell Res. Cell Res. 2011, 317, 1746–1762. [Google Scholar] [CrossRef]
- Yan, C.; Wang, H.; Boyd, D.D. KiSS-1 represses 92-kDa type IV collagenase expression by down-regulating NF-kappa B binding to the promoter as a consequence of Ikappa Balpha -induced block of p65/p50 nuclear translocation. J. Biol. Chem. 2001, 276, 1164–1172. [Google Scholar] [CrossRef] [PubMed]
- Torricelli, M.; Novembri, R.; Conti, N.; De Falco, G.; De Bonis, M.; Petraglia, F. Correlation with placental kisspeptin in postterm pregnancy and apoptosis. Reprod. Sci. 2012, 19, 1133–1137. [Google Scholar] [CrossRef]
- De Pedro, M.A.; Morán, J.; Díaz, I.; Murias, L.; Fernández-Plaza, C.; González, C.; Díaz, E. Circadian Kisspeptin expression in human term placenta. Placenta 2015, 36, 1337–1339. [Google Scholar] [CrossRef]
- Cao, Y.; Li, Z.; Jiang, W.; Ling, Y.; Kuang, H. Reproductive functions of Kisspeptin/KISS1R Systems in the Periphery. Reprod. Biol. Endocrinol. 2019, 17, 65. [Google Scholar] [CrossRef]
- Liu, H.; Li, Y.; Zhang, J.; Rao, M.; Liang, H.; Liu, G. The defect of both angiogenesis and lymphangiogenesis is involved in preeclampsia. Placenta 2015, 36, 279–286. [Google Scholar] [CrossRef]
- Tomimatsu, T.; Mimura, K.; Matsuzaki, S.; Endo, M.; Kumasawa, K.; Kimura, T. Preeclampsia: Maternal Systemic Vascular Disorder Caused by Generalized Endothelial Dysfunction Due to Placental Antiangiogenic Factors. Int. J. Mol. Sci. 2019, 20, 4246. [Google Scholar] [CrossRef]
- Stevens, D.U.; Smits, M.P.; Bulten, J.; Spaanderman, M.E.A.; van Vugt, J.M.G.; Al-Nasiry, S. Prevalence of hypertensive disorders in women after preeclamptic pregnancy associated with decidual vasculopathy. Hypertens. Pregnancy 2015, 34, 332–341. [Google Scholar] [CrossRef]
- Sharkey, A.M.; Cooper, J.C.; Balmforth, J.R.; McLaren, J.; Clark, D.E.; Charnock-Jones, D.S.; Morris, N.H.; Smith, S.K. Maternal plasma levels of vascular endothelial growth factor in normotensive pregnancies and in pregnancies complicated by pre-eclampsia. Eur. J. Clin. Investig. 1996, 26, 1182–1185. [Google Scholar] [CrossRef] [PubMed]
- Poniedziałek-Czajkowska, E.; Mierzyński, R. Could Vitamin D Be Effective in Prevention of Preeclampsia? Nutrients 2021, 13, 3854. [Google Scholar] [CrossRef] [PubMed]
- Mezei, Z.; Zamani-Forooshani, O.; Csabafi, K.; Szikszai, B.; Papp, E.; Ónodi, Á.; Török, D.; Leprán, Á.; Telegdy, G.; Szabó, G. The effect of kisspeptin on the regulation of vascular tone. Can. J. Physiol. Pharmacol. 2015, 93, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Maguire, J.J.; Kirby, H.R.; Mead, E.J.; Kuc, R.E.; d’Anglemont de Tassigny, X.; Colledge, W.H.; Davenport, A.P. Inotropic action of the puberty hormone kisspeptin in rat, mouse and human: Cardiovascular distribution and characteristics of the kisspeptin receptor. PLoS ONE 2011, 6, e27601. [Google Scholar] [CrossRef]
- Sato, K.; Shirai, R.; Hontani, M.; Shinooka, R.; Hasegawa, A.; Kichise, T.; Yamashita, T.; Yoshizawa, H.; Watanabe, R.; Matsuyama, T.-A.; et al. Potent Vasoconstrictor Kisspeptin-10 Induces Atherosclerotic Plaque Progression and Instability: Reversal by its Receptor GPR54 Antagonist. J. Am. Heart Assoc. 2017, 6, e005790. [Google Scholar] [CrossRef]
- Gestational Hypertension and Preeclampsia: ACOG Practice Bulletin Summary, Number 222. Obstet. Gynecol. Gynecol. 2020, 135, 1492–1495. [CrossRef] [PubMed]
- Aneman, I.; Pienaar, D.; Suvakov, S.; Simic, T.P.; Garovic, V.D.; McClements, L. Mechanisms of Key Innate Immune Cells in Early- and Late-Onset Preeclampsia. Front. Immunol. 2020, 11, 1864. [Google Scholar] [CrossRef]
- Fantone, S.; Mazzucchelli, R.; Giannubilo, S.R.; Ciavattini, A.; Marzioni, D.; Tossetta, G. AT-rich interactive domain 1A protein expression in normal and pathological pregnancies complicated by preeclampsia. Histochem. Cell Biol. 2020, 154, 339–346. [Google Scholar] [CrossRef]
- Opichka, M.A.; Rappelt, M.W.; Gutterman, D.D.; Grobe, J.L.; McIntosh, J.J. Vascular Dysfunction in Preeclampsia. Cells 2021, 10, 3055. [Google Scholar] [CrossRef]
- Fang, L.; Gao, Y.; Wang, Z.; Li, Y.; Yan, Y.; Wu, Z.; Cheng, J.-C.; Sun, Y.-P. EGF stimulates human trophoblast cell invasion by downregulating ID3-mediated KISS1 expression. Cell Commun. Signal. 2021, 19, 101. [Google Scholar] [CrossRef]
- Al-Kaabi, M.A.; Hamdan, F.B.; Al-Matubsi, H. Maternal plasma kisspeptin-10 level in preeclamptic pregnant women and its relation in changing their reproductive hormones. J. Obstet. Gynaecol. Res. 2020, 46, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Adali, E.; Kurdoglu, Z.; Kurdoglu, M.; Kamaci, M.; Kolusari, A.; Yildizhan, R. Metastin levels in pregnancies complicated by pre-eclampsia and their relation with disease severity. J. Matern. Fetal Neonatal Med. 2012, 25, 2671–2675. [Google Scholar] [CrossRef] [PubMed]
- Ziyaraa, M.A.; Hamdan, F.B.; Mousa, L.R. Correlation of Kisspeptin-10 level and fetal well-being in preeclamptic patients. Taiwan J. Obstet. Gynecol. 2016, 55, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Madazli, R.; Bulut, B.; Tuten, A.; Aydin, B.; Demirayak, G.; Kucur, M. First-trimester maternal serum metastin, placental growth factor and chitotriosidase levels in pre-eclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2012, 164, 146–149. [Google Scholar] [CrossRef]
- Matjila, M.; Millar, R.; van der Spuy, Z.; Katz, A. Elevated placental expression at the maternal-fetal interface but diminished maternal circulatory kisspeptin in preeclamptic pregnancies. Pregnancy Hypertens. 2016, 6, 79–87. [Google Scholar] [CrossRef]
- Armstrong, R.A.; Reynolds, R.M.; Leask, R.; Shearing, C.H.; Calder, A.A.; Riley, S.C. Decreased serum levels of kisspeptin in early pregnancy are associated with intra-uterine growth restriction and pre-eclampsia. Prenat. Diagn. 2009, 29, 982–985. [Google Scholar] [CrossRef]
- Qiao, C.; Wang, C.; Zhao, J.; Liu, C.; Shang, T. Elevated expression of KiSS-1 in placenta of Chinese women with early-onset preeclampsia. PLoS ONE 2012, 7, e48937. [Google Scholar] [CrossRef]
- Zhang, H.; Long, Q.; Ling, L.; Gao, A.; Li, H.; Lin, Q. Elevated expression of KiSS-1 in placenta of preeclampsia and its effect on trophoblast. Reprod. Biol. 2011, 11, 99–115. [Google Scholar] [CrossRef]
- Vazquez-Alaniz, F.; Galaviz-Hernandez, C.; Marchat, L.A.; Salas-Pacheco, J.M.; Chairez-Hernandez, I.; Guijarro-Bustillos, J.J.; Mireles-Ordaz, A. Comparative expression profiles for KiSS-1 and REN genes in preeclamptic and healthy placental tissues. Eur. J. Obstet. Gynecol. Reprod. Biol. 2011, 159, 67–71. [Google Scholar] [CrossRef]
- Ibanoglu, M.C.; Oskovi-Kaplan, Z.A.; Ozgu-Erdinc, A.S.; Kara, O.; Sahin, D. Comparison of the Kisspeptin levels in early onset preeclampsia and late-onset preeclampsia. Arch. Gynecol. Obs. 2022. ahead of print. [Google Scholar] [CrossRef]
- Fetal Growth Restriction: ACOG Practice Bulletin, Number 227. Obstet. Gynecol. 2021, 137, e16–e28. [CrossRef] [PubMed]
- Lees, C.C.; Stampalija, T.; Baschat, A.; da Silva Costa, F.; Ferrazzi, E.; Figueras, F.; Hecher, K.; Kingdom, J.; Poon, L.C.; Salomon, L.J.; et al. ISUOG Practice Guidelines: Diagnosis and management of small-for-gestational-age fetus and fetal growth restriction. Ultrasound Obstet. Gynecol. 2020, 56, 298–312. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, S.; Torbe, A.; Borowski, D.; Breborowicz, G.; Czajkowski, K.; Huras, H.; Kajdy, A.; Kalinka, J.; Kosinska-Kaczynska, K.; Leszczynska-Gorzelak, B.; et al. Polish Society of Gynecologists and Obstetricians Recommendations on diagnosis and management of fetal growth restriction. Ginekol. Pol. 2020, 91, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Zur, R.L.; Kingdom, J.C.; Parks, W.T.; Hobson, S.R. The Placental Basis of Fetal Growth Restriction. Obstet. Gynecol. Clin. N. Am. 2020, 47, 81–98. [Google Scholar] [CrossRef]
- Baschat, A.A. Planning management and delivery of the growth-restricted fetus. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 49, 53–65. [Google Scholar] [CrossRef]
- Logie, J.J.; Denison, F.C.; Riley, S.C.; Ramaesh, T.; Forbes, S.; Norman, J.E.; Reynolds, R.M. Evaluation of kisspeptin levels in obese pregnancy as a biomarker for pre-eclampsia. Clin. Endocrinol. 2012, 76, 887–893. [Google Scholar] [CrossRef]
- Smets, E.M.L.; Deurloo, K.L.; Go, A.T.J.I.; van Vugt, J.M.G.; Blankenstein, M.A.; Oudejans, C.B.M. Decreased plasma levels of metastin in early pregnancy are associated with small for gestational age neonates. Prenat. Diagn. 2008, 28, 299–303. [Google Scholar] [CrossRef]
- Kimyon Comert, G.; Esin, S.; Caglar, G.S.; Yirci, B.; Ozdemir, S.; Demirtas, S.; Kandemir, O. The correlation between birth weight and insulin-like growth factor-binding protein-1 (IGFBP-1), kisspeptin-1 (KISS-1), and three-dimensional fetal volume. J. Matern. Fetal Neonatal Med. 2019, 32, 2152–2158. [Google Scholar] [CrossRef]
- Vogel, J.P.; Chawanpaiboon, S.; Moller, A.-B.; Watananirun, K.; Bonet, M.; Lumbiganon, P. The global epidemiology of preterm birth. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 52, 3–12. [Google Scholar] [CrossRef]
- Arrowsmith, S.; Wray, S. Oxytocin: Its mechanism of action and receptor signalling in the myometrium. J. Neuroendocr. 2014, 26, 356–369. [Google Scholar] [CrossRef]
- Cecati, M.; Sartini, D.; Campagna, R.; Biagini, A.; Ciavattini, A.; Emanuelli, M.; Giannubilo, S.R. Molecular analysis of endometrial inflammation in preterm birth. Cell Mol. Biol. 2017, 63, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Uvnäs Moberg, K.; Handlin, L.; Kendall-Tackett, K.; Petersson, M. Oxytocin is a principal hormone that exerts part of its effects by active fragments. Med. Hypotheses 2019, 133, 109394. [Google Scholar] [CrossRef] [PubMed]
- Seymour, A.J.; Scott, V.; Augustine, R.A.; Bouwer, G.T.; Campbell, R.E.; Brown, C.H. Development of an excitatory kisspeptin projection to the oxytocin system in late pregnancy. J. Physiol. 2017, 595, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Torricelli, M.; Galleri, L.; Voltolini, C.; Biliotti, G.; Florio, P.; De Bonis, M.; Petraglia, F. Changes of placental Kiss-1 mRNA expression and maternal/cord kisspeptin levels at preterm delivery. Reprod. Sci. 2008, 15, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Nijher, G.M.K.; Chaudhri, O.B.; Ramachandran, R.; Murphy, K.G.; Zac-Varghese, S.E.K.; Fowler, A.; Chinthapalli, K.; Patterson, M.; Thompson, E.L.; Williamson, C.; et al. The effects of kisspeptin-54 on blood pressure in humans and plasma kisspeptin concentrations in hypertensive diseases of pregnancy. Br. J. Clin. Pharmacol. 2010, 70, 674–681. [Google Scholar] [CrossRef]
- Farina, A.; Sekizawa, A.; Purwosunu, Y.; Rizzo, N.; Banzola, I.; Concu, M.; Morano, D.; Giommi, F.; Bevini, M.; Mabrook, M.; et al. Quantitative distribution of a panel of circulating mRNA in preeclampsia versus controls. Prenat. Diagn. 2006, 26, 1115–1120. [Google Scholar] [CrossRef]
- Whitehead, C.L.; McNamara, H.; Walker, S.P.; Alexiadis, M.; Fuller, P.J.; Vickers, D.K.; Hannan, N.J.; Hastie, R.; Tuohey, L.; Kaitu’u-Lino, T.J.; et al. Identifying late-onset fetal growth restriction by measuring circulating placental RNA in the maternal blood at 28 weeks’ gestation. Am. J. Obstet. Gynecol. 2016, 214, 521.e1–521.e8. [Google Scholar] [CrossRef]
- Mark, P.J.; Jones, M.L.; Lewis, J.L.; Waddell, B.J.; Smith, J.T. Kiss1 and Kiss1r mRNA expression in the rat placenta: Changes with gestational age and regulation by glucocorticoids. Placenta 2013, 34, 657–662. [Google Scholar] [CrossRef]
- Whitehead, C.L.; Walker, S.P.; Ye, L.; Mendis, S.; Kaitu’u-Lino, T.J.; Lappas, M.; Tong, S. Placental specific mRNA in the maternal circulation are globally dysregulated in pregnancies complicated by fetal growth restriction. J. Clin. Endocrinol. Metab. 2013, 98, E429–E436. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szydełko-Gorzkowicz, M.; Poniedziałek-Czajkowska, E.; Mierzyński, R.; Sotowski, M.; Leszczyńska-Gorzelak, B. The Role of Kisspeptin in the Pathogenesis of Pregnancy Complications: A Narrative Review. Int. J. Mol. Sci. 2022, 23, 6611. https://doi.org/10.3390/ijms23126611
Szydełko-Gorzkowicz M, Poniedziałek-Czajkowska E, Mierzyński R, Sotowski M, Leszczyńska-Gorzelak B. The Role of Kisspeptin in the Pathogenesis of Pregnancy Complications: A Narrative Review. International Journal of Molecular Sciences. 2022; 23(12):6611. https://doi.org/10.3390/ijms23126611
Chicago/Turabian StyleSzydełko-Gorzkowicz, Magdalena, Elżbieta Poniedziałek-Czajkowska, Radzisław Mierzyński, Maciej Sotowski, and Bożena Leszczyńska-Gorzelak. 2022. "The Role of Kisspeptin in the Pathogenesis of Pregnancy Complications: A Narrative Review" International Journal of Molecular Sciences 23, no. 12: 6611. https://doi.org/10.3390/ijms23126611
APA StyleSzydełko-Gorzkowicz, M., Poniedziałek-Czajkowska, E., Mierzyński, R., Sotowski, M., & Leszczyńska-Gorzelak, B. (2022). The Role of Kisspeptin in the Pathogenesis of Pregnancy Complications: A Narrative Review. International Journal of Molecular Sciences, 23(12), 6611. https://doi.org/10.3390/ijms23126611