
The Use of Pro-Angiogenic and/or Pro-Hypoxic miRNAs as Tools to Monitor Patients with Diffuse Gliomas

 , , , and
, , , and
Abstract
1. Introduction
2. Results
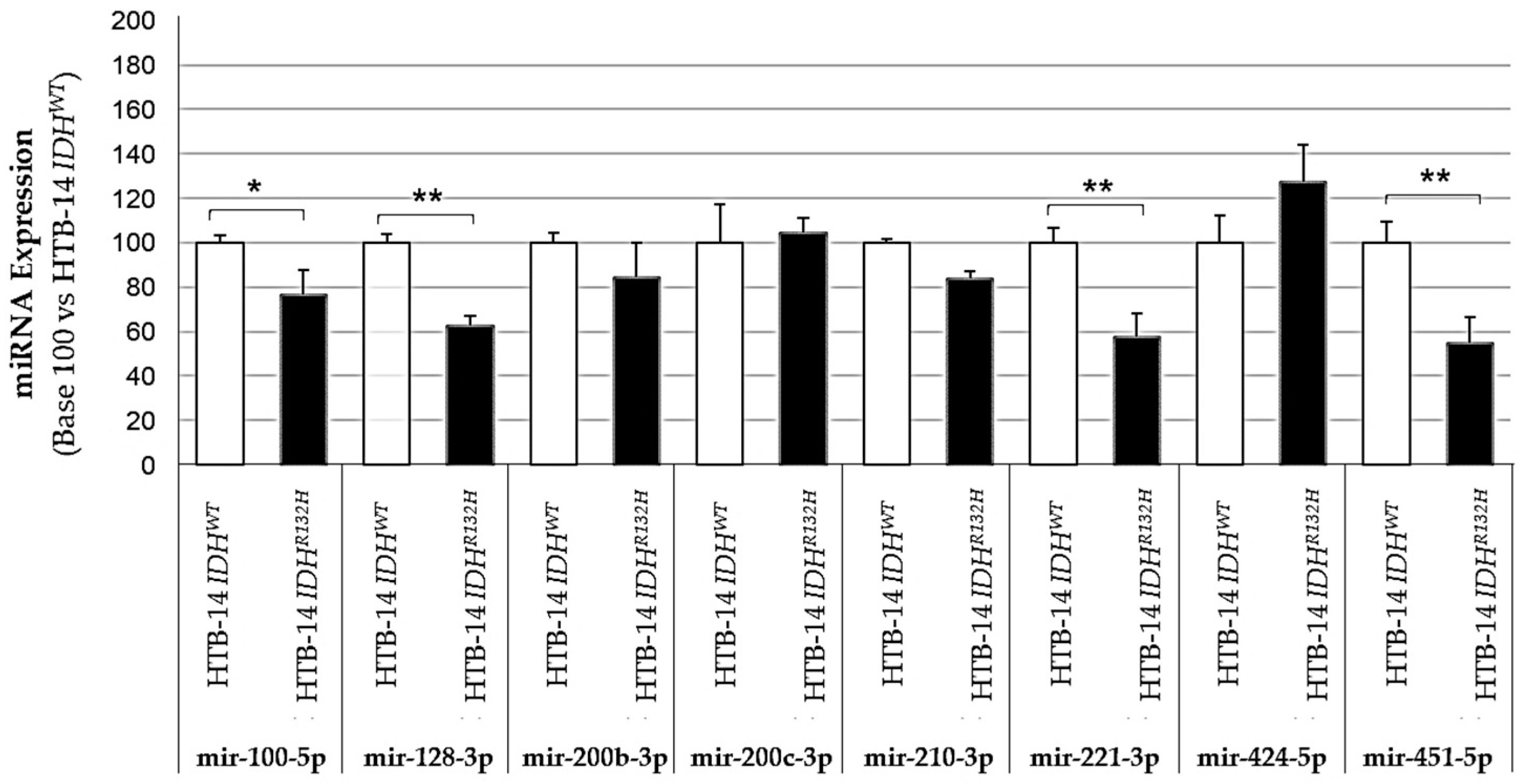
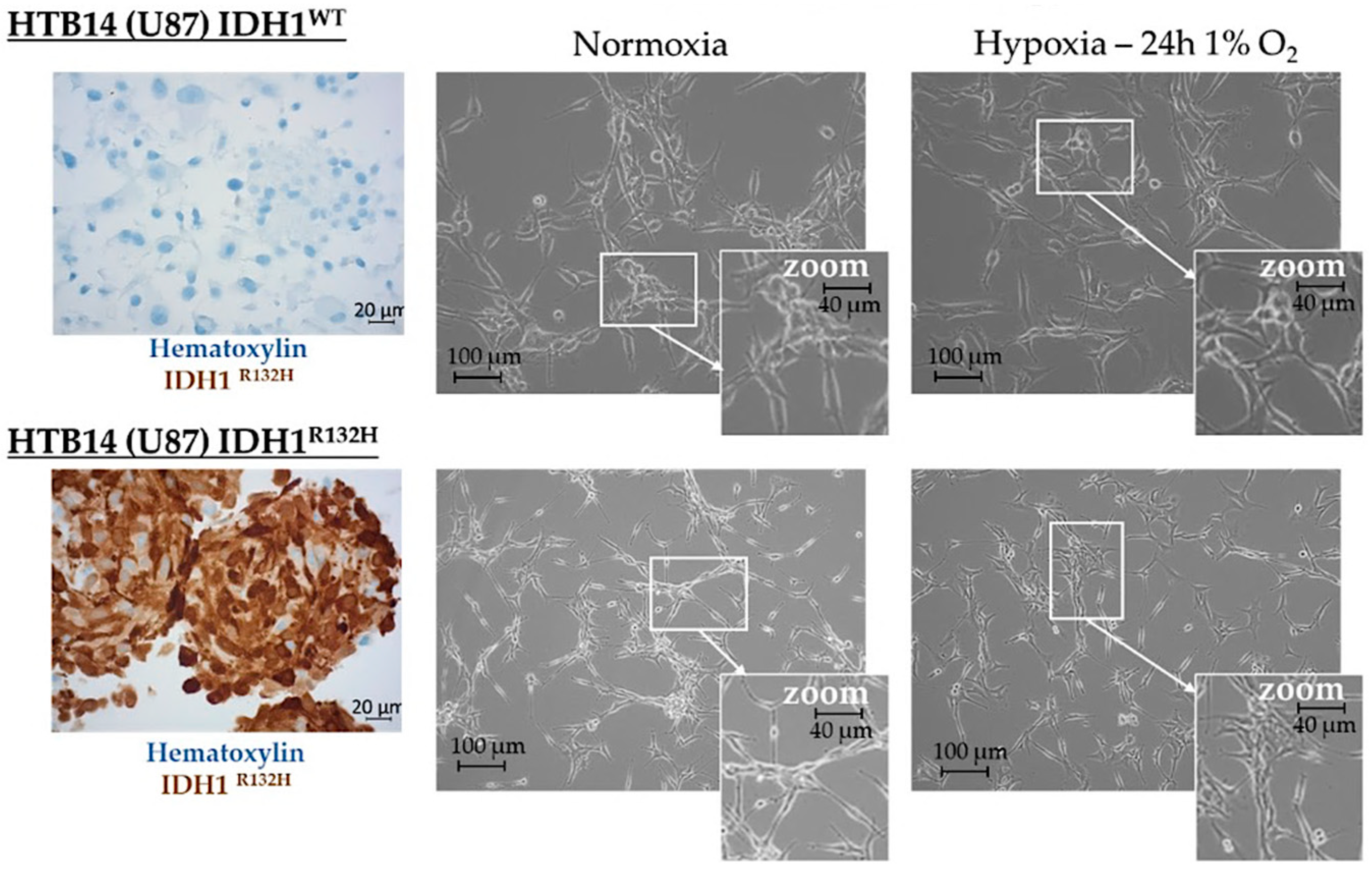
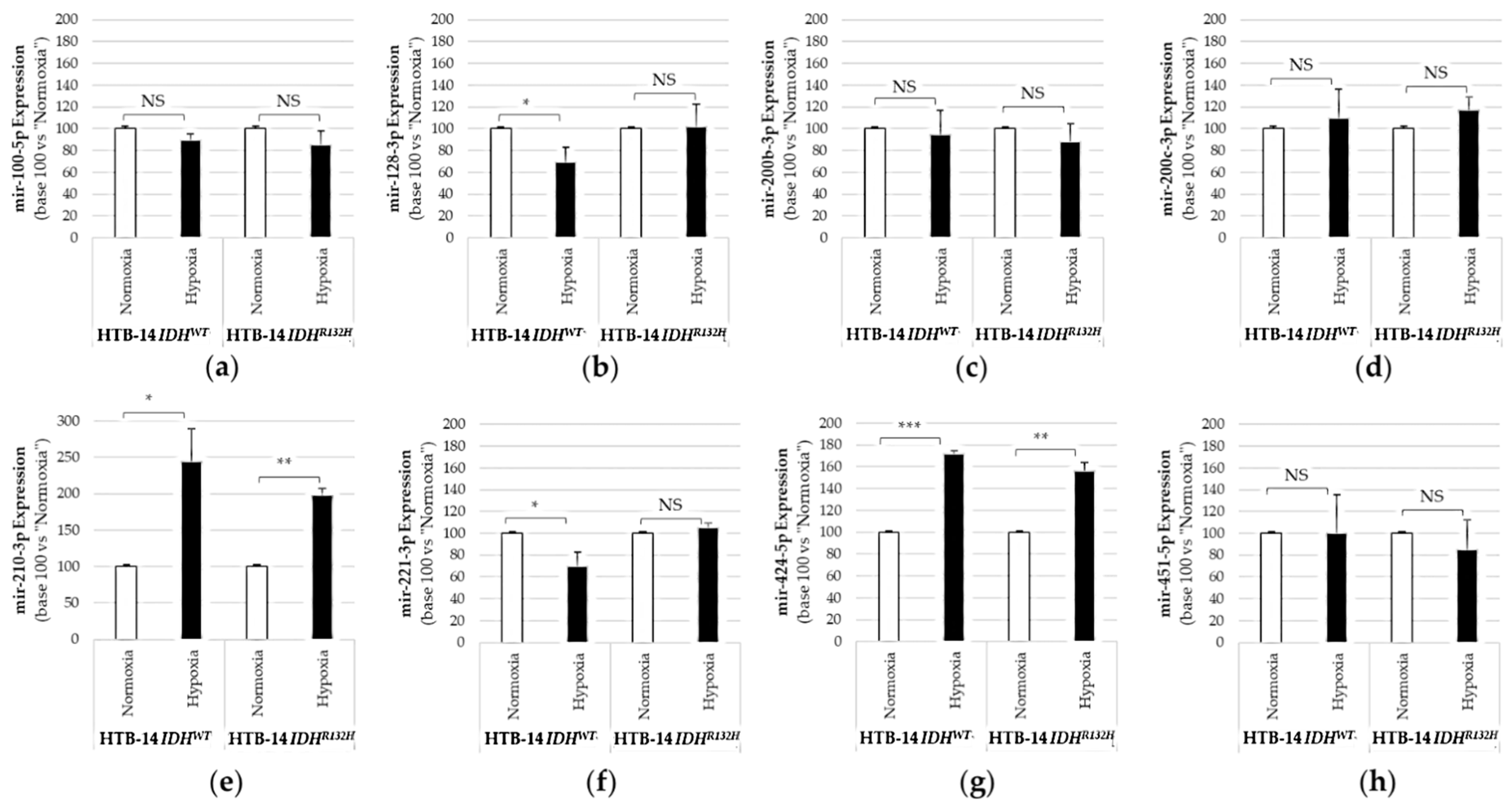
2.1. Expression of the Pro-Angiogenic and/or Pro-Hypoxic miRNAs Studied Is Affected by IDH Mutation and Hypoxia In Vitro
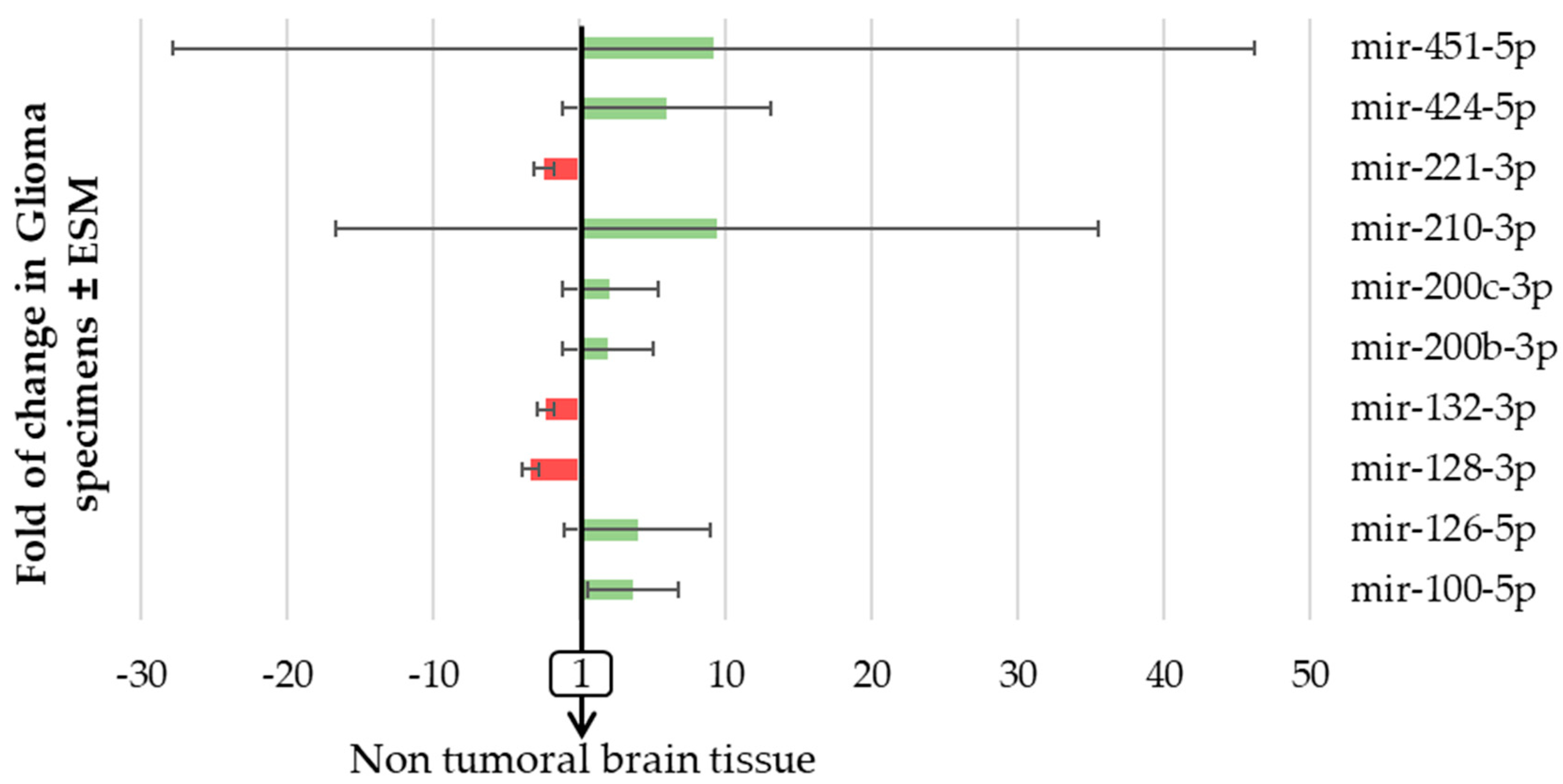
2.2. The Expression of Pro-Angiogenic and/or Pro-Hypoxic miRNAs Is Strongly Affected in Patients with Glioma
2.3. The Expression of Pro-Angiogenic and/or Pro-Hypoxic miRNAs Is Correlated with the 2016 WHO Classification
2.4. The Expression of Pro-Angiogenic and/or Pro-Hypoxic miRNAs Predicts Overall (OS) and Progression-Free (PFS) Survival in Patients with Glioma in Univariate Analysis
2.5. Mir-128-3p Predicts a Poorer PFS in Patients with AA-IDHMUT or AO-IDHMUT and mir-100-5p Predicts a Poorer PFS in Patients with AA-IDHMUT
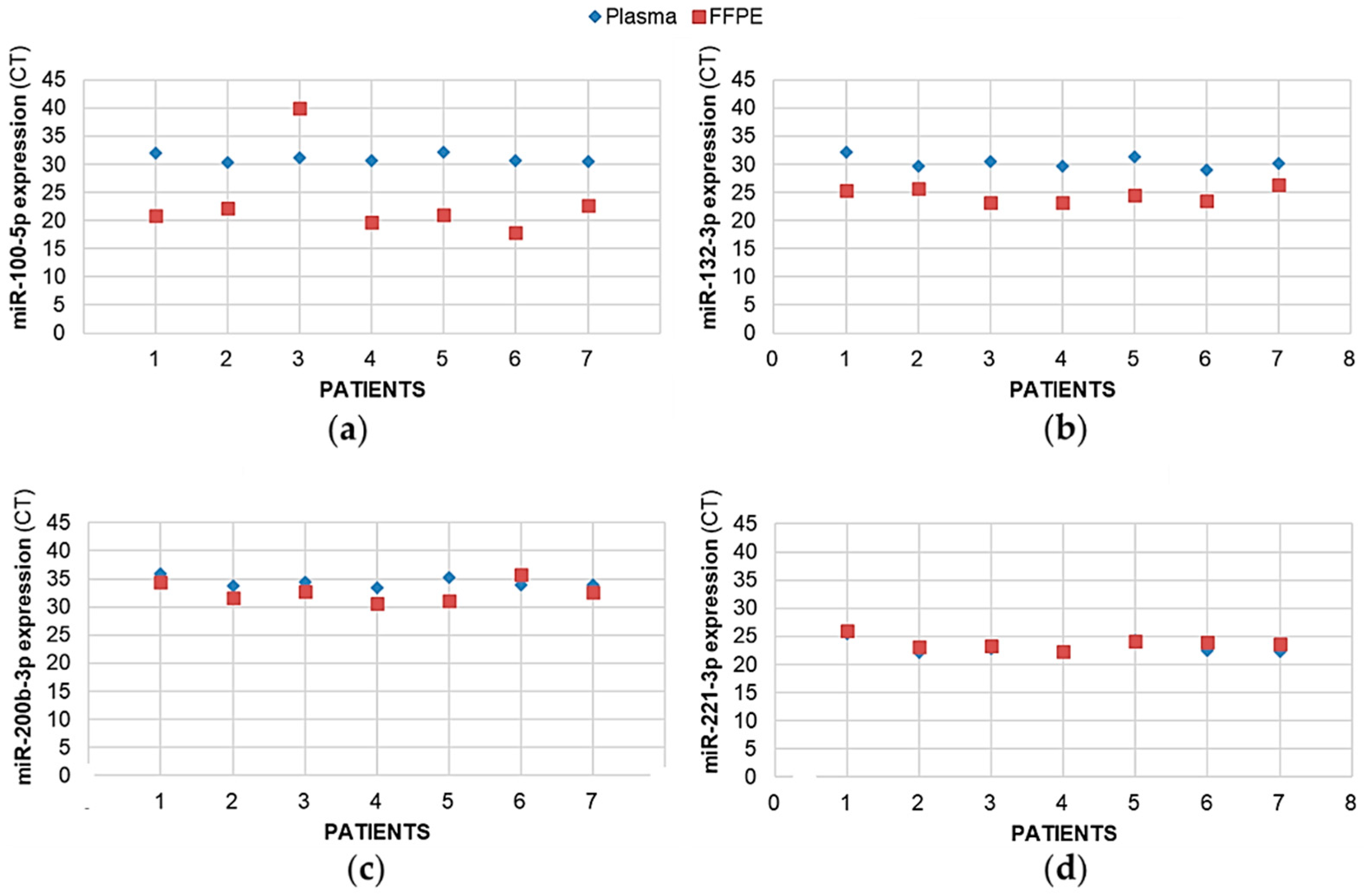
2.6. For the Same Patient, Expression of Plasma miRNAs Coincides with the Expression of Tumoral miRNAs
3. Discussion
4. Materials and Methods
4.1. Tissue Samples and Patient Characteristics
4.2. Cell Culture and Hypoxia Treatment
4.3. DNA and miRNA Extraction
4.4. Quantitative Real-Time Reverse Transcription: PCR
4.5. IDH1 and IDH2 Mutations Assay
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Torrisi, F.; Alberghina, C.; D’Aprile, S.; Pavone, A.M.; Longhitano, L.; Giallongo, S.; Tibullo, D.; Di Rosa, M.; Zappalà, A.; Cammarata, F.P.; et al. The Hallmarks of Glioblastoma: Heterogeneity, Intercellular Crosstalk and Molecular Signature of Invasiveness and Progression. Biomedicines 2022, 10, 806. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef]
- Komori, T. The 2016 WHO Classification of Tumours of the Central Nervous System: The major points of revision. Neurol Med. Chir. 2017, 57, 301–311. [Google Scholar] [CrossRef]
- Iuchi, T.; Sugiyama, T.; Ohira, M.; Kageyama, H.; Yokoi, S.; Sakaida, T.; Hasegawa, Y.; Setoguchi, T.; Itami, M. Clinical significance of the 2016 WHO classification in Japanese patients with gliomas. Brain Tumor. Pathol. 2018, 35, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro-Oncology 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Liu, Y.; Cai, S.J.; Qian, M.; Ding, J.; Larion, M.; Gilbert, M.R.; Yang, C. IDH mutation in glioma: Molecular mechanisms and potential therapeutic targets. Br. J. Cancer 2020, 122, 1580–1589. [Google Scholar] [CrossRef] [PubMed]
- van den Bent, M.J.; Dubbink, H.J.; Marie, Y.; Brandes, A.A.; Taphoorn, M.J.; Wesseling, P.; Frenay, M.; Tijssen, C.C.; Lacombe, D.; Idbaih, A.; et al. IDH1 and IDH2 mutations are prognostic but not predictive for outcome in anaplastic oligodendroglial tumors: A report of the European Organization for Research and Treatment of Cancer Brain Tumor Group. Clin. Cancer Res. 2010, 16, 1597–1604. [Google Scholar] [CrossRef] [PubMed]
- Beyer, S.; Fleming, J.; Meng, W.; Singh, R.; Haque, S.J.; Chakravarti, A. The Role of miRNAs in Angiogenesis, Invasion and Metabolism and Their Therapeutic Implications in Gliomas. Cancers 2017, 9, 85. [Google Scholar] [CrossRef] [PubMed]
- Turcan, S.; Rohle, D.; Goenka, A.; Walsh, L.A.; Fang, F.; Yilmaz, E.; Campos, C.; Fabius, A.W.; Lu, C.; Ward, P.S.; et al. IDH1 mutation is sufficient to establish the glioma hypermethylator phenotype. Nature 2012, 483, 479–483. [Google Scholar] [CrossRef]
- Levallet, G.; Creveuil, C.; Bekaert, L.; Péres, E.; Planchard, G.; Lecot-Cotigny, S.; Guillamo, J.S.; Emery, E.; Zalcman, G.; Lechapt-Zalcman, E. Promoter Hypermethylation of Genes Encoding for RASSF/Hippo Pathway Members Reveals Specific Alteration Pattern in Diffuse Gliomas. J. Mol. Diagn. 2019, 21, 695–704. [Google Scholar] [CrossRef]
- Cheng, W.; Ren, X.; Zhang, C.; Han, S.; Wu, A. Expression and prognostic value of microRNAs in lower-grade glioma depends on IDH1/2 status. J. Neurooncol. 2017, 132, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Miska, E.A. How microRNAs control cell division, differentiation and death. Curr. Opin. Genet. Dev. 2005, 15, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.C.; Kwok, W.K.; Chen, Z.; Ng, H.K. Oncogenic role of microRNAs in brain tumors. Acta Neuropathol. 2009, 117, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Li, R.; Liu, Y.; Yang, P.; Wang, Z.; Zhang, C.; Bao, Z.; Zhang, W.; You, Y.; Jiang, T. MicroRNA expression patterns in the malignant progression of gliomas and a 5-micro-RNA signature for prognosis. Oncotarget 2014, 5, 12908–12915. [Google Scholar] [CrossRef]
- Rezaei, O.; Honarmand, K.; Nateghinia, S.; Taheri, M.; Ghafouri-Fard, S. miRNA signature in glioblastoma: Potential biomarkers and therapeutic targets. Exp. Mol. Pathol. 2020, 117, 104550. [Google Scholar] [CrossRef]
- Ames, H.; Halushka, M.K.; Rodriguez, F.J. miRNA Regulation in Gliomas: Usual Suspects in Glial Tumorigenesis and Evolving Clinical Applications. J. Neuropathol. Exp. Neurol. 2017, 76, 246–254. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, J.; Xue, Q.; Wang, J.; Zhao, L.; Han, K.; Zhang, D.; Hou, L. Prognostic Significance of MicroRNAs in Glioma: A Systematic Review and Meta-Analysis. Biomed. Res. Int. 2019, 2019, 4015969. [Google Scholar] [CrossRef]
- Fasanaro, P.; Greco, S.; Lorenzi, M.; Pescatori, M.; Brioschi, M.; Kulshreshtha, R.; Banfi, C.; Stubbs, A.; Calin, G.A.; Ivan, M.; et al. An integrated approach for experimental target identification of hypoxia-induced miR-210. J. Biol Chem. 2009, 284, 35134–35143. [Google Scholar] [CrossRef]
- Alaiti, M.A.; Ishikawa, M.; Masuda, H.; Simon, D.I.; Jain, M.K.; Asahara, T.; Costa, M.A. Up-regulation of miR-210 by vascular endothelial growth factor in ex vivo expanded CD34+ cells enhances cell-mediated angiogenesis. J. Cell Mol. Med. 2012, 16, 2413–2421. [Google Scholar] [CrossRef]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef]
- Teplyuk, N.M.; Mollenhauer, B.; Gabriely, G.; Giese, A.; Kim, E.; Smolsky, M.; Kim, R.Y.; Saria, M.G.; Pastorino, S.; Kesari, S.; et al. MicroRNAs in cerebrospinal fluid identify glioblastoma and metastatic brain cancers and reflect disease activity. Neuro-oncology 2012, 14, 689–700. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Jiang, Y.; Liu, L.; Zuo, Z.; Zeng, C. Circulating MicroRNAs as Promising Diagnostic Biomarkers for Patients with Glioma: A Meta-Analysis. Front. Neurol. 2021, 11, 610163. [Google Scholar] [CrossRef] [PubMed]
- Shimono, Y.; Zabala, M.; Cho, R.W.; Lobo, N.; Dalerba, P.; Qian, D.; Diehn, M.; Liu, H.; Panula, S.P.; Chiao, E.; et al. Downregulation of miRNA-200c links breast cancer stem cells with normal stem cells. Cell 2009, 138, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, G.; Subramanian, I.V.; Adhikari, N.; Zhang, X.; Joshi, H.P.; Basi, D.; Chandrashekhar, Y.S.; Hall, J.L.; Roy, S.; Zeng, Y.; et al. Hypoxia-induced microRNA-424 expression in human endothelial cells regulates HIF-α isoforms and promotes angiogenesis. J. Clin. Investig. 2010, 120, 4141–4154. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.C.; Yoon, S.; Jeong, Y.; Yoon, J.; Baek, K. Regulation of vascular endothelial growth factor signaling by miR-200b. Mol. Cells 2011, 32, 77–82. [Google Scholar] [CrossRef]
- Katare, R.; Riu, F.; Mitchell, K.; Gubernator, M.; Campagnolo, P.; Cui, Y.; Fortunato, O.; Avolio, E.; Cesselli, D.; Beltrami, A.P.; et al. Transplantation of human pericyte progenitor cells improves the repair of infarcted heart through activation of an angiogenic program involving micro-RNA-132. Circ. Res. 2011, 109, 894–906. [Google Scholar] [CrossRef]
- González-Gómez, P.; Sánchez, P.; Mira, H. MicroRNAs as Regulators of Neural Stem Cell-Related Pathways in Glioblastoma Multiforme. Mol. Neurobiol. 2011, 44, 235–249. [Google Scholar] [CrossRef]
- Quintavalle, C.; Garofalo, M.; Zanca, C.; Romano, G.; Iaboni, M.; del Basso De Caro, M.; Martinez-Montero, J.C.; Incoronato, M.; Nuovo, G.; Croce, C.M.; et al. miR-221/222 overexpession in human glioblastoma increases invasiveness by targeting the protein phosphate PTPμ. Oncogene 2012, 31, 858–868. [Google Scholar] [CrossRef]
- Ma, X.; Yoshimoto, K.; Guan, Y.; Hata, N.; Mizoguchi, M.; Sagata, N.; Murata, H.; Kuga, D.; Amano, T.; Nakamizo, A.; et al. Associations between microRNA expression and mesenchymal marker gene expression in glioblastoma. Neuro-Oncology 2012, 14, 1153–1162. [Google Scholar] [CrossRef][Green Version]
- Shang, C.; Hong, Y.; Guo, Y.; Liu, Y.H.; Xue, Y.X. MiR-210 up-regulation inhibits proliferation and induces apoptosis in glioma cells by targeting SIN3A. Med. Sci. Monit. 2014, 20, 2571–2577. [Google Scholar]
- Luan, Y.; Zhang, S.; Zuo, L.; Zhou, L. Overexpression of miR-100 inhibits cell proliferation, migration, and chemosensitivity in human glioblastoma through FGFR3. Onco. Targets Ther. 2015, 8, 3391–3400. [Google Scholar] [PubMed]
- Han, I.B.; Kim, M.; Lee, S.H.; Kim, J.K.; Kim, S.H.; Chang, J.H.; Teng, Y.D. Down-regulation of MicroRNA-126 in Glioblastoma and its Correlation with Patient Prognosis: A Pilot Study. Anticancer Res. 2016, 36, 6691–6697. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Wang, L.; Li, T.; Zhu, M.; Zhang, C.; Chen, L.; Zhao, P.; Zhou, H.; Yu, S.; Yang, X. The role of miR-451 in the switching between proliferation and migration in malignant glioma cells: AMPK signaling, mTOR modulation and Rac1 activation required. Int. J. Oncol. 2017, 50, 1989–1999. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Vera, Y.M.; Marchat, L.A.; Gallardo-Rincón, D.; Ruiz-García, E.; Astudillo-De La Vega, H.; Echavarría-Zepeda, R.; López-Camarillo, C. AngiomiRs: MicroRNAs driving angiogenesis in cancer (Review). Int. J. Mol. Med. 2019, 43, 657–670. [Google Scholar] [CrossRef]
- Behl, T.; Kumar, C.; Makkar, R.; Gupta, A.; Sachdeva, M. Intercalating the Role of MicroRNAs in Cancer: As Enemy or Protector. Asian Pac. J. Cancer Prev. 2020, 21, 593–598. [Google Scholar] [CrossRef]
- Molina-Pinelo, S.; Suárez, R.; Pastor, M.D.; Nogal, A.; Márquez-Martín, E.; Martín-Juan, J.; Carnero, A.; Paz-Ares, L. Association between the miRNA signatures in plasma and bronchoalveolar fluid in respiratory pathologies. Dis. Markers 2012, 32, 221–230. [Google Scholar] [CrossRef]
- Papagiannakopoulos, T.; Friedmann-Morvinski, D.; Neveu, P.; Dugas, J.C.; Gill, R.M.; Huillard, E.; Liu, C.; Zong, H.; Rowitch, D.H.; Barres, B.A.; et al. Pro-neural miR-128 is a glioma tumor suppressor that targets mitogenic kinases. Oncogene 2012, 15, 1884–1895. [Google Scholar] [CrossRef]
- Li, Y.; Yan, C.; Fan, J.; Hou, Z.; Han, Y. MiR-221-3p targets Hif-1α to inhibit angiogenesis in heart failure. Lab. Investig. 2021, 101, 104–115. [Google Scholar] [CrossRef]
- Zhou, X.U.; Qi, L.; Tong, S.; Cui, Y.U.; Chen, J.; Huang, T.; Chen, Z.; Zu, X.B. miR-128 downregulation promotes growth and metastasis of bladder cancer cells and involves VEGF-C upregulation. Oncol. Lett. 2015, 10, 3183–3190. [Google Scholar] [CrossRef]
- Foote, M.B.; Papadopoulos, N.; Diaz, L.A., Jr. Genetic classification of gliomas: Refining histopathology. Cancer Cell 2015, 28, 9–11. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, G.; Zhou, Q.; Li, Z. Plasma microRNA-720 may predict prognosis and diagnosis in glioma patients. Biosci. Rep. 2020, 40, BSR20201449. [Google Scholar] [CrossRef] [PubMed]
- Agostini, M.; Knight, R.A. Mir-34: From bench to bedside. Oncotarget 2014, 5, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.; Chen, P.H.; Ho, K.H.; Shih, C.M.; Cheng, C.H.; Lin, C.W.; Cheng, K.T.; Liu, A.J.; Chen, K.C. The microRNA-302b-inhibited insulin-like growth factor-binding protein 2 signaling pathway induces glioma cell apoptosis by targeting nuclear factor IA. PLoS ONE 2017, 12, e0173890. [Google Scholar] [CrossRef] [PubMed]
- Li, C.G.; Pu, M.F.; Li, C.Z.; Gao, M.; Liu, M.X.; Yu, C.Z.; Yan, H.; Peng, C.; Zhao, Y.; Li, Y.; et al. MicroRNA-1304 suppresses human non-small cell lung cancer cell growth in vitro by targeting heme oxygenase-1. Acta Pharmacol. Sin. 2017, 38, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.Q.; Yin, T.K.; Li, Y.X.; Zhang, J.H.; Gu, J.J. miR-130b regulates the proliferation, invasion and apoptosis of glioma cells via targeting of CYLD. Oncol. Rep. 2017, 38, 167–174. [Google Scholar] [CrossRef]
- An, T.; Fan, T.; Zhang, X.Q.; Liu, Y.F.; Huang, J.; Liang, C.; Lv, B.H.; Wang, Y.Q.; Zhao, X.G.; Liu, J.X.; et al. Comparison of Alterations in miRNA Expression in Matched Tissue and Blood Samples during Spinal Cord Glioma Progression. Sci. Rep. 2019, 9, 9169–9184. [Google Scholar] [CrossRef]
- Cui, C.; Cui, Q. The relationship of human tissue microRNAs with those from body fluids. Sci. Rep. 2020, 10, 5644–5657. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, J.; Wang, Z.; Ruan, C.; Wang, L.; Guo, H. Serum miR-100 is a potential biomarker for detection and outcome prediction of glioblastoma patients. Cancer Biomark. 2019, 24, 43–49. [Google Scholar] [CrossRef]
- Adlakha, Y.K.; Saini, N. Brain microRNAs and insights into biological functions and therapeutic potential of brain enriched miRNA128. Mol. Cancer 2014, 21, 13–33. [Google Scholar]
- TissueAtlas. Available online: https://ccb-web.cs.uni-saarland.de/tissueatlas/patterns (accessed on 27 April 2022).
- Wang, X.W.; Labussière, M.; Valable, S.; Pérès, E.A.; Guillamo, J.S.; Bernaudin, M.; Sanson, M. IDH1(R132H) mutation increases U87 glioma cell sensitivity to radiation therapy in hypoxia. Biomed. Res. Int. 2014, 2014, 198697. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mir-200c-3p | mir-210-3p | mir-100-5p | mir-126-5p | mir-132-3p | mir-221-3p | mir-424-5p | mir-128-3p | mir-451-5p | |
| mir-200b-3p | 0.707 | 0.596 | 0.713 | 0.740 | 0.700 | 0.784 | 0.793 | 0.385 | 0.619 |
| mir-200c-3p | 0.494 | 0.591 | 0.778 | 0.726 | 0.791 | 0.764 | 0.355 | 0.641 | |
| mir-210-3p | 0.435 | 0.550 | 0.344 | 0.543 | 0.549 | 0.078 | 0.464 | ||
| mir-100-5p | 0.642 | 0.627 | 0.607 | 0.741 | 0.535 | 0.514 | |||
| mir-126-5p | 0.729 | 0.797 | 0.781 | 0.434 | 0.749 | ||||
| mir-132-3p | 0.822 | 0.725 | 0.592 | 0.546 | |||||
| mir-221-3p | 0.728 | 0.381 | 0.621 | ||||||
| mir-424-5p | 0.549 | 0.702 | |||||||
| mir-128-3p | 0.539 |
| IDH1/2-WT (n = 37) | IDH1/2-MUT (n = 60) | p | |||||
|---|---|---|---|---|---|---|---|
| Median (%) | First Quartile (%) | Third Quartile (%) | Median (%) | First Quartile (%) | Third Quartile (%) | ||
| mir-200b-3p | 250.5 | 82.9 | 450.0 | 59.3 | 21.8 | 97.8 | <0.001 |
| mir-200c-3p | 262.4 | 122.0 | 449.0 | 64.8 | 20.9 | 138.9 | <0.001 |
| mir-210-3p | 1109.0 | 380.1 | 1891.2 | 126.7 | 67.2 | 201.3 | <0.001 |
| mir-100-5p | 362.9 | 189.1 | 597.7 | 238.6 | 141.2 | 410.4 | 0.086 |
| mir-126-5p | 451.0 | 175.7 | 1194.3 | 119.6 | 38.0 | 294.6 | <0.001 |
| mir-132-3p | 32.1 | 18.4 | 56.0 | 20.0 | 9.2 | 36.7 | 0.024 |
| mir-221-3p | 35.7 | 17.4 | 90.4 | 8.5 | 3.0 | 18.5 | <0.001 |
| mir-424-5p | 699.8 | 257.9 | 1324.0 | 215.4 | 104.8 | 410.5 | <0.001 |
| mir-128-3p | 4.3 | 1.2 | 21.9 | 9.9 | 5.4 | 37.8 | 0.027 |
| mir-451-5p | 322.7 | 67.4 | 1104.3 | 73.5 | 25.9 | 310.6 | 0.0010 |
| No Microvascular Proliferation (n = 40) | Microvascular Proliferation (n = 57) | p | |||||
|---|---|---|---|---|---|---|---|
| Median (%) | First Quartile (%) | Third Quartile (%) | Median (%) | First Quartile (%) | Third Quartile (%) | ||
| mir-200b-3p | 62.0 | 28.6 | 115.0 | 99.0 | 26.0 | 386.4 | 0.021 |
| mir-200c-3p | 57.7 | 21.0 | 162.4 | 138.7 | 71.3 | 359.7 | 0.010 |
| mir-210-3p | 117.4 | 53.3 | 154.0 | 498.0 | 192.7 | 1568.4 | <0.001 |
| mir-100-5p | 251.9 | 176.8 | 420.4 | 267.5 | 124.8 | 579.3 | 0.96 |
| mir-126-5p | 120.2 | 43.8 | 334.4 | 275.7 | 117.5 | 688.3 | 0.0073 |
| mir-132-3p | 26.4 | 9.5 | 42.3 | 25.5 | 13.0 | 49.3 | 0.51 |
| mir-221-3p | 8.6 | 3.8 | 20.6 | 19.5 | 7.8 | 63.2 | 0.0063 |
| mir-424-5p | 192.2 | 114.6 | 366.2 | 447.5 | 179.9 | 1075.5 | 0.0063 |
| mir-128-3p | 10.0 | 5.4 | 42.5 | 5.8 | 2.2 | 26.4 | 0.087 |
| mir-451-5p | 59.9 | 25.9 | 261.2 | 256.1 | 51.8 | 879.9 | 0.0068 |
| O (n = 11) | AO (n = 16) | A (n = 18) | AA (n = 7) | GB-IDHMUT (n = 8) | GB-IDHWT (n = 37) | p | |
|---|---|---|---|---|---|---|---|
| Median (%) | |||||||
| mir-200b-3p | 37.9 | 35.0 | 58.9 | 96.6 | 63.5 | 250.5 | <0.001 |
| mir-200c-3p | 45.0 | 88.9 | 66.7 | 166.1 | 53.0 | 262.4 | <0.001 |
| mir-210-3p | 98.1 | 139.1 | 110.5 | 118.2 | 228.1 | 1109.0 | <0.001 |
| mir-100-5p | 208.3 | 241.2 | 350.8 | 242.7 | 231.6 | 362.9 | 0.28 |
| mir-126-5p | 105.9 | 274.3 | 113.3 | 131.8 | 103.5 | 451.0 | <0.001 |
| mir-132-3p | 15.6 | 18.6 | 23.1 | 47.8 | 15.2 | 32.1 | 0.10 |
| mir-221-3p | 4.6 | 8.1 | 8.5 | 13.6 | 8.8 | 35.7 | <0.001 |
| mir-424-5p | 115.0 | 271.1 | 203.9 | 574.3 | 274.9 | 699.8 | 0.0012 |
| mir-128-3p | 7.1 | 12.9 | 15.2 | 9.7 | 10.6 | 4.3 | 0.28 |
| mir-451-5p | 71.9 | 313.9 | 53.9 | 58.2 | 116.3 | 322.7 | 0.0077 |
| HR † | IC95% | p | Adjusted HR ‡ | IC95% | p | |||
|---|---|---|---|---|---|---|---|---|
| OS | ||||||||
| mir-200b-3p | 1.08 | 1.05 | 1.12 | <0.001 | 1.03 | 0.98 | 1.08 | 0.30 |
| mir-200c-3p | 1.06 | 1.03 | 1.09 | <0.001 | 1.02 | 0.98 | 1.05 | 0.42 |
| mir-210-3p | 1.015 | 1.008 | 1.02 | <0.001 | 1.01 | 0.997 | 1.01 | 0.23 |
| mir-100-5p | 1.02 | 0.90 | 1.15 | 0.80 | 0.92 | 0.81 | 1.06 | 0.25 |
| mir-126-5p | 1.08 | 1.03 | 1.12 | <0.001 | 0.99 | 0.94 | 1.05 | 0.83 |
| mir-132-3p | 1.04 | 1.00 | 1.08 | 0.055 | 1.02 | 0.98 | 1.07 | 0.38 |
| mir-221-3p | 1.07 | 1.04 | 1.11 | <0.001 | 1.01 | 0.97 | 1.05 | 0.50 |
| mir-424-5p | 1.07 | 1.02 | 1.13 | 0.0038 | 1.00 | 0.95 | 1.06 | 0.94 |
| mir-128-3p | 1.02 | 0.97 | 1.07 | 0.40 | 1.00 | 0.96 | 1.04 | 0.99 |
| mir-451-5p | 1.008 | 1.003 | 1.01 | 0.0022 | 1.003 | 0.997 | 1.01 | 0.31 |
| PFS | ||||||||
| mir-200b-3p | 1.07 | 1.03 | 1.11 | <0.001 | 1.02 | 0.96 | 1.07 | 0.56 |
| mir-200c-3p | 1.06 | 1.03 | 1.09 | <0.001 | 1.02 | 0.98 | 1.06 | 0.44 |
| mir-210-3p | 1.013 | 1.007 | 1.02 | <0.001 | 1.01 | 0.997 | 1.01 | 0.23 |
| mir-100-5p | 1.01 | 0.90 | 1.12 | 0.92 | 0.97 | 0.86 | 1.09 | 0.62 |
| mir-126-5p | 1.06 | 1.02 | 1.11 | 0.007 | 0.99 | 0.93 | 1.04 | 0.63 |
| mir-132-3p | 1.04 | 1.00 | 1.08 | 0.053 | 1.02 | 0.98 | 1.06 | 0.41 |
| mir-221-3p | 1.06 | 1.03 | 1.09 | <0.001 | 1.00 | 0.97 | 1.04 | 0.90 |
| mir-424-5p | 1.07 | 1.02 | 1.12 | 0.0034 | 1.00 | 0.95 | 1.06 | 0.93 |
| mir-128-3p | 1.01 | 0.98 | 1.05 | 0.45 | 0.99 | 0.96 | 1.03 | 0.76 |
| mir-451-5p | 1.007 | 1.00 | 1.01 | 0.0077 | 1.003 | 0.996 | 1.01 | 0.40 |
| mir-100-5p | mir-128-3p | |||||||
|---|---|---|---|---|---|---|---|---|
| HR | IC95% | p | HR | IC95% | p | |||
| O | 0.75 | 0.28 | 2.00 | 0.56 | 0.95 | 0.75 | 1.21 | 0.66 |
| A | 0.79 | 0.57 | 1.11 | 0.18 | 0.97 | 0.72 | 1.30 | 0.82 |
| AO | 0.99 | 0.776 | 1.26 | 0.94 | 1.33 | 1.046 | 1.68 | 0.020 |
| AA | 5.12 | 1.84 | 14.24 | 0.0018 | 1.31 | 1.06 | 1.60 | 0.010 |
| GB-IDHMUT | 0.68 | 0.42 | 1.08 | 0.10 | 0.93 | 0.65 | 1.31 | 0.66 |
| GB-IDHWT | 1.05 | 0.90 | 1.23 | 0.52 | 0.99 | 0.95 | 1.03 | 0.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levallet, G.; Dubois, F.; Leclerc, A.; Petit, E.; Bekaert, L.; Faisant, M.; Creveuil, C.; Emery, E.; Zalcman, G.; Lechapt-Zalcman, E. The Use of Pro-Angiogenic and/or Pro-Hypoxic miRNAs as Tools to Monitor Patients with Diffuse Gliomas. Int. J. Mol. Sci. 2022, 23, 6042. https://doi.org/10.3390/ijms23116042
Levallet G, Dubois F, Leclerc A, Petit E, Bekaert L, Faisant M, Creveuil C, Emery E, Zalcman G, Lechapt-Zalcman E. The Use of Pro-Angiogenic and/or Pro-Hypoxic miRNAs as Tools to Monitor Patients with Diffuse Gliomas. International Journal of Molecular Sciences. 2022; 23(11):6042. https://doi.org/10.3390/ijms23116042
Chicago/Turabian StyleLevallet, Guénaëlle, Fatéméh Dubois, Arthur Leclerc, Edwige Petit, Lien Bekaert, Maxime Faisant, Christian Creveuil, Evelyne Emery, Gérard Zalcman, and Emmanuèle Lechapt-Zalcman. 2022. "The Use of Pro-Angiogenic and/or Pro-Hypoxic miRNAs as Tools to Monitor Patients with Diffuse Gliomas" International Journal of Molecular Sciences 23, no. 11: 6042. https://doi.org/10.3390/ijms23116042
APA StyleLevallet, G., Dubois, F., Leclerc, A., Petit, E., Bekaert, L., Faisant, M., Creveuil, C., Emery, E., Zalcman, G., & Lechapt-Zalcman, E. (2022). The Use of Pro-Angiogenic and/or Pro-Hypoxic miRNAs as Tools to Monitor Patients with Diffuse Gliomas. International Journal of Molecular Sciences, 23(11), 6042. https://doi.org/10.3390/ijms23116042