
Baicalin Alleviates Short-Term Lincomycin-Induced Intestinal and Liver Injury and Inflammation in Infant Mice




Abstract
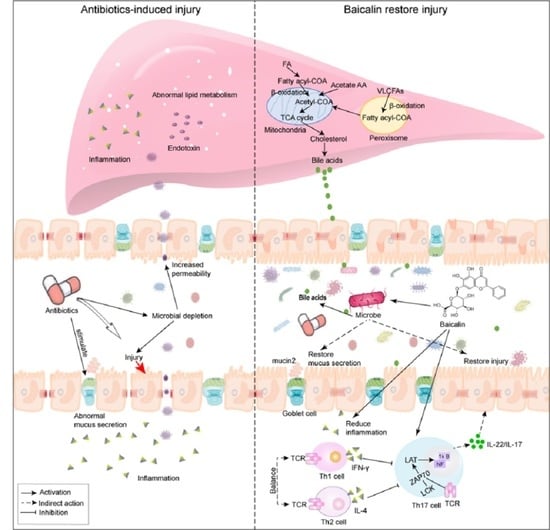
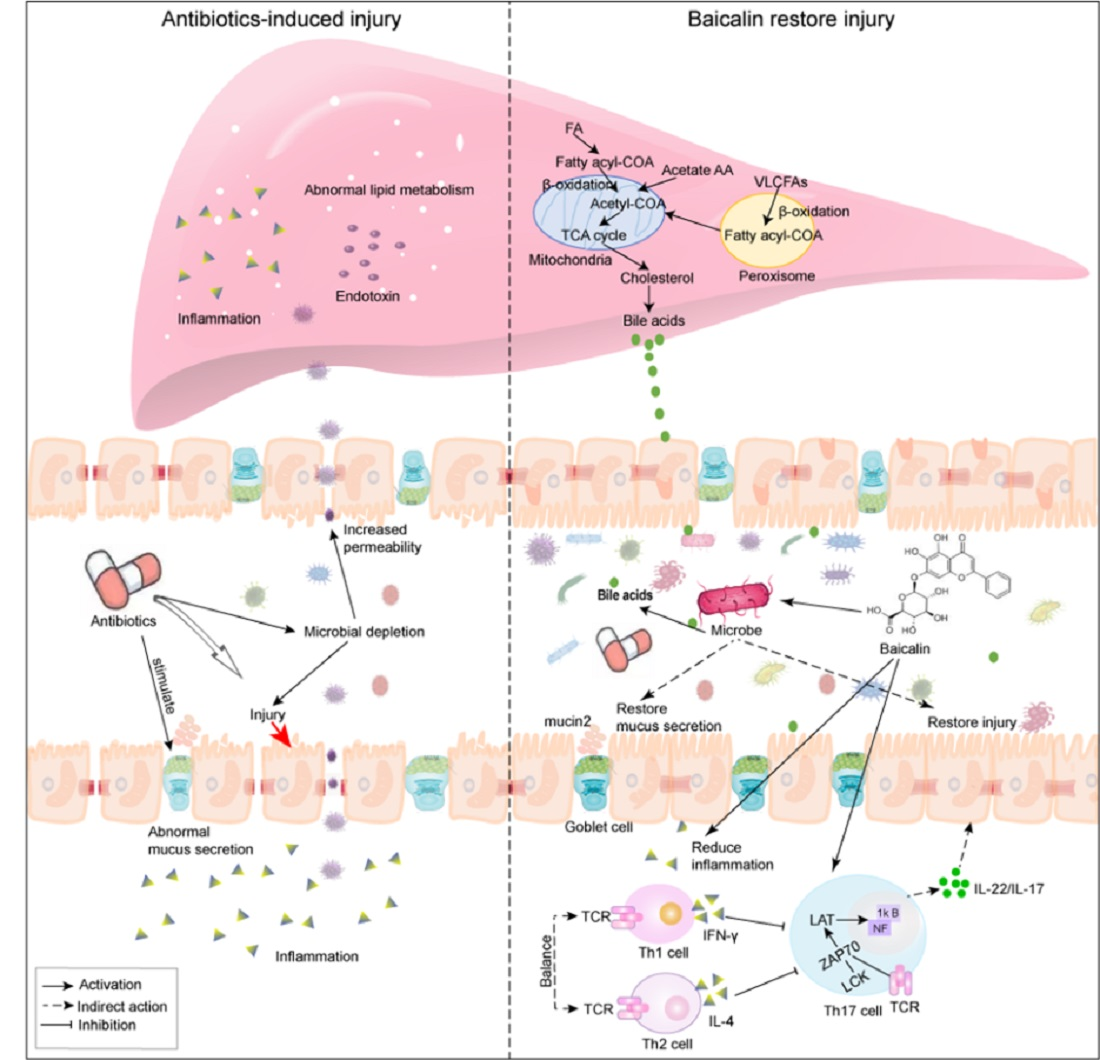
1. Introduction
2. Results
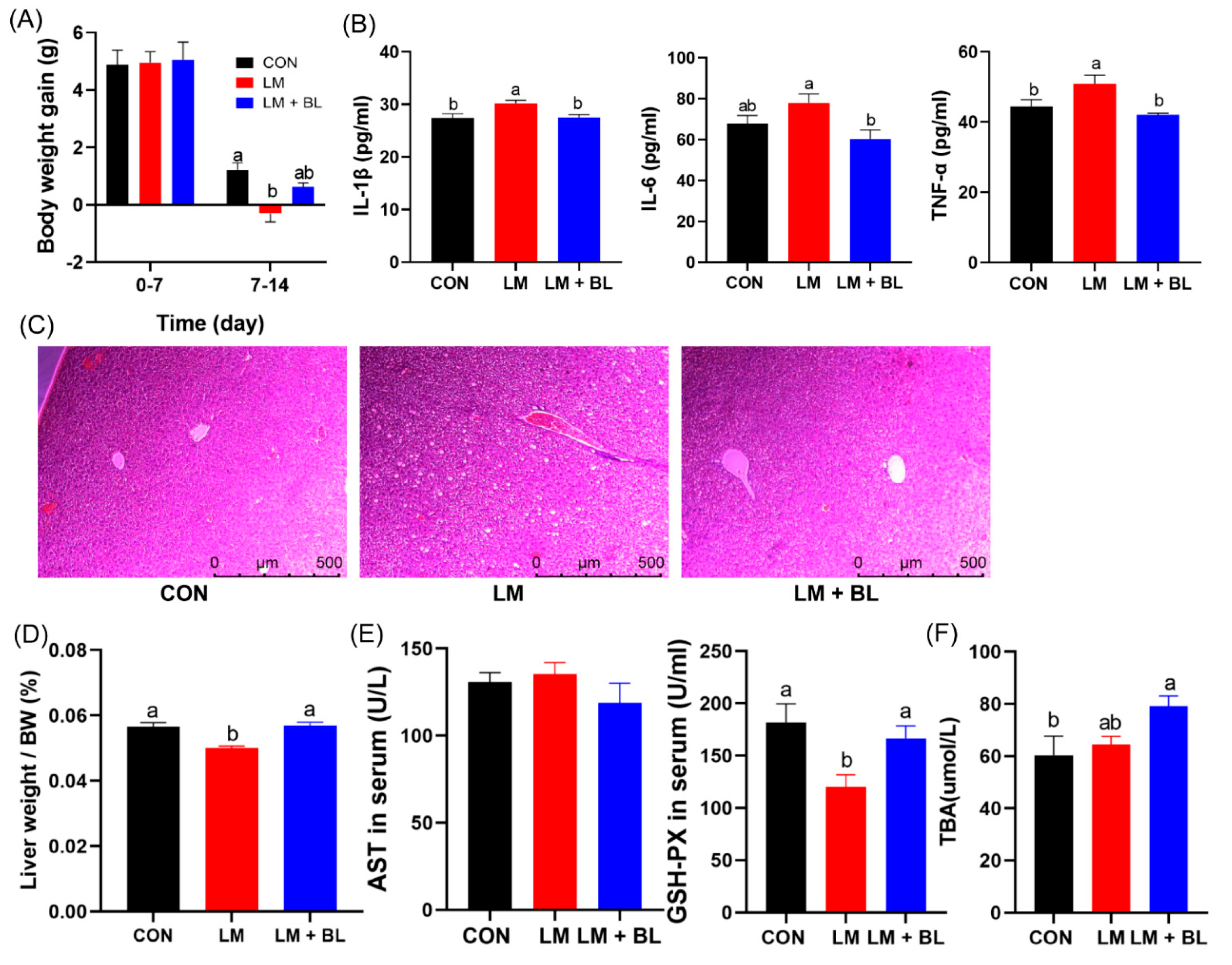
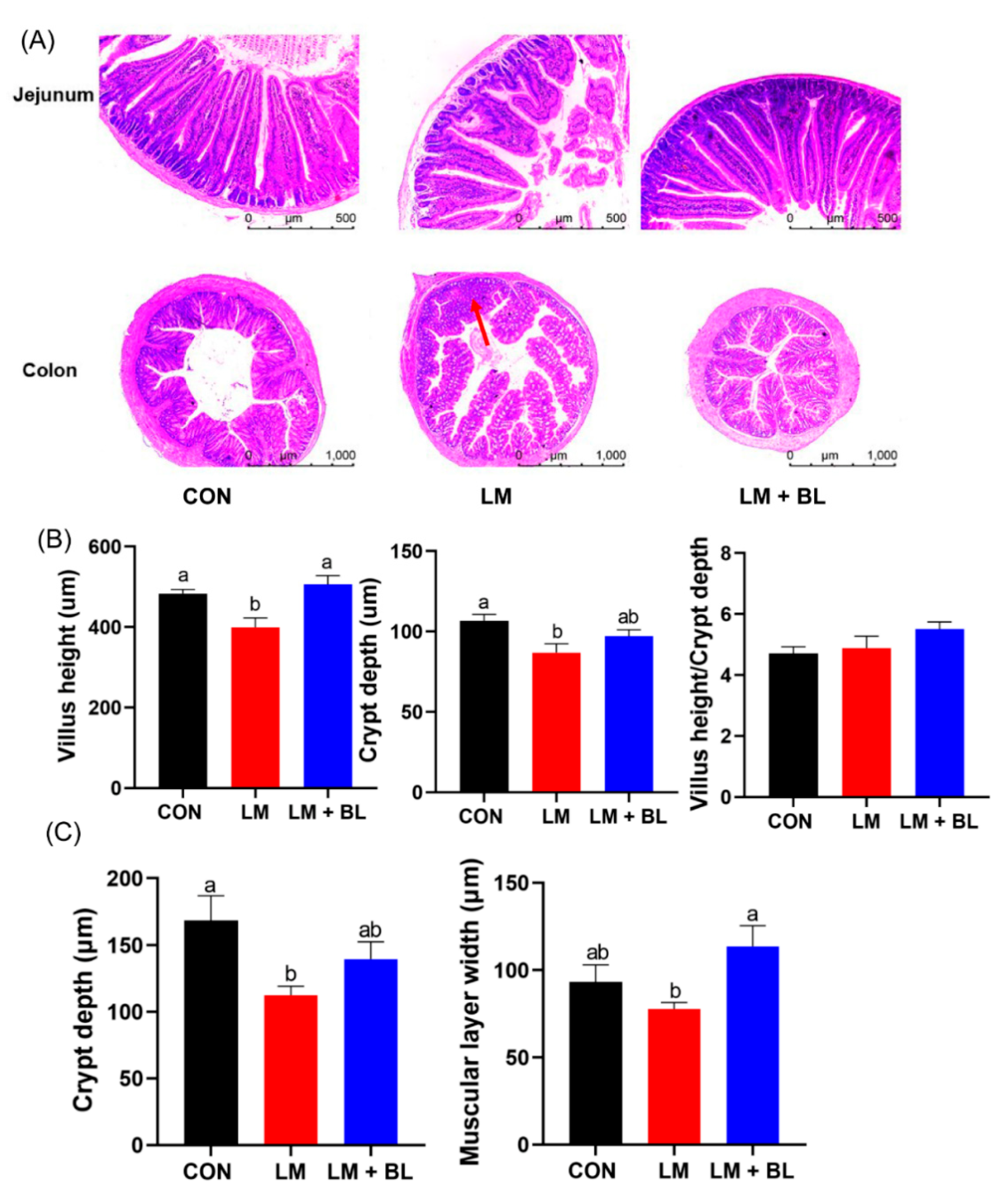
2.1. Growth Performance and Intestinal and Liver Morphologies
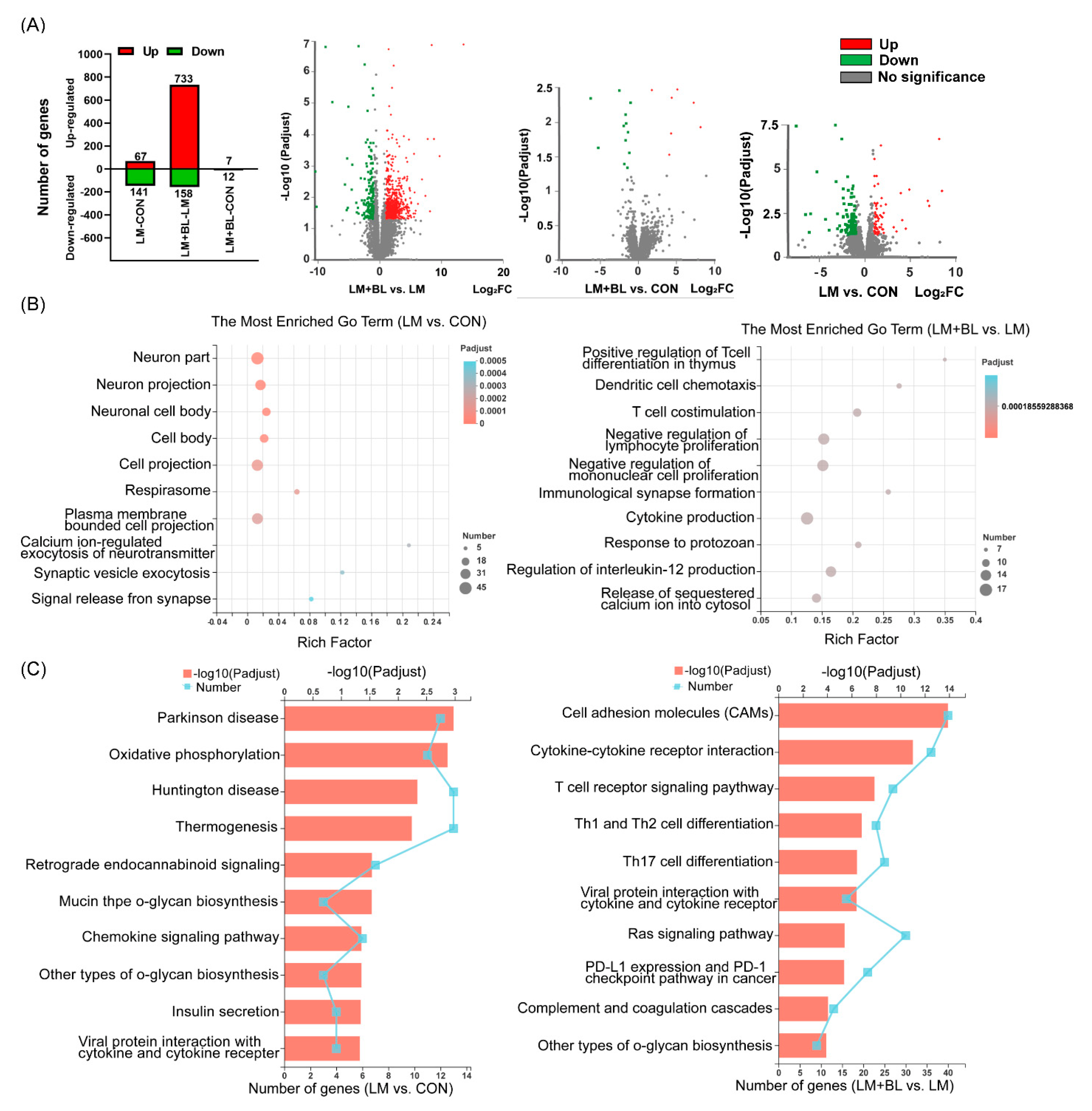
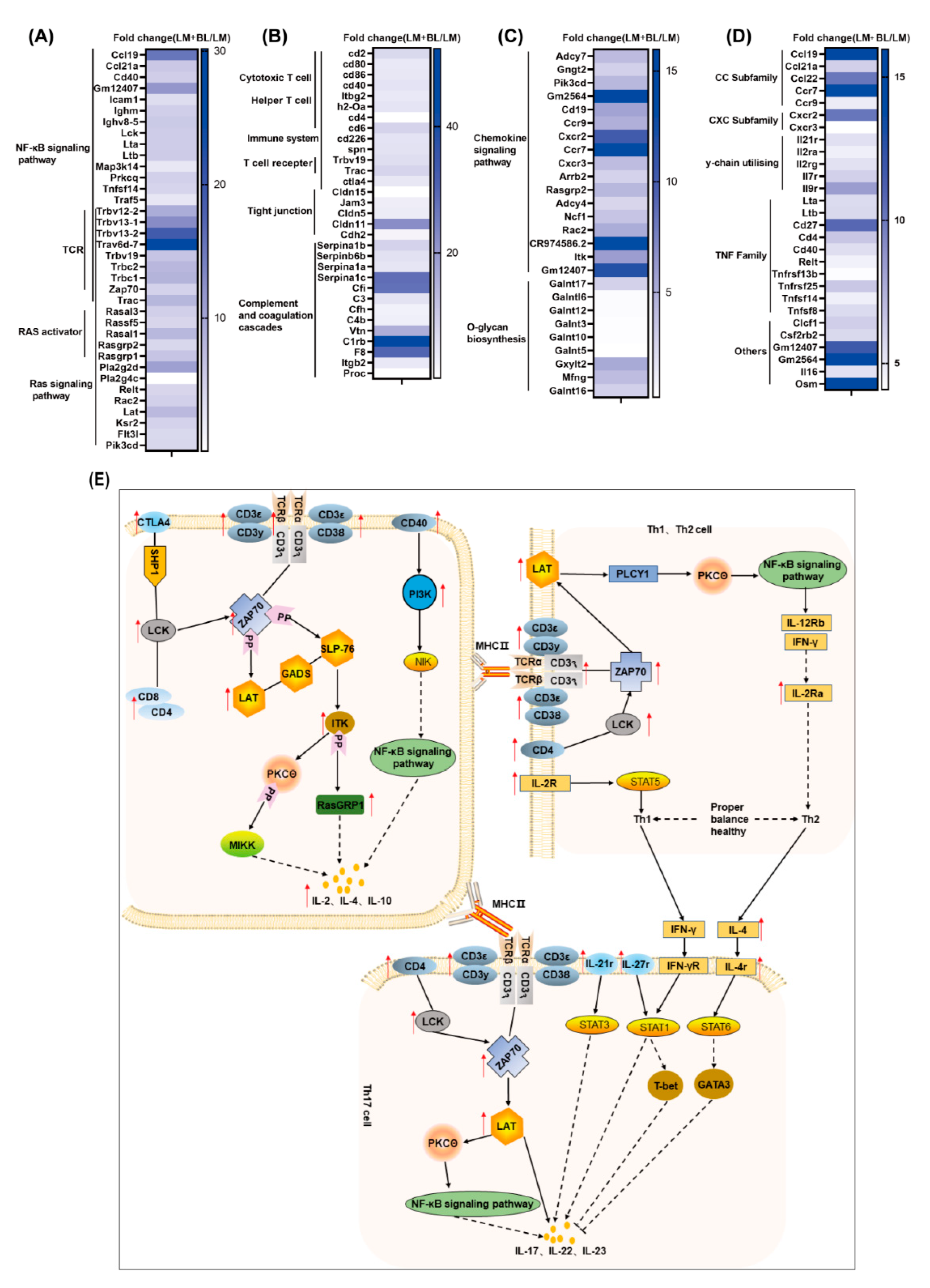
2.2. Colon Transcriptome
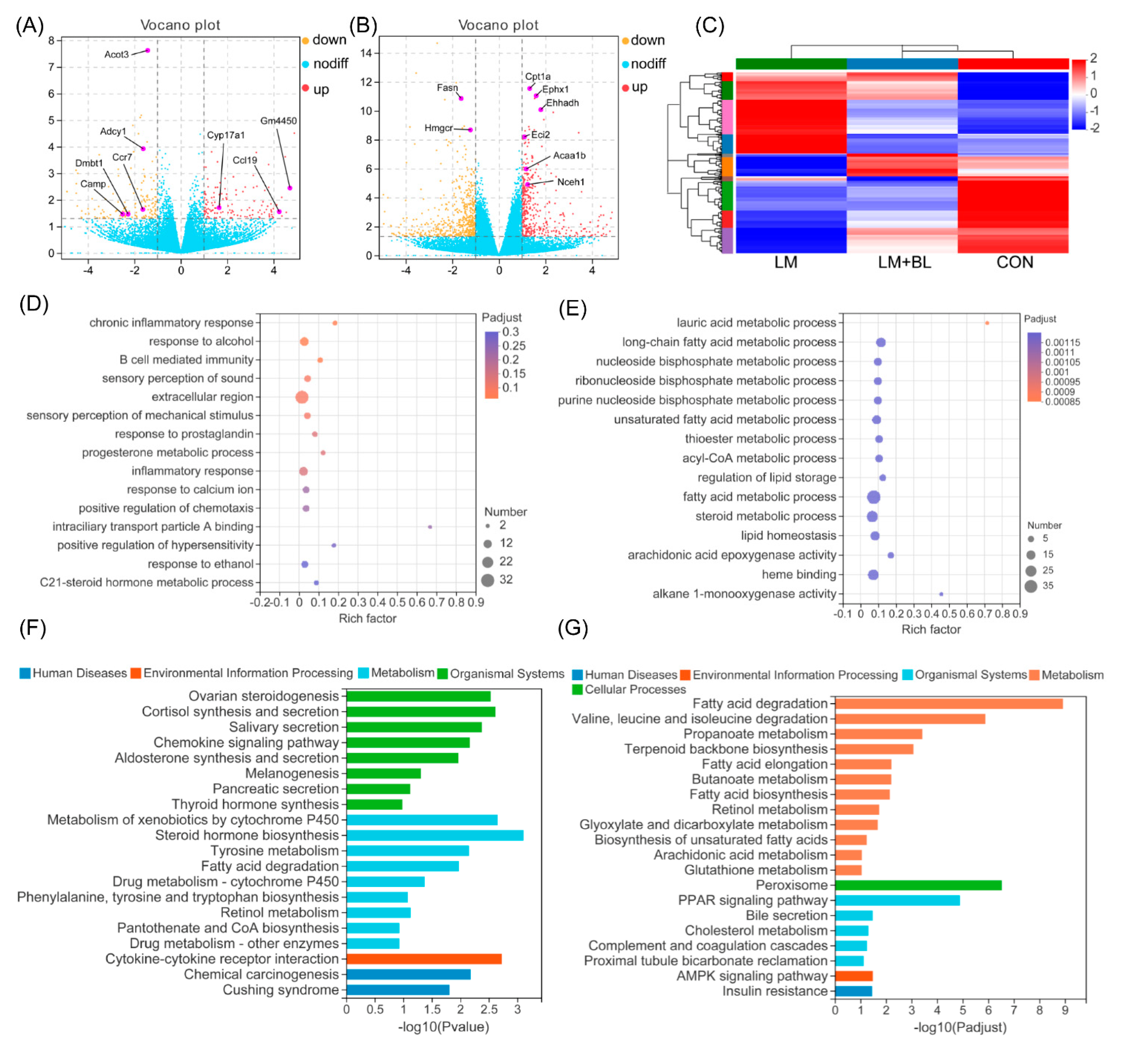
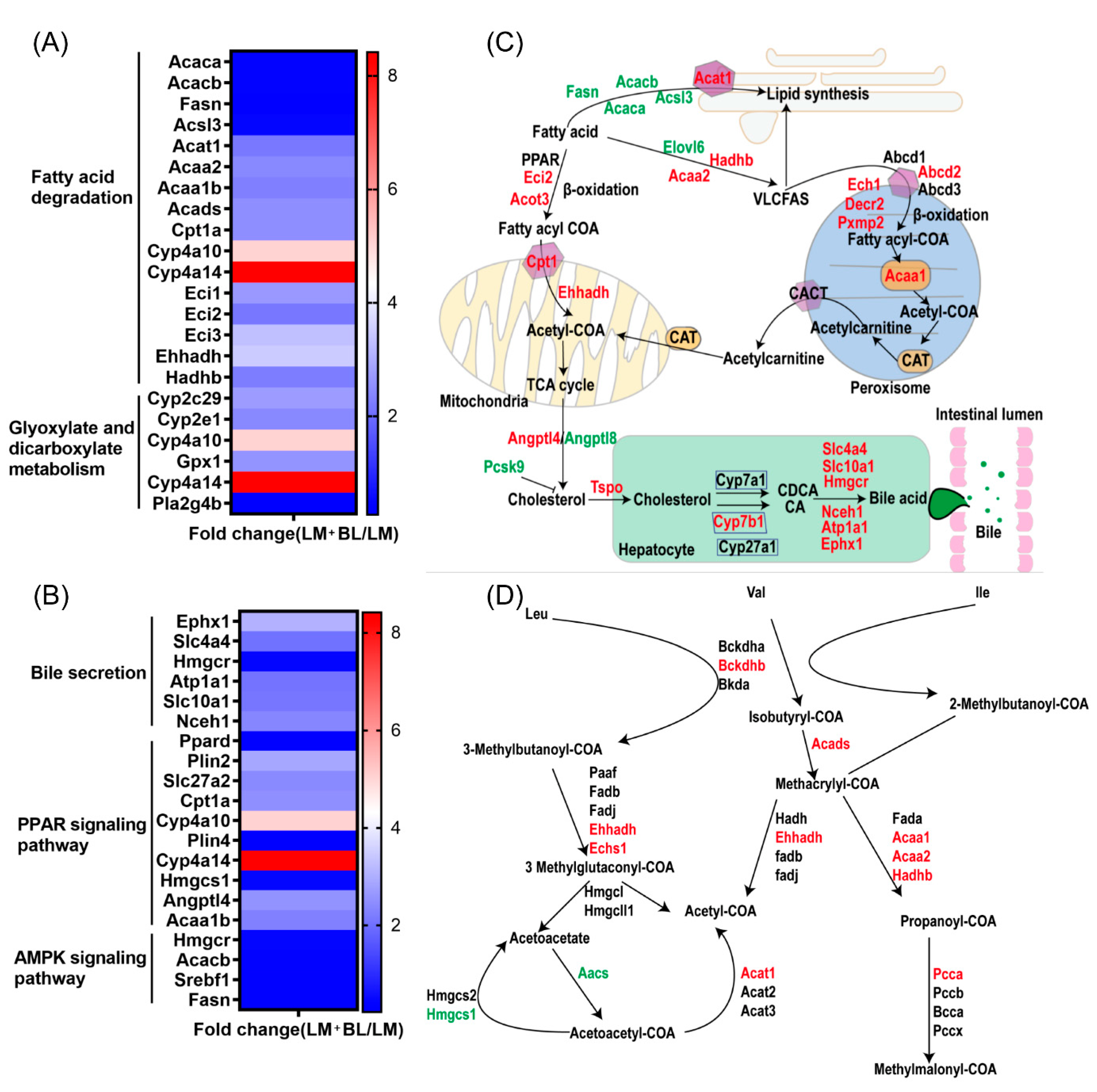
2.3. Liver Transcriptome
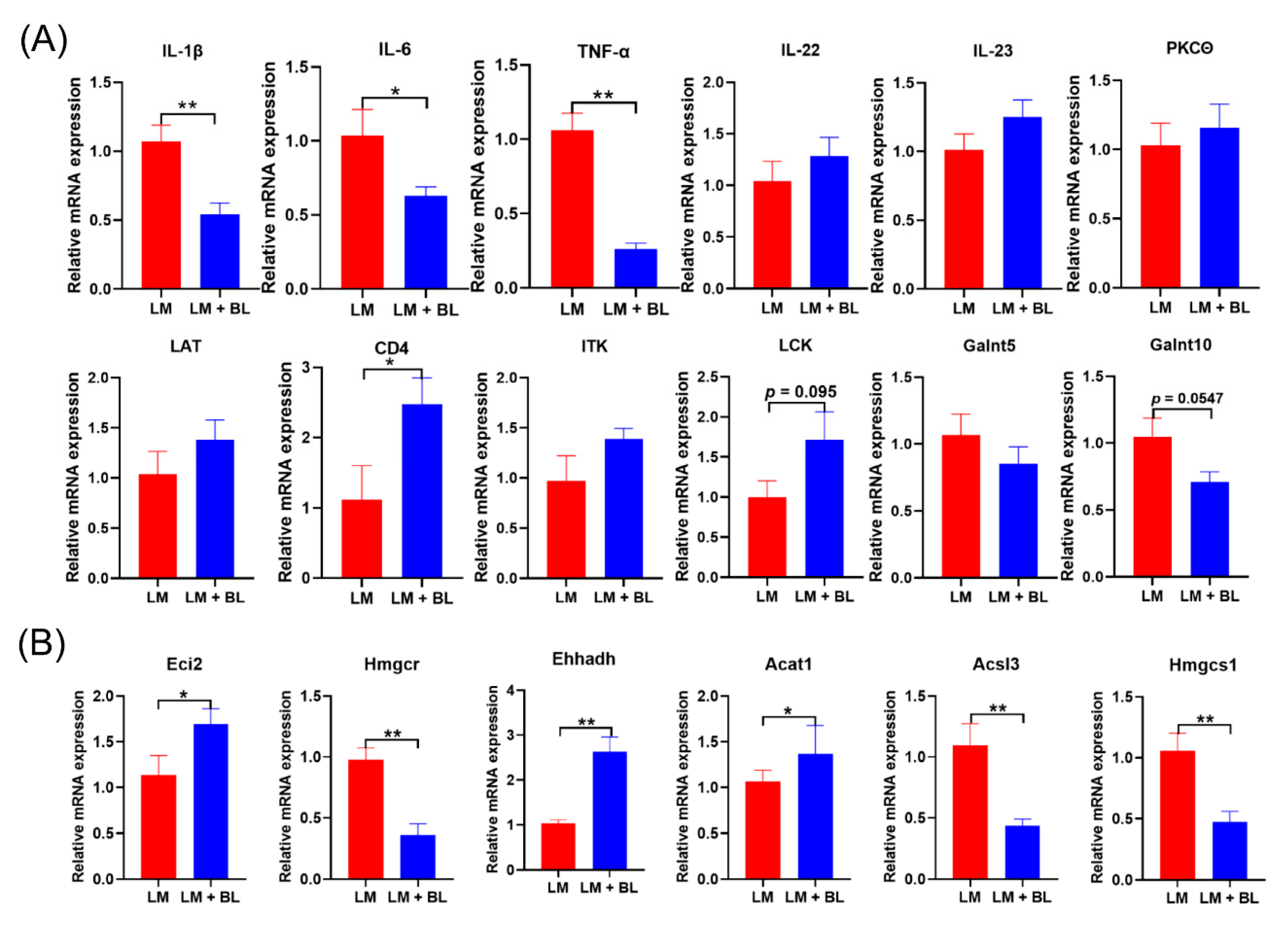
2.4. Validation of the RNA-Seq Results by qPCR
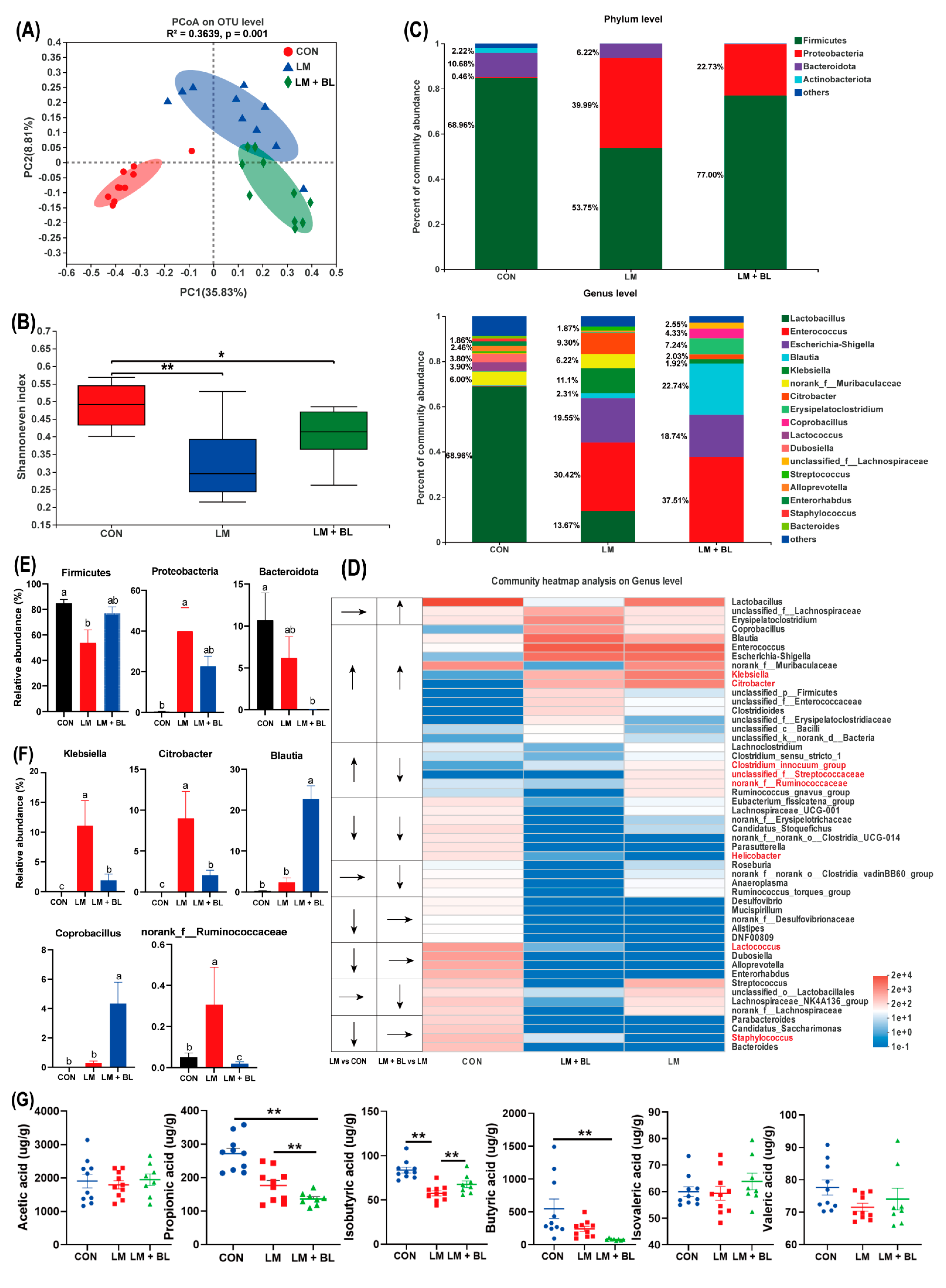
2.5. Colon Microbiome
3. Discussion
4. Materials and Methods
4.1. Mice and Treatment
4.2. 16S rRNA Gene Sequencing Analysis
4.3. Transcriptome Analysis
4.4. Relative Concentrations of Short-Chain Fatty Acids
4.5. Intestinal and Hepatic Morphology
4.6. Gene Expression Related to Tight Junction Proteins and Inflammatory Cytokines
4.7. Inflammatory Cytokines and GSH-PX, AST in Serum, and TBA in Liver
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maier, L.; Goemans, C.V.; Wirbel, J.; Kuhn, M.; Eberl, C.; Pruteanu, M.; Müller, P.; Garcia-Santamarina, S.; Cacace, E.; Zhang, B.; et al. Unravelling the collateral damage of antibiotics on gut bacteria. Nature 2021, 599, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Koliarakis, I.; Athanasakis, E.; Sgantzos, M.; Mariolis-Sapsakos, T.; Xynos, E.; Chrysos, E.; Souglakos, J.; Tsiaoussis, J. Intestinal Microbiota in Colorectal Cancer Surgery. Cancers 2020, 12, 3011. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Du, W.; Xu, J. Retrospective analysis of adverse reactions of lincomycin. Chin. J. New Drugs Clin. Remedies 2003, 22, 571–574. [Google Scholar]
- Skirecki, T.; Borkowska-Zielinska, U.; Zlotorowicz, M.; Hoser, G. Sepsis Immunopathology: Perspectives of Monitoring and Modulation of the Immune Disturbances. Arch. Immunol. Ther. Exp. 2012, 60, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhong, R.; Han, H.; Yi, B.; Yin, J.; Chen, L.; Zhang, H. Short-Term Lincomycin Exposure Depletion of Murine Microbiota Affects Short-Chain Fatty Acids and Intestinal Morphology and Immunity. Antibiotics 2020, 9, 907. [Google Scholar] [CrossRef]
- Ferrajolo, C.; Capuano, A.; Verhamme, K.M.C.; Schuemie, M.; Rossi, F.; Stricker, B.H.; Sturkenboom, M.C.J.M. Drug-induced hepatic injury in children: A case/non-case study of suspected adverse drug reactions in VigiBase. Br. J. Clin. Pharmacol. 2010, 70, 721–728. [Google Scholar] [CrossRef]
- Andrade, R.J.; Tulkens, P.M. Hepatic safety of antibiotics used in primary care. J. Antimicrob. Chemother. 2011, 66, 1431–1446. [Google Scholar] [CrossRef]
- Ferrajolo, C.; Verhamme, K.M.C.; Trifirò, G.; Jong, G.W.; Picelli, G.; Giaquinto, C.; Mazzaglia, G.; Stricker, B.H.; Rossi, F.; Capuano, A.; et al. Antibiotic-Induced Liver Injury in Paediatric Outpatients: A Case-Control Study in Primary Care Databases. Drug Saf. 2016, 40, 305–315. [Google Scholar] [CrossRef]
- Guo, W.; Zhou, X.; Li, X.; Zhu, Q.; Peng, J.; Zhu, B.; Zheng, X.; Lu, Y.; Yang, D.; Wang, B.; et al. Depletion of Gut Microbiota Impairs Gut Barrier Function and Antiviral Immune Defense in the Liver. Front. Immunol. 2021, 12, 897. [Google Scholar] [CrossRef]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J. Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef]
- Liao, P.; Li, Y.; Li, M.; Chen, X.; Yuan, D.; Tang, M.; Xu, K. Baicalin alleviates deoxynivalenol-induced intestinal inflammation and oxidative stress damage by inhibiting NF-κB and increasing mTOR signaling pathways in piglets. Food Chem. Toxicol. 2020, 140, 111326. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Xu, L.-Z.; Zhao, S.; Shen, Z.-F.; Shen, H.; Zhan, L.-B. Protective effect of baicalin on the regulation of Treg/Th17 balance, gut microbiota and short-chain fatty acids in rats with ulcerative colitis. Appl. Microbiol. Biotechnol. 2020, 104, 5449–5460. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Xiao, Z.; Yan, H.; Lu, X.; Zhang, X.; Luo, L.; Long, C.; Zhu, Y. Baicalin suppresses Th1 and Th17 responses and promotes Treg response to ameliorate sepsis-associated pancreatic injury via the RhoA-ROCK pathway. Int. Immunopharmacol. 2020, 86, 106685. [Google Scholar] [CrossRef]
- Yang, J.-Y.; Li, M.; Zhang, C.-L.; Liu, D. Pharmacological properties of baicalin on liver diseases: A narrative review. Pharmacol. Rep. 2021, 73, 1230–1239. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Liu, T.; Xing, L.; Ji, G. Baicalin and puerarin reverse epithelial-mesenchymal transition via the TGF-β1/Smad3 pathway in vitro. Exp. Ther. Med. 2018, 16, 1968–1974. [Google Scholar] [CrossRef]
- Miao, Z.; Cheng, R.; Zhang, Y.; Liang, H.; Jiang, F.; Shen, X.; Chen, G.; Zhang, Q.; He, F.; Li, M. Antibiotics can cause weight loss by impairing gut microbiota in mice and the potent benefits of lactobacilli. Biosci. Biotechnol. Biochem. 2019, 84, 411–420. [Google Scholar] [CrossRef]
- Wang, L.-S.; Wang, J.-Y.; Jian-Yao, W.; Cao, X.; Zhang, R.; Li-Sheng, W.; Xu, Z.-L.; Zhang, D.-G. Protective effect of baicalin against experimental colitis via suppression of oxidant stress and apoptosis. Pharmacogn. Mag. 2016, 12, 225–234. [Google Scholar] [CrossRef]
- Hunter, D.J. Pharmacologic therapy for osteoarthritis—the era of disease modification. Nat. Rev. Rheumatol. 2010, 7, 13–22. [Google Scholar] [CrossRef]
- Alonso, M.N.; Wong, M.T.; Zhang, A.L.; Winer, D.; Suhoski, M.M.; Tolentino, L.L.; Gaitan, J.; Davidson, M.G.; Kung, T.H.; Galel, D.M.; et al. TH1, TH2, and TH17 cells instruct monocytes to differentiate into specialized dendritic cell subsets. Blood 2011, 118, 3311–3320. [Google Scholar] [CrossRef]
- Nakagawa, T.; Hirano, T.; Nakagawa, N.; Yoshizaki, K.; Kishimoto, T. Effect of recombinant IL 2 and gamma-IFN on proliferation and differentiation of human B cells. J. Immunol. 1985, 134, 959–966. [Google Scholar]
- Osborne, L.C.; A Duthie, K.; Seo, J.H.; Gascoyne, R.D.; Abraham, N. Selective ablation of the YxxM motif of IL-7Rα suppresses lymphomagenesis but maintains lymphocyte development. Oncogene 2010, 29, 3854–3864. [Google Scholar] [CrossRef] [PubMed]
- Truong, A.D.; Hong, Y.H.; Lillehoj, H.S. RNA-seq Profiles of Immune Related Genes in the Spleen of Necrotic Enteritis-afflicted Chicken Lines. Asian-Australasian J. Anim. Sci. 2015, 28, 1496–1511. [Google Scholar] [CrossRef] [PubMed]
- Holmes, T.D.; Wilson, E.B.; Black, E.V.I.; Benest, A.V.; Vaz, C.; Tan, B.; Tanavde, V.M.; Cook, G.P. Licensed human natural killer cells aid dendritic cell maturation via TNFSF14/LIGHT. Proc. Natl. Acad. Sci. USA 2014, 111, E5688–E5696. [Google Scholar] [CrossRef] [PubMed]
- Valatas, V.; Kolios, G.; Bamias, G. TL1A (TNFSF15) and DR3 (TNFRSF25): A Co-stimulatory System of Cytokines With Diverse Functions in Gut Mucosal Immunity. Front. Immunol. 2019, 10, 583. [Google Scholar] [CrossRef]
- Fava, V.M.; Cobat, A.; Van Thuc, N.; Latini, A.C.P.; Stefani, M.M.A.; Belone, A.F.; Ba, N.N.; Orlova, M.; Manry, J.; Mira, M.; et al. Association of TNFSF8 Regulatory Variants With Excessive Inflammatory Responses but not Leprosy Per Se. J. Infect. Dis. 2014, 211, 968–977. [Google Scholar] [CrossRef]
- von Andrian, U.H.; Chambers, J.D.; McEvoy, L.M.; Bargatze, R.F.; E Arfors, K.; Butcher, E.C. Two-step model of leukocyte-endothelial cell interaction in inflammation: Distinct roles for LECAM-1 and the leukocyte beta 2 integrins in vivo. Proc. Natl. Acad. Sci. USA 1991, 88, 7538–7542. [Google Scholar] [CrossRef]
- Tian, D.; Li, Y.; Li, X.; Tian, Z. Aloperine inhibits proliferation, migration and invasion and induces apoptosis by blocking the Ras signaling pathway in human breast cancer cells. Mol. Med. Rep. 2018, 18, 3699–3710. [Google Scholar] [CrossRef]
- Abdel-Salam, B.K.A.-H. Comparing effects of interleukin-2 and interleukin-4 on the expression of MHC class II, CD80 and CD86 on polymorphonuclear neutrophils. Egypt. J. Med. Hum. Genet. 2010, 11, 115–120. [Google Scholar] [CrossRef]
- Urban, D.; Thanabalasingam, U.; Stibenz, D.; Kaufmann, J.; Meyborg, H.; Fleck, E.; Gräfe, M.; Stawowy, P. CD40/CD40L interaction induces E-selectin dependent leukocyte adhesion to human endothelial cells and inhibits endothelial cell migration. Biochem. Biophys. Res. Commun. 2011, 404, 448–452. [Google Scholar] [CrossRef]
- Rosenthal, R.; Milatz, S.; Krug, S.M.; Oelrich, B.; Schulzke, J.-D.; Amasheh, S.; Günzel, D.; Fromm, M. Claudin-2, a component of the tight junction, forms a paracellular water channel. J. Cell Sci. 2010, 123, 1913–1921. [Google Scholar] [CrossRef]
- Zeissig, S.; Bürgel, N.; Günzel, D.; Richter, J.; Mankertz, J.; Wahnschaffe, U.; Kroesen, A.J.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Changes in expression and distribution of claudin 2, 5 and 8 lead to discontinuous tight junctions and barrier dysfunction in active Crohn’s disease. Gut 2007, 56, 61–72. [Google Scholar] [CrossRef] [PubMed]
- El-Husseini, A.A.; Sheashaa, H.A.; Sabry, A.A.; Moustafa, F.E.; Sobh, M.A. Acute Postinfectious Crescentic Glomerulonephritis: Clinicopathologic Presentation and Risk Factors. Int. Urol. Nephrol. 2005, 37, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Kremer, V.; Ligtenberg, M.A.; Zendehdel, R.; Seitz, C.; Duivenvoorden, A.; Wennerberg, E.; Colón, E.; Scherman-Plogell, A.-H.; Lundqvist, A. Genetic engineering of human NK cells to express CXCR2 improves migration to renal cell carcinoma. J. Immunother. Cancer 2017, 5, 73. [Google Scholar] [CrossRef] [PubMed]
- Pageon, S.V.; Tabarin, T.; Yamamoto, Y.; Ma, Y.; Nicovich, P.R.; Bridgeman, J.S.; Cohnen, A.; Benzing, C.; Gao, Y.; Crowther, M.D.; et al. Functional role of T-cell receptor nanoclusters in signal initiation and antigen discrimination. Proc. Natl. Acad. Sci. USA 2016, 113, E5454–E5463. [Google Scholar] [CrossRef]
- Wang, H.; Zeng, X.; Fan, Z.; Lim, B. RhoH modulates pre-TCR and TCR signalling by regulating LCK. Cell. Signal. 2011, 23, 249–258. [Google Scholar] [CrossRef]
- Shimamura, M.; Huang, Y.-Y.; Kobayashi, M.; Goji, H. Altered production of immunoregulatory cytokines by invariant V 19 TCR-bearing cells dependent on the duration and intensity of TCR engagement. Int. Immunol. 2008, 21, 179–185. [Google Scholar] [CrossRef]
- Zhao, J.-L.; Wang, X.; Wang, Y.-S. Relationships between Th1/Th2 cytokine profiles and chest radiographic manifestations in childhood Mycoplasma pneumoniae pneumonia. Ther. Clin. Risk Manag. 2016, 12, 1683–1692. [Google Scholar] [CrossRef]
- Loria-Cervera, E.N.; Sosa-Bibiano, E.I.; Villanueva-Lizama, L.E.; Van Wynsberghe, N.R.; Schountz, T.; Andrade-Narvaez, F.J. Cloning and sequence analysis of Peromyscus yucatanicus (Rodentia) Th1 (IL-12p35, IFN-γ and TNF) and Th2 (IL-4, IL-10 and TGF-β) cytokines. Cytokine 2014, 65, 48–55. [Google Scholar] [CrossRef]
- Mejía, D.; Salvatierra, G.; Maximiliano, J.; Rímac, R.; Carhuaricra, D.; Almeyda, M.; Luna, L.; Rosadio, R.; Maturrano, L. Expresión de citoquinas Th1 (IL-2, IL-12, IFN-γ, TNF-α), Th2 (IL-4, IL-10, TGF-β) y Th17 (IL-17) en linfocitos circulantes de cuyes inoculados con una cepa de campo de Salmonella Typhimurium. Rev. De Investig. Vet. Del Perú 2019, 30, 1750–1761. [Google Scholar] [CrossRef]
- Schulze-Koops, H.; Lipsky, P.E.; Davis, L.S. Human memory T cell differentiation into Th2-like effector cells is dependent on IL-4 and CD28 stimulation and inhibited by TCR ligation. Eur. J. Immunol. 1998, 28, 2517–2529. [Google Scholar] [CrossRef]
- Torres, K.C.; Dutra, W.O.; Gollob, K.J. Endogenous IL-4 and IFN-γ are essential for expression of Th2, but not Th1 cytokine message during the early differentiation of human CD4+ T helper cells. Hum. Immunol. 2004, 65, 1328–1335. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lin, X.; Liu, X.; Ma, Z.; Li, Y. Baicalin regulates Treg/Th17 cell imbalance by inhibiting autophagy in allergic rhinitis. Mol. Immunol. 2020, 125, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.R.; Berthoud, T.K.; Kumar, A.; Angel, J.B. IL-23 signaling in Th17 cells is inhibited by HIV infection and is not restored by HAART: Implications for persistent immune activation. PLoS ONE 2017, 12, e0186823. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Cui, Y.; Shen, X.; Wang, S.; Yang, G.-B. IL-17A and IL-17F repair HIV-1 gp140 damaged Caco-2 cell barriers by upregulating tight junction genes. Microbes Infect. 2019, 21, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Feng, D.; Wang, H.; Hong, F.; Bertola, A.; Wang, F.-S.; Gao, B. Interleukin-22 induces hepatic stellate cell senescence and restricts liver fibrosis in mice. Hepatology 2012, 56, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. The Intestinal Crypt, A Prototype Stem Cell Compartment. Cell 2013, 154, 274–284. [Google Scholar] [CrossRef]
- Van der Flier, L.G.; Clevers, H. Stem Cells, Self-Renewal, and Differentiation in the Intestinal Epithelium. Annu. Rev. Physiol. 2009, 71, 241–260. [Google Scholar] [CrossRef]
- Barker, N.; Clevers, H. Tracking Down the Stem Cells of the Intestine: Strategies to Identify Adult Stem Cells. Gastroenterology 2007, 133, 1755–1760. [Google Scholar] [CrossRef]
- Thin, L.; Lawrance, I.; Spilsbury, K.; Kava, J.; Olynyk, J. Detection of liver injury in IBD using transient elastography. J. Crohn’s Colitis 2014, 8, 671–677. [Google Scholar] [CrossRef]
- Ooi, K.; Shiraki, K.; Sakurai, Y.; Morishita, Y.; Nobori, T. Clinical significance of abnormal lipoprotein patterns in liver diseases. Int. J. Mol. Med. 2005, 15, 655–660. [Google Scholar] [CrossRef]
- Benedetti, E.; D’Angelo, B.; Cristiano, L.; Di Giacomo, E.; Fanelli, F.; Moreno, S.; Cecconi, F.; Fidoamore, A.; Antonosante, A.; Falcone, R.; et al. Involvement of peroxisome proliferator-activated receptor β/δ (PPAR β/δ) in BDNF signaling during aging and in Alzheimer disease: Possible role of 4-hydroxynonenal (4-HNE). Cell Cycle 2014, 13, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Feng, J.; Jiang, D.; Zhou, X.; Jiang, Q.; Cai, M.; Wang, X.; Shan, T.; Wang, Y. AMPK regulates lipid accumulation in skeletal muscle cells through FTO-dependent demethylation of N6-methyladenosine. Sci. Rep. 2017, 7, 41606. [Google Scholar] [CrossRef] [PubMed]
- Chaumontet, C.; Even, P.C.; Schwarz, J.; Simonin-Foucault, A.; Piedcoq, J.; Fromentin, G.; Azzout-Marniche, D.; Tomé, D. High dietary protein decreases fat deposition induced by high-fat and high-sucrose diet in rats. Br. J. Nutr. 2015, 114, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Migita, T.; Takayama, K.; Urano, T.; Obinata, D.; Ikeda, K.; Soga, T.; Takahashi, S.; Inoue, S. ACSL3 promotes intratumoral steroidogenesis in prostate cancer cells. Cancer Sci. 2017, 108, 2011–2021. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Liang, K.; Zhao, S.; Jia, W.; Liu, Y.; Wu, H.; Lv, J.; Cao, C.; Chen, T.; Zhuang, S.; et al. Chemoproteomics reveals baicalin activates hepatic CPT1 to ameliorate diet-induced obesity and hepatic steatosis. Proc. Natl. Acad. Sci. USA 2018, 115, E5896–E5905. [Google Scholar] [CrossRef]
- Campbell, C.; McKenney, P.T.; Konstantinovsky, D.; Isaeva, O.; Schizas, M.; Verter, J.; Mai, C.; Jin, W.-B.; Guo, C.-J.; Violante, S.; et al. Bacterial metabolism of bile acids promotes generation of peripheral regulatory T cells. Nature 2020, 581, 475–479. [Google Scholar] [CrossRef]
- Hang, S.; Paik, D.; Yao, L.; Kim, E.; Trinath, J.; Lu, J.; Ha, S.; Nelson, B.N.; Kelly, S.P.; Wu, L.; et al. Bile acid metabolites control TH17 and Treg cell differentiation. Nature 2019, 576, 143–148. [Google Scholar] [CrossRef]
- Thibaut, M.M.; Bindels, L.B. Crosstalk between bile acid-activated receptors and microbiome in entero-hepatic inflammation. Trends Mol. Med. 2022, 28, 223–236. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef]
- Quraishi, M.N.; Acharjee, A.; Beggs, A.D.; Horniblow, R.; Tselepis, C.; Gkoutos, G.; Ghosh, S.; E Rossiter, A.; Loman, N.; van Schaik, W.; et al. A Pilot Integrative Analysis of Colonic Gene Expression, Gut Microbiota, and Immune Infiltration in Primary Sclerosing Cholangitis-Inflammatory Bowel Disease: Association of Disease With Bile Acid Pathways. J. Crohn’s Colitis 2020, 14, 935–947. [Google Scholar] [CrossRef]
- Sinha, S.R.; Haileselassie, Y.; Nguyen, L.P.; Tropini, C.; Wang, M.; Becker, L.S.; Sim, D.; Jarr, K.; Spear, E.T.; Singh, G.; et al. Dysbiosis-Induced Secondary Bile Acid Deficiency Promotes Intestinal Inflammation. Cell Host Microbe 2020, 27, 659–+. [Google Scholar] [CrossRef]
- Song, X.; Sun, X.; Oh, S.F.; Wu, M.; Zhang, Y.; Zheng, W.; Geva-Zatorsky, N.; Jupp, R.; Mathis, D.; Benoist, C.; et al. Microbial bile acid metabolites modulate gut RORγ+ regulatory T cell homeostasis. Nature 2020, 577, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zong, X.; Xiao, X.; Cheng, Y.; Fu, J.; Lu, Z.; Jin, M.; Wang, F.; Wang, Y. Multi-Omics Analysis of the Gut-Liver Axis Reveals the Mechanism of Liver Injury in Colitis Mice. Front. Immunol. 2022, 12, 773070. [Google Scholar] [CrossRef] [PubMed]
- Ueyama, J.; Nadai, M.; Kanazawa, H.; Iwase, M.; Nakayama, H.; Hashimoto, K.; Yokoi, T.; Baba, K.; Takagi, K.; Takagi, K.; et al. Endotoxin from various gram-negative bacteria has differential effects on function of hepatic cytochrome P450 and drug transporters. Eur. J. Pharmacol. 2005, 510, 127–134. [Google Scholar] [CrossRef]
- Koroleva, E.P.; Halperin, S.; Gubernatorova, E.O.; Macho-Fernandez, E.; Spencer, C.M.; Tumanov, A.V. Citrobacter rodentium -induced colitis: A robust model to study mucosal immune responses in the gut. J. Immunol. Methods 2015, 421, 61–72. [Google Scholar] [CrossRef]
- Youn, Y.; Lee, S.W.; Cho, H.-H.; Park, S.; Chung, H.-S.; Seo, J.W. Antibiotics-Associated Hemorrhagic Colitis Caused by Klebsiella oxytoca: Two Case Reports. Pediatr. Gastroenterol. Hepatol. Nutr. 2018, 21, 141–146. [Google Scholar] [CrossRef]
- Wang, S.; Huang, M.; You, X.; Zhao, J.; Chen, L.; Wang, L.; Luo, Y.; Chen, Y. Gut microbiota mediates the anti-obesity effect of calorie restriction in mice. Sci. Rep. 2018, 8, 13037. [Google Scholar] [CrossRef]
- Evans, C.C.; LePard, K.J.; Kwak, J.W.; Stancukas, M.C.; Laskowski, S.; Dougherty, J.; Moulton, L.; Glawe, A.; Wang, Y.; Leone, V.; et al. Exercise Prevents Weight Gain and Alters the Gut Microbiota in a Mouse Model of High Fat Diet-Induced Obesity. PLoS ONE 2014, 9, e92193. [Google Scholar] [CrossRef]
- Shang, Q.; Song, G.; Zhang, M.; Shi, J.; Xu, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan improves metabolic syndrome in association with increased Akkermansia population in the gut microbiota of high-fat diet-fed mice. J. Funct. Foods 2017, 28, 138–146. [Google Scholar] [CrossRef]
- Zhang, M.; Zou, X.; Zhao, D.; Zhao, F.; Li, C. Pork Meat Proteins Alter Gut Microbiota and Lipid Metabolism Genes in the Colon of Adaptive Immune-Deficient Mice. Mol. Nutr. Food Res. 2020, 64, 1901105. [Google Scholar] [CrossRef]
- Wu, W.; Xie, J.; Zhang, H. Dietary fibers influence the intestinal SCFAs and plasma metabolites profiling in growing pigs. Food Funct. 2016, 7, 4644–4654. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liu, L.; Long, S.; Pan, L.; Piao, X. Effect of organic acids and essential oils on performance, intestinal health and digestive enzyme activities of weaned pigs. Anim. Feed Sci. Technol. 2018, 235, 110–119. [Google Scholar] [CrossRef]
- Fan, P.; Liu, P.; Song, P.; Chen, X.; Ma, X. Moderate dietary protein restriction alters the composition of gut microbiota and improves ileal barrier function in adult pig model. Sci. Rep. 2017, 7, srep43412. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Nucleotide Sequence Primers (5′-3′) | Reverse Nucleotide Sequence Primers (5′-3′) | Product Size (bp) |
|---|---|---|---|
| GAPDH | ACCACAGTCCATGCCATCAC | TCCACCACCCTGTTGCTGTA | 172 |
| IL-1β | TCGCAGCAGCACATCAACAAGAG | AGGTCCACGGGAAAGACACAGG | 97 |
| IL-6 | AGAGGATACCACTCCCAACAGACC | AGCCACTCCTTCTGTGACTCCAG | 97 |
| TNF-α | GGACTAGCCAGGAGGGAGAACAG | GCCAGTGAGTGAAAGGGACAGAAC | 103 |
| Claudin-1 | AGATACAGTGCAAAGTCTTCGA | CAGGATGCCAATTACCATCAAG | 86 |
| ZO-1 | CTGGTGAAGTCTCGGAAAAATG | CATCTCTTGCTGCCAAACTATC | 97 |
| Occuldin | TGCTTCATCGCTTCCTTAGTAA | GGGTTCACTCCCATTATGTACA | 155 |
| Mucin1 | ACGTGAAGTCACAGCTTATACA | AGGGCAAGGAAATAGACGATAG | 193 |
| Mucin2 | TGCTGACGAGTGGTTGGTGAATG | TGATGAGGTGGCAGACAGGAGAC | 137 |
| IL-22 | GCAGATAACAACACAGATGTCC | GTCTTCCAGGGTGAAGTTGAG | 111 |
| IL-23 | CAGCGGGACATATGAATCTACT | TTGAAGATGTCAGAGTCAAGCA | 184 |
| PKCƟ | CGCCGACAGAGCACTCATCAAC | CAGTTCCCAGAGAGAAGGCAAATCC | 144 |
| CD4 | CAGCATGGCAAAGGTGTATTAA | GACTGAAGGTCACTTTGAACAC | 202 |
| LCK | AGGGAGAAGTGGTGAAACATTA | AATCCGGGAAAAGTGATACGAG | 85 |
| LAT | CAGCAGAATTCAGATGATGAGC | TTCACGTAATCTTCACACGACT | 200 |
| ITK | CTACCTGGAAAAAGCTTGTGTC | CGTGGAGCTGGTATATTGATCA | 136 |
| Galnt5 | ACGCAGGCAGAGAGTGACAGG | GCAACAGCAGCAGCAGTAGGAG | 114 |
| Galnt10 | CCCAACACCAGCATCATCATCCC | GACCAACACAATCTCCGCCACTAG | 117 |
| Eci2 | CTTGTCACTGAAGTTTTCCCTG | CTTTGGTTTTCTGGAGACGAAG | 234 |
| Hmgcr | GGTGCAAAGTTCCTTAGTGATG | GAATAGACACACCACGTTCATG | 112 |
| Ehhadh | GAATGGACTCCAGAAAGCTAGT | CGCTGTATTTCATCTACCAAGC | 150 |
| Acat1 | AGGTAGCTCAGTCGGTAGTGTTGG | TGGATCGTTTCTCGGTTCGTTTCTC | 138 |
| Acsl3 | CGGAAATCATGGATCGGATCTA | GTGGAGTACTACACCCTTTTGA | 132 |
| Hmgcs1 | AAGCACAGCCACCGAGCATATTC | ACCATCCCACCCCACACTGAAG | 150 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Zhong, R.; Tang, S.; Han, H.; Chen, L.; Zhang, H. Baicalin Alleviates Short-Term Lincomycin-Induced Intestinal and Liver Injury and Inflammation in Infant Mice. Int. J. Mol. Sci. 2022, 23, 6072. https://doi.org/10.3390/ijms23116072
Zhang S, Zhong R, Tang S, Han H, Chen L, Zhang H. Baicalin Alleviates Short-Term Lincomycin-Induced Intestinal and Liver Injury and Inflammation in Infant Mice. International Journal of Molecular Sciences. 2022; 23(11):6072. https://doi.org/10.3390/ijms23116072
Chicago/Turabian StyleZhang, Shunfen, Ruqing Zhong, Shanlong Tang, Hui Han, Liang Chen, and Hongfu Zhang. 2022. "Baicalin Alleviates Short-Term Lincomycin-Induced Intestinal and Liver Injury and Inflammation in Infant Mice" International Journal of Molecular Sciences 23, no. 11: 6072. https://doi.org/10.3390/ijms23116072
APA StyleZhang, S., Zhong, R., Tang, S., Han, H., Chen, L., & Zhang, H. (2022). Baicalin Alleviates Short-Term Lincomycin-Induced Intestinal and Liver Injury and Inflammation in Infant Mice. International Journal of Molecular Sciences, 23(11), 6072. https://doi.org/10.3390/ijms23116072