
Investigation of Chemical Constituents of Eranthis longistipitata (Ranunculaceae): Coumarins and Furochromones

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
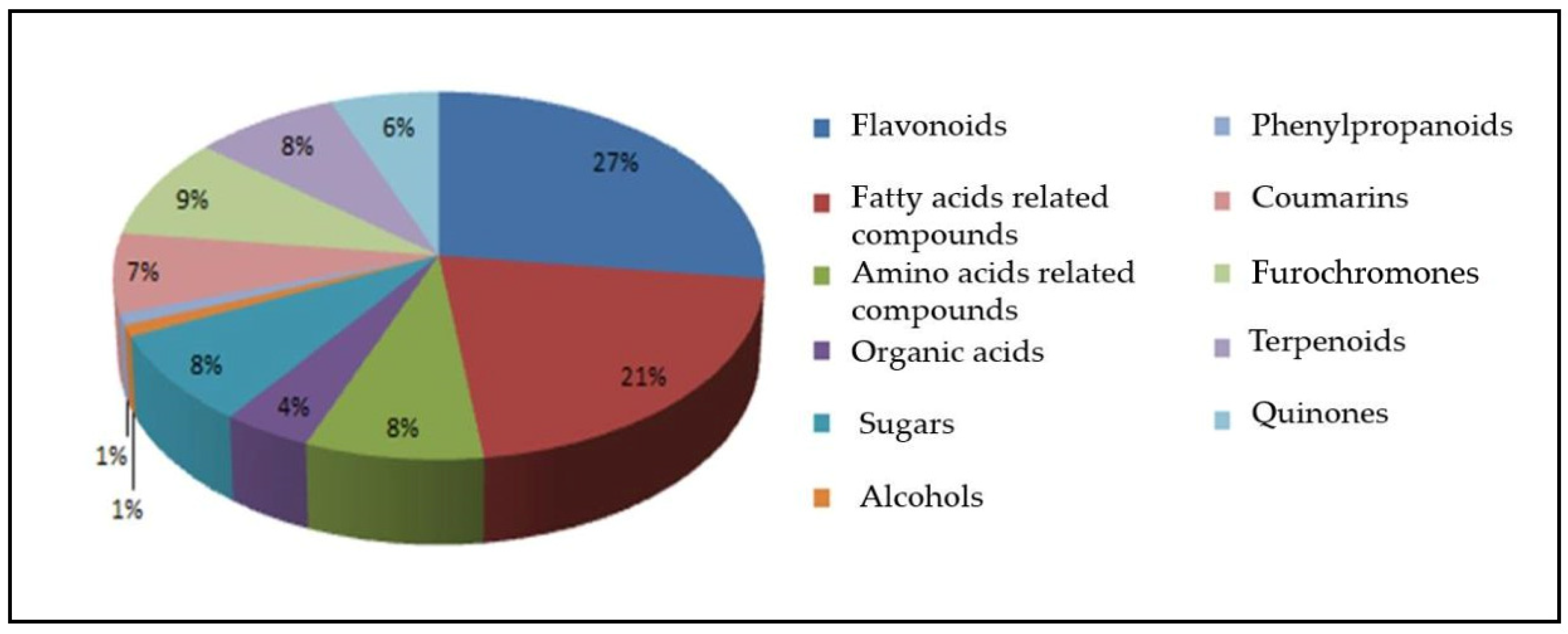
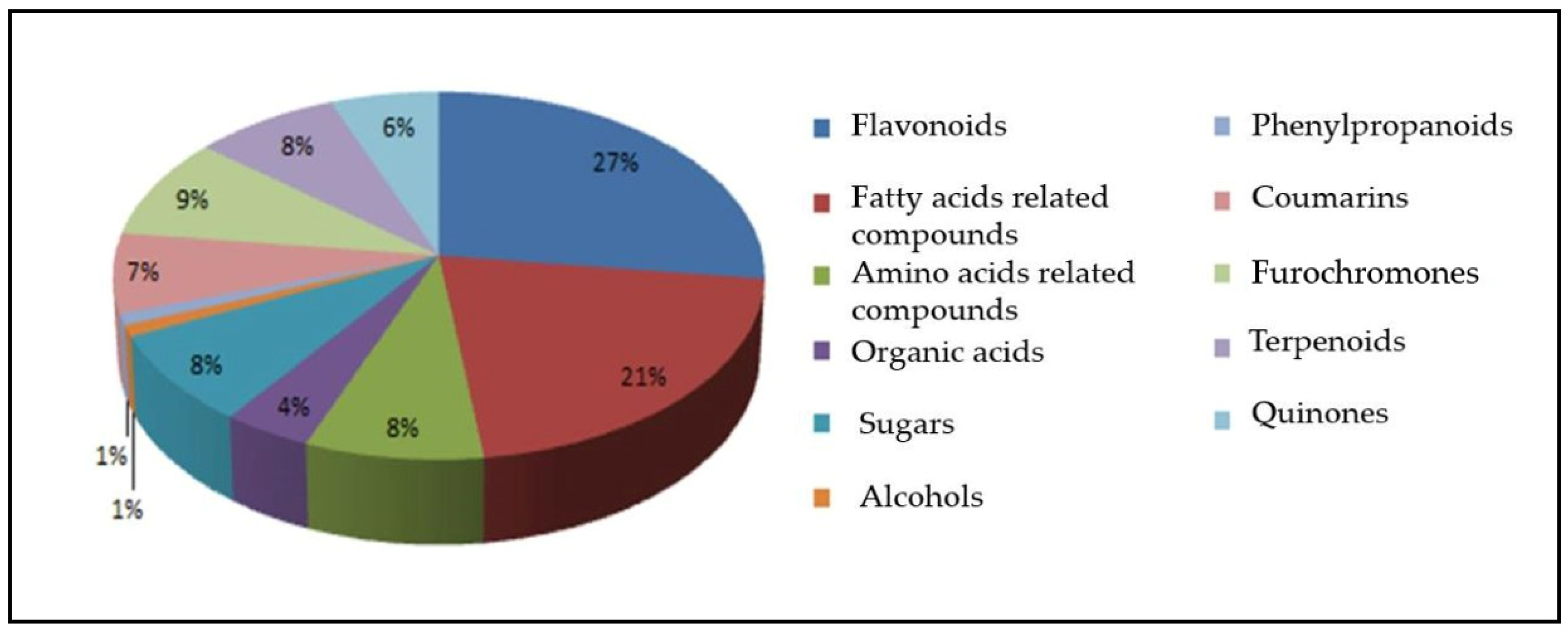
2. Results
3. Discussion
3.1. Flavonoids
3.2. Fatty-Acid–Related Compounds
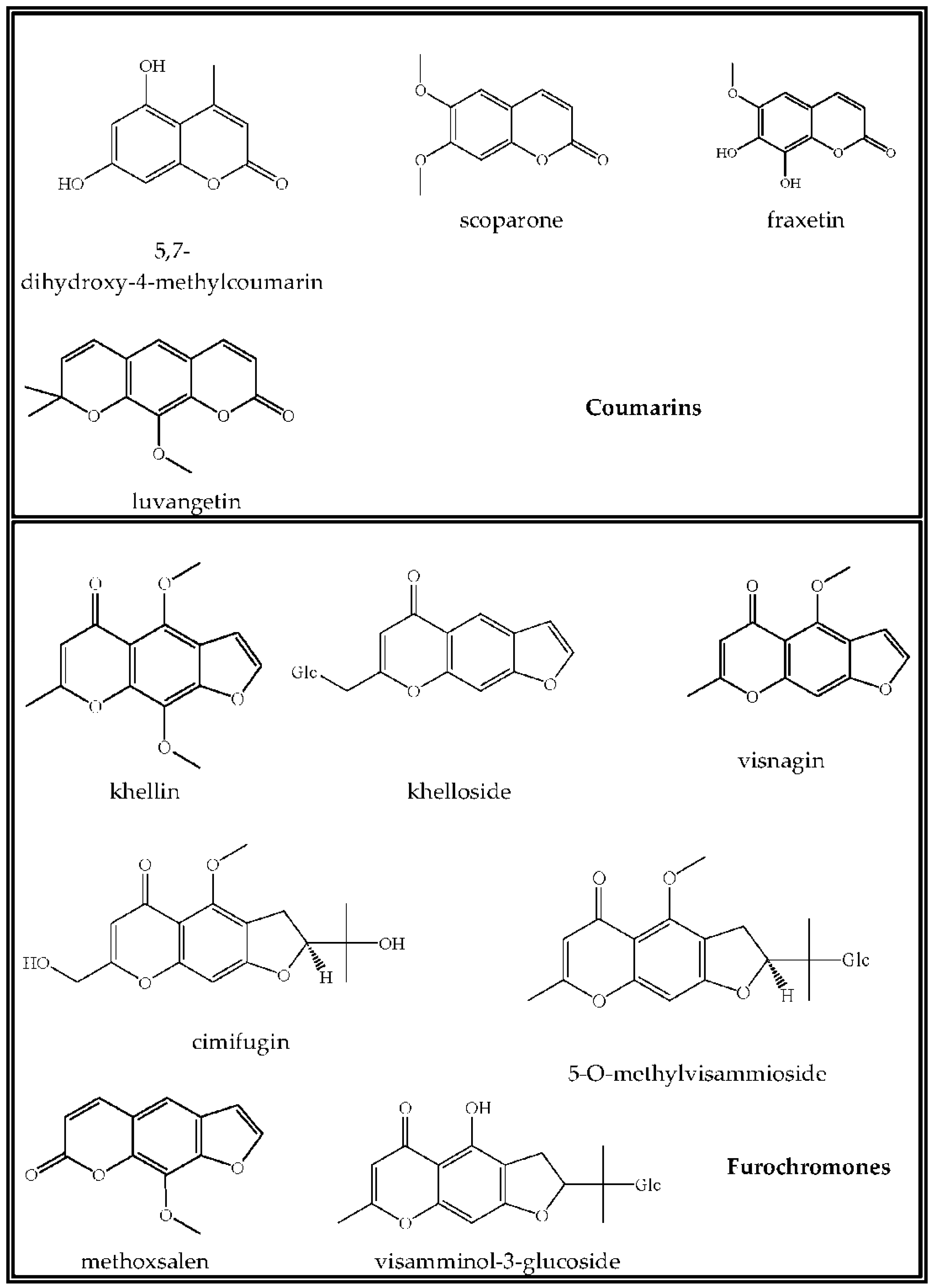
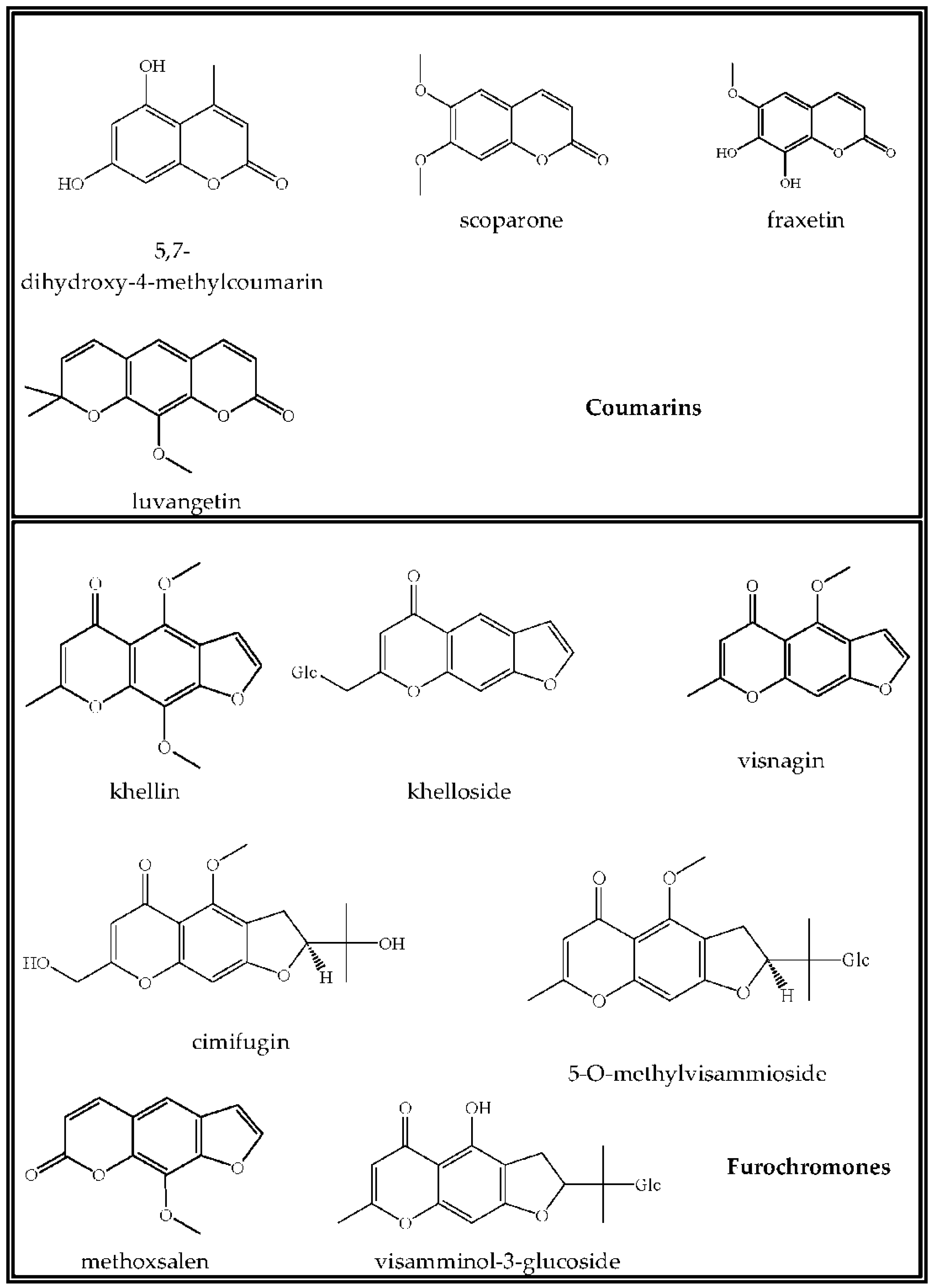
3.3. Coumarins
3.4. Chromones
4. Materials and Methods
4.1. Plant Material and Preparation of the Extract
4.2. MS Settings and the Spectral Library
4.3. Chemicals
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doppler, M.; Kluger, B.; Bueschl, C.; Schneider, C.; Krska, R.; Delcambre, S.; Hiller, K.; Lemmens, M.; Schuhmacher, R. Stable isotope-assisted evaluation of different extraction solvents for untargeted metabolomics of plants. Int. J. Mol. Sci. 2016, 17, 1017. [Google Scholar] [CrossRef] [Green Version]
- Abuzaid, H.; Amin, E.; Moawad, A.; Abdelmohsen, U.R.; Hetta, M.; Mohammed, R. Liquid Chromatography High Resolution Mass Spectrometry Analysis, Phytochemical and Biological Study of Two Aizoaceae Plants: A New Kaempferol Derivative from Trianthema portulacastrum L. Pharmacogn. Res. 2020, 12, 212. [Google Scholar] [CrossRef]
- Wang, W.; Lu, A.M.; Ren, Y.; Endress, M.E.; Chen, Z.D. Phylogeny and classification of Ranunculales: Evidence from four molecular loci and morphological data. Perspect. Plant Ecol. Evol. Syst. 2009, 11, 81–110. [Google Scholar] [CrossRef]
- Oh, A.; Oh, B.U. The speciation history of northern- and southern-sourced Eranthis (Ranunculaceae) species on the Korean peninsula and surrounding areas. Ecol. Evol. 2019, 9, 2907–2919. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Jeon, M.J.; Ma, S.H.; Wahlsteen, E.; Amundsen, K.; Kim, J.H.; Suh, J.K.; Chang, J.S.; Joung, Y.H. Phylogeny and genetic variation in the genus Eranthis using nrITS and cpIS singlenucleotide polymorphisms. Hortic. Environ. Biotechnol. 2019, 60, 239–252. [Google Scholar] [CrossRef]
- Erst, A.S.; Sukhorukov, A.P.; Mitrenina, E.Y.; Skaptsov, M.V.; Kostikova, V.A.; Chernisheva, O.A.; Troshkina, V.; Kushunina, M.; Krivenko, D.A.; Ikeda, H.; et al. An integrative taxonomic approach reveals a new species of Eranthis (Ranunculaceae) in North Asia. PhytoKeys 2020, 140, 75–100. [Google Scholar] [CrossRef]
- Mitrenina, E.Y.; Erst, A.S.; Peruzzi, L.; Skaptsov, M.V.; Ikeda, H.; Nikulin, V.Y.; Wang, W. Karyotype and genome size variation in white-flowered Eranthis sect. Shibateranthis (Ranunculaceae). PhytoKeys 2021, in press. [Google Scholar]
- Watanabe, K.; Mimaki, Y.; Sakuma, C.; Sashida, Y. Eranthisaponins A and B, two new bisdesmosidic triterpene saponins from the tubers of Eranthis cilicica. J. Nat. Prod. 2003, 66, 879–882. [Google Scholar] [CrossRef]
- Watanabe, K.; Mimaki, Y.; Fukaya, H.; Matsuo, Y. Cycloartane and oleanane glycosides from the tubers of Eranthis cilicica. Molecules 2019, 24, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostikova, V.A.; Erst, A.S.; Kuznetsov, A.A.; Gureyeva, I.I. Levels of phenolic compounds in leaves of Eranthis sibirica, E. stellata, and E. tanhoensis (Ranunculaceae). Ukr. J. Ecol. 2020, 10, 232–237. [Google Scholar] [CrossRef]
- Kostikova, V.A.; Chernonosov, A.A.; Kuznetsov, A.A.; Petrova, N.V.; Krivenko, D.A.; Chernysheva, O.A.; Wang, W.; Erst, A.S. Identification of Flavonoids in the Leaves of Eranthis longistipitata (Ranunculaceae) by Liquid Chromatography with High-Resolution Mass Spectrometry (LC-HRMS). Plants 2021, 10, 2146. [Google Scholar] [CrossRef]
- Egger, K. Khellolglucosid in Eranthis hiemalis. Z. Naturforsch. 1961, 16, 697–702. [Google Scholar] [CrossRef]
- Wada, H.; Gaino, M.; Saito, S. Furochromones of Erantis pinnatifida. Phytochemistry 1974, 13, 297–299. [Google Scholar] [CrossRef]
- Junior, P. Eranthin and eranthin-β-d-glucoside: Two new chromones from Eranthis hiemalis. Phytochemistry 1979, 18, 2053–2054. [Google Scholar] [CrossRef]
- Kopp, B.; Kubelka, E.; Reich, C.; Robien, W.; Kubelka, W. 4-H-Chromenone glycosides from Eranthis hyemalis (L.) Salisbury. Helv. Chim. Acta. 1991, 74, 611–616. [Google Scholar] [CrossRef]
- Kuroda, M.; Uchida, S.; Watanabe, K.; Mimaki, Y. Chromones from the tubers of Eranthis cilicica and their antioxidant activity. Phytochemistry 2009, 70, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Maltsev, Y.; Maltseva, K. Fatty acids of microalgae: Diversity and applications. Rev. Environ. Sc. Biotechnol. 2021, 20, 515–547. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, K.; Kulikovskiy, M.; Maltseva, S. Influence of light conditions on microalgae growth and content of lipids, carotenoids and fatty acid composition. Biology 2021, 10, 1060. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Hirai, S.; Takahashi, H.; Goto, T.; Ohyane, C.; Tsugane, T.; Konishi, C.; Fujii, T.; Inai, S.; Iijima, Y.; et al. 9-oxo-10(E),12(E)-Octadecanoic acid derived from tomato is a potent PPAR α agonist to decrease triglyceride accumulation in mouse primary hepatocytes. Mol. Nutr. Food Res. 2011, 55, 585–593. [Google Scholar] [CrossRef]
- Taki, N.; Sasaki-Sekimoto, Y.; Obayashi, T.; Kikuta, A.; Kobayashi, K.; Ainai, T.; Yagi, K.; Sakurai, N.; Suzuki, H.; Masuda, T.; et al. 12-Oxo-phytodienoic acid triggers expression of a distinct set of genes and plays a role in wond-induced gene expression in Arabidopsis. Plant. Physiol. 2005, 139, 1268–1283. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Savchenko, T.V.; Zastrijnaja, O.M.; Klimov, V.V. Oxylipins and plant abiotic stress resistance. Biochemistry 2014, 79, 362–375. [Google Scholar] [CrossRef] [PubMed]
- Taki-Nakano, N.; Kotera, J.; Ohta, H. 12-Oxo-phytodienoic acid, a plant-derived oxylipin, attenuates lipopolysaccharide-induced inflammation in microglia. Biochem. Biophys. Res. Commun. 2016, 473, 1288–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aitzetmüller, K. An unusual fatty acid pattern in Eranthis seed oil. Lipids 1996, 31, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Hui, Y.; Wang, X.; Yu, Z.; Fan, X.; Cui, B.; Zhao, T.; Mao, L.; Feng, H.; Lin, L.; Yu, Q.; et al. Scoparone as a therapeutic drug in liver diseases: Pharmacology, pharmacokinetics and molecular mechanisms of action. Pharm. Res. 2020, 160, 105170. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, Q.; Wang, Q.; Li, Y.; Zhang, B. Fraxetin inhibits the proliferation of RL95-2 cells through regulation of metabolism. Int. J. Clin. Exp. Pathol. 2020, 13, 1500–1505. [Google Scholar] [PubMed]
- Küpeli Akkol, E.; Genç, Y.; Karpuz, B.; Sobarzo-Sánchez, E.; Capasso, R. Coumarins and Coumarin-Related Compounds in Pharmacotherapy of Cancer. Cancers 2020, 12, 1959. [Google Scholar] [CrossRef]
- Srivastava, S.D.; Halwe, K.; Srivastava, S.K. New coumarins from Limonia crenulata. Fitoterapia 1997, 68, 410–412. [Google Scholar]
- Chakraborty, D.P.; Sen, M.; Bose, P.K. On the antibiotic activity of some natural coumarins. Trans. Bose Rees. Inst. 1961, 24, 31. [Google Scholar]
- Xiong, Y.; Huang, G.; Yao, Z.; Zhao, C.; Zhu, X.; Wu, Q.; Zhou, X.; Li, J. Screening effective antifungal substances from the bark and leaves of Zanthoxylum avicennae by the bioactivity-guided isolation method. Molecules 2019, 24, 4207. [Google Scholar] [CrossRef] [Green Version]
- Goel, R.K.; Maiti, R.N.; Manickam, M.; Ray, A.B. Antiulcer activity of naturally occurring pyrano-coumarin and isocoumarins and their effecton prostanoid synthesis using human colonic mucosa. Indian J. Exp. Biol. 1997, 35, 1080–1083. [Google Scholar]
- Harborne, J.B.; Baxter, H.; Moss, G.P. Phytochemical dictionary A Handbook of Bioactive Compounds from Plants; Taylor and Francis: London, UK, 1999; pp. 381–389. [Google Scholar]
- Harborne, J.B. Biochemistry of Phenolic Compounds; Academic Press: New York, NY, USA, 1964; 88p. [Google Scholar]
- Cao, P.; Pu, X.F.; Peng, S.L.; Zhang, X.R.; Ding, L.S. Chemical constituents from Cimicifuga foetida. J. Asian Nat. Prod. Res. 2005, 7, 145–149. [Google Scholar] [CrossRef]
- Lu, L.; Chen, J.C.; Li, Y.; Qing, C.; Wang, Y.Y.; Nian, Y.; Qiu, M.H. Studies on the constituents of Cimicifuga foetida collected in Guizhou province and their cytotoxic activities. Chem. Pharm. Bull. 2012, 60, 571–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huyen, C.T.T.; Luyen, B.T.T.; Khan, G.J.; Oanh, H.V.; Hung, T.M.H.; Li, H.J.; Li, P. Chemical comstituents from Cimicifuga dahurica and their anti-proliferative effects on MCF-7 breast cancer cells. Molecules 2018, 23, 1083. [Google Scholar] [CrossRef] [Green Version]
- Shi, Q.Q.; Lu, J.; Peng, X.R.; Li, D.S.; Zhou, L.; Qiu, M.H. Cimitriteromone A-G, macromolecular triterpenoid-chromone hybrids from rhizomes of Cimicifuga foetida. J. Org. Chem. 2018, 83, 10359–10369. [Google Scholar] [CrossRef]
- Shi, Q.Q.; Gao, Y.; Lu, J.; Zhou, L.; Qiu, M.H. Two new triterpenoid-chromone hybrids from the rhizomes of Actaea cimicifuga L. (syn. Cimicifuga foetida L.) and their cytotoxic activities. Nat. Prod. Res. 2022, 36, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Al-Khateb, E.H.; Al-Shamma, A.A.; Nehar, B.A. A study of the furocoumarin derivative of Ruta chalepensis L. (Rutaceae). Iraqi J. Pharm. Sci. 2006, 15, 10–15. [Google Scholar] [CrossRef]
- Kaboodi, P.S.; Moghadamnia, A.A.; Bakhshi, D.; Sefidgar, A.A. A study of phytochemical properties of various extracts of Ammi majus fruit using GC-MS technique. Ecol. Environ. Conserv. 2017, 23, 150–155. [Google Scholar]
- Alam, F.; Khan, G.N.; Asad, M.H.H.B. Psoralea corylifolia L.: Ethnobotanical, biological, and chemical aspects: A review. Phytother. Res. 2018, 32, 597–615. [Google Scholar] [CrossRef]
- Liu, R.; Wu, S.; Sun, A. Separation and purification of four chromones from radix saposhnikoviae by high-speed counter-current chromatography. Phytochem. Anal. 2008, 19, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, A.; Matos, M.J.; Garrido, J.; Uriarte, E.; Borges, F. Chromones: A valid scaffold in medicinal chemistry. Chem. Rev. 2014, 114, 4960–4992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keri, R.S.; Budagumpi, S.; Pai, R.K.; Balakrishna, G. Chromones as a privileged scaffold in drug discovery: A review. Eur. J. Med. Chem. 2014, 78, 340–374. [Google Scholar] [CrossRef] [PubMed]
- Vanguru, M.; Merugu, R.; Garimella, S.; Laxminarayana, E. A review on the synthetic methodologies of chromones. Asian J. Pharm. Clin. Res. 2018, 11, 9–16. [Google Scholar] [CrossRef]
- Travaini, M.L.; Sosa, G.M.; Ceccarelli, E.A.; Walter, H.; Cantrell, C.L.; Carrillo, N.J.; Dayan, F.E.; Meepagala, K.M.; Duke, S.O. Khellin and visnagin, furanochromones from Ammi visnaga (L.) Lam., as potential bioherbicides. Agric. Food Chem. 2016, 64, 9475–9487. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.S.; Chin, Y.W.; Lee, K.; Kim, Y.W.; Choi, B.Y.; Keum, Y.S. Identification of 4-O-β-d-glucosyl-5-O-methylvisamminol as a novel epigenetic suppressor of histone H3 phosphorylation at Ser10 and its interaction with 14-3-3Ɛ. Bioorg. Med. Chem. Lett. 2014, 24, 4763–4767. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, X.; Yu, X.; Liu, H.; Tao, Y.; Jiang, G.; Hong, M. Cimifugin suppresses allergic inflammation by reducing epithelial derived initiative key factors via regulating tight junctions. J. Cell. Mol. Med. 2017, 21, 2926–2936. [Google Scholar] [CrossRef] [PubMed]
- Langlet, O.F.J. Über Chromosomen verhältnisse und systematic der Ranunculaceae. Svensk. Bot. Tidskr. 1932, 26, 318–400. [Google Scholar]
- Tamura, M. Ranunculaceae. In The families and genera of vascular plants, Vol. II Flowering plants, Dicotyledons, Magnoliid, Hamamelid and Caryophyllid Families; Kubitzki, K., Rohwer, J.G., Bittrichet, V., Eds.; Springer: Berlin, Germany, 1993; Volume 2, pp. 563–583. [Google Scholar]
- Hoot, S.B. Phylogeny of the Ranunculaceae based on epidermal microcharacters and micromorphology. Syst. Bot. 1991, 16, 741–755. [Google Scholar] [CrossRef]
- Maior, M.C.; Dobrotă, C. Natural compounds with important medical potential found in Helleborus sp. Cent. Europ. J. Biol. 2013, 8, 272–285. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.C.; Peng, Y.; Yang, M.S.; Xiao, P.G. A preliminary pharmacophylogenetic study of tribe Cimicifugeae (Ranunculaceae). J. Syst. Evol. 2008, 46, 516–536. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Identified Compounds | tR (min) | Calculated Mass | Measured Mass | Delta Mass [Da] | Delta Mass [ppm] | MzCloud Score | Mode |
|---|---|---|---|---|---|---|---|
| Flavonoids | |||||||
| Quercetin * | 11.29 | 302.0426 | 302.0423 | −0.00035 | −1.15 | 99.9 | Positive |
| Isoquercitrin (quercetin-3-O-β-d-glucoside) | 13.01 | 464.0954 | 464.0958 | 0.00035 | 0.75 | 99.2 | Negative |
| Hyperoside * (quercetin 3-O-β-d-galactoside) | 10.95 | 464.0954 | 464.0952 | −0.00024 | −0.51 | 98.2 | Positive |
| Reynoutrin (quercetin-3-O-β-d-xylopyranoside) | 10.95 | 434.0849 | 434.0846 | −0.00031 | −0.72 | 98.7 | Positive |
| Quercetin-6-O-β-d- xylopyranosyl-β-d- glucopyranoside | 11.65 | 596.13773 | 596.13714 | −0.0006 | −1 | 98.1 | Negative |
| Quercetin 3-sambubioside (quercetin-3-O-[β-d-xylosyl- (1→2)-β-d-glucoside]) | 11.66 | 596.1377 | 596.1371 | −0.00060 | −1.00 | 98.1 | Negative |
| Peltatoside (quercetin-3-(6-O-α-l- arabinopyranosyl)-β-d- glucopyranoside)) | 10.13 | 596.1377 | 596.1368 | −0.00084 | −1.42 | - | Positive |
| Rutin * (quercetin 3-O-β-d- rutinoside) | 12.48 | 610.1533 | 610.1524 | −0.00089 | −1.46 | 98.9 | Positive |
| Kaempferol * | 11.90 | 286.0477 | 286.0475 | −0.00023 | −0.80 | 99.0 | Positive |
| Juglalin (kaempferol 3-O-α-l- arabinopyranoside) | 11.90 | 418.0900 | 418.0896 | −0.00033 | −0.79 | 79.8 | Positive |
| Trifolin (kaempferol-3-O-β-d-galactoside) | 11.90 | 448.1005 | 448.1003 | −0.00024 | −0.54 | 98.5 | Positive |
| Carlinoside (luteolin 6-C-β-d- glucopyranoside-8-C-α-l- arabinopyranoside) | 12.56 | 580.1428 | 580.1425 | −0.00023 | −0.40 | - | Negative |
| Cianidanol [(+)-catechin] | 16.53 | 290.0790 | 290.0789 | −0.00003 | −0.11 | - | Positive |
| Auriculoside (7,3′,5′-trihydroxy-4′-methoxyflavan-3′-glucoside) | 19.78 | 450.1526 | 450.1521 | −0.00045 | −0.99 | - | Positive |
| 6-Methoxytaxifolin | 14.55 | 334.0688 | 334.0690 | 0.00016 | 0.48 | - | Negative |
| Aromadendrin ((+)-dihydrokaempferol) | 21.50 | 288.0633 | 288.0633 | −0.00003 | −0.11 | - | Positive |
| Aspalathin | 15.45 | 452.1318 | 452.1317 | −0.00014 | −0.32 | - | Positive |
| Phloridzin (phloretin-2′-O-β-glucoside) | 16.23 | 436.1369 | 436.1368 | −0.00013 | −0.30 | - | Positive |
| Phloretin (dihydroxy naringenin) | 20.80 | 274.0841 | 274.0839 | −0.00019 | −0.69 | - | Positive |
| Fatty acids-related compounds | |||||||
| 12-Oxo-phytodienoic acid | 36.84 | 292.2038 | 292.2037 | −0.00013 | −0.43 | 87.3 | Positive |
| 15-OxoEDE (15-Oxo-11Z,13E-eicosadienoic acid) | 47.47 | 322.2507 | 322.2508 | 0.00006 | 0.2 | 91.7 | Positive |
| 9-oxo-ODA (9-Oxo-10(E),12(E)-octadecadienoic acid) | 39.22 | 294.2194 | 294.2194 | −0.00006 | −0.21 | 95.3 | Positive |
| 9S,13R-12-Oxo-phytodienoic acid | 36.47 | 292.2038 | 292.2037 | −0.00009 | −0.3 | 87.2 | Positive |
| Linolenic acid ethyl ester | 46.47 | 306.2558 | 306.2558 | −0.00008 | −0.28 | 98.3 | Positive |
| Palmitoleic Acid | 45.38 | 254.2245 | 254.2245 | −0.00006 | −0.24 | 86.7 | Positive |
| α-Eleostearic acid | 49.77 | 278.2245 | 278.2244 | −0.00017 | −0.62 | 99.1 | Positive |
| α-Linolenic acid | 45.50 | 278.2245 | 278.2244 | −0.00012 | −0.45 | 99.2 | Positive |
| (+/−)13-HODE (13-hydroxyoctadecadienoic acid) | 38.17 | 296.2351 | 296.2352 | 0.0001 | 0.35 | 92.8 | Negative |
| (15Z)-9,12,13-Trihydroxy-15- octadecenoic acid | 24.63 | 330.2406 | 330.2408 | 0.00019 | 0.57 | 68.5 | Negative |
| 13(S)-HOTrE (13-OH-9Z,11E,15Z-octadecatrienoic acid) | 36.23 | 294.2194 | 294.2195 | 0.00009 | 0.29 | 92.1 | Negative |
| 16-Hydroxyhexadecanoic acid | 44.84 | 272.2351 | 272.2353 | 0.0002 | 0.73 | 89.5 | Negative |
| Corchorifatty acid F | 23.35 | 328.2249 | 328.2249 | 0.00002 | 0.06 | 93 | Negative |
| Pinolenic acid | 45.76 | 278.2245 | 278.2245 | −0.0006 | −0.21 | 94.5 | Negative |
| Amino acid-related compounds | |||||||
| d-(+)-Pyroglutamic Acid | 1.56 | 129.0425 | 129.0428 | 0.00021 | 1.63 | 98.2 | Positive |
| d-(+)-Tryptophan | 6.96 | 204.0898 | 204.0898 | −0.0003 | −0.13 | 99.2 | Positive |
| Isoleucine | 1.93 | 131.0946 | 131.0947 | 0.00014 | 1.10 | 99.2 | Positive |
| l-Phenylalanine | 4.45 | 165.0789 | 165.0790 | 0.00003 | 0.19 | 98.6 | Positive |
| L-Tyrosine | 1.93 | 181.0738 | 181.0740 | 0.00015 | 0.81 | 98.4 | Positive |
| d-(-)-Glutamine | 1.56 | 146.0691 | 146.0684 | −0.00073 | −4.99 | 66.9 | Negative |
| Organic acids | |||||||
| Citric acid | 2.00 | 192.027 | 192.0264 | −0.00056 | −2.93 | 99.6 | Negative |
| d-α-Hydroxyglutaric acid | 1.99 | 148.0371 | 148.0364 | −0.00071 | −4.81 | 68.8 | Negative |
| Gluconic acid | 1.61 | 196.0583 | 196.0577 | −0.00057 | −2.89 | 99 | Negative |
| Sugars | |||||||
| α-Lactose | 1.61 | 342.1162 | 342.1160 | −0.00014 | −0.41 | 81.8 | Positive |
| d-(+)-Galactose | 1.61 | 180.0633 | 180.0629 | −0.00047 | −2.61 | 60.5 | Negative |
| α.α-Trehalose | 1.64 | 342.1162 | 342.1160 | −0.00019 | −0.55 | 98.9 | Negative |
| Phenylpropanoid | |||||||
| 6-Gingerol | 28.34 | 294.1831 | 294.1830 | −0.00004 | −0.14 | 81.8 | Positive |
| Coumarins | |||||||
| 5,7-Dihydroxy-4-methylcoumarin | 17.32 | 192.0422 | 192.0423 | 0.00007 | 0.35 | 99.9 | Positive |
| Scoparone (6,7-dimethoxycoumarin) | 24.79 | 206.0579 | 206.0579 | 0.00002 | 0.12 | 98.8 | Positive |
| Fraxetin (7,8-dihydroxy-6-methoxycoumarin) | 11.39 | 208.0371 | 208.0371 | −0.00005 | −0.24 | 96.1 | Positive |
| Luvangetin (10-methoxy-2,2-dimethylpyrano[3,2-g]chromen-8-one) | 32.62 | 258.0892 | 258.0891 | −0.00005 | −0.19 | 93.6 | Positive |
| Furochromones | |||||||
| Khelloside (7-hydroxymethyl-4-methoxy-5H-furo[3,2-g](1)benzopyran-5-one glucoside) | 12.02 | 408.1056 | 408.1048 | −0.00077 | −1.9 | 99.5 | Positive |
| Khellin (4,9-dimethoxy-7-methyl-5H-furo[3,2-g]chromen-5-one) | 21.003 | 260.0684 | 260.0680 | −0.00039 | −1.5 | 99.8 | Positive |
| Visnagin (4-methoxy-7-methyl-5H-furo[3,2-g]chromen-5-one) | 21.138 | 230.0579 | 230.0576 | −0.00023 | −0.98 | 99.3 | Positive |
| Cimifugin ((2S)-7-(hydroxymethyl)-2-(1-hydroxy-1-methyl-ethyl)-4-methoxy-2,3-dihydrofuro[3,2-g]chromen-5-one) | 13.544 | 306.1103 | 306.1097 | −0.00061 | −1.98 | 95.1 | Positive |
| Methoxsalen (9-methoxyfuro[3,2-g]chromen-7-one) | 27.757 | 216.0422 | 216.0422 | 0 | 0.02 | 99.3 | Positive |
| 5-O-Methylvisammioside (4-O-β-d-glucosyl-5-O-methylvisamminol) | 14.538 | 452.1682 | 452.1677 | −0.00048 | −1.05 | 99.1 | Positive |
| Visamminol-3′-O-glucoside (4-hydroxy-2-(2-hydroxypropan-2-yl)-methyl-2,3-dihydrofuro[3,2-g] chromen-5-one) | 16.894 | 438.1526 | 438.1523 | −0.00022 | −0.5 | 98.2 | Positive |
| ID | Compound (ID) | Structure | Sourse | Ref. |
|---|---|---|---|---|
| CHROMONES | ||||
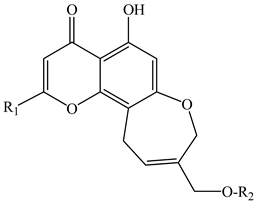 | ||||
| 1 | 8,11-dihydro-5-hydroxy-2,9-dihydroxymethyl-4H-pyrano[2,3-g][1]benzoxepin-4-one | R1 = O-CH2-OH R2 = H | E. cilicica (tubers) | [16] |
| 2 | Eranthin (5-hydroxy-9-hydroxymethyl-2-methyl-8,11-dihydro-4H-pyrano[2,3-g][1]benzoxepin-4-one) | R1 = CH3 R2 = H | E. hyemalis (rhizome) | [14] |
| 3 | Eranthin-β-d-glucoside (9-{[(β-d-glucopyranosvl)oxy]methyl}-8,11-dihydro-5-hydroxy-2-methyl-4H-pyrano[2,3-g][1]benzoxepin-4-one) | R1 = CH3 R2 = β-d-glucopyranosyl | E. hyemalis (rhizome) E. hyemalis (tubers) | [14,15] |
| 4 | Eranthin 9-β-d-glucopyranosyl-(1→6)-β-d-glucopyranoside | R1 = CH3 R2 = glucopyranosyl-(1→6)-glucopyranoside | E. cilicica (tubers) E. hyemalis (tubers) | [15,16] |
| 5 | Eranthin β-d-gentiobioside (9-{[(β-d-gentiobiosyl)oxy]methyl}-8,11-dihydro-5-hydroxy-2-methyl-4H-pyrano[2,3-g][1]benzoxepin-4-one) | R1 = CH3 R2 = β-D-gentiobiosyl | E. hyemalis (tubers) | [15] |
| 6 | 2-C-Hydroxyeranthin β-d-glucopyranoside (9-{[(β-d-glucopyranosyl)oxy]methyl}-8,11-dihydro-5-hydroxy-2-(hydroxymethyl-4H-pyrano[2,3-g][1]benzoxepin-4-one) | R1 = CH2-OH R2 = β-d-glucopyranosyl | E. hyemalis (tubers) | [15] |
 | ||||
| 7 | 5,7-dihydroxy-8-[(2E)-4-hydroxy-3-methylbut-2-enyl]-2-methyl-4H-1-benzopyran-4-one | R1 = CH3 R2 = R3=H | E. cilicica (tubers) | [16] |
| 8 | 5,7-dihydroxy-2-hydroxymethyl-8-[(2E)-4-hydroxy-3-methylbut-2-enyl]-4H-1-benzopyran-4-one | R1 = CH2-OH R2 = R3=H | E. cilicica (tubers) | [16] |
| 9 | 7-[(β-d-glucopyranosyl)oxy]-5-hydroxy-8-[(2E)-4-hydroxy-3-methylbut-2-enyl]-2-methyl-4H-1-benzopyran-4-one | R1 = CH3 R2 = β-d-glucopyranosyl R3 = H | E. cilicica (tubers) | [16] |
| 10 | 7-[(β-d-glucopyranosyl)oxy]-5-hydroxy-2-hydroxymethyl-8-[(2E)-4-hydroxy-3-methylbut-2-enyl]-4H-1-benzopyran-4-one | R1 = CH2-OH R2 = β-d-glucopyranosyl R3 = H | E. cilicica (tubers) | [16] |
| 11 | 7,8-Secoeranthin β-d-glucoside (8-{(2E)-4-[(β-d-glucopyranosyl)oxy]-3-methylbut-2-enyl}-5,7-dihydroxy-2-methyl-4H-1-benzopyran-4-one) | R1 = CH3 R2 = H R3 = β-D-glucopyranosyl | E. hyemalis (tubers) | [15] |
| 12 | 2-C-Hydroxy-7,8-Secoeranthin β-d-glucoside (8-{(2E)-4-[(β-d-glucopyranosyl)oxy]-3-methylbut-2-enyl}-5,7-dihydroxy-2-(hydroxymethyl)-4H-1-benzopyran-4-one) | R1 = CH2-OH R2 = H R3 = β-D-glucopyranosyl | E. hyemalis (tubers) | [15] |
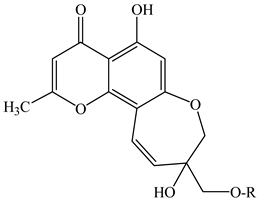 | ||||
| 13 | 9-[(O-β-d-glucopyranosyl-(1→6)-β-d-glucopyranosyl)oxy]methyl-8,11-dihydro-5,9-dihydroxy-2-methyl-4H-pyrano[2,3-g][1]benzoxepin-4-one | R = glucopyranosyl-(1→6)-glucopyranosyl | E. cilicica (tubers) | [16] |
| 14 | 8,11-dihydro-5,9-dihydroxy-9-hydroxymethyl-2-methyl-4H-pyrano[2,3-g][1]benzoxepin-4-one | R = H | E. cilicica (tubers) | [16] |
| FUROCHROMONES | ||||
 | ||||
| 15 | Cimifugin (2S)-7-(hydroxymethyl)-2-(2-hydroxypropan-2-yl)-4-methoxy-2,3-dihydrofuro[3,2g]chromen-5-one) | R1 = H R2 = O-CH3 | E. pinnatifida (leaves, stems) E. cilicica (tubers) | [13,16] |
| 16 | Cimicifugin β-d-glucopyranoside (7-{[(β-d-glucopyranosy1)oxy]methyl}-2,3-dihydro-2-(l-hydroxy-1-methylethyl)-4-methoxy-5H-furo[3,2-g][1]benzopyran-5-one) | R1 = β-D-glucopyranosyl R2 = O-CH3 | E. hyemalis | [15] |
| 17 | Norcimifugin (2S)-4-hydroxy-7-(hydroxymethyl)-2-(2-hydroxypropan-2-yl)-2,3-dihydrofuro[3,2-g]-chromen-5-one) | R1 = H R2 = OH | E. pinnatifida (leaves, stems) | [13] |
 | ||||
| 18 | Visnagin (4-methoxy-7-methyl-5H-furo[3,2-g]chromen-5-one) | R1 = R2=H R3 = O-CH3 | E. hyemalis | [33] |
| 19 | Khellin (4,9-dimethoxy-7-methyl-5H-furo[3,2-g]chromen-5-one) | R1 = O-CH3 R2 = H R3 = O-CH3 | E. hyemalis | [33] |
| 20 | Khellol (7-(hydroxymethyl)-4-methoxyfuro[3,2-g]chromen-5-one) | R1 = H R2 = CH2-OH R3 = O-CH3 | E. pinnatifida (leaves, stems) | [13] |
| 21 | Khellol glucoside (khellinin; 2-hydroxymethyl-5-methoxyfuranochrome glucoside) | R1 = H R2 = CH2-β-d-glucopyranoside R3 = O-CH3 | E. hyemalis (leaves, flowers) | [12] |
| 22 | Norkhellol (4-hydroxy-7-(hydroxymethyl)-5H-furo[3,2-g][1]benzopyran-5-one) | R1 = H R2 = CH2-OH R3 = OH | E. pinnatifida (leaves, stems) | [13] |
| 23 | Norammiol (4-hydroxy-7(hydroxymethyl)-9-methoxy-5H-furo[3,2-g][1]-benzopyran-5-one) | R1 = O-CH3 R2 = CH2-OH R3 = OH | E. pinnatifida (leaves, stems) | [13] |
| 24 | 7-[(O-β-d-glucopyranosyl-(1→6)-β-d-glucopyranosyl)oxy]methyl-4-hydroxy-5H-furo[3,2-g][1]benzopyran-5-one (24) | R1 = H R2 = CH2-O-glucopyranosyl-(1→6)-glucopyranosyl R3 = OH | E. cilicica (tubers) | [16] |
| Sample No. | Locality; Coordinates | Habitat | Date |
|---|---|---|---|
| 1 | Kyrgyzstan, Chuya region, Issyk-atinskii district, Karandolot tract; 42°44′22″ N, 74°55’50″ E | foot of the mount | 22 March 2019 |
| 2 | Kyrgyzstan, Talas region, Kara-Buurinskii district, west of Kirovskoe reservoir; 42°37′57″ N, 71°34′47″ E | steppe | 26 March 2019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erst, A.S.; Chernonosov, A.A.; Petrova, N.V.; Kulikovskiy, M.S.; Maltseva, S.Y.; Wang, W.; Kostikova, V.A. Investigation of Chemical Constituents of Eranthis longistipitata (Ranunculaceae): Coumarins and Furochromones. Int. J. Mol. Sci. 2022, 23, 406. https://doi.org/10.3390/ijms23010406
Erst AS, Chernonosov AA, Petrova NV, Kulikovskiy MS, Maltseva SY, Wang W, Kostikova VA. Investigation of Chemical Constituents of Eranthis longistipitata (Ranunculaceae): Coumarins and Furochromones. International Journal of Molecular Sciences. 2022; 23(1):406. https://doi.org/10.3390/ijms23010406
Chicago/Turabian StyleErst, Andrey S., Alexander A. Chernonosov, Natalia V. Petrova, Maxim S. Kulikovskiy, Svetlana Yu. Maltseva, Wei Wang, and Vera A. Kostikova. 2022. "Investigation of Chemical Constituents of Eranthis longistipitata (Ranunculaceae): Coumarins and Furochromones" International Journal of Molecular Sciences 23, no. 1: 406. https://doi.org/10.3390/ijms23010406
APA StyleErst, A. S., Chernonosov, A. A., Petrova, N. V., Kulikovskiy, M. S., Maltseva, S. Y., Wang, W., & Kostikova, V. A. (2022). Investigation of Chemical Constituents of Eranthis longistipitata (Ranunculaceae): Coumarins and Furochromones. International Journal of Molecular Sciences, 23(1), 406. https://doi.org/10.3390/ijms23010406