
Platelet Extracellular Vesicles Are Taken up by Canine T Lymphocytes but Do Not Play a Role in Their Proliferation, Differentiation and Cytokine Production In Vitro


Abstract
:1. Introduction
2. Results
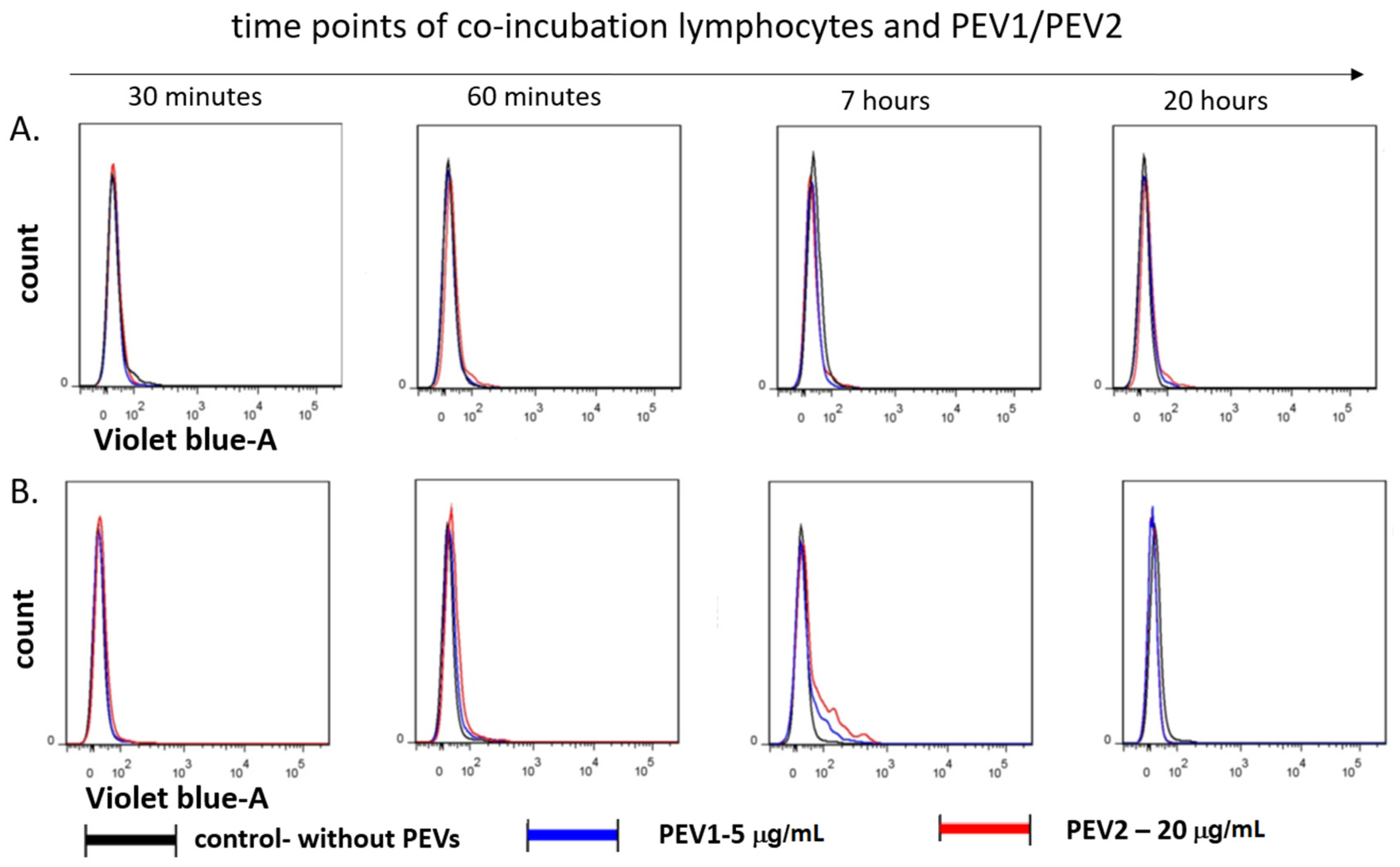
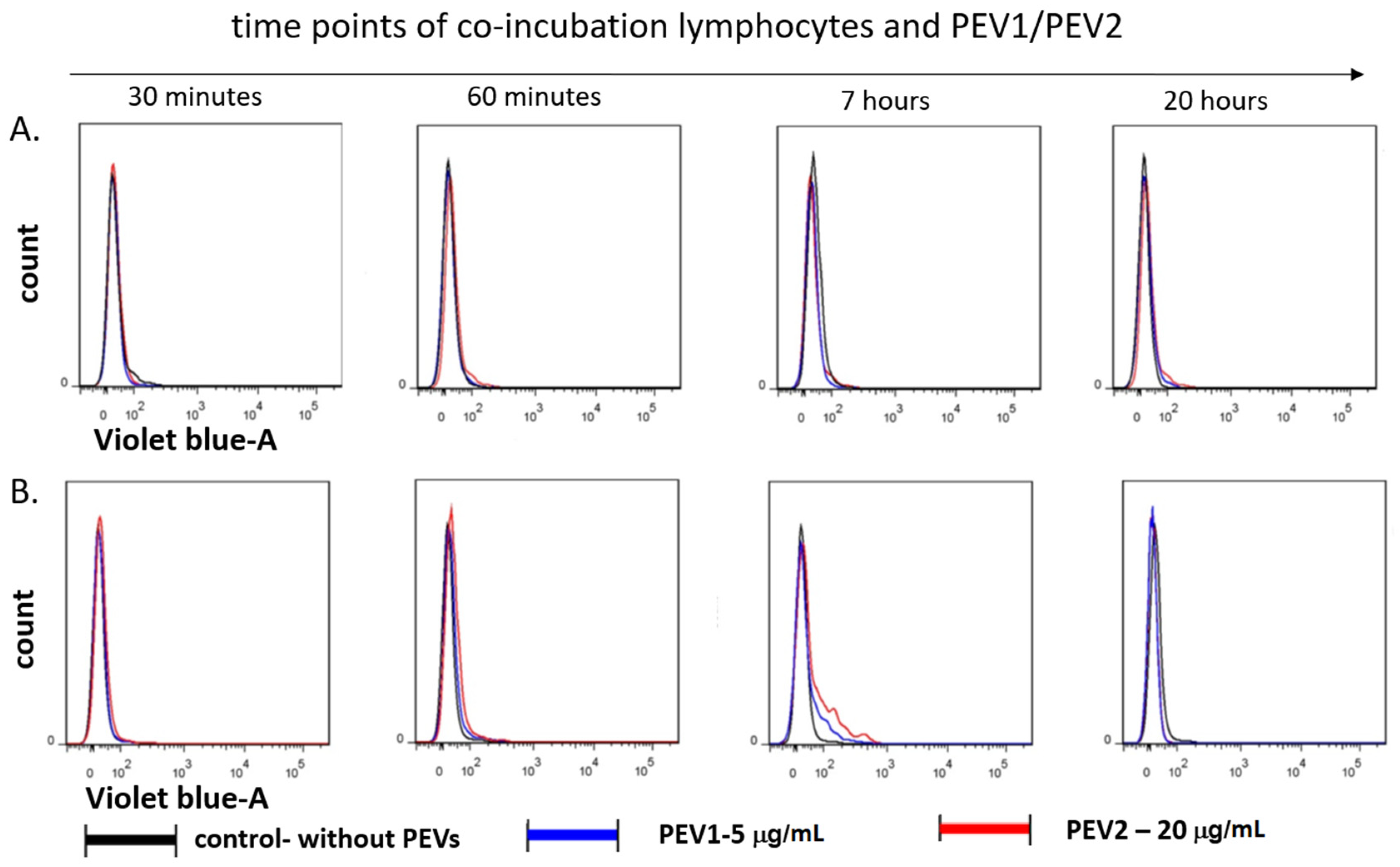
2.1. Platelet-Derived Extracellular Vesicles (PEVs) Are Taken up By Lymphocytes
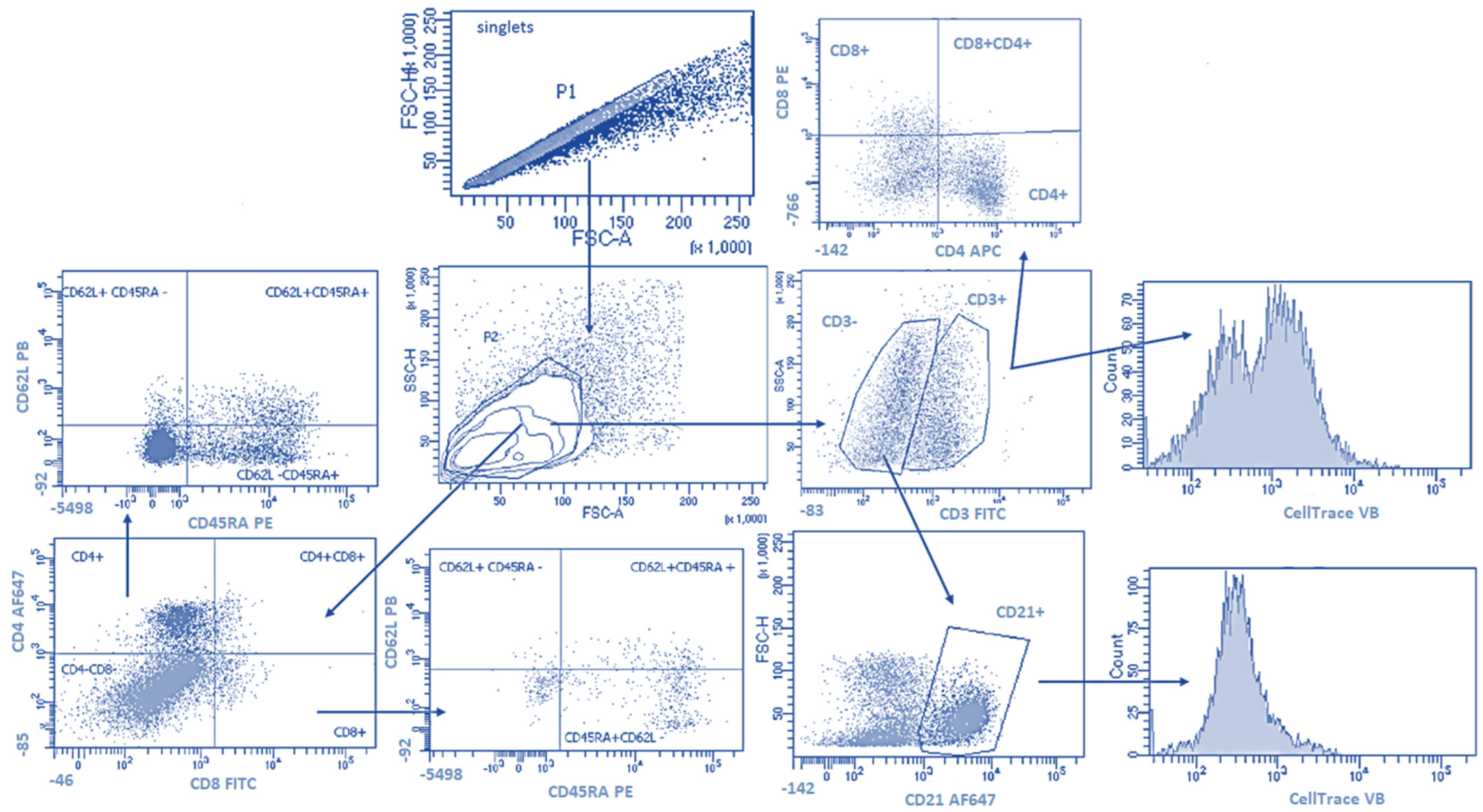
2.2. PEV Interactions with Lymphocyte Subsets, Their Proliferation and Cytokine Production
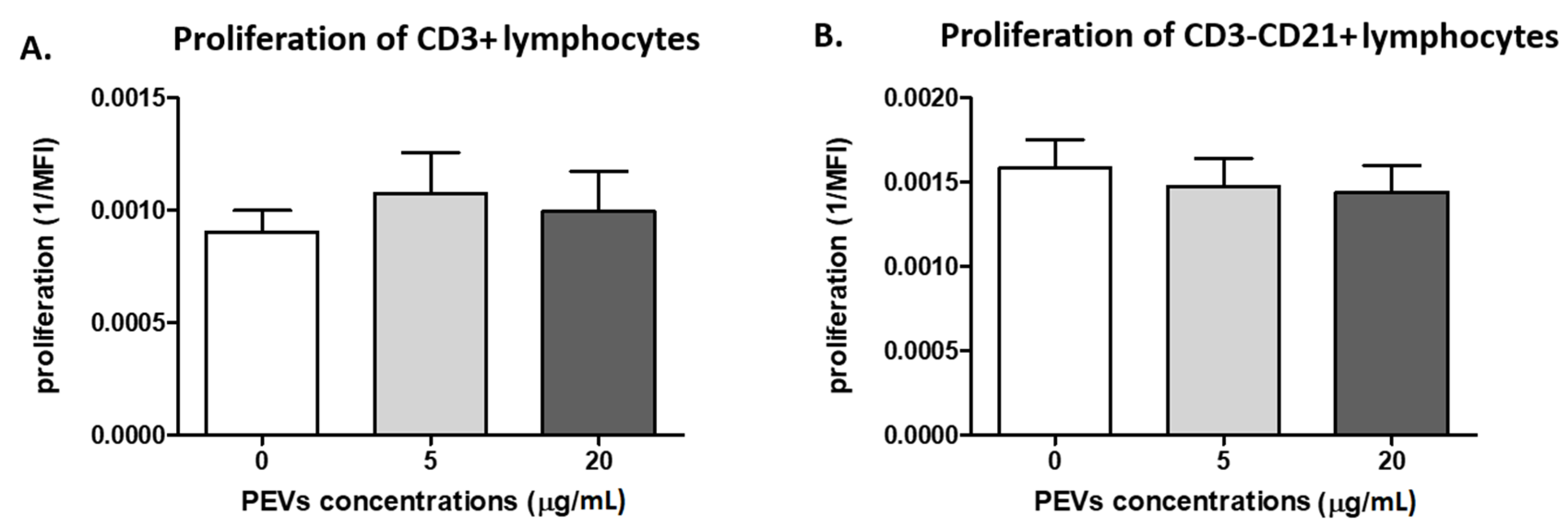
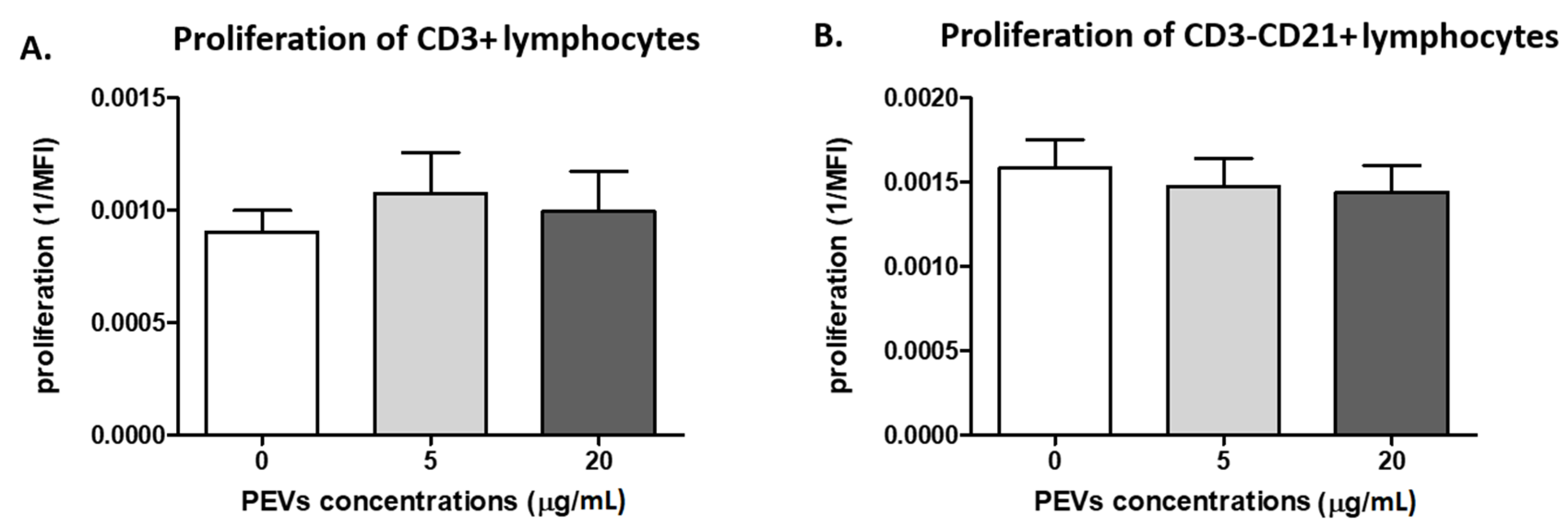
2.2.1. PEV Does Not Influence Lymphocyte Proliferation
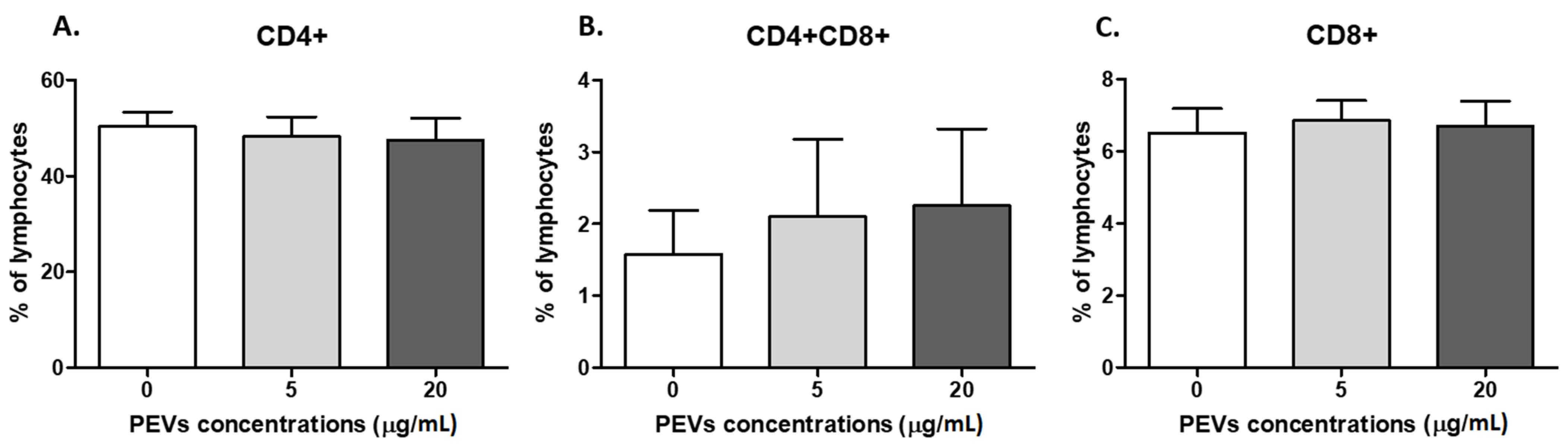
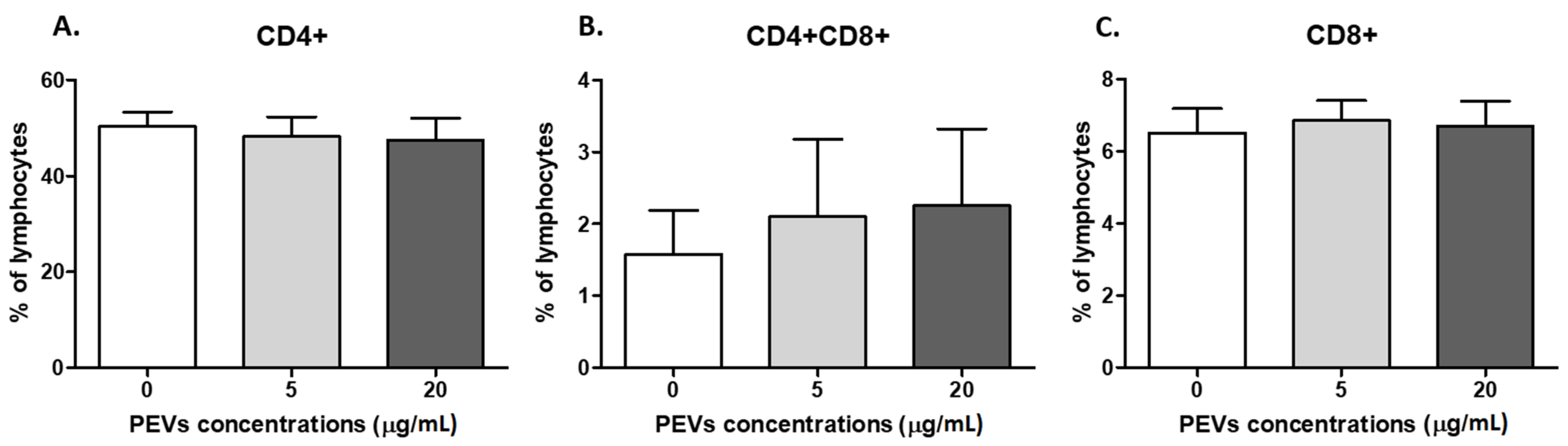
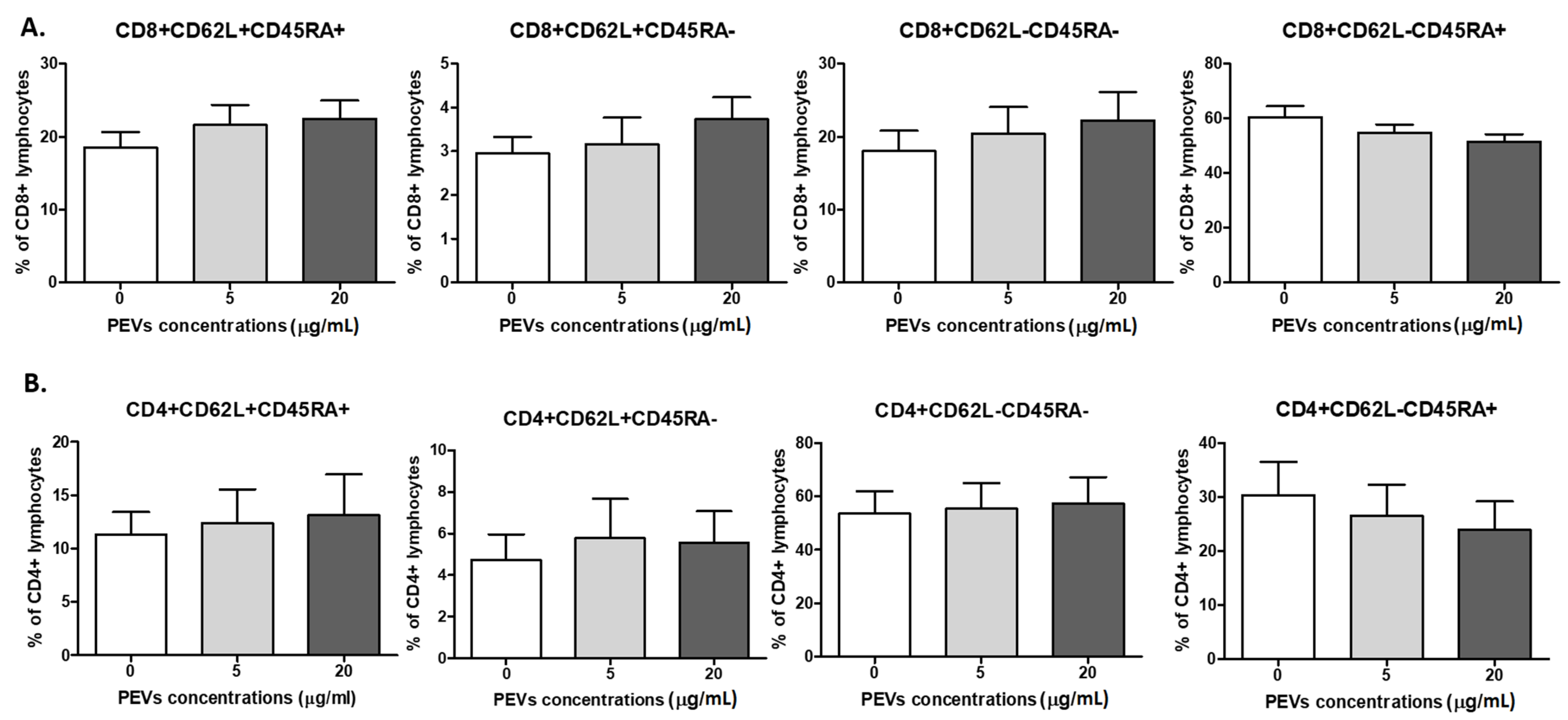
2.2.2. PEV Has No Impact on Lymphocyte Immunophenotype
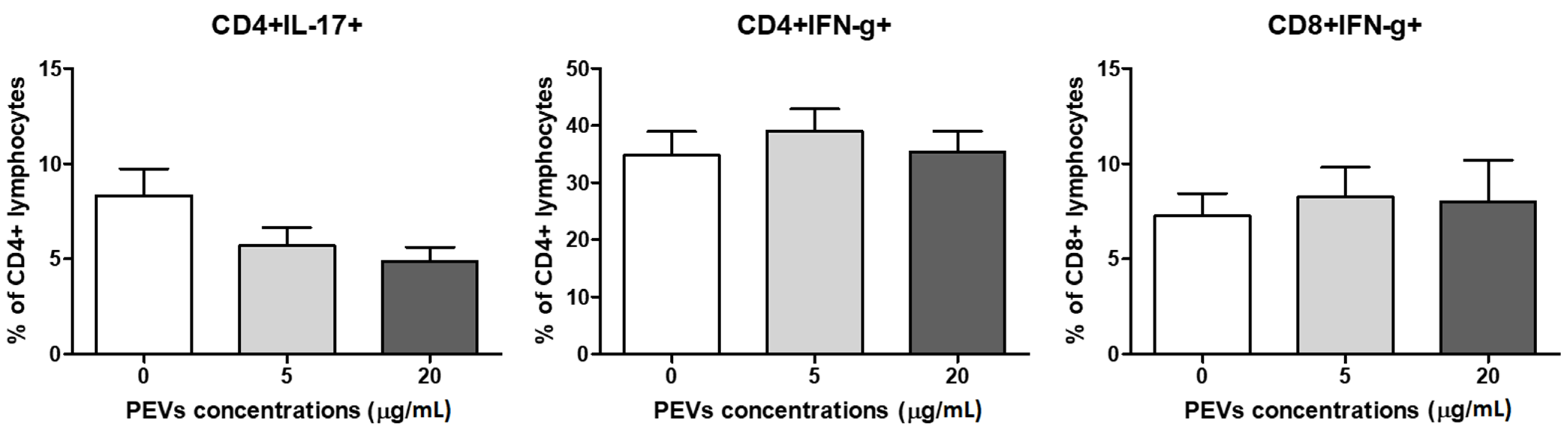
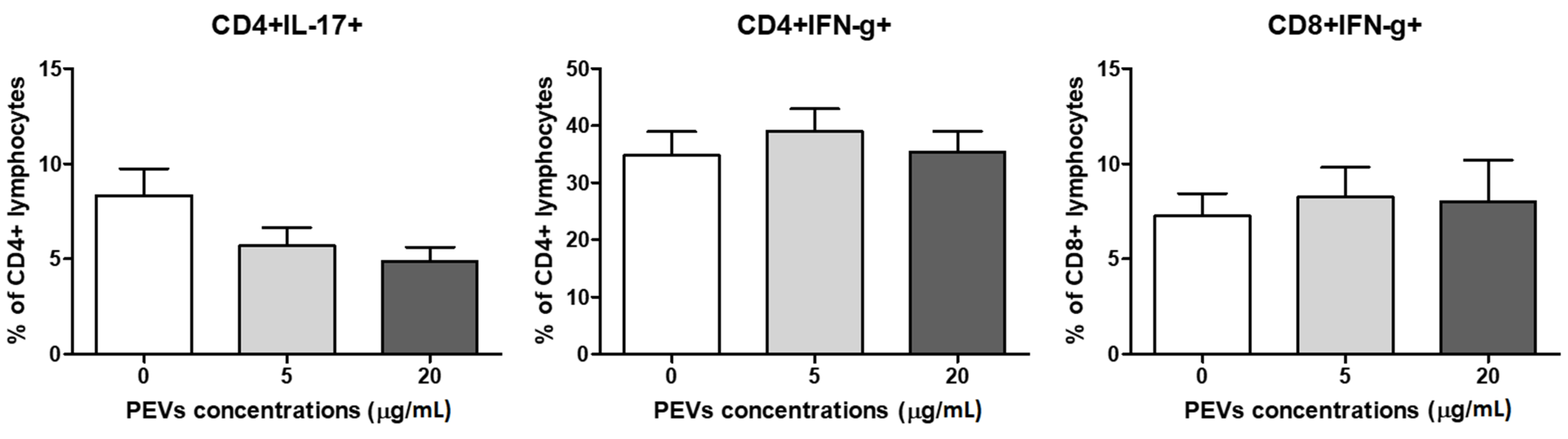
2.2.3. PEV Does Not Influence Lymphocytes T Cytokine Production
3. Discussion
4. Materials and Methods
4.1. Animals and Blood Samples
4.2. PEV Isolation and Staining
4.3. Cells Isolation and Culture
4.4. Cell Staining
4.5. Flow Cytometry Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ratajczak, M.Z.; Ratajczak, J. Extracellular microvesicles/exosomes: Discovery, disbelief, acceptance, and the future? Leukemia 2020, 34, 3126–3135. [Google Scholar] [CrossRef] [PubMed]
- Maumus, M.; Rozier, P.; Boulestreau, J.; Jorgensen, C.; Noël, D. Mesenchymal Stem Cell-Derived Extracellular Vesicles: Opportunities and Challenges for Clinical Translation. Front. Bioeng. Biotechnol. 2020, 10, 997. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the international society for extracellular vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazar, S.; Goldfinger, L.E. Platelets and extracellular vesicles and their cross talk with cancer. Blood 2021, 10, 3192–3200. [Google Scholar] [CrossRef] [PubMed]
- Wojtukiewicz, M.Z.; Sierko, E.; Hempel, D.; Tucker, S.C.; Honn, K. Platelets and cancer angiogenesis nexus. Cancer Metastasis Rev. 2017, 2, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Tableros, V.; Gomez, Y.; Camussi, G.; Brizzi, M.F. Extracellular vesicles: New players in lymphomas. Int. J. Mol. Sci. 2018, 21, 41. [Google Scholar] [CrossRef] [Green Version]
- Żmigrodzka, M.; Guzera, M.; Miśkiewicz, A.; Jagielski, D.; Winnicka, A. The biology of extracellular vesicles with focus on platelet microparticles and their role in cancer development and progression. Tumour Biol. 2016, 11, 14391–14401. [Google Scholar] [CrossRef] [Green Version]
- Sedgwick, A.E.; D’Souza-Schorey, C. The biology of extracellular microvesicles. Traffic 2018, 19, 319–327. [Google Scholar] [CrossRef]
- Stahl, P.D.; Raposo, G. Extracellular vesicles: Exosomes and microvesicles, integrators of homeostasis. Physiology 2019, 3, 169–177. [Google Scholar] [CrossRef]
- Ratajczak, J.; Wysoczynski, M.; Hayek, F.; Janowska-Wieczorek, A.; Ratajczak, M.Z. Membrane-derived microvesicles: Important and underappreciated mediators of cell-to-cell communication. Leukemia 2006, 20, 1487–1495. [Google Scholar] [CrossRef]
- Menck, K.; Bleckmann, A.; Wachter, A.; Hennies, B.; Ries, L.; Schulz, M.; Balkenhol, M.; Pukrop, T.; Schatlo, B.; Rost, U.; et al. Characterisation of tumour-derived microvesicles in cancer patients’ blood and correlation with clinical outcome. J. Extracell. Vesicles 2017, 1, 1340745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menck, K.; Sivaloganathan, S.; Bleckmann, A.; Binder, C. Microvesicles in Cancer: Small Size, Large Potential. Int. J. Mol. Sci. 2020, 28, 5373. [Google Scholar] [CrossRef] [PubMed]
- Żmigrodzka, M.; Witkowska-Piłaszewicz, O.; Winnicka, A. Platelets Extracellular Vesicles as Regulators of Cancer Progression-An Updated Perspective. Int. J. Mol. Sci. 2020, 22, 5195. [Google Scholar] [CrossRef] [PubMed]
- Laroche, M.; Dunois, C.; Vissac, A.M.; Amiral, J. Update on functional and genetic laboratory assays for the detection of platelet microvesicles. Platelets 2017, 3, 235–241. [Google Scholar] [CrossRef]
- Wolf, P. The nature and significance of platelet products in human plasma. Br. J. Haematol. 1967, 3, 269–288. [Google Scholar] [CrossRef]
- Melki, I.; Tessandier, N.; Zufferey, A.; Boilard, E. Platelet microvesicles in health and disease. Platelets 2017, 3, 214–221. [Google Scholar] [CrossRef]
- van der Pol, E.; Harrison, P. From platelet dust to gold dust: Physiological importance and detection of platelet microvesicles. Platelets 2017, 3, 211–213. [Google Scholar] [CrossRef] [Green Version]
- Aatonen, M.; Grönholm, M.; Siljander, P.R. Platelet-derived microvesicles: Multitalented participants in intercellular communication. Semin Thromb. Hemost. 2012, 1, 102–113. [Google Scholar] [CrossRef]
- Puhm, F.; Boilard, E.; Machlus, K.R. Platelet Extracellular Vesicles: Beyond the Blood. Arterioscler Thromb. Vasc. Biol. 2021, 1, 87–96. [Google Scholar] [CrossRef]
- Snir, O.; Wilsgård, L.; Latysheva, N.; Wahlund, C.J.E.; Braekkan, S.K.; Hindberg, K.; Hansen, J.B. Plasma levels of platelet-derived microvesicles are associated with risk of future venous thromboembolism. J. Thromb. Haemost. 2022, 4, 899–908. [Google Scholar] [CrossRef]
- Anene, C.; Graham, A.M.; Boyne, J.; Roberts, W. Platelet microparticle delivered microRNA-Let-7a promotes the angiogenic switch. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 8, 2633–2643. [Google Scholar] [CrossRef] [PubMed]
- Lazar, S.; Goldfinger, L.E. Platelet Microparticles and miRNA Transfer in Cancer Progression: Many Targets, Modes of Action, and Effects Across Cancer Stages. Front. Cardiovasc. Med. 2018, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Baj-Krzyworzeka, M.; Majka, M.; Pratico, D.; Ratajczak, J.; Vilaire, G.; Kijowski, J.; Reca, R.; Janowska-Wieczorek, A.; Ratajczak, M.Z. Platelet-derived microparticles stimulate proliferation, survival, adhesion, and chemotaxis of hematopoietic cells. Exp. Hematol. 2002, 5, 450–459. [Google Scholar] [CrossRef]
- Dovizio, M.; Bruno, A.; Contursi, A.; Grande, R.; Patrignani, P. Platelets and extracellular vesicles in cancer: Diagnostic and therapeutic implications. Cancer Metastasis Rev. 2018, 37, 455–467. [Google Scholar] [CrossRef] [PubMed]
- Dymicka-Piekarska, V.; Gryko, M.; Lipska, A.; Korniluk, A.; Siergiejko, E.; Kemona, H. Platelet-derived microparticles in patients with colorectal cancer. J. Cancer Ther. 2012, 6, 898–901. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Qi, J.; Zhao, S.; Shen, W.; Dai, L.; Han, W.; Huang, M.; Wang, Z.; Ruan, C.; Wu, D.; et al. Clinical significance of circulating microparticles in Ph- myeloproliferative neoplasms. Oncol. Lett. 2017, 2, 2531–2536. [Google Scholar] [CrossRef] [Green Version]
- Bjørnetrø, T.; Steffensen, L.A.; Vestad, B.; Brusletto, B.S.; Olstad, O.K.; Trøseid, A.M.; Aass, H.C.D.; Haug, K.B.F.; Llorente, A.; Bøe, S.O.; et al. Uptake of circulating extracellular vesicles from rectal cancer patients and differential responses by human monocyte cultures. FEBS Open Bio 2021, 11, 724–740. [Google Scholar] [CrossRef]
- Czystowska-Kuzmicz, M.; Sosnowska, A.; Nowis, D.; Ramji, K.; Szajnik, M.; Chlebowska-Tuz, J.; Wolinska, E.; Gaj, P.; Grazul, M.; Pilch, Z.; et al. Small extracellular vesicles containing arginase-1 suppress T-cell responses and promote tumor growth in ovarian carcinoma. Nat. Commun. 2019, 5, 3000. [Google Scholar] [CrossRef]
- Pando, A.; Reagan, J.L.; Quesenberry, P.; Fast, L.D. Extracellular vesicles in leukemia. Leuk Res. 2018, 64, 52–60. [Google Scholar] [CrossRef]
- Hazan-Halevy, I.; Rosenblum, D.; Weinstein, S.; Bairey, O.; Raanani, P.; Peer, D. Cell-specific uptake of mantle cell lymphoma-derived exosomes by malignant and non-malignant B-lymphocytes. Cancer Lett. 2015, 1, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Yenigürbüz, F.D.; Kızmazoğlu, D.; Ateş, H.; Erdem, M.; Tufekci, O.; Yilmaz, S.; Oren, H. Analysis of apoptotic, platelet-derived, endothelial-derived, and tissue factor-positive microparticles of children with acute lymphoblastic leukemia during induction therapy. Blood Coagul. Fibrinolysis 2019, 4, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Yao, B.; Qu, S.; Hu, R.; Gao, W.; Jin, S.; Ju, J.; Zhao, Q. Delivery of platelet TPM3 mRNA into breast cancer cells via microvesicles enhances metastasis. FEBS Open Bio 2019, 12, 2159–2169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaftian, M.; Yari, F.; Ghasemzadeh, M.; Fallah, A.V.; Haghighi, M. Induction of apoptosis in cancer cells of pre-B ALL patients after exposure to platelets, platelet-derived microparticles and soluble CD40 ligand. Cell J. 2018, 1, 120–126. [Google Scholar]
- Michael, J.V.; Wurtzel, J.G.T.; Mao, G.F.; Rao, A.K.; Kolpakov, M.A.; Sabri, A.; Hoffman, N.E.; Rajan, S.; Tomar, D.; Madesh, M.; et al. Platelet microparticles infiltrating solid tumors transfer miRNAs that suppress tumor growth. Blood 2017, 5, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saber, S.H.; Ali, H.E.A.; Gaballa, R.; Gaballah, M.; Ali, H.I.; Zerfaoui, M.; Abd Elmageed, Z.Y. Exosomes are the driving force in preparing the soil for the metastatic seeds: Lessons from the prostate cancer. Cells 2020, 3, 564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, L.M.; Wang, M.Z. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells 2019, 7, 727. [Google Scholar] [CrossRef] [Green Version]
- Castaman, G.; Yu-Feng, L.; Battistin, E.; Rodeghiero, F. Characterization of a novel bleeding disorder with isolated prolonged bleeding time and deficiency of platelet microvesicle generation. Br. J. Haematol. 1997, 96, 458–463. [Google Scholar] [CrossRef]
- Berckmans, R.J.; Nieuwland, R.; Böing, A.N.; Romijn, F.P.; Hack, C.E.; Sturk, A. Cell-derived microparticles circulate in healthy humans and support low grade thrombin generation. Thromb. Haemost. 2001, 4, 639–646. [Google Scholar]
- Żmigrodzka, M.; Witkowska-Piłaszewicz, O.; Rzepecka, A.; Cywińska, A.; Jagielski, D.; Winnicka, A. Extracellular Vesicles in the Blood of Dogs with Cancer-A Preliminary Study. Animals 2019, 19, 575. [Google Scholar] [CrossRef] [Green Version]
- Spakova, T.; Janockova, J.; Rosocha, J. Characterization and Therapeutic Use of Extracellular Vesicles Derived from Platelets. Int. J. Mol. Sci. 2021, 18, 9701. [Google Scholar] [CrossRef]
- Jayachandran, M.; Miller, V.M.; Heit, J.A.; Owen, W.G. Methodology for isolation, identification and characterization of microvesicles in peripheral blood. J. Immunol. Methods 2012, 375, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, R.; Gröger, M.; Rauscher, S.; Fendl, B.; Eichhorn, T.; Fischer, M.B.; Spittler, A.; Weber, V. Differential Interaction of Platelet-Derived Extracellular Vesicles with Leukocyte Subsets in Human Whole Blood. Sci. Rep. 2018, 1, 6598. [Google Scholar] [CrossRef] [PubMed]
- Bebesi, T.; Kitka, D.; Gaál, A.; Szigyártó, I.C.; Deák, R.; Beke-Somfai, T.; Koprivanacz, K.; Juhász, T.; Bóta, A.; Varga, Z.; et al. Storage conditions determine the characteristics of red blood cell derived extracellular vesicles. Sci. Rep. 2022, 1, 977. [Google Scholar] [CrossRef] [PubMed]
- Wisgrill, L.; Lamm, C.; Hartmann, J.; Preißing, F.; Dragosits, K.; Bee, A.; Hell, L.; Thaler, J.; Ay, C.; Pabinger, I.; et al. Peripheral blood microvesicles secretion is influenced by storage time, temperature, and anticoagulants. Cytom. Part A 2016, 89, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Cauwenberghs, S.; Feijge, M.A.; Harper, A.G.; Sage, S.O.; Curvers, J.; Heemskerk, J.W. Shedding of procoagulant microparticles from unstimulated platelets by integrin-mediated destabilization of actin cytoskeleton. FEBS Lett. 2006, 580, 5313–5320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazawa, B.; Trivedi, A.; Togarrati, P.P.; Potter, D.; Baimukanova, G.; Vivona, L.; Lin, M.; Lopez, E.; Callcut, R.; Srivastava, A.; et al. Regulation of endothelial cell permeability by platelet-derived extracellular vesicles. J. Trauma Acute Care Surg. 2019, 86, 931–942. [Google Scholar] [CrossRef]
- Hess, J.R.; Lelkens, C.C.; Holcomb, J.B.; Scalea, T.M. Advances in military, field, and austere transfusion medicine in the last decade. Transfus. Apher. Sci. 2013, 49, 380–386. [Google Scholar] [CrossRef]
- Vasina, E.M.; Cauwenberghs, S.; Staudt, M.; Feijge, M.A.; Weber, C.; Koenen, R.R.; Heemskerk, J.W. Aging- and activation-induced platelet microparticles suppress apoptosis in monocytic cells and differentially signal to proinflammatory mediator release. Am. J. Blood Res. 2013, 2, 107–123. [Google Scholar]
- Guo, J.; Feng, C.; Zhang, B.; Zhang, S.; Shen, X.; Zhu, J.; Zhao, X. Extraction and identification of platelet-derived microparticles. Mol. Med. Rep. 2019, 20, 2916–2921. [Google Scholar] [CrossRef]
- Aatonen, M.T.; Ohman, T.; Nyman, T.A.; Laitinen, S.; Grönholm, M.; Siljander, P.R. Isolation and characterization of platelet-derived extracellular vesicles. J. Extracell. Vesicles 2014, 3, 24692. [Google Scholar] [CrossRef]
- Italiano, J.E., Jr.; Mairuhu, A.T.; Flaumenhaft, R. Clinical relevance of microparticles from platelets and megakaryocytes. Curr. Opin. Hematol. 2010, 6, 578–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heijnen, H.F.; Schiel, A.E.; Fijnheer, R.; Geuze, H.J.; Sixma, J.J. Activated platelets release two types of membrane vesicles: Microvesicles by surface shedding and exosomes derived from exocytosis of multivesicular bodies and alpha-granules. Blood 1999, 11, 3791–3799. [Google Scholar] [CrossRef]
- Ferreira, P.M.; Bozbas, E.; Tannetta, S.D.; Alroqaiba, N.; Zhou, R.; Crawley, J.T.B.; Gibbins, J.M.; Jones, C.I.; Ahnström, J.; Yaqoob, P. Mode of induction of platelet-derived extracellular vesicles is a critical determinant of their phenotype and function. Sci. Rep. 2020, 1, 18061. [Google Scholar] [CrossRef] [PubMed]
- Arraud, N.; Linares, R.; Tan, S.; Gounou, C.; Pasquet, J.M.; Mornet, S.; Brisson, A.R. Extracellular vesicles from blood plasma: Determination of their morphology, size, phenotype and concentration. J. Thromb. Haemost. 2014, 5, 614–627. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.K.; Le, A.; Chavakis, T.; Rumbaut, R.E.; Thiagarajan, P. Developmental endothelial locus-1 (Del-1) mediates clearance of platelet microparticles by the endothelium. Circulation 2012, 13, 1664–1672. [Google Scholar] [CrossRef] [Green Version]
- Namba, M.; Tanaka, A.; Shimada, K.; Ozeki, Y.; Uehata, S.; Sakamoto, T.; Nishida, Y.; Nomura, S.; Yoshikawa, J. Circulating platelet-derived microparticles are associated with atherothrombotic events: A marker for vulnerable blood. Arterioscler. Thromb. Vasc. Biol. 2007, 1, 255–256. [Google Scholar] [CrossRef]
- Fendl, B.; Weiss, R.; Fischer, M.B.; Spittler, A.; Weber, V. Characterization of extracellular vesicles in whole blood: Influence of pre-analytical parameters and visualization of vesicle-cell interactions using imaging flow cytometry. Biochem. Biophys. Res. Commun. 2016, 1, 168–173. [Google Scholar] [CrossRef]
- Dinkla, S.; van Cranenbroek, B.; van der Heijden, W.A.; He, X.; Wallbrecher, R.; Dumitriu, I.E.; van der Ven, A.J.; Bosman, G.J.; Koenen, H.J.; Joosten, I. Platelet microparticles inhibit IL-17 production by regulatory T cells through P-selectin. Blood 2016, 21, 1976–1986. [Google Scholar] [CrossRef] [Green Version]
- Sadallah, S.; Amicarella, F.; Eken, C.; Iezzi, G.; Schifferli, J.A. Ectosomes released by platelets induce differentiation of CD4+T cells into T regulatory cells. Thromb. Haemost. 2014, 112, 1219–1229. [Google Scholar]
- Waqas, M.Y.; Javid, M.A.; Nazir, M.M.; Niaz, N.; Nisar, M.F.; Manzoor, Z.; Bhatti, S.A.; Hameed, S.; Khaliq, M.H. Extracellular vesicles and exosome: Insight from physiological regulatory perspectives. J. Physiol. Biochem. 2022. online ahead of print. [Google Scholar] [CrossRef]
- Flaumenhaft, R.; Dilks, J.R.; Richardson, J.; Alden, E.; Patel-Hett, S.R.; Battinelli, E.; Klement, G.L.; Sola-Visner, M.; Italiano, J.E., Jr. Megakaryocyte-derived microparticles: Direct visualization and distinction from platelet-derived microparticles. Blood 2009, 5, 1112–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moritz, A.; Walcheck, B.K.; Weiss, D.J. Flow cytometric detection of activated platelets in the dog. Vet. Clin. Pathol. 2003, 32, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Żmigrodzka, M.; Guzera, M.; Winnicka, A. Flow cytometric assessment of activation of peripheral blood platelets in dogs with normal platelet count and asymptomatic thrombocytopenia. Pol. J. Vet. Sci. 2016, 19, 407–414. [Google Scholar] [CrossRef]
- Czernek, L.; Chworos, A.; Duechler, M. The Uptake of Extracellular Vesicles is Affected by the Differentiation Status of Myeloid Cells. Scand. J. Immunol. 2015, 82, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhys, H.I.; Dell’Accio, F.; Pitzalis, C.; Moore, A.; Norling, L.V.; Perretti, M. Neutrophil Microvesicles from Healthy Control and Rheumatoid Arthritis Patients Prevent the Inflammatory Activation of Macrophages. EBioMedicine 2018, 29, 60–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Clone | Isotype | Host Species | Fluorochrome | Catalog Number | Supplier | Dilution |
|---|---|---|---|---|---|---|---|
| CD3: CD8 | CA17.2A12/ YCATE55.9 | IgG1 | mouse | FITC:PE | DC047 | BioRad | 1:5 |
| CD4: CD8 | YKIX302.9/ YCATE55.9 | IgG2a | rat | FITC:PE | DC048 | BioRad | 1:5 |
| CD4 | YKIX302.9 | IgG2a | rat | AF647 | MCA1038A647 | BioRad | 1:5 |
| CD8 | YCATE55.9 | IgG1 | rat | FITC | MCA1039F | BioRad | 1:5 |
| CD21 | CA2.1D6 | IgG1 | mouse | AF647 | MCA1781A647 | BioRad | 1:5 |
| CD62L | FMC46 | IgG2b | mouse | PB | MCA1076PB | BioRad | 1:5 |
| CD45RA | CA4.1D3 | IgG1 | mouse | - | MA5-16612 | Invitrogen | 1:5 |
| IgG1 | M1-14D12 | IgG1 | rat | PE | 12-4015-82 | Invitrogen | 1:5 |
| IFNγ | CC302 | IgG1 | mouse | AF647 | MCA1783A647 | BioRad | 1:5 |
| IL-17A | eBio17B7 | IgG2a,κ | rat | PE | 12-7177-81 | Invitrogen | 1:20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Żmigrodzka, M.; Witkowska-Piłaszewicz, O.; Pingwara, R.; Winnicka, A. Platelet Extracellular Vesicles Are Taken up by Canine T Lymphocytes but Do Not Play a Role in Their Proliferation, Differentiation and Cytokine Production In Vitro. Int. J. Mol. Sci. 2022, 23, 5504. https://doi.org/10.3390/ijms23105504
Żmigrodzka M, Witkowska-Piłaszewicz O, Pingwara R, Winnicka A. Platelet Extracellular Vesicles Are Taken up by Canine T Lymphocytes but Do Not Play a Role in Their Proliferation, Differentiation and Cytokine Production In Vitro. International Journal of Molecular Sciences. 2022; 23(10):5504. https://doi.org/10.3390/ijms23105504
Chicago/Turabian StyleŻmigrodzka, Magdalena, Olga Witkowska-Piłaszewicz, Rafał Pingwara, and Anna Winnicka. 2022. "Platelet Extracellular Vesicles Are Taken up by Canine T Lymphocytes but Do Not Play a Role in Their Proliferation, Differentiation and Cytokine Production In Vitro" International Journal of Molecular Sciences 23, no. 10: 5504. https://doi.org/10.3390/ijms23105504
APA StyleŻmigrodzka, M., Witkowska-Piłaszewicz, O., Pingwara, R., & Winnicka, A. (2022). Platelet Extracellular Vesicles Are Taken up by Canine T Lymphocytes but Do Not Play a Role in Their Proliferation, Differentiation and Cytokine Production In Vitro. International Journal of Molecular Sciences, 23(10), 5504. https://doi.org/10.3390/ijms23105504