
Order vs. Disorder: Cholesterol and Omega-3 Phospholipids Determine Biomembrane Organization

 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. Composition of Lipid Mixtures
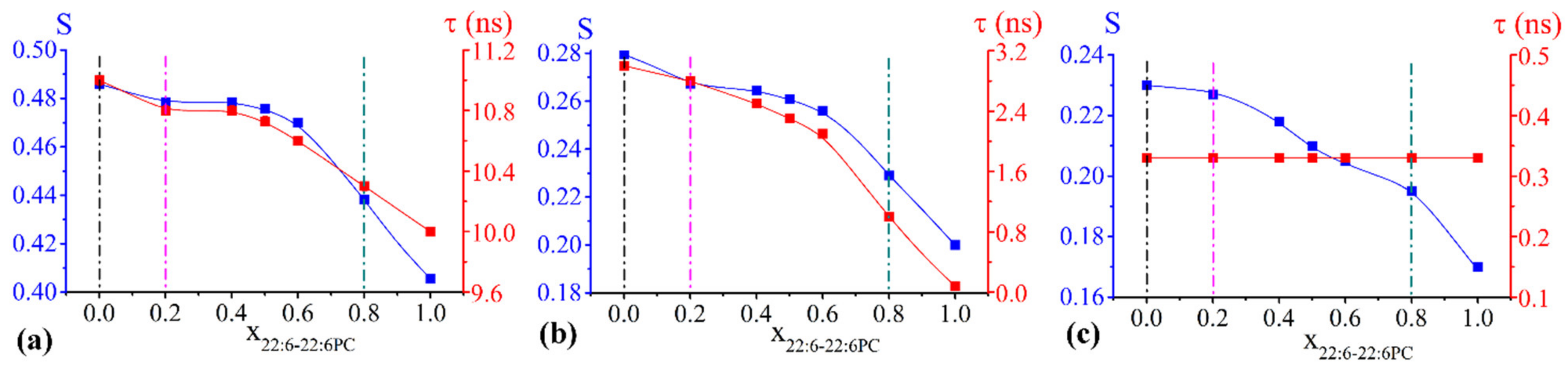
2.2. EPR Results: Microstructural Features of Vesicles Containing Omega-3 Phospholipids
2.2.1. Chol/POPC/22:6–22:6PC Lipid Bilayers
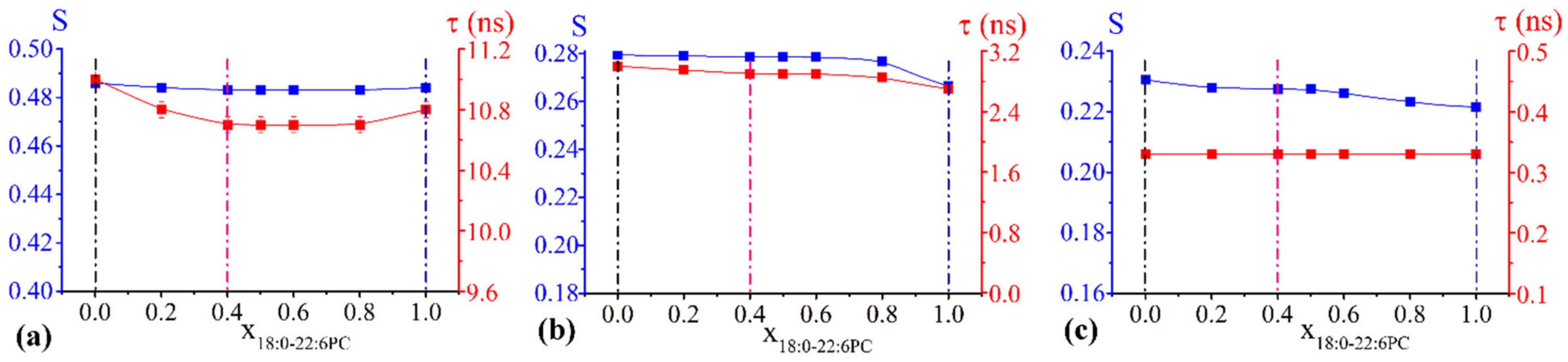
2.2.2. Chol/POPC/18:0–22:6PC Lipid Bilayers
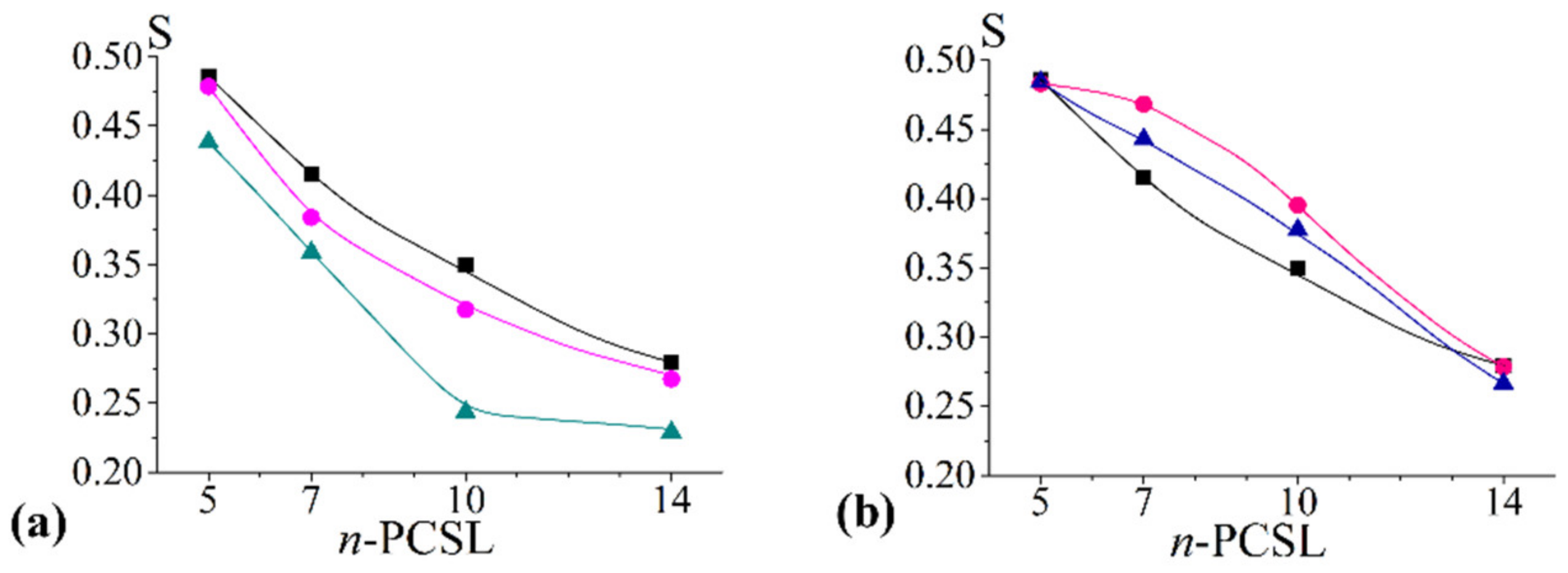
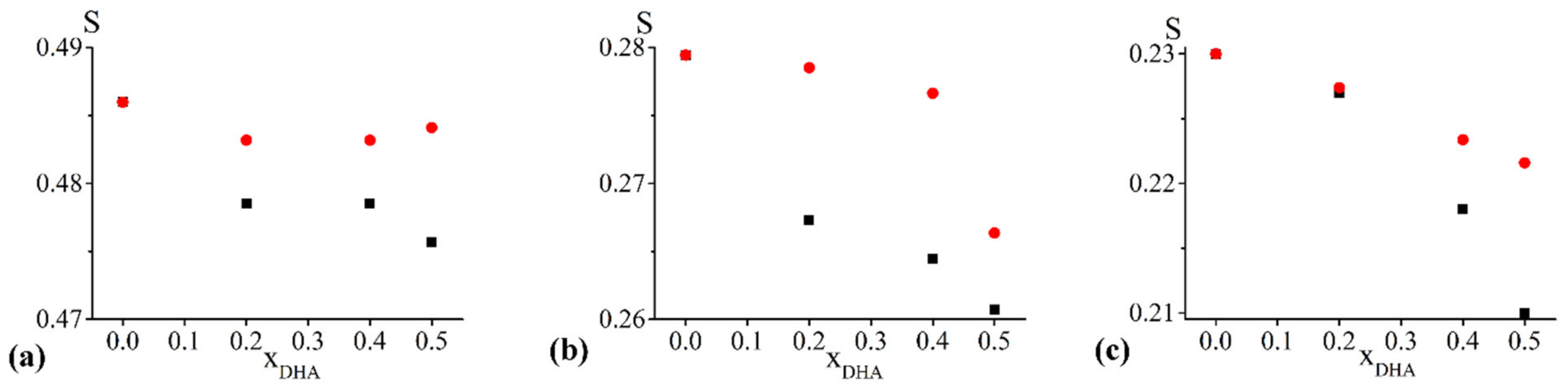
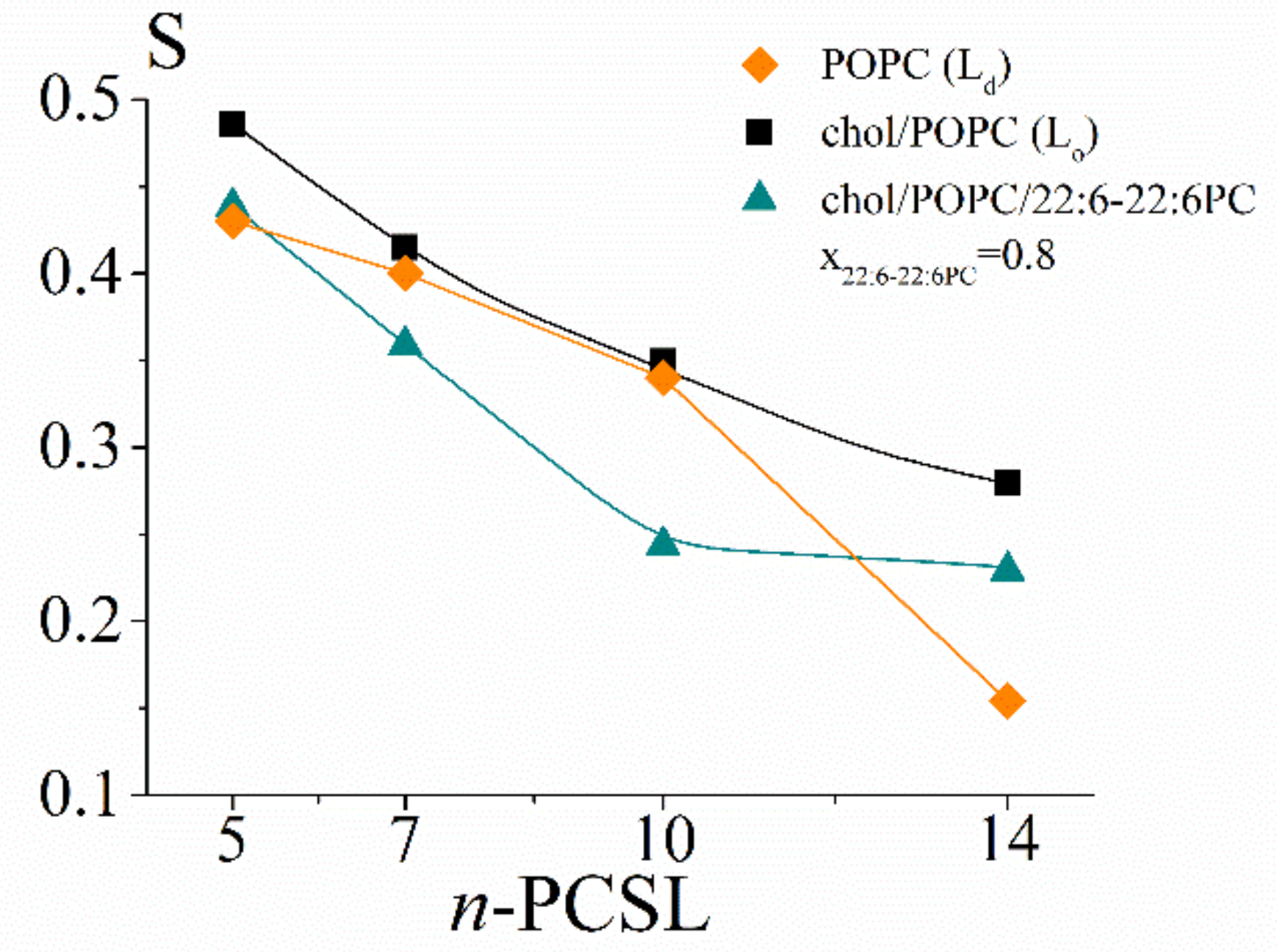
2.2.3. S Profiles
- concerning the chol/POPC/22:6–22:6PC system, we considered two lipid compositions: one well below (x22:6–22:6PC = 0.2) and the other above (x22:6–22:6PC = 0.8) the threshold (x22:6–22:6PC = 0.6) at which a dramatic change of the spectral features was observed (Figure 1);
- concerning the chol/POPC/18:0–22:6PC system, we chose the lipid mixtures with a DHA content as close as possible to those of chol/POPC/22:6–22:6PC systems, i.e., x18:0–22:6PC = 0.4 and x18:0–22:6PC = 1 (the last corresponding to chol/18:0–22:6PC system), in order to allow a direct analysis of the effects of PUFA molecular distribution (i.e., monopolyenes vs. dipolyenes)
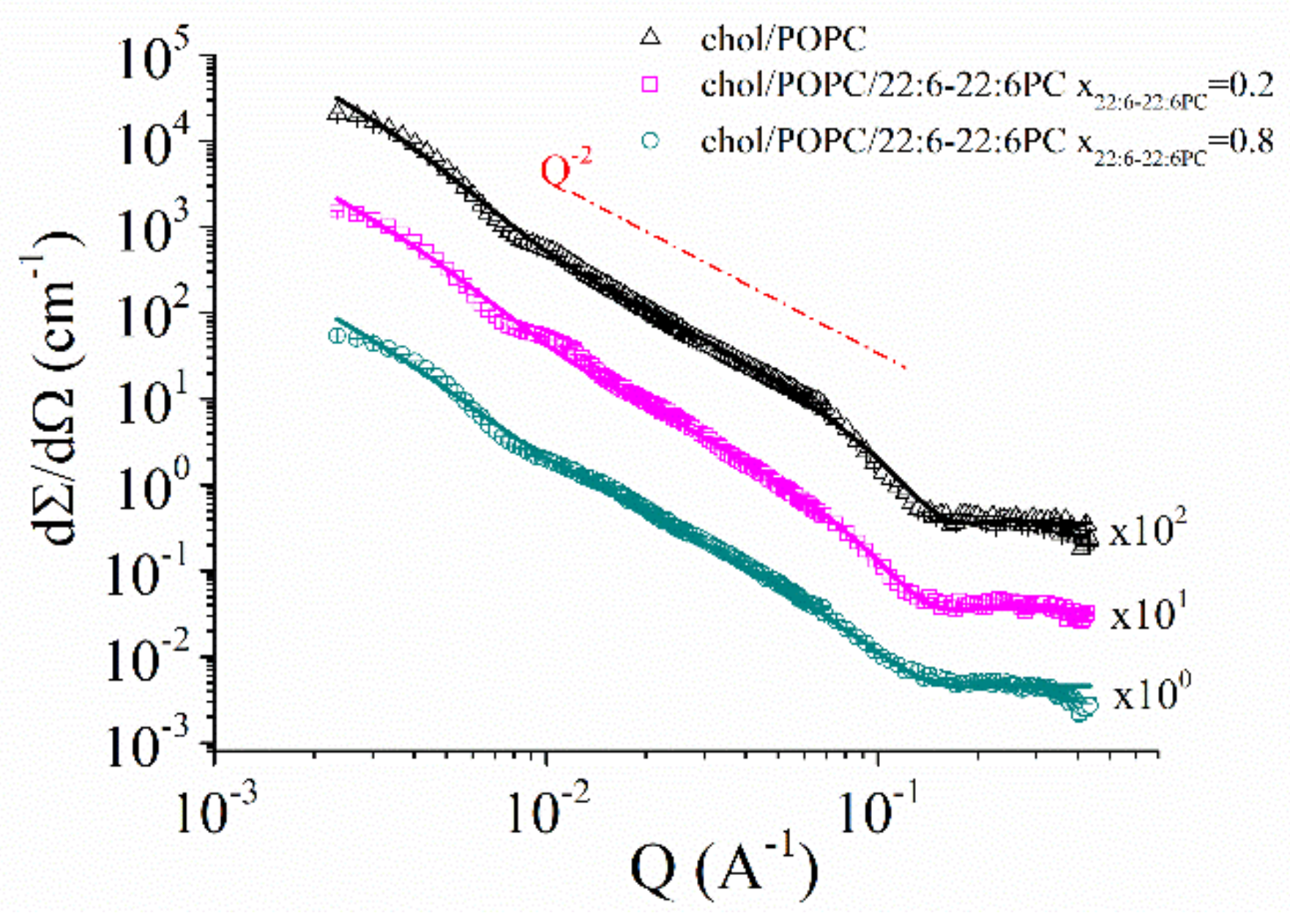
2.3. SANS Results: Morphology of chol/POPC/22:6–22:6PC Aggregates
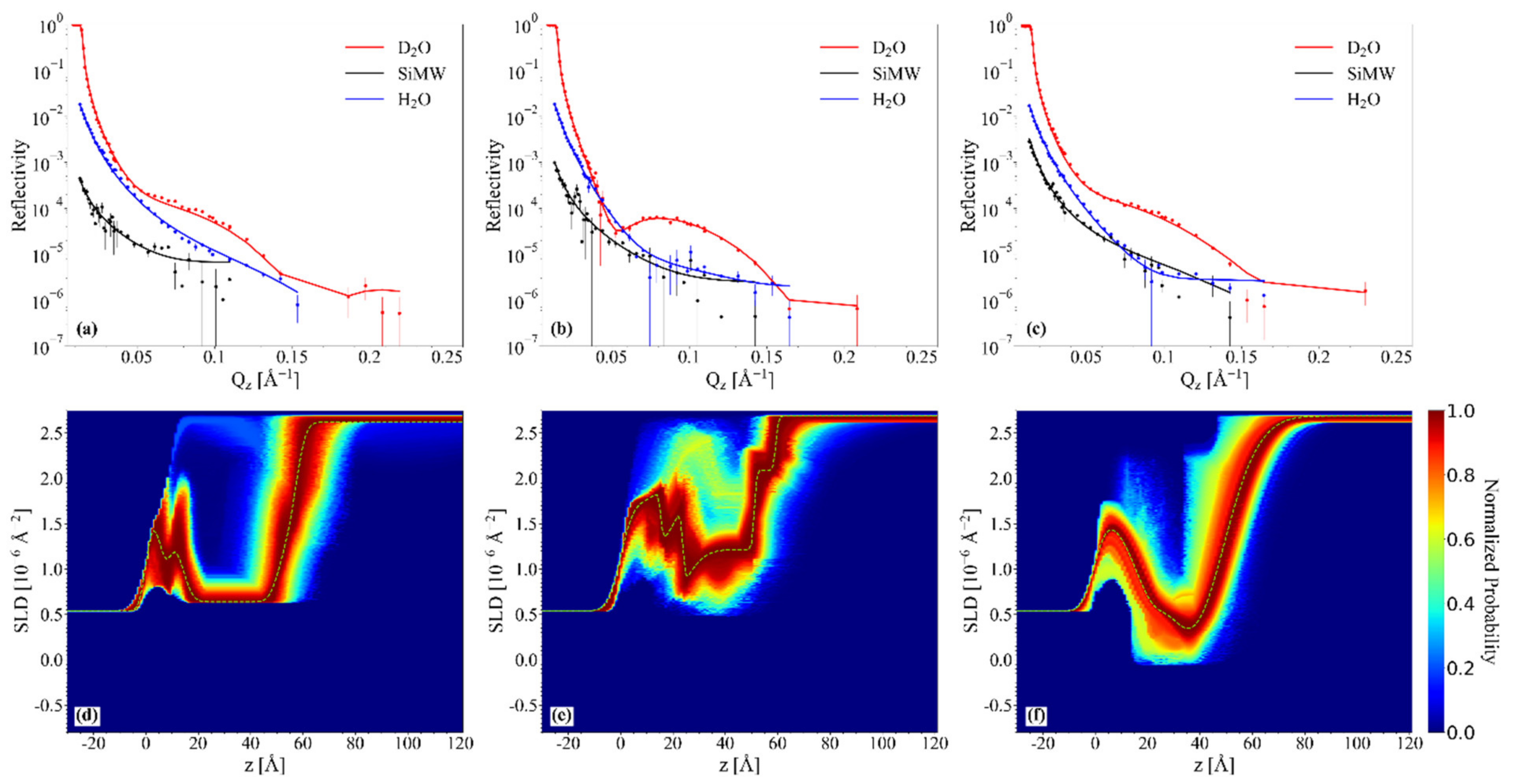
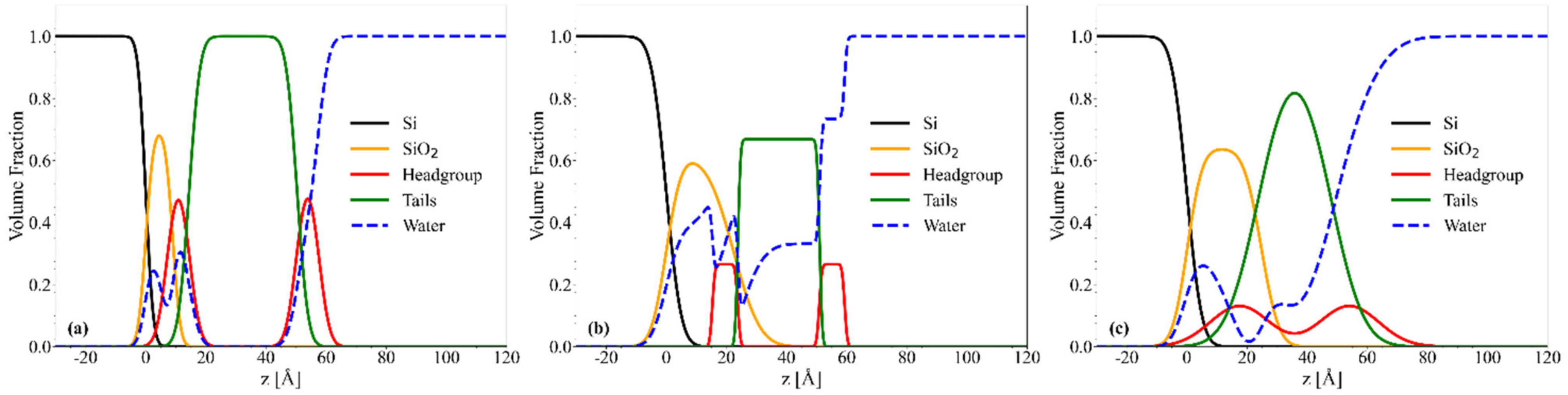
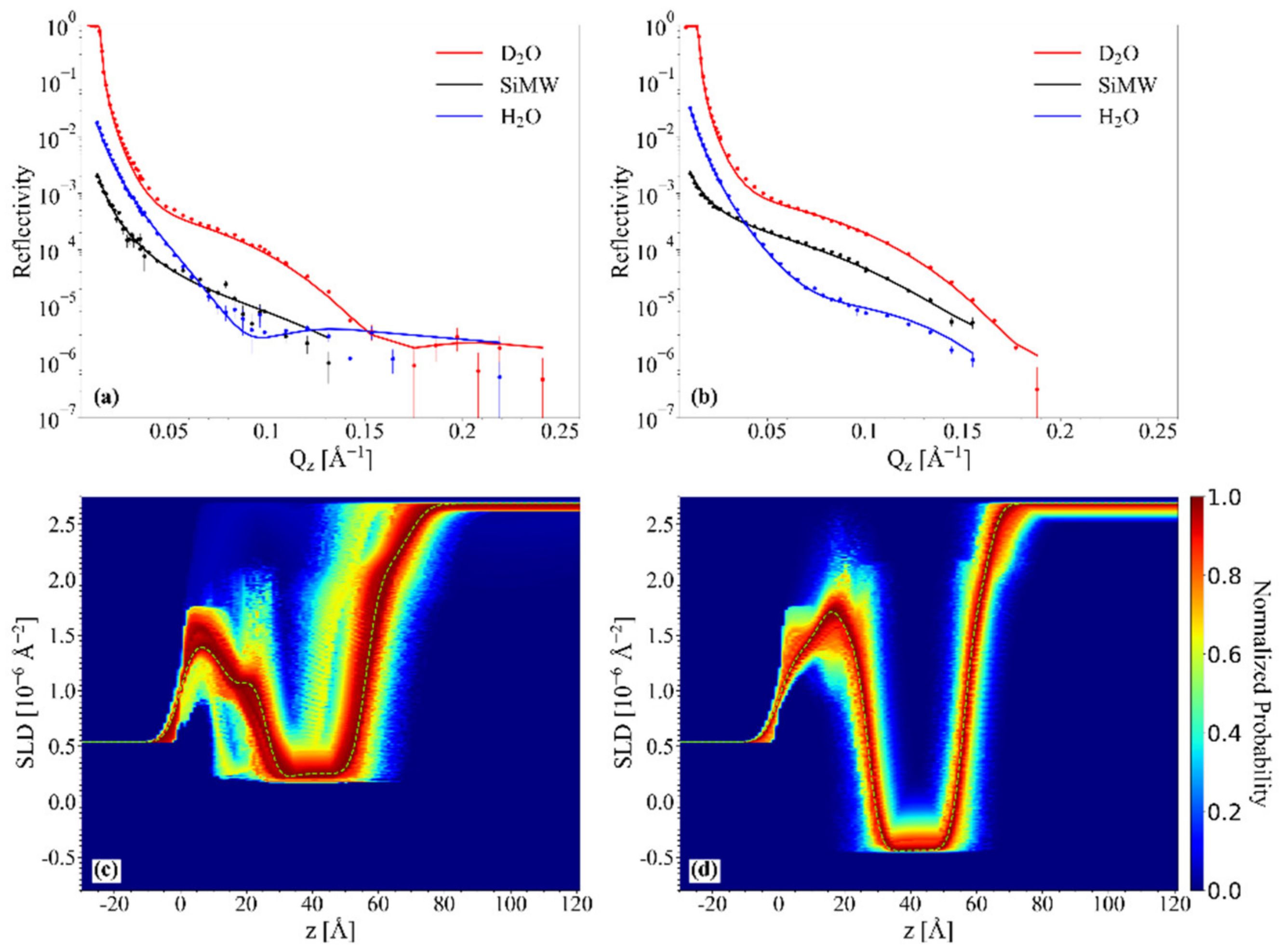
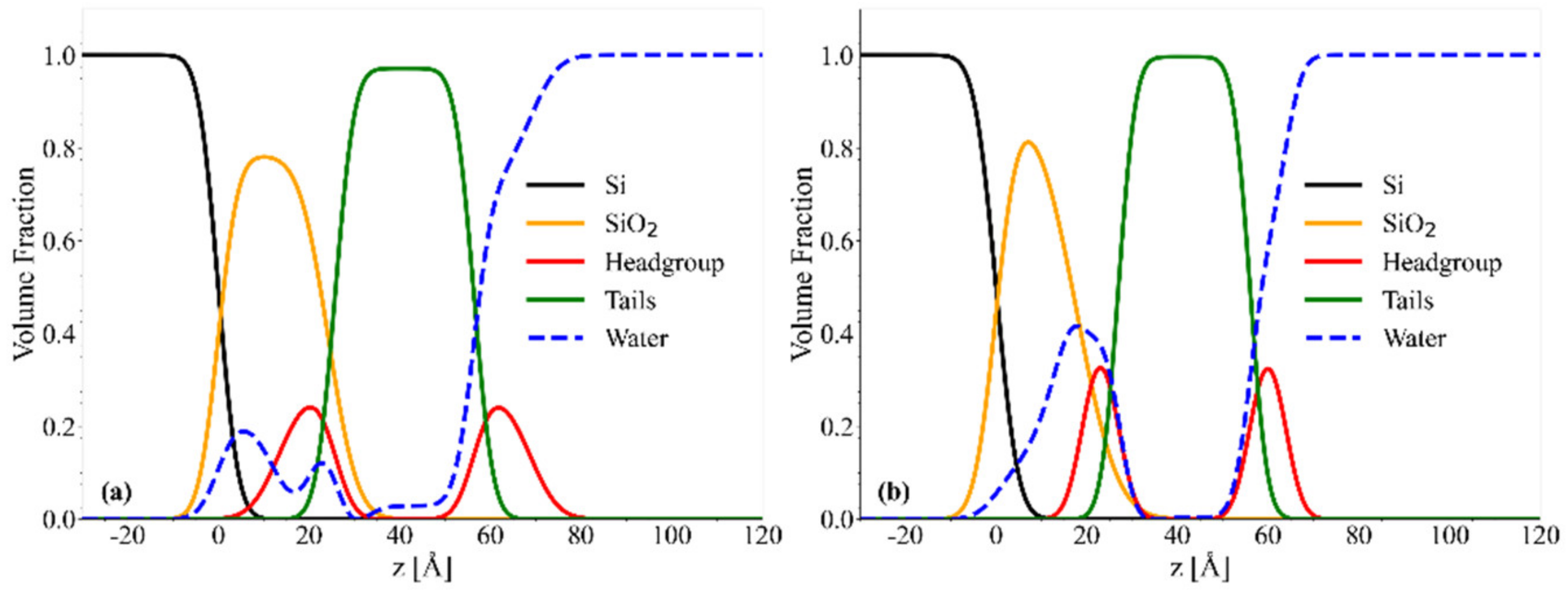
2.4. NR Results: Mesostructure of Supported Lipid Bilayers Containing Omega-3 Phospholipids
2.4.1. Chol/POPC/22:6–22:6PC
2.4.2. Chol/POPC/18:0–22:6PC
3. Discussion
3.1. The Nature of the Polyunsaturated Lipid, Rather than the Overall DHA Content, Defines the Lipid Membrane Organization
3.2. Cholesterol Suppresses Morphological Changes in 22:6–22:6PC-Rich Systems
3.3. Cholesterol Is Highly Soluble in 22:6–22:6PC-Rich Systems
3.4. Domain Formation Does Not Occur in Lipid Mixtures Formed by Cholesterol and Polyunsaturated Phosphocholines
3.5. Cholesterol Appears to Preserve a Canonical In-Leaflet Location in Polyunsaturated Systems
3.6. The Co-Presence of Cholesterol and Symmetric Polyunsaturated Phospholipids Induces the Formation of a New Disordered Lipid Organization within the Bilayer
4. Materials and Methods
4.1. Materials
4.2. Sample Preparation
4.3. Electron Paramagnetic Resonance (EPR) Spectroscopy
4.4. Analysis of EPR Spectra
- the gii values for the coupling between the electron spin and the magnetic field. Those giving the best fitting for all spectra were: gii = 2.0075, 2006, 2.003 in the case of n-PCSL spectra, and gii = 2.014, 2.006, 2.0015 in the case of CNO spectra;
- the Aii values for the coupling between the electron spin and the nitrogen nuclear spin. They were also maintained constant for each spin probe, since the same values well fitted all the spectra: Aii = 7.1 G, 7.1 G, 29.7 G, <A> = (Axx + Ayy + Azz)/3 = 14.7 G, in the case of 5-PCSL spectra; Aii = 7.1 G, 7.1 G, 29.0 G, <A> = 14.4 G, in the case of 7-PCSL spectra; Aii = 7.1 G, 7.1 G, 28.5 G, <A> = 14.2 G, in the case of 10-PCSL spectra; Aii = 7.1, 7.1, 28.0 G, <A> = 14.1 G, in the case of 14-PCSL spectra; Aii = 5.0, 5.0, 33.0 G, <A> = 14.3 G, in the case of CNO spectra;
- the correlation time for the rotational diffusion motion of the probes, τ. To improve the fitting between the experimental and the computed line shapes, it was necessary to include in the calculation an anisotropy of motion (different parallel and perpendicular τ values, namely τpar and τperp), also considering a tilt of the rotational axis. However, in the case of all the spin probes, the fitting was good by taking constant both the τpar value (13.2 ns, indicative of the steric hindrance of the chain in its parallel direction with respect to the p-orbital hosting the unpaired electron) and the tilt angle (70°). Therefore, the main parameter τ changing from one to another system is τperp;
- the order parameter, indicated as S, which measures the orientational ordering of the labelled segment of the acyl chain with respect to the normal to the bilayer surface. This parameter changes from one system to another.
4.5. Small Angle Neutron Scattering (SANS)
4.6. Analysis of SANS Data
4.7. Neutron Reflectivity (NR)
4.8. Analysis of NR Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kučerka, N.; Gallová, J.; Uhríková, D. The membrane structure and function affected by water. Chem. Phys. Lipids 2019, 221, 140–144. [Google Scholar] [CrossRef]
- Frias, M.A.; Disalvo, E.A. Breakdown of classical paradigms in relation to membrane structure and functions. Biochim. Biophys. Acta (BBA)-Biomembr. 2020, 1863, 183512. [Google Scholar] [CrossRef]
- Rosa, A.S.; Disalvo, E.A.; Frias, M.A. Water Behavior at the Phase Transition of Phospholipid Matrixes Assessed by FTIR Spec-troscopy. J. Phys. Chem. B 2020, 124, 6236–6244. [Google Scholar] [CrossRef]
- Semrau, S.; Schimdt, T. Membrane heterogeneity—From lipid domains to curvature effects. RSC Adv. 2009, 5, 3174–3186. [Google Scholar] [CrossRef]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggeling, C. The mystery of membrane organization: Composition, regulation and roles of lipid rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef]
- Sezgin, E.; Gutmann, T.; Buhl, T.; Dirkx, R.; Grzybek, M.; Coskun, Ü.; Solimena, M.; Simons, K.; Levental, I.; Schwille, P. Adaptive Lipid Packing and Bioactivity in Membrane Domains. PLoS ONE 2015, 10, e0123930. [Google Scholar] [CrossRef]
- Harayama, T.; Riezman, H. Understanding the diversity of membrane lipid composition. Nat. Rev. Mol. Cell Biol. 2018, 19, 281–296. [Google Scholar] [CrossRef]
- Phillips, R.; Ursell, T.; Wiggins, P.; Sens, P. Emerging roles for lipids in shaping membrane-protein function. Nature 2009, 459, 379–385. [Google Scholar] [CrossRef]
- Dyall, S.C. Long-chain omega-3 fatty acids and the brain: A review of the independent and shared effects of EPA, DPA and DHA. Front. Aging Neurosci. 2015, 7, 52. [Google Scholar] [CrossRef]
- Reyes-Corral, M.; Sola-Idigora, N.; de la Puerta, R.; Montaner, J.; Ybot-Gonzalez, P. Nutraceuticals in the Prevention of Neonatal Hypoxia-Ischemia: A Comprehensive Review of their Neuroprotective Properties, Mechanisms of Action and Future Direc-tions. Int. J. Mol. Sci. 2021, 22, 2524. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: Nutrition or pharmacology? Br. J. Clin. Pharmacol. 2013, 75, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Suzumura, A.; Terao, R.; Kaneko, H. Protective Effects and Molecular Signaling of n-3 Fatty Acids on Oxidative Stress and Inflammation in Retinal Diseases. Antioxid.-Basel 2020, 9, 920. [Google Scholar] [CrossRef] [PubMed]
- Zgorzynska, E.; Dziedzic, B.; Markiewicz, M.; Walczewska, A. Omega-3 PUFAs Suppress IL-1β-Induced Hyperactivity of Immunoproteasomes in Astrocytes. Int. J. Mol. Sci. 2021, 22, 5410. [Google Scholar] [CrossRef]
- Gray, B.; Steyn, F.; Davies, P.S.W.; Vitetta, L. Omega-3 fatty acids: A review of the effects on adiponectin and leptin and potential implications for obesity management. Eur. J. Clin. Nutr. 2013, 67, 1234–1242. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.-S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Fuentes, N.R.; Kim, E.; Fan, Y.-Y.; Chapkin, R.S. Omega-3 fatty acids, membrane remodeling and cancer prevention. Mol. Asp. Med. 2018, 64, 79–91. [Google Scholar] [CrossRef]
- Liput, K.; Lepczyński, A.; Ogłuszka, M.; Nawrocka, A.; Poławska, E.; Grzesiak, A.; Ślaska, B.; Pareek, C.; Czarnik, U.; Pierzchała, M. Effects of Dietary n–3 and n–6 Polyunsaturated Fatty Acids in Inflammation and Cancerogenesis. Int. J. Mol. Sci. 2021, 22, 6965. [Google Scholar] [CrossRef]
- Hishikawa, D.; Valentine, W.J.; Iizuka-Hishikawa, Y.; Shindou, H.; Shimizu, T. Metabolism and functions of docosahexaenoic acid-containing membrane glycerophospholipids. FEBS Lett. 2017, 591, 2730–2744. [Google Scholar] [CrossRef]
- Pinot, M.; Vanni, S.; Pagnotta, S.; Lacas-Gervais, S.; Payet, L.-A.; Ferreira, T.; Gautier, R.; Goud, B.; Antonny, B.; Barelli, H. Lipid cell biology. Polyunsaturated phospholipids facilitate membrane deformation and fission by endocytic proteins. Science 2014, 345, 693–697. [Google Scholar] [CrossRef]
- Leng, X.; Kinnun, J.J.; Shaikh, S.R.; Feller, S.E.; Wassall, S.R. All N-3 PUFA are not the Same: A Comparison of DPA, EPA and DHA by MD Simulations. Biophys. J. 2016, 110, 583a. [Google Scholar] [CrossRef]
- Feller, S.E.; Gawrisch, K.; MacKerell, A.D. Polyunsaturated Fatty Acids in Lipid Bilayers: Intrinsic and Environmental Contributions to Their Unique Physical Properties. J. Am. Chem. Soc. 2001, 124, 318–326. [Google Scholar] [CrossRef] [PubMed]
- De Santis, A.; Vitiello, G.; Appavou, M.-S.; Scoppola, E.; Fragneto, G.; Barnsley, L.C.; Clifton, L.A.; Ottaviani, M.F.; Paduano, L.; Krauss, I.R.; et al. Not just a fluidifying effect: Omega-3 phospholipids induce formation of non-lamellar structures in biomembranes. Soft Matter 2020, 16, 10425–10438. [Google Scholar] [CrossRef] [PubMed]
- Weiner, M.D.; Feigenson, G.W. Presence and Role of Midplane Cholesterol in Lipid Bilayers Containing Registered or Antiregistered Phase Domains. J. Phys. Chem. B 2018, 122, 8193–8200. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, D.; Heberle, F.A.; Nickels, J.D.; Pabst, G.; Katsaras, J. On scattered waves and lipid domains: Detecting membrane rafts with X-rays and neutrons. Soft Matter 2015, 11, 9055–9072. [Google Scholar] [CrossRef] [PubMed]
- Maxfield, F.R.; van Meer, G. Cholesterol, the central lipid of mammalian cells. Curr. Opin. Cell Biol. 2010, 22, 422–429. [Google Scholar] [CrossRef]
- Cornelius, F. Modulation of Na,K-ATPase and Na-ATPase Activity by Phospholipids and Cholesterol. I. Steady-State Kinetics. Biochemistry 2001, 40, 8842–8851. [Google Scholar] [CrossRef]
- De Meyer, F.J.-M.; Rodgers, J.M.; Willems, T.F.; Smit, B. Molecular Simulation of the Effect of Cholesterol on Lipid-Mediated Protein-Protein Interactions. Biophys. J. 2010, 99, 3629–3638. [Google Scholar] [CrossRef]
- Hooper, N.M. Detergent-insoluble glycosphingolipid/cholesterol-rich membrane domains, lipid rafts and caveolae (Review). Mol. Membr. Biol. 1999, 16, 145–156. [Google Scholar] [CrossRef]
- Kusumi, A.; Fujiwara, T.; Tsunoyama, T.A.; Kasai, R.; Liu, A.; Hirosawa, K.M.; Kinoshita, M.; Matsumori, N.; Komura, N.; Ando, H.; et al. Defining raft domains in the plasma membrane. Traffic 2019, 21, 106–137. [Google Scholar] [CrossRef]
- Silvius, J.R. Role of cholesterol in lipid raft formation: Lessons from lipid model systems. Biochim. et Biophys. Acta (BBA)-Biomembr. 2003, 1610, 174–183. [Google Scholar] [CrossRef]
- Helms, J.B.; Zurzolo, C. Lipids as Targeting Signals: Lipid Rafts and Intracellular Trafficking. Traffic 2004, 5, 247–254. [Google Scholar] [CrossRef]
- Laude, A.; Prior, I.A. Plasma membrane microdomains: Organization, function and trafficking (Review). Mol. Membr. Biol. 2004, 21, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Alwarawrah, M.; Dai, J.; Huang, J. A Molecular View of the Cholesterol Condensing Effect in DOPC Lipid Bilayers. J. Phys. Chem. B 2010, 114, 7516–7523. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Alwarawrah, M.; Huang, J. Instability of Cholesterol Clusters in Lipid Bilayers and The Cholesterol’s Umbrella Effect. J. Phys. Chem. B 2009, 114, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Hung, W.-C.; Lee, M.-T.; Chen, F.-Y.; Huang, H.W. The Condensing Effect of Cholesterol in Lipid Bilayers. Biophys. J. 2007, 92, 3960–3967. [Google Scholar] [CrossRef]
- Leeb, F.; Maibaum, L. Spatially Resolving the Condensing Effect of Cholesterol in Lipid Bilayers. Biophys. J. 2018, 115, 2179–2188. [Google Scholar] [CrossRef]
- Róg, T.; Pasenkiewicz-Gierula, M.; Vattulainen, I.; Karttunen, M. Ordering effects of cholesterol and its analogues. Biochim. Biophys. Acta (BBA)-Biomembr. 2009, 1788, 97–121. [Google Scholar] [CrossRef]
- Chakraborty, S.; Doktorova, M.; Molugu, T.R.; Heberle, F.A.; Scott, H.L.; Dzikovski, B.; Nagao, M.; Stingaciu, L.-R.; Standaert, R.F.; Barrera, F.N.; et al. How cholesterol stiffens unsaturated lipid membranes. Proc. Natl. Acad. Sci. USA 2020, 117, 21896–21905. [Google Scholar] [CrossRef]
- Stillwell, W.; Shaikh, S.R.; Zerouga, M.; Siddiqui, R.; Wassall, S.R. Docosahexaenoic acid affects cell signaling by altering lipid rafts. Reprod. Nutr. Dev. 2005, 45, 559–579. [Google Scholar] [CrossRef]
- Sherratt, S.C.R.; Mason, R.P. Eicosapentaenoic acid and docosahexaenoic acid have distinct membrane locations and lipid in-teractions as determined by X-ray diffraction. Chem. Phys. Lipids 2018, 212, 73–79. [Google Scholar] [CrossRef]
- Wassall, S.R.; Leng, X.; Canner, S.W.; Pennington, E.R.; Kinnun, J.; Cavazos, A.T.; Dadoo, S.; Johnson, D.; Heberle, F.; Katsaras, J.; et al. Docosahexaenoic acid regulates the formation of lipid rafts: A unified view from experiment and simulation. Biochim. Biophys. Acta (BBA)-Biomembr. 2018, 1860, 1985–1993. [Google Scholar] [CrossRef] [PubMed]
- Mihailescu, M.; Soubias, O.; Worcester, D.; White, S.H.; Gawrisch, K. Structure and Dynamics of Cholesterol-Containing Polyunsaturated Lipid Membranes Studied by Neutron Diffraction and NMR. J. Membr. Biol. 2010, 239, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Kučerka, N.; Marquardt, D.; Harroun, T.A.; Nieh, M.-P.; Wassall, S.R.; Katsaras, J. The Functional Significance of Lipid Diversity: Orientation of Cholesterol in Bilayers Is Determined by Lipid Species. J. Am. Chem. Soc. 2009, 131, 16358–16359. [Google Scholar] [CrossRef] [PubMed]
- Marrink, S.J.; de Vries, A.H.; Harroun, T.A.; Katsaras, J.; Wassall, S.R. Cholesterol Shows Preference for the Interior of Polyunsaturated Lipid Membranes. J. Am. Chem. Soc. 2007, 130, 10–11. [Google Scholar] [CrossRef]
- Shaikh, S.R.; Cherezov, V.; Caffrey, M.; Soni, S.P.; LoCascio, D.; Stillwell, W.; Wassall, S.R. Molecular Organization of Cholesterol in Unsaturated Phosphatidylethanolamines: X-ray Diffraction and Solid State 2H NMR Reveal Differences with Phosphatidylcholines. J. Am. Chem. Soc. 2006, 128, 5375–5383. [Google Scholar] [CrossRef]
- Marquardt, D.; Kucerka, N.; Wassall, S.R.; Harroun, T.; Katsaras, J. Cholesterol’s location in lipid bilayers. Chem. Phys. Lipids 2016, 199, 17–25. [Google Scholar] [CrossRef]
- Marquardt, D.; Heberle, F.A.; Greathouse, D.V.; Koeppe, R.E.; Standaert, R.F.; Van Oosten, B.J.; Harroun, T.A.; Kinnun, J.J.; Williams, J.A.; Wassall, S.R.; et al. Lipid bilayer thickness determines cholesterol’s location in model membranes. Soft Matter 2016, 12, 9417–9428. [Google Scholar] [CrossRef]
- Ollila, O.S.; Pabst, G. Atomistic resolution structure and dynamics of lipid bilayers in simulations and experiments. Biochim. Biophys. Acta (BBA)-Biomembr. 2016, 1858, 2512–2528. [Google Scholar] [CrossRef]
- Perlmutter, J.D.; Sachs, J.N. Interleaflet Interaction and Asymmetry in Phase Separated Lipid Bilayers: Molecular Dynamics Simulations. J. Am. Chem. Soc. 2011, 133, 6563–6577. [Google Scholar] [CrossRef]
- Aghaaminiha, M.; Farnoud, A.M.; Sharma, S. Quantitative relationship between cholesterol distribution and ordering of lipids in asymmetric lipid bilayers. Soft Matter 2021, 17, 2742–2752. [Google Scholar] [CrossRef]
- Duraisamy, Y.; Lambert, D.; O’Neill, C.A.; Padfield, P.J. Differential incorporation of docosahexaenoic acid into distinct cholesterol-rich membrane raft domains. Biochem. Biophys. Res. Commun. 2007, 360, 885–890. [Google Scholar] [CrossRef]
- Martín, V.; Fabelo, N.; Santpere, G.; Puig, B.; Marín, R.; Ferrer, I.; Díaz, M. Lipid Alterations in Lipid Rafts from Alzheimer’s Disease Human Brain Cortex. J. Alzheimer’s Dis. 2010, 19, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Konyakhina, T.M.; Feigenson, G.W. Phase diagram of a polyunsaturated lipid mixture: Brain sphingomyelin/1-stearoyl-2-docosahexaenoyl-sn-glycero-3-phosphocholine/cholesterol. Biochim. Biophys. Acta (BBA)-Biomembr. 2015, 1858, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, S.R.; Dumaual, A.C.; Castillo, A.; LoCascio, D.; Siddiqui, R.A.; Stillwell, W.; Wassall, S.R. Oleic and Docosahexaenoic Acid Differentially Phase Separate from Lipid Raft Molecules: A Comparative NMR, DSC, AFM, and Detergent Extraction Study. Biophys. J. 2004, 87, 1752–1766. [Google Scholar] [CrossRef] [PubMed]
- Wassall, S.R.; Stillwell, W. Polyunsaturated fatty acid–cholesterol interactions: Domain formation in membranes. Biochim. Biophys. Acta (BBA)-Biomembr. 2009, 1788, 24–32. [Google Scholar] [CrossRef]
- Rappolt, M.; Vidal, M.; Kriechbaum, M.; Steinhart, M.; Amenitsch, H.; Bernstorff, S.; Laggner, P. Structural, dynamic and mechanical properties of POPC at low cholesterol concentration studied in pressure/temperature space. Eur. Biophys. J. 2003, 31, 575–585. [Google Scholar] [CrossRef]
- Scheidt, H.A.; Müller, P.; Herrmann, A.; Huster, D. The Potential of Fluorescent and Spin-labeled Steroid Analogs to Mimic Natural Cholesterol. J. Biol. Chem. 2003, 278, 45563–45569. [Google Scholar] [CrossRef]
- Perlstein, M.; Ottaviani, M.F.; Aserin, A.; Garti, N. Structural effects on cosolubilization of dendrimer and propofol in water dilutable microemulsions as delivery vehicle. Colloids Surfaces A: Physicochem. Eng. Asp. 2016, 497, 257–264. [Google Scholar] [CrossRef]
- Prigat, Y.; Fattori, A.; Shames, A.I.; Ottaviani, M.F.; Garti, N. Micro-characterization of modified microemulsions loaded with gossypol, pure and extracted from cottonseed. Colloids Surfaces B Biointerfaces 2019, 180, 487–494. [Google Scholar] [CrossRef]
- Levy, D.; Briggman, K.A. Cholesterol/Phospholipid Interactions in Hybrid Bilayer Membranes. Langmuir 2007, 23, 7155–7161. [Google Scholar] [CrossRef]
- M’Baye, G.; Mély, Y.; Duportail, G.; Klymchenko, A.S. Liquid Ordered and Gel Phases of Lipid Bilayers: Fluorescent Probes Reveal Close Fluidity but Different Hydration. Biophys. J. 2008, 95, 1217–1225. [Google Scholar] [CrossRef] [PubMed]
- Doucet, M.; Cho, J.H.; Alina, G.; Bakker, J.; Bouwman, W.; Butler, P.; Campbell, K.; Gonzales, M.; Heenan, R.; Jackson, A.; et al. SasView, version 4.1; Zenodo: Meyrin, Switzerland, 2017. [Google Scholar]
- Kotlarchyk, M.; Ritzau, S.M. Paracrystal model of the high-temperature lamellar phase of a ternary microemulsion system. J. Appl. Crystallogr. 1991, 24, 753–758. [Google Scholar] [CrossRef]
- Lolicato, F.; Joly, L.; Martinez-Seara, H.; Fragneto, G.; Scoppola, E.; Bombelli, F.B.; Vattulainen, I.; Akola, J.; Maccarini, M. The Role of Temperature and Lipid Charge on Intake/Uptake of Cationic Gold Nanoparticles into Lipid Bilayers. Small 2019, 15, 1805046. [Google Scholar] [CrossRef] [PubMed]
- Scoppola, E.; Micciulla, S.; Kuhrts, L.; Maestro, A.; Campbell, R.A.; Konovalov, O.V.; Fragneto, G.; Schneck, E. Reflectometry reveals accumulation of surfactants impurities at bare oil/water interfaces. Molecules 2019, 24, 4113. [Google Scholar] [CrossRef] [PubMed]
- Merlino, A.; Vitiello, G.; Grimaldi, M.; Sica, F.; Busi, E.; Basosi, R.; D’Ursi, A.M.; Fragneto, G.; Paduano, L.; D’Errico, G. Destabilization of Lipid Membranes by a Peptide Derived from Glycoprotein gp36 of Feline Immunodeficiency Virus: A Combined Molecular Dynamics/Experimental Study. J. Phys. Chem. B 2012, 116, 401–412. [Google Scholar] [CrossRef]
- Vega, M.; Lurio, L.B.; Lal, J.; Karapetrova, E.A.; Gaillard, E.R. Structure of supported DPPC/cholesterol bilayers studied via X-ray reflectivity. Phys. Chem. Chem. Phys. 2020, 22, 19089–19099. [Google Scholar] [CrossRef]
- Levental, I.; Levental, K.R.; Heberle, F.A. Lipid Rafts: Controversies Resolved, Mysteries Remain. Trends Cell Biol. 2020, 30, 341–353. [Google Scholar] [CrossRef]
- Gawrisch, K.; Eldho, N.V.; Holte, L.L. The structure of DHA in phospholipid membranes. Lipids 2003, 38, 445–452. [Google Scholar] [CrossRef]
- Saiz, L.; Klein, M.L. Structural Properties of a Highly Polyunsaturated Lipid Bilayer from Molecular Dynamics Simulations. Biophys. J. 2001, 81, 204–216. [Google Scholar] [CrossRef][Green Version]
- Miyashita, K. Paradox of omega-3 PUFA oxidation. Eur. J. Lipid Sci. Technol. 2014, 116, 268–1279. [Google Scholar] [CrossRef]
- Brzustowicz, M.R.; Cherezov, V.; Zerouga, M.; Caffrey, M.; Stillwell, W.; Wassall, S.R. Controlling Membrane Cholesterol Content. A Role for Polyunsaturated (Docosahexaenoate) Phospholipids. Biochemistry 2002, 41, 12509–12519. [Google Scholar] [CrossRef] [PubMed]
- Brzustowicz, M.R.; Cherezov, V.; Caffrey, M.; Stillwell, W.; Wassall, S.R. Molecular Organization of Cholesterol in Polyunsaturated Membranes: Microdomain Formation. Biophys. J. 2002, 82, 285–298. [Google Scholar] [CrossRef]
- Ibarguren, M.; Alonso, A.; Tenchov, B.G.; Goñi, F.M. Quantitation of cholesterol incorporation into extruded lipid bilayers. Biochim. Biophys. Acta (BBA)-Biomembr. 2010, 1798, 1735–1738. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Buboltz, J.T.; Feigenson, G.W. Maximum solubility of cholesterol in phosphatidylcholine and phosphatidylethanolamine bilayers. Biochim. Biophys. Acta (BBA)-Biomembr. 1999, 1417, 89–100. [Google Scholar] [CrossRef]
- Freeman, R.; Finean, J.B. Cholesterol: Lecithin association at molecular ratios of up to 2:1. Chem. Phys. Lipids 1975, 14, 313–320. [Google Scholar] [CrossRef]
- Gershfeld, N.L. Equilibrium Studies of Lecithin-Cholesterol Interactions: I. Stoichiometry of Lecithin-Cholesterol Complexes in Bulk Systems. Biophys. J. 1978, 22, 469–488. [Google Scholar] [CrossRef]
- Collins, J.J.; Phillips, M.C. The stability and structure of cholesterol-rich codispersions of cholesterol and phosphatidylcholine. J. Lipid Res. 1982, 23, 291–298. [Google Scholar] [CrossRef]
- Cimato, A.; Facorro, G.; Sarrasague, M.M. Determining the fluid ordered and disordered phases in a pulmonary surfactant by electron spin resonance technique. Respir. Physiol. Neurobiol. 2019, 271, 103309. [Google Scholar] [CrossRef]
- Collado, M.I.; Goñi, F.M.; Alonso, A.A.; Marsh, D. Domain Formation in Sphingomyelin/Cholesterol Mixed Membranes Studied by Spin-Label Electron Spin Resonance Spectroscopy. Biochemistry 2005, 44, 4911–4918. [Google Scholar] [CrossRef]
- Ionova, I.V.; Livshits, V.A.; Marsh, D. Phase Diagram of Ternary Cholesterol/Palmitoylsphingomyelin/Palmitoyloleoyl-Phosphatidylcholine Mixtures: Spin-Label EPR Study of Lipid-Raft Formation. Biophys. J. 2012, 102, 1856–1865. [Google Scholar] [CrossRef]
- Stillwell, W.; Wassall, S.R. Docosahexaenoic acid: Membrane properties of a unique fatty acid. Chem. Phys. Lipids 2003, 126, 1–27. [Google Scholar] [CrossRef]
- Shaikh, S.R.; Cherezov, V.; Caffrey, M.; Stillwell, W.; Wassall, S.R. Interaction of Cholesterol with a Docosahexaenoic Acid-Containing Phosphatidylethanolamine: Trigger for Microdomain/Raft Formation? Biochemistry 2003, 42, 12028–12037. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, S.R.; LoCascio, D.S.; Soni, S.P.; Wassall, S.R.; Stillwell, W. Oleic- and docosahexaenoic acid-containing phosphatidylethanolamines differentially phase separate from sphingomyelin. Biochim. Biophys. Acta (BBA)-Biomembr. 2009, 1788, 2421–2426. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, S.R.; Rockett, B.D.; Salameh, M.; Carraway, K. Docosahexaenoic Acid Modifies the Clustering and Size of Lipid Rafts and the Lateral Organization and Surface Expression of MHC Class I of EL4 Cells. J. Nutr. 2009, 139, 1632–1639. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.A.; Batten, S.E.; Harris, M.; Rockett, B.D.; Shaikh, S.R.; Stillwell, W.; Wassall, S.R. Docosahexaenoic and Eicosapentaenoic Acids Segregate Differently between Raft and Nonraft Domains. Biophys. J. 2012, 103, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Goh, M.W.S.; Tero, R. Cholesterol-induced microdomain formation in lipid bilayer membranes consisting of completely miscible lipids. Biochim. Biophys. Acta (BBA)-Biomembr. 2021, 1863, 183626. [Google Scholar] [CrossRef]
- Pasenkiewicz-Gierula, M.; Baczynski, K.; Markiewicz, M.; Murzyn, K. Computer modelling studies of the bilayer/water interface. Biochim. Biophys. Acta Biomembr. 2016, 1858, 2305–2321. [Google Scholar] [CrossRef]
- Huang, J.; Feigenson, G.W. A Microscopic Interaction Model of Maximum Solubility of Cholesterol in Lipid Bilayers. Biophys. J. 1999, 76, 2142–2157. [Google Scholar] [CrossRef]
- Huster, D.; Arnold, K.; Gawrisch, K. Influence of Docosahexaenoic Acid and Cholesterol on Lateral Lipid Organization in Phospholipid Mixtures. Biochemistry 1998, 37, 17299–17308. [Google Scholar] [CrossRef]
- Shaikh, S.R.; Kinnun, J.J.; Leng, X.; Williams, J.A.; Wassall, S.R. How polyunsaturated fatty acids modify molecular organization in membranes: Insight from NMR studies of model systems. Biochim. Biophys. Acta Biomembr. 2015, 1848, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Bagatolli, L.; Maggio, B.; Aguilar, F.; Sotomayor, C.P.; Fidelio, G.D. Laurdan properties in glycosphingolipid-phospholipid mixtures: A comparative fluorescence and calorimetric study. Biochim. Biophys. Acta (BBA)-Biomembr. 1997, 1325, 80–90. [Google Scholar] [CrossRef]
- Fidorra, M.; Duelund, L.; Leidy, C.; Simonsen, A.C.; Bagatolli, L. Absence of Fluid-Ordered/Fluid-Disordered Phase Coexistence in Ceramide/POPC Mixtures Containing Cholesterol. Biophys. J. 2006, 90, 4437–4451. [Google Scholar] [CrossRef] [PubMed]
- Leekumjorn, S.; Sum, A.K. Molecular Characterization of Gel and Liquid-Crystalline Structures of Fully Hydrated POPC and POPE Bilayers. J. Phys. Chem. B 2007, 111, 6026–6033. [Google Scholar] [CrossRef]
- Budil, D.E.; Lee, S.; Saxena, S.; Freed, J.H. Nonlinear-Least-Squares Analysis of Slow-Motion EPR Spectra in One and Two Dimensions Using a Modified Levenberg-Marquardt Algorithm. J. Magn. Reson. Ser. A 1996, 120, 155–189. [Google Scholar] [CrossRef]
- Feoktystov, A.V.; Frielinghaus, H.; Di, Z.; Jaksch, S.; Pipich, V.; Appavou, M.-S.; Babcock, E.; Hanslik, R.; Engels, R.; Kemmerling, G.; et al. KWS-1 high-resolution small-angle neutron scattering instrument at JCNS: Current state. J. Appl. Crystallogr. 2015, 48, 61–70. [Google Scholar] [CrossRef]
- Frielinghaus, H.; Feoktystov, A.V.; Berts, I.; Mangiapia, G. KWS-1: Small-angle scattering diffractometer. J. Large-Scale Res. Facil. 2015, 1, A28. [Google Scholar] [CrossRef]
- Mattauch, S.; Koutsioumpas, A.; Rücker, U.; Korolkov, D.; Fracassi, V.; Daemen, J.; Schmitz, R.; Bussmann, K.; Suxdorf, F.; Wagener, M.; et al. The high-intensity reflectometer of the Jülich Centre for Neutron Science: MARIA. J. Appl. Crystallogr. 2018, 51, 646–654. [Google Scholar] [CrossRef]
- Fragneto, G.; Thomas, R.K.; Rennie, A.R.; Penfold, J. Neutron Reflection from Hexadecyltrimethylammonium Bromide Adsorbed on Smooth and Rough Silicon Surfaces. Langmuir 1996, 12, 6036–6043. [Google Scholar] [CrossRef]
- Scoppola, E.; Gochev, G.G.; Drnec, J.; Pithan, L.; Novikov, D.; Schneck, E. Investigating the Conformation of Surface-Adsorbed Proteins with Standing-Wave X-ray Fluorescence. Biomacromolecules 2021, 22, 5195–5203. [Google Scholar] [CrossRef]
- De la Serna, J.B.; Schütz, G.J.; Eggeling, C.; Cebecauer, M. There Is No Simple Model of the Plasma Membrane Organization. Front. Cell Dev. Biol. 2016, 4, 106. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molar Fraction of Polyunsaturated Lipids with Respect to Total Phosphocholines | Molar Fraction with Respect to Total Lipids | Molar Fraction of DHA with Respect to Total Phosphocholine Tails | ||||
|---|---|---|---|---|---|---|
| Cholesterol | POPC | 22:6–22:6PC | 18:0–22:6PC | |||
| chol/POPC | x22:6–22:6PC = 0x18:0–22:6PC = 0 | 0.4 | 0.6 | 0 | 0 | xDHA = 0 |
| chol/POPC/ 22:6–22:6PC | x22:6–22:6PC = 0.2 | 0.4 | 0.48 | 0.12 | 0 | xDHA = 0.2 |
| x22:6–22:6PC = 0.4 | 0.4 | 0.36 | 0.24 | 0 | xDHA = 0.4 | |
| x22:6–22:6PC = 0.5 | 0.4 | 0.3 | 0.3 | 0 | xDHA = 0.5 | |
| x22:6–22:6PC = 0.6 | 0.4 | 0.24 | 0.36 | 0 | xDHA = 0.6 | |
| x22:6–22:6PC = 0.8 | 0.4 | 0.12 | 0.48 | 0 | xDHA = 0.8 | |
| chol/22:6–22:6PC | x22:6–22:6PC = 1 | 0.4 | 0 | 0.6 | 0 | xDHA = 1 |
| chol/POPC/ 18:0–22:6PC | x18:0–22:6PC = 0.2 | 0.4 | 0.48 | 0 | 0.12 | xDHA = 0.1 |
| x18:0–22:6PC = 0.4 | 0.4 | 0.36 | 0 | 0.24 | xDHA = 0.2 | |
| x18:0–22:6PC = 0.5 | 0.4 | 0.3 | 0 | 0.3 | xDHA = 0.25 | |
| x18:0–22:6PC = 0.6 | 0.4 | 0.24 | 0 | 0.36 | xDHA = 0.3 | |
| x18:0–22:6PC = 0.8 | 0.4 | 0.12 | 0 | 0.48 | xDHA = 0.4 | |
| chol/18:0–22:6PC | x18:0–22:6PC = 1 | 0.4 | 0 | 0 | 0.6 | xDHA = 0.5 |
| Thickness (Å) | N Layers | <D> (Å) | σD/<D> | Polydispersity on Thickness | |
|---|---|---|---|---|---|
| chol/POPC | 38.09 ± 0.05 | 4.424 ± 0.005 | 105.14 ± 0.06 | 1.311 ± 0.002 | 0.05 |
| chol/POPC/22:6–22:6PC x22:6–22:6PC = 0.2 | 38.67 ± 0.06 | 3.801 ± 0.003 | 94.73 ± 0.06 | 1.801 ± 0.002 | 0.1 |
| chol/POPC/22:6–22:6PC x22:6–22:6PC = 0.8 | 34.60 ± 0.09 | 2.698 ± 0.006 | 110.1 ± 0.2 | 1.877 ± 0.008 | 0.05 |
| Headrgroup Thickness (Å) | Tail Thickness (Å) | Bilayer Roughness (Å) | Area/Lipid (Å2) | |
|---|---|---|---|---|
| chol/POPC | 7.2 (6.8–8.1) | 35.8 (33.6–37.8) | 2.9 (2.4–5.6) | 45 ± 1 |
| chol/POPC/22:6–22:6PC x22:6–22:6PC = 0.2 | 8.5 (7.7–9.3) | 27.2 (25.9–30.5) | 0.7 (0.6–3.0) | 84 ± 3 |
| chol/POPC/22:6–22:6PC x22:6–22:6PC = 0.8 | 13.0 (13.0–13.0) | 23.3 (22.7–25.5) | 8.7 (7.2–9.2) | 60 ± 4 |
| chol/POPC/18:0–22:6PC x22:6–22:6PC = 0.4 | 12.4 (11.3–12.5) | 30.6 (29.1–32.4) | 3.4 (2.8–4.5) | 77 ± 3 |
| 18:0–22:6PC/chol | 7.8 (7.1–8.7) | 29.1 (28.5–29.8) | 3.0 (2.5–3.6) | 61 ± 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Santis, A.; Scoppola, E.; Ottaviani, M.F.; Koutsioubas, A.; Barnsley, L.C.; Paduano, L.; D’Errico, G.; Russo Krauss, I. Order vs. Disorder: Cholesterol and Omega-3 Phospholipids Determine Biomembrane Organization. Int. J. Mol. Sci. 2022, 23, 5322. https://doi.org/10.3390/ijms23105322
de Santis A, Scoppola E, Ottaviani MF, Koutsioubas A, Barnsley LC, Paduano L, D’Errico G, Russo Krauss I. Order vs. Disorder: Cholesterol and Omega-3 Phospholipids Determine Biomembrane Organization. International Journal of Molecular Sciences. 2022; 23(10):5322. https://doi.org/10.3390/ijms23105322
Chicago/Turabian Stylede Santis, Augusta, Ernesto Scoppola, Maria Francesca Ottaviani, Alexandros Koutsioubas, Lester C. Barnsley, Luigi Paduano, Gerardino D’Errico, and Irene Russo Krauss. 2022. "Order vs. Disorder: Cholesterol and Omega-3 Phospholipids Determine Biomembrane Organization" International Journal of Molecular Sciences 23, no. 10: 5322. https://doi.org/10.3390/ijms23105322
APA Stylede Santis, A., Scoppola, E., Ottaviani, M. F., Koutsioubas, A., Barnsley, L. C., Paduano, L., D’Errico, G., & Russo Krauss, I. (2022). Order vs. Disorder: Cholesterol and Omega-3 Phospholipids Determine Biomembrane Organization. International Journal of Molecular Sciences, 23(10), 5322. https://doi.org/10.3390/ijms23105322