
Activated ERK Signaling Is One of the Major Hub Signals Related to the Acquisition of Radiotherapy-Resistant MDA-MB-231 Breast Cancer Cells


 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
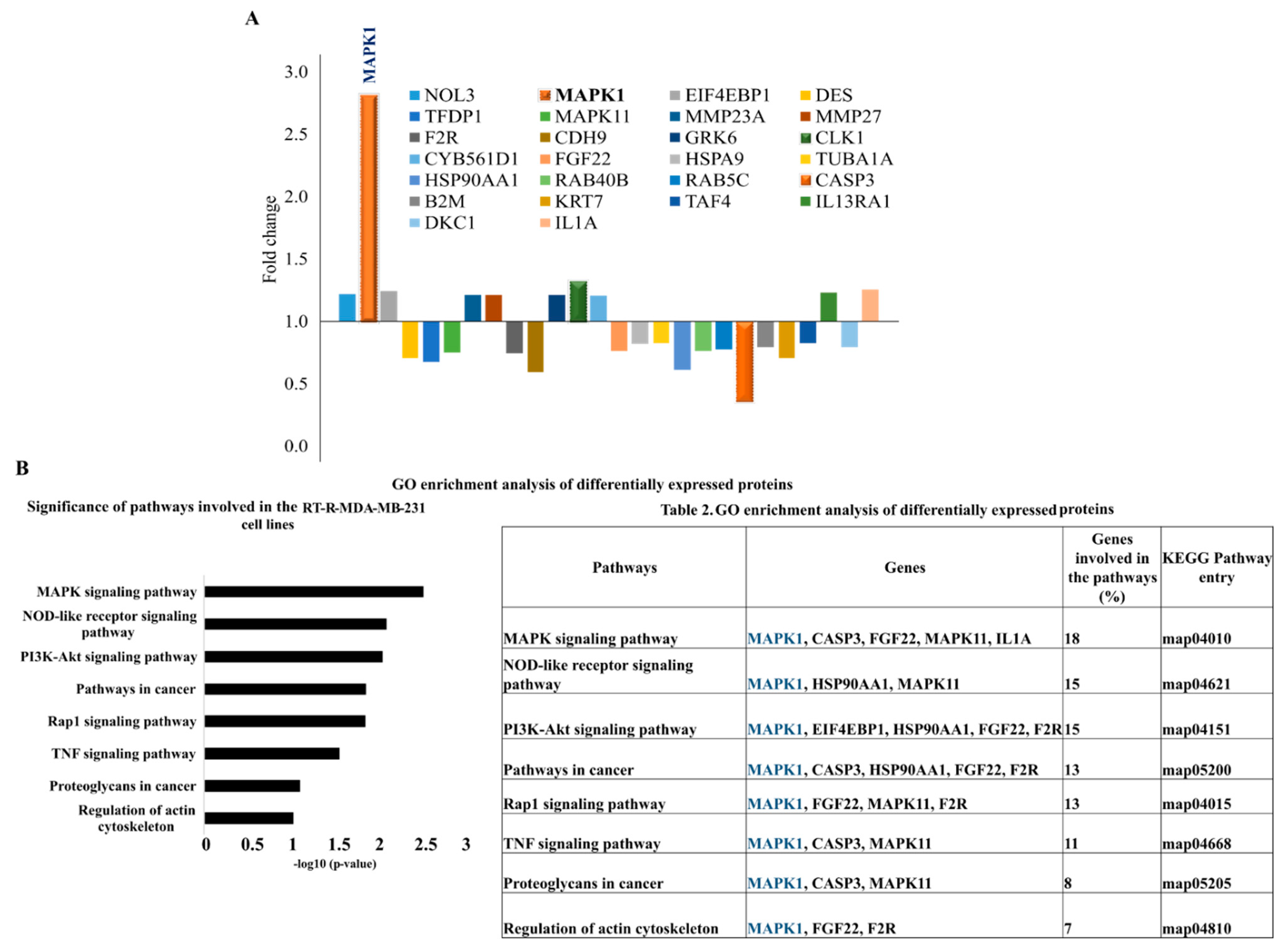
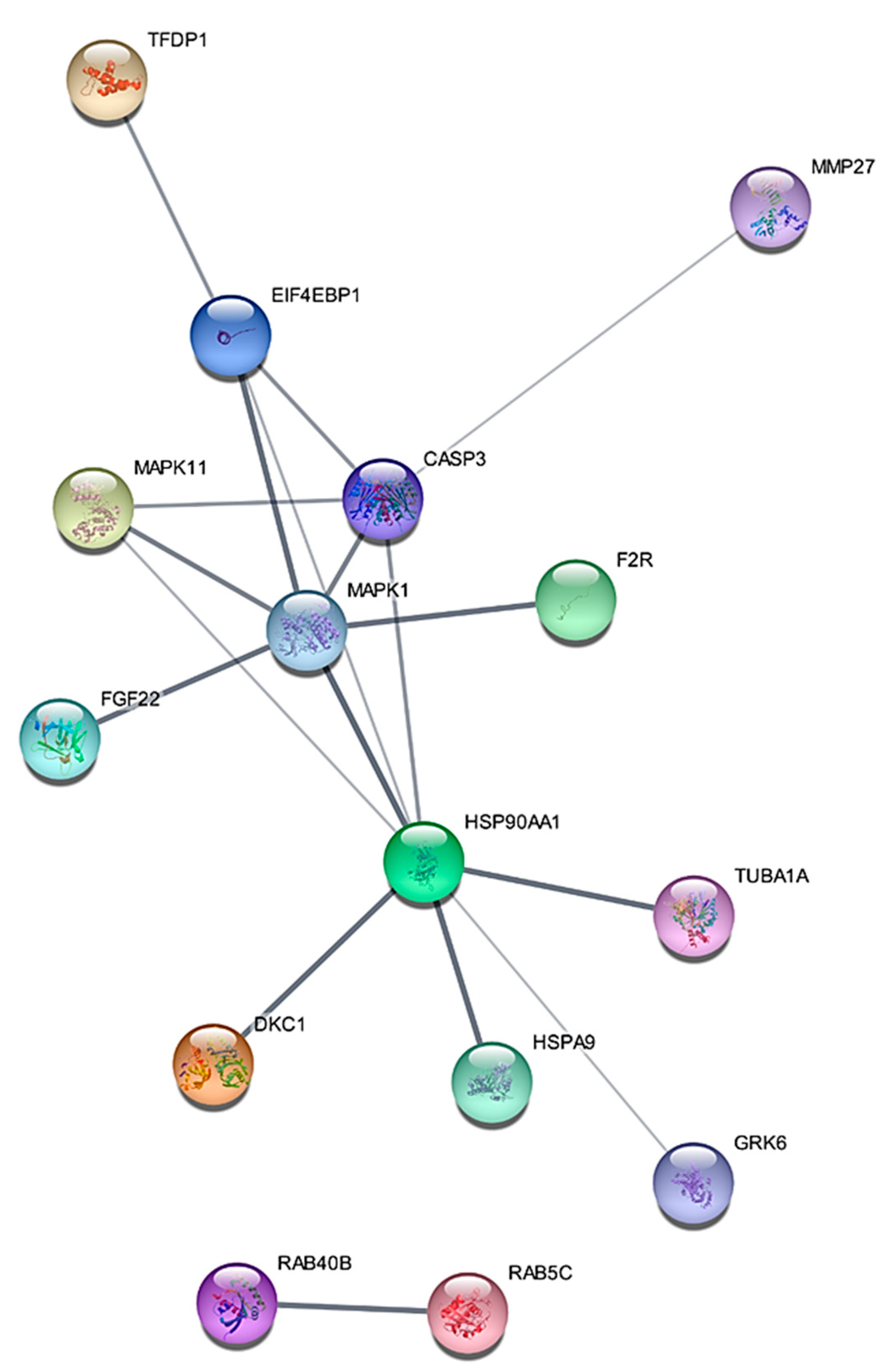
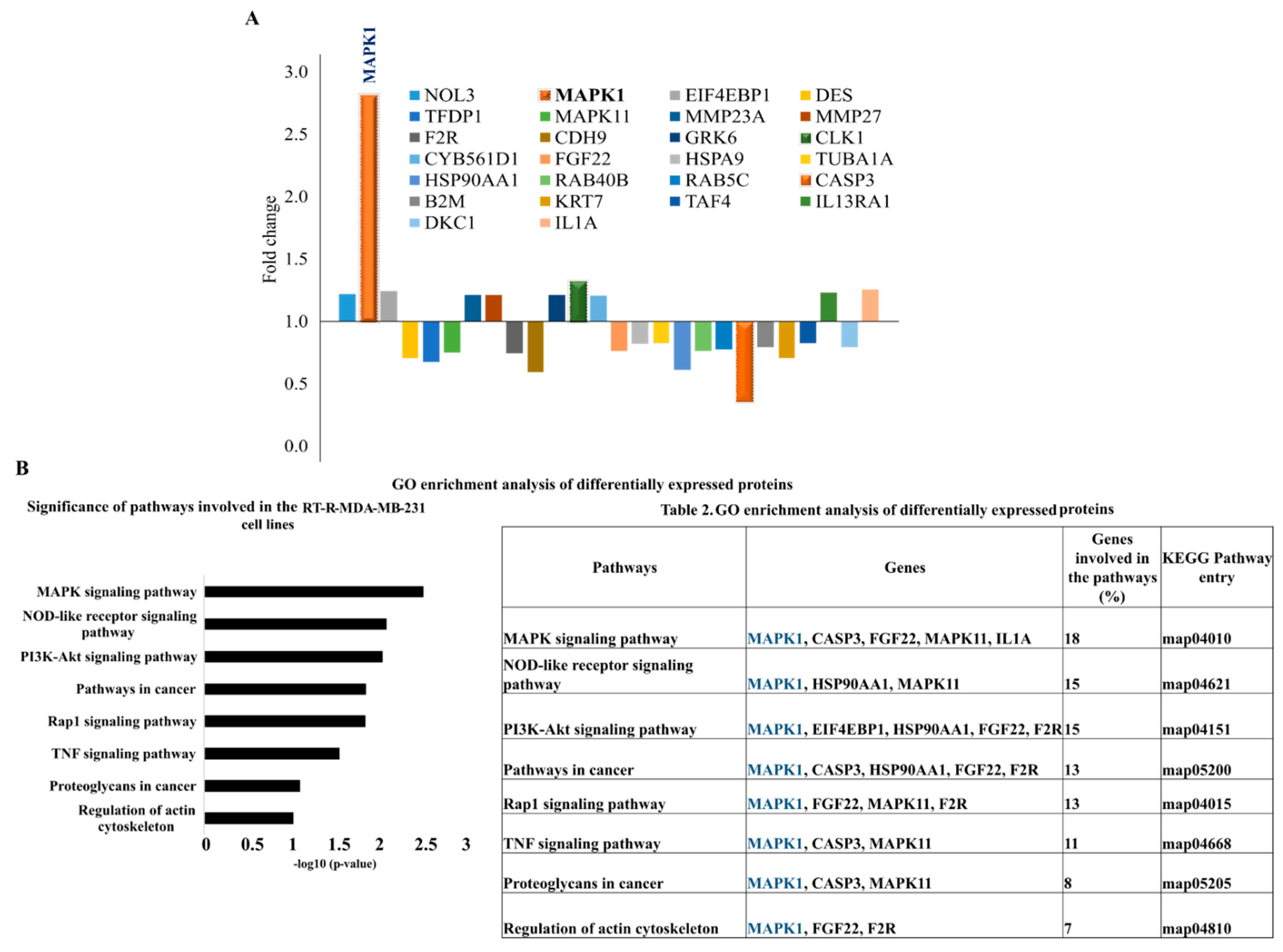
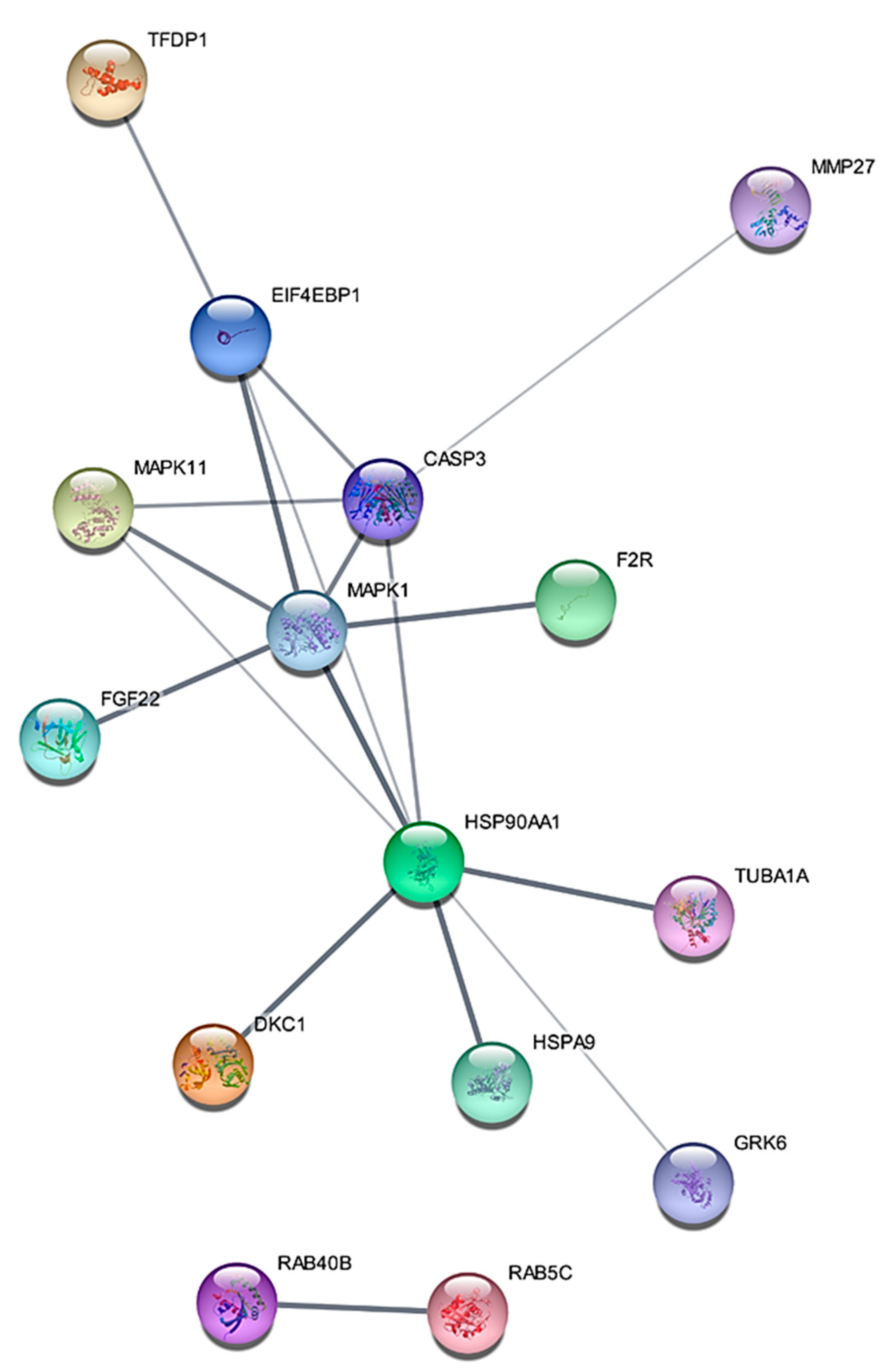
2.1. Proteomic Profiling of RT-R-MDA-MB-231 Cell Lines
2.2. MAPK1 Is the Most Important Signaling Pathway in Acquiring Radio-Resistant RT-R-MDA-MB-231 Cells
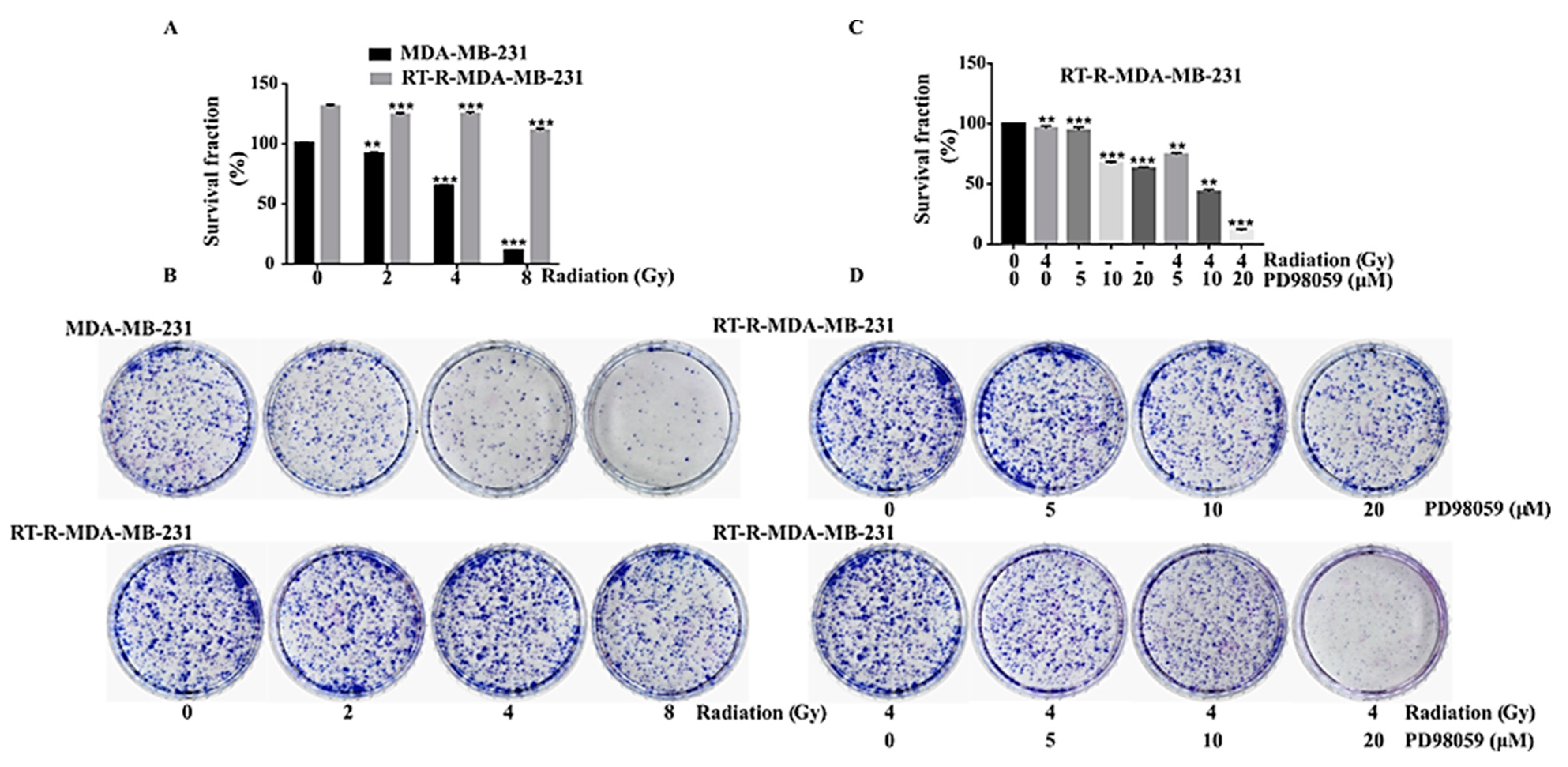
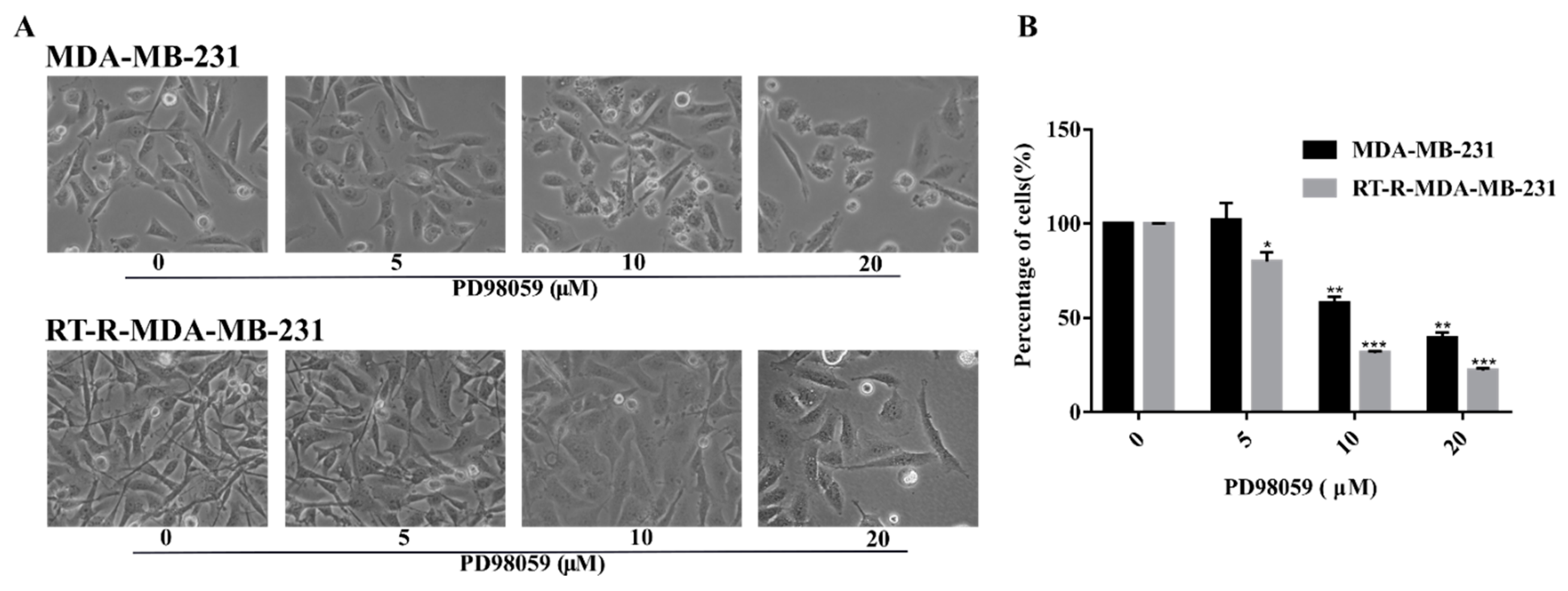
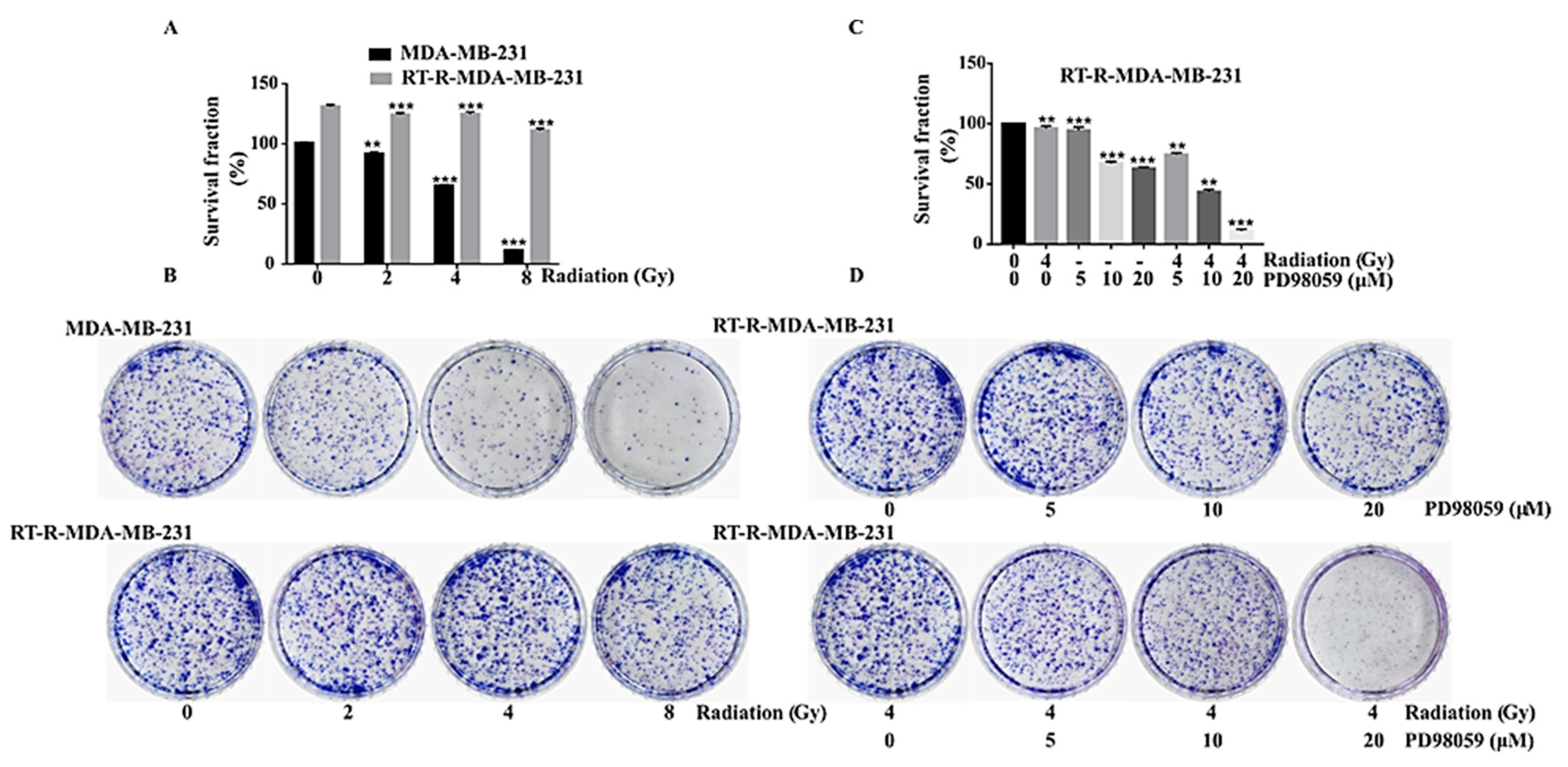
2.3. ERK Signaling Was Important in the Cell Survival of RT-R-MDA-MB-231 Cells, and the Inhibition of MEK/ERK Signaling Reversed the Radio-Resistance of MDA-MB-231 Cells
2.4. Inhibition of ERK Signaling Reversed the Radio-Resistance of RT-R-MDA-MB-231 Cells
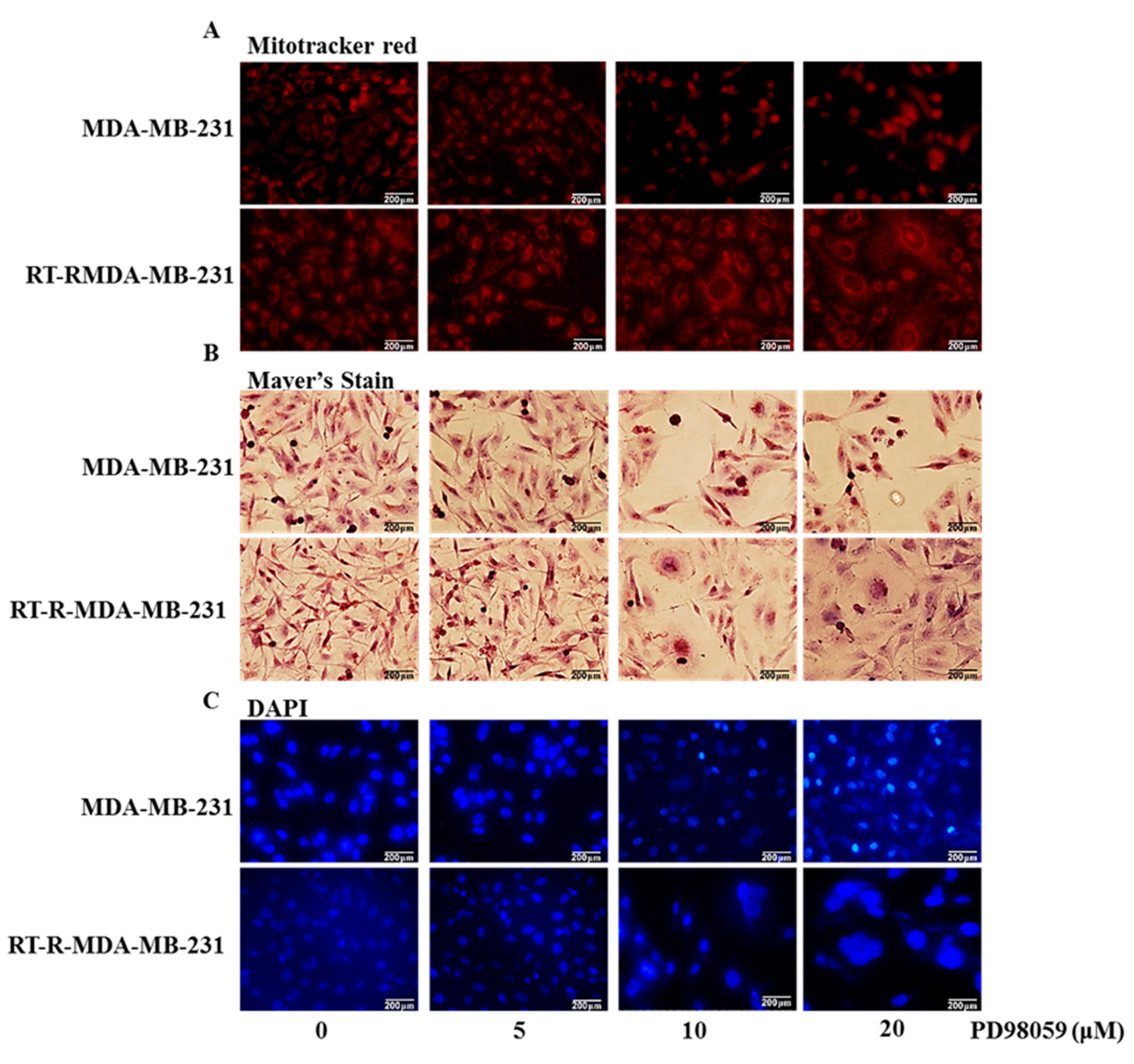
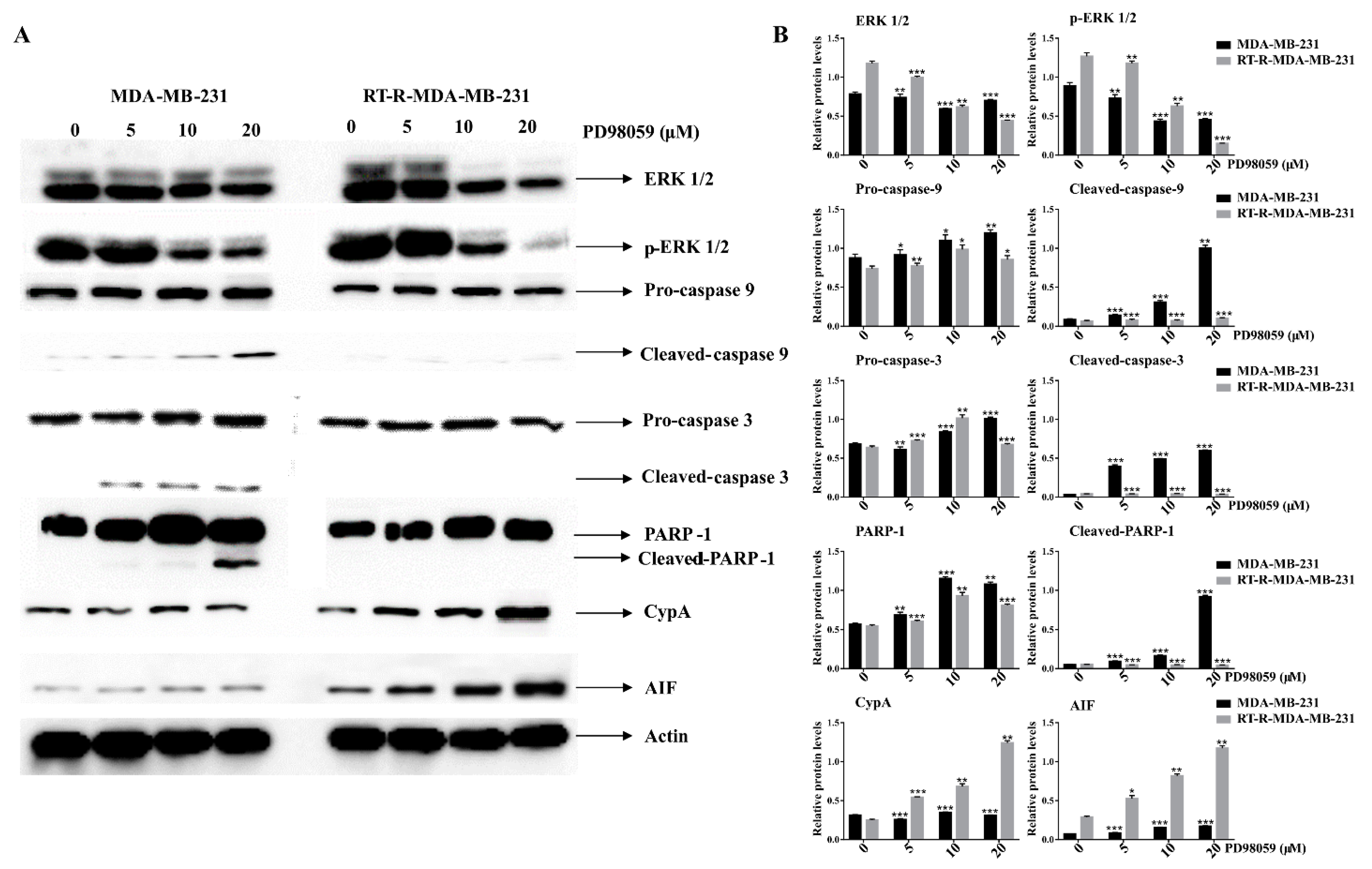
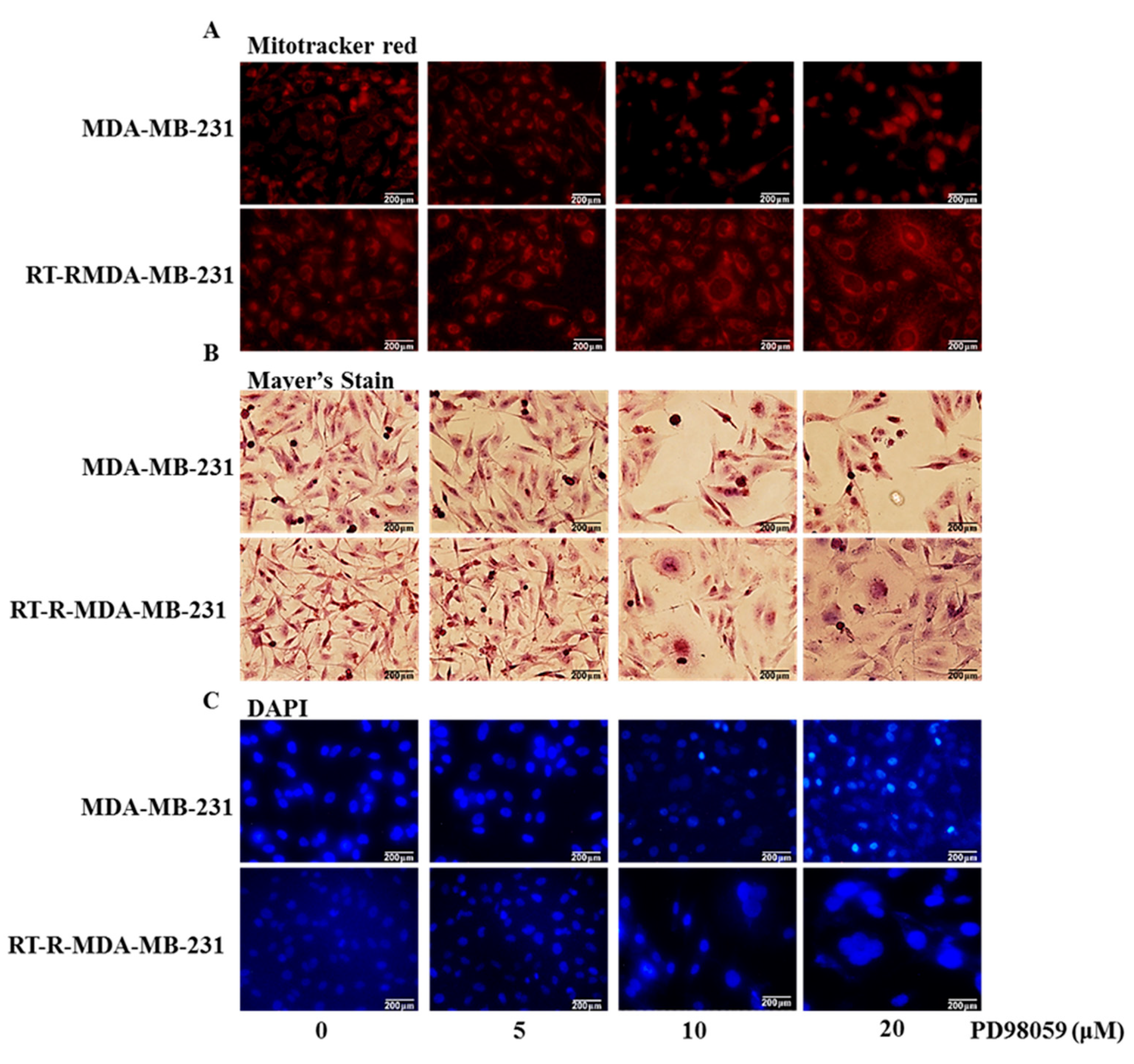
2.5. Inhibition of ERK Signaling-Induced Necroptosis of RT-R-MDA-MB-231 Cells While It Induced the Apoptosis of p-MDA-MB-231 Cells
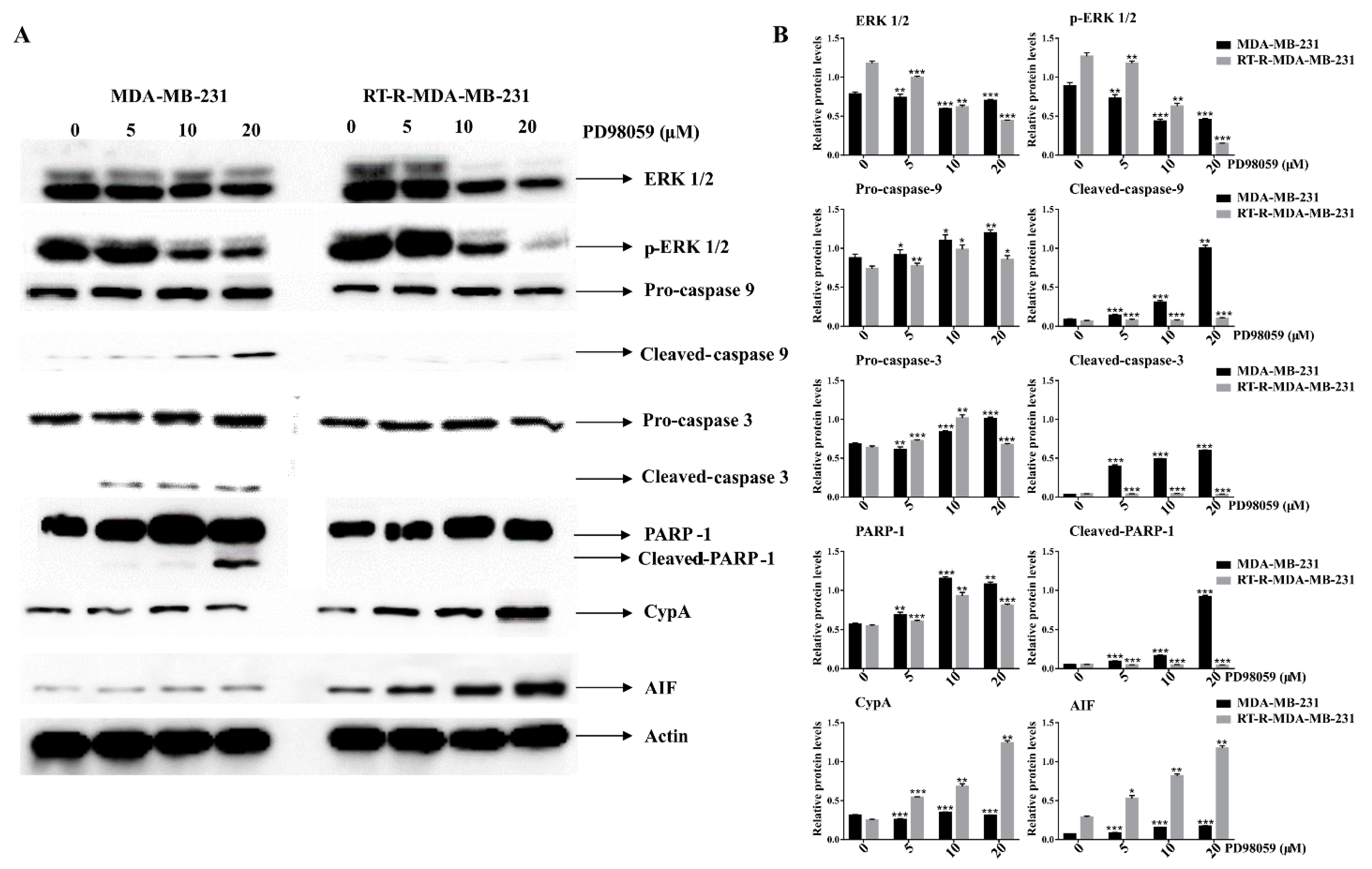
2.6. ERK Inhibition Induced Caspase Activation and PARP-1 Cleavages in p-MDA-MB-231 Cells, While It Did Increase the Expression of Cyclophilin A (CypA) and AIF in RT-R-MDA-MB-231 Cells
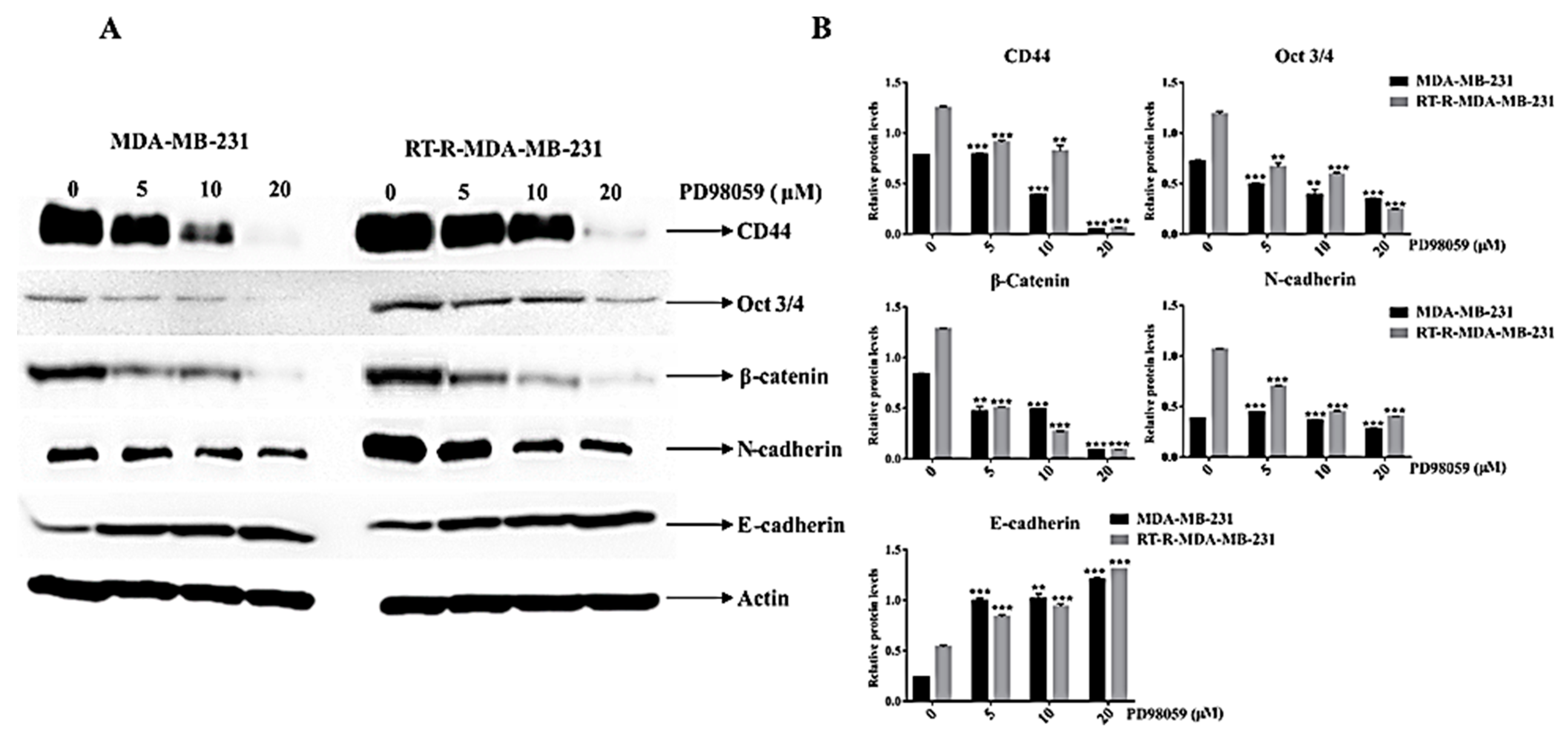
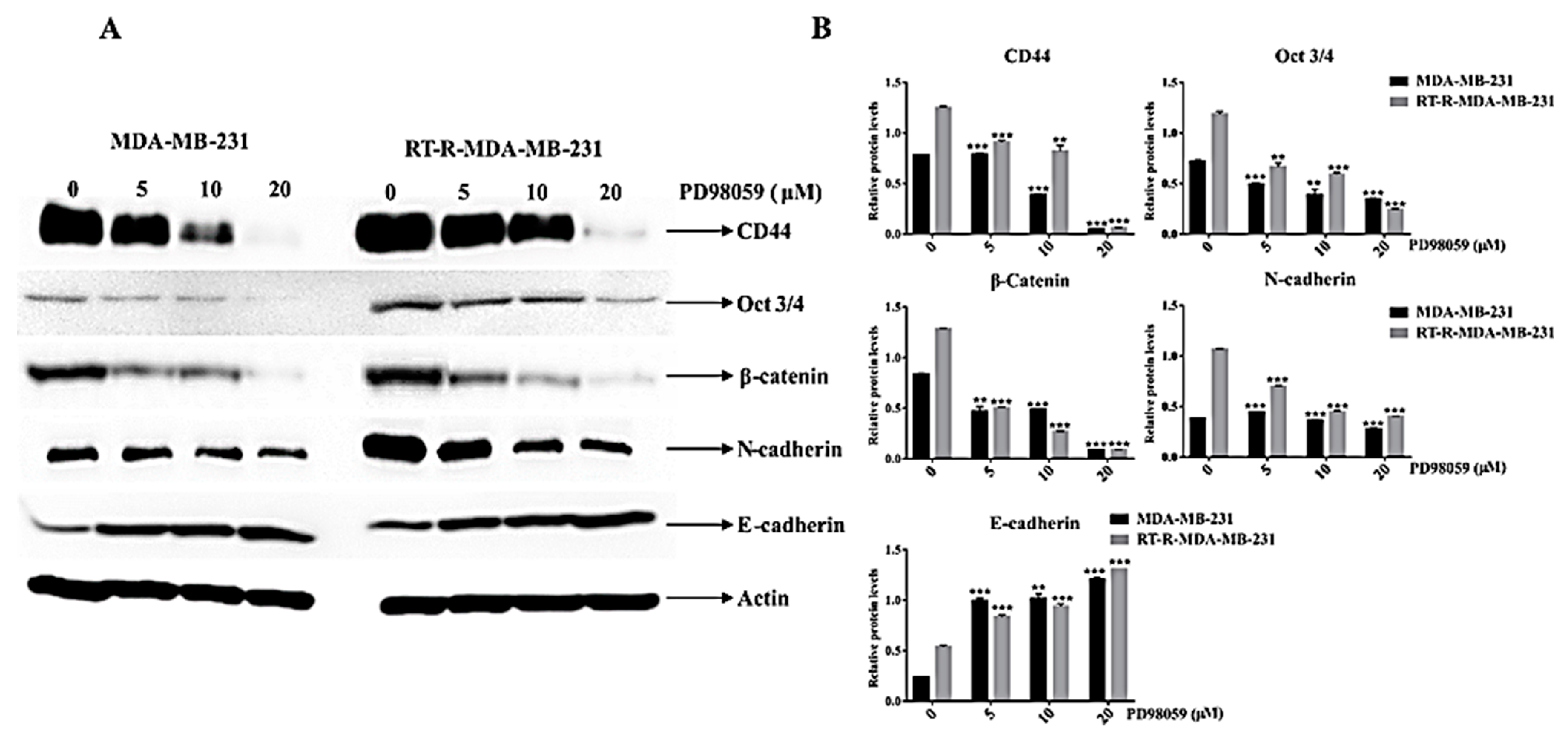
2.7. ERK Inhibition Reduced the Expression of Cancer Stem Cell (CSC) Markers (CD44 and Oct ¾) and the EMT Phenotype, Which Is Closely Related to the Radio-Resistance of RT-R-MDA-MB-231 Cells
3. Material and Methods
3.1. Protein Array Analysis
3.2. Bioinformatics Analysis
3.3. Cell Culture
3.4. Cell Viability Assay
3.5. Colony Formation Assay
3.6. Cytochemical Staining Methods
3.6.1. Mitotracker Red Analysis
3.6.2. Hematoxylin Staining
3.6.3. DAPI (4′,6-diamidino-2-phenylindole) Staining
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CSCs | Cancer stem cells |
| CypA | Cyclophilin A |
| AIF | Apoptosis-inducing factor |
| EMT | Epithelial-to-mesenchymal transition |
| ERK | Extracellular-signal-regulated kinase |
| DDR | Damage response |
| IR | Irradiation |
| PI3K | Phosphatidylinositol 3-kinase |
| MAPK | Mitogen-activated protein kinase |
| Wnt | Wingless-related integration site |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| STRING | Search Tool for the Retrieval of Interacting Genes |
| DMSO | Dimethyl sulfoxide is an organosulfur |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| PBS | Phosphate-buffered saline |
| DAPI | 4′,6-diamidino-2-phenylindole |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| SDS | Sodium dodecyl sulphate |
| RT | Radiotherapy |
| RT-R | Radiotherapy-resistant |
| SD | Standard deviation |
| CLK1 | CDC-like kinases |
| FGF22 | Fibroblast growth factor 22 |
| F2R | Coagulation factor II (thrombin) receptor |
| Gy | Gray |
| CCD | Charge-coupled device |
| JAK2 | Janus-activated kinase 2 |
| PI3K-Akt | Phosphatidylinositol 3-kinase (PI3K)-protein kinase B (AKT) |
| STAT | Signal transducer and activator of transcription |
| NF-κB | Nuclear factor-κB |
| EGFR | Epidermal growth factor receptor |
| SOS | Son of Sevenless |
| B-raf | B-Rapidly Accelerated Fibrosarcoma |
| Ras | Rat sarcoma |
| MEK | Mitogen-activated protein kinase kinase |
| CASP9 | Caspase 9 |
| IFIT2 | interferon-induced protein with tetratricopeptide repeats 2 |
| TRIB3 | Tribbles homolog 3 |
| ESM-1 | Endothelial cell-specific molecule-1 |
| DLX2 | Vertebrate distal-less homeobox 2 |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, C.L.; Olin, J.L. Triple negative breast cancer: A brief review of its characteristics and treatment options. J. Pharm. Pract. 2012, 25, 319–323. [Google Scholar] [CrossRef]
- Brownlee, Z.; Garg, R.; Listo, M.; Zavitsanos, P.; Wazer, D.E.; Huber, K.E. Late complications of radiation therapy for breast cancer: Evolution in techniques and risk over time. Gland Surg. 2018, 7, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Langlands, F.E.; Horgan, K.; Dodwell, D.D.; Smith, L. Breast cancer subtypes: Response to radiotherapy and potential radiosensitisation. Br. J. Radiol. 2013, 86, 20120601. [Google Scholar] [CrossRef]
- Marampon, F.; Ciccarelli, C.; Zani, B.M. Biological Rationale for Targeting MEK/ERK Pathways in Anti-Cancer Therapy and to Potentiate Tumour Responses to Radiation. Int. J. Mol. Sci. 2019, 20, 2530. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Ding, L.; Li, Q.; Yao, H. miR-668 enhances the radioresistance of human breast cancer cell by targeting IκBα. Breast Cancer 2017, 24, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Wei, H.; Ling, R.; Lv, Y. Opposite effects of TNF-α on proliferation via ceramide in MDA-MB-231 and MCF-7 breast cancer cell lines. Int. J. Clin. Exp. Med. 2018, 11, 9239–9247. [Google Scholar]
- Ko, Y.S.; Jin, H.; Lee, J.S.; Park, S.W.; Chang, K.C.; Kang, K.M.; Jeong, B.K.; Kim, H.J. Radioresistant breast cancer cells exhibit increased resistance to chemotherapy and enhanced invasive properties due to cancer stem cells. Oncol. Rep. 2018, 40, 3752–3762. [Google Scholar] [CrossRef]
- Kopf, E.; Zharhary, D. Antibody arrays—An emerging tool in cancer proteomics. Int. J. Biochem. Cell Biol. 2007, 39, 1305–1317. [Google Scholar] [CrossRef]
- Bernichon, E.; Vallard, A.; Wang, Q.; Attignon, V.; Pissaloux, D.; Bachelot, T.; Heudel, P.E.; Ray-Coquard, I.; Bonnet, E.; de la Fouchardière, A.; et al. Genomic alterations and radioresistance in breast cancer: An analysis of the ProfiLER protocol. Ann. Oncol. 2017, 28, 2773–2779. [Google Scholar] [CrossRef] [PubMed]
- Strasser-Wozak, E.M.; Hartmann, B.L.; Geley, S.; Sgonc, R.; Böck, G.; Santos, A.J.; Hattmannstorfer, R.; Wolf, H.; Pavelka, M.; Kofler, R. Irradiation induces G2/M cell cycle arrest and apoptosis in p53-deficient lymphoblastic leukemia cells without affecting Bcl-2 and Bax expression. Cell Death Differ. 1998, 5, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Bernhard, E.J.; Maity, A.; Muschel, R.J.; McKenna, W.G. Effects of ionizing radiation on cell cycle progression. Radiat. Environ. Biophys. 1995, 34, 79–83. [Google Scholar] [CrossRef]
- Pawlik, T.M.; Keyomarsi, K. Role of cell cycle in mediating sensitivity to radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2004, 59, 928–942. [Google Scholar] [CrossRef] [PubMed]
- Meier, K.E.; Gause, K.C.; Wisehart-Johnson, A.E.; Gore, A.C.S.; Finley, E.L.; Jones, L.G.; Bradshaw, C.D.; McNair, A.F.; Ella, K.M. Effects of Propranolol on Phosphatidate Phosphohydrolase and Mitogen-Activated Protein Kinase Activities in A7r5 Vascular Smooth Muscle Cells. Cell. Signal. 1998, 10, 415–426. [Google Scholar] [CrossRef]
- Munshi, A.; Ramesh, R. Mitogen-activated protein kinases and their role in radiation response. Genes Cancer 2013, 4, 401–408. [Google Scholar] [CrossRef]
- Dudley, D.T.; Pang, L.; Decker, S.J.; Bridges, A.J.; Saltiel, A.R. A synthetic inhibitor of the mitogen-activated protein kinase cascade. Proc. Natl. Acad. Sci. USA 1995, 92, 7686. [Google Scholar] [CrossRef] [Green Version]
- Favata, M.F.; Horiuchi, K.Y.; Manos, E.J.; Daulerio, A.J.; Stradley, D.A.; Feeser, W.S.; Van Dyk, D.E.; Pitts, W.J.; Earl, R.A.; Hobbs, F.; et al. Identification of a Novel Inhibitor of Mitogen-activated Protein Kinase Kinase. J. Biol. Chem. 1998, 273, 18623–18632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebolt-Leopold, J.S.; Dudley, D.T.; Herrera, R.; Becelaere, K.V.; Wiland, A.; Gowan, R.C.; Tecle, H.; Barrett, S.D.; Bridges, A.; Przybranowski, S.; et al. Blockade of the MAP kinase pathway suppresses growth of colon tumors in vivo. Nat. Med. 1999, 5, 810. [Google Scholar] [CrossRef]
- Marampon, F.; Gravina, G.L.; Di Rocco, A.; Bonfili, P.; Di Staso, M.; Fardella, C.; Polidoro, L.; Ciccarelli, C.; Festuccia, C.; Popov, V.M.; et al. MEK/ERK inhibitor U0126 increases the radiosensitivity of rhabdomyosarcoma cells in vitro and in vivo by downregulating growth and DNA repair signals. Mol. Cancer Ther. 2011, 10, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, N.F.D.; Silva, R.F.; Pinto, M.M.; Silva, E.B.; Tasat, D.R.; Amaral, A. Active caspase-3 expression levels as bioindicator of individual radiosensitivity. An. Acad. Bras. Cienc. 2017, 89, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Samudio, I.; Konopleva, M.; Hail, N., Jr.; Shi, Y.-X.; McQueen, T.; Hsu, T.; Evans, R.; Honda, T.; Gribble, G.W.; Sporn, M.; et al. 2-Cyano-3,12-dioxooleana-1,9-dien-28-imidazolide (CDDO-Im) Directly Targets Mitochondrial Glutathione to Induce Apoptosis in Pancreatic Cancer. J. Biol. Chem. 2005, 280, 36273–36282. [Google Scholar] [CrossRef] [Green Version]
- Christofferson, D.E.; Yuan, J. Cyclophilin A release as a biomarker of necrotic cell death. Cell Death Differ. 2010, 17, 1942–1943. [Google Scholar] [CrossRef]
- Shepard, D.M.; Ferris, M.C.; Olivera, G.H.; Mackie, T.R. Optimizing the Delivery of Radiation Therapy to Cancer Patients. SIAM Rev. 1999, 41, 721–744. [Google Scholar] [CrossRef] [Green Version]
- Cannan, W.J.; Pederson, D.S. Mechanisms and Consequences of Double-Strand DNA Break Formation in Chromatin. J.Cell. Physiol. 2016, 231, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Schulz, A.; Meyer, F.; Dubrovska, A. Cancer Stem Cells and Radioresistance: DNA Repair and Beyond. Cancers 2019, 11, 862. [Google Scholar] [CrossRef] [Green Version]
- Gray, M.; Turnbull, A.K.; Ward, C.; Meehan, J.; Martínez-Pérez, C.; Bonello, M.; Pang, L.Y.; Langdon, S.P.; Kunkler, I.H.; Murray, A.; et al. Development and characterisation of acquired radioresistant breast cancer cell lines. Radiat. Oncol. 2019, 14, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ediriweera, M.K.; Moon, J.Y.; Nguyen, Y.T.; Cho, S.K. 10-Gingerol Targets Lipid Rafts Associated PI3K/Akt Signaling in Radio-Resistant Triple Negative Breast Cancer Cells. Molecules 2020, 25, 3164. [Google Scholar] [CrossRef]
- Jin, H.; Rugira, T.; Ko, Y.S.; Park, S.W. ESM-1 Overexpression is Involved in Increased Tumorigenesis of Radiotherapy-Resistant Breast Cancer Cells. Cancers 2020, 12, 1363. [Google Scholar] [CrossRef]
- Lu, L.; Dong, J.; Wang, L.; Xia, Q.; Zhang, D.; Kim, H.; Yin, T.; Fan, S.; Shen, Q. Activation of STAT3 and Bcl-2 and reduction of reactive oxygen species (ROS) promote radioresistance in breast cancer and overcome of radioresistance with niclosamide. Oncogene 2018, 37, 5292–5304. [Google Scholar] [CrossRef]
- Bravatà, V.; Cammarata, F.P.; Minafra, L.; Musso, R.; Pucci, G.; Spada, M.; Fazio, I.; Russo, G.; Forte, G.I. Gene Expression Profiles Induced by High-dose Ionizing Radiation in MDA-MB-231 Triple-negative Breast Cancer Cell Line. Cancer Genom. Proteom. 2019, 16, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.J.; Baek, G.Y.; Park, H.R.; Jo, S.K.; Jung, U. Smad2/3-Regulated Expression of DLX2 Is Associated with Radiation-Induced Epithelial-Mesenchymal Transition and Radioresistance of A549 and MDA-MB-231 Human Cancer Cell Lines. PLoS ONE 2016, 11, e0147343. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Gao, Y.; Zhang, X.; Wang, J.; Ding, D.; Zhang, Y.; Zhang, J.; Chen, H. Niclosamide sensitizes triple-negative breast cancer cells to ionizing radiation in association with the inhibition of Wnt/β-catenin signaling. Oncotarget 2016, 7, 42126–42138. [Google Scholar] [CrossRef]
- Lin, F.; Luo, J.; Gao, W.; Wu, J.; Shao, Z.; Wang, Z.; Meng, J.; Ou, Z.; Yang, G. COX-2 promotes breast cancer cell radioresistance via p38/MAPK-mediated cellular anti-apoptosis and invasiveness. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2013, 34, 2817–2826. [Google Scholar] [CrossRef] [PubMed]
- Koh, S.Y.; Moon, J.Y.; Unno, T. Baicalein Suppresses Stem Cell-Like Characteristics in Radio- and Chemoresistant MDA-MB-231 Human Breast Cancer Cells through Up-Regulation of IFIT2. Nutrients 2019, 11, 624. [Google Scholar] [CrossRef] [Green Version]
- Lammering, G.; Valerie, K.; Lin, P.S.; Hewit, T.H.; Schmidt-Ullrich, R.K. Radiation-induced activation of a common variant of EGFR confers enhanced radioresistance. Radiother. Oncol. J. Eur. Soc. Ther. Radiol. Oncol. 2004, 72, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Jung, S.Y.; Ahn, J.; Hwang, S.G.; Woo, H.J.; An, S.; Nam, S.Y.; Lim, D.S.; Song, J.Y. Quantitative proteomic analysis of single or fractionated radiation-induced proteins in human breast cancer MDA-MB-231 cells. Cell Biosci. 2015, 5, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, W.; Fan, M.; Huang, M.; Li, J.J.; Wang, Y. Targeted Profiling of Heat Shock Proteome in Radioresistant Breast Cancer Cells. Chem. Res. Toxicol. 2019, 32, 326–332. [Google Scholar] [CrossRef]
- Lee, Y.C.; Wang, W.L.; Chang, W.C.; Huang, Y.H.; Hong, G.C.; Wang, H.L.; Chou, Y.H. Tribbles Homolog 3 Involved in Radiation Response of Triple Negative Breast Cancer Cells by Regulating Notch1 Activation. Cancers 2019, 11, 127. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Li, L.; Zhu, H.; Chen, H.; Wei, N.; Dai, M.; Ni, Q.; Guo, X. Association between breast cancer cell migration and radiosensitivity in vitro. Oncol. Lett. 2019, 18, 6877–6884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Kuai, F.; Chen, Z.; Fu, D.; Liu, J.; Wu, Y.; Zhong, J. miR-634 Decreases the Radioresistance of Human Breast Cancer Cells by Targeting STAT3. Cancer Biother. Radiopharm. 2020, 35, 241–248. [Google Scholar] [CrossRef]
- Li, J.Y.; Li, Y.Y.; Jin, W.; Yang, Q.; Shao, Z.M.; Tian, X.S. ABT-737 reverses the acquired radioresistance of breast cancer cells by targeting Bcl-2 and Bcl-xL. J. Exp. Clin. Cancer Res. 2012, 31, 102. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, Y.T.; Moon, J.Y.; Ediriweera, M.K.; Cho, S.K. Phenethyl Isothiocyanate Suppresses Stemness in the Chemo- and Radio-Resistant Triple-Negative Breast Cancer Cell Line MDA-MB-231/IR Via Downregulation of Metadherin. Cancers 2020, 12, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oommen, D.; Prise, K.M. KNK437, abrogates hypoxia-induced radioresistance by dual targeting of the AKT and HIF-1α survival pathways. Biochem. Biophys. Res. Commun. 2012, 421, 538–543. [Google Scholar] [CrossRef]
- Kuger, S.; Cörek, E.; Polat, B.; Kämmerer, U.; Flentje, M.; Djuzenova, C.S. Novel PI3K and mTOR Inhibitor NVP-BEZ235 Radiosensitizes Breast Cancer Cell Lines under Normoxic and Hypoxic Conditions. Breast Cancer Basic Clin. Res. 2014, 8, 39–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holler, M.; Grottke, A.; Mueck, K.; Manes, J.; Jücker, M.; Rodemann, H.P.; Toulany, M. Dual Targeting of Akt and mTORC1 Impairs Repair of DNA Double-Strand Breaks and Increases Radiation Sensitivity of Human Tumor Cells. PLoS ONE 2016, 11, e0154745. [Google Scholar] [CrossRef]
- Chen, X.; Ma, N.; Zhou, Z.; Wang, Z.; Hu, Q.; Luo, J.; Mei, X.; Yang, Z.; Zhang, L.; Wang, X.; et al. Estrogen Receptor Mediates the Radiosensitivity of Triple-Negative Breast Cancer Cells. Med Sci. Monit. Int. Med J. Exp. Clin. Res. 2017, 23, 2674–2683. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Han, D.; Zhang, T.; Cheng, G.; Lu, Y.; Wang, J.; Zhao, H.; Zhao, Z. Hypoxia-induced autophagy promotes EGFR loss in specific cell contexts, which leads to cell death and enhanced radiosensitivity. Int. J. Biochem. Cell. Biol. 2019, 111, 12–18. [Google Scholar] [CrossRef]
- Arnold, K.M.; Opdenaker, L.M.; Flynn, N.J.; Appeah, D.K.; Sims-Mourtada, J. Radiation induces an inflammatory response that results in STAT3-dependent changes in cellular plasticity and radioresistance of breast cancer stem-like cells. Int. J. Radiat. Biol. 2020, 96, 434–447. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Choi, J.H.; Nam, J.S. Targeting Cancer Stem Cells in Triple-Negative Breast Cancer. Cancers 2019, 11, 965. [Google Scholar] [CrossRef] [Green Version]
- Ko, Y.S.; Jung, E.J.; Go, S.-I.; Jeong, B.K.; Kim, G.S.; Jung, J.-M.; Hong, S.C.; Kim, C.W.; Kim, H.J.; Lee, W.S. Polyphenols Extracted from Artemisia annua L. Exhibit Anti-Cancer Effects on Radio-Resistant MDA-MB-231 Human Breast Cancer Cells by Suppressing Stem Cell Phenotype, β-Catenin, and MMP-9. Molecules 2020, 25, 1916. [Google Scholar] [CrossRef]
- Qin, J.; Xin, H.; Nickoloff, B.J. Specifically targeting ERK1 or ERK2 kills melanoma cells. J. Transl. Med. 2012, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Ciccarelli, C.; Di Rocco, A.; Gravina, G.L.; Mauro, A.; Festuccia, C.; Del Fattore, A.; Berardinelli, P.; De Felice, F.; Musio, D.; Bouché, M.; et al. Disruption of MEK/ERK/c-Myc signaling radiosensitizes prostate cancer cells in vitro and in vivo. J. Cancer Res. Clin. Oncol. 2018, 144, 1685–1699. [Google Scholar] [CrossRef] [PubMed]
- Matsui, W.H. Cancer stem cell signaling pathways. Medicine 2016, 95, S8–S19. [Google Scholar] [CrossRef]
- Tuasha, N.; Petros, B. Heterogeneity of Tumors in Breast Cancer: Implications and Prospects for Prognosis and Therapeutics. Scientifica 2020, 2020, 4736091. [Google Scholar] [CrossRef] [PubMed]
- Moncharmont, C.; Levy, A.; Gilormini, M.; Bertrand, G.; Chargari, C.; Alphonse, G.; Ardail, D.; Rodriguez-Lafrasse, C.; Magné, N. Targeting a cornerstone of radiation resistance: Cancer stem cell. Cancer Lett. 2012, 322, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.H.; Zhou, L.; Ng, K.Y.; Wong, T.L.; Lee, T.K.; Sharma, R.; Loong, J.H.; Ching, Y.P.; Yuan, Y.-F.; Xie, D.; et al. PRMT6 Regulates RAS/RAF Binding and MEK/ERK-Mediated Cancer Stemness Activities in Hepatocellular Carcinoma through CRAF Methylation. Cell Rep. 2018, 25, 690–701. [Google Scholar] [CrossRef] [Green Version]
- Skvortsov, S.; Debbage, P.; Lukas, P.; Skvortsova, I. Crosstalk between DNA repair and cancer stem cell (CSC) associated intracellular pathways. Semin. Cancer Biol. 2015, 31, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Chai, X.; Chu, H.; Yang, X.; Meng, Y.; Shi, P.; Gou, S. Metformin Increases Sensitivity of Pancreatic Cancer Cells to Gemcitabine by Reducing CD133+ Cell Populations and Suppressing ERK/P70S6K Signaling. Sci. Rep. 2015, 5, 14404. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.-J.; Wu, D.-W.; Wang, Y.-C.; Chen, C.-Y.; Lee, H. PAK1 confers chemoresistance and poor outcome in non-small cell lung cancer via β-catenin-mediated stemness. Sci. Rep. 2016, 6, 34933. [Google Scholar] [CrossRef]
- Tung, S.L.; Huang, W.C.; Hsu, F.C.; Yang, Z.P.; Jang, T.H.; Chang, J.W.; Chuang, C.M.; Lai, C.R.; Wang, L.H. miRNA-34c-5p inhibits amphiregulin-induced ovarian cancer stemness and drug resistance via downregulation of the AREG-EGFR-ERK pathway. Oncogenesis 2017, 6, e326. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.; Yang, Z.; Xie, L.; DeWitt, J.P.; Chen, Y. Cancer therapy in the necroptosis era. Cell Death Differ. 2016, 23, 748–756. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, A.P.; Silva, S.N.; Reichert, A.; Lima, F.; Júnior, E.; Rueff, J. Effects of polymorphic DNA genes involved in BER and caspase pathways on the clinical outcome of myeloproliferative neoplasms under treatment with hydroxyurea. Mol. Med. Rep. 2018, 18, 5243–5255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.Y.; Xuan, Y.; Jin, X.Y.; Tian, X.; Wu, R. CASP-9 gene functional polymorphisms and cancer risk: A large-scale association study plus meta-analysis. Genet. Mol. Res. GMR 2013, 12, 3070–3078. [Google Scholar] [CrossRef]
- Araki, S.; Dairiki, R.; Nakayama, Y.; Murai, A.; Miyashita, R.; Iwatani, M.; Nomura, T.; Nakanishi, O. Inhibitors of CLK protein kinases suppress cell growth and induce apoptosis by modulating pre-mRNA splicing. PLoS ONE 2015, 10, e0116929. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Kim, J.H.; Carver, K.; Su, Y.; Weremowicz, S.; Mulvey, L.; Yamamoto, S.; Brennan, C.; Mei, S.; Long, H.; et al. CLK2 Is an Oncogenic Kinase and Splicing Regulator in Breast Cancer. Cancer Res. 2015, 75, 1516–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, M. FGFR inhibitors: Effects on cancer cells, tumor microenvironment and whole-body homeostasis (Review). Int. J. Mol. Med. 2016, 38, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmanian, N.; Hosseinimehr, S.J.; Khalaj, A. The paradox role of caspase cascade in ionizing radiation therapy. J. Biomed. Sci. 2016, 23, 88. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.K.; Park, J.I. MEK1/2 Inhibitors: Molecular Activity and Resistance Mechanisms. Semin. Oncol. 2015, 42, 849–862. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathways | Genes | Genes Involved in the Pathways (%) | KEGG Pathway Entry |
|---|---|---|---|
| MAPK signaling pathway | MAPK1, CASP3, FGF22, MAPK11, IL1A | 18 | map04010 |
| NOD-like receptor signaling pathway | MAPK1, HSP90AA1, MAPK11 | 15 | map04621 |
| PI3K-Akt signaling pathway | MAPK1, EIF4EBP1, HSP90AA1, FGF22, F2R | 15 | map04151 |
| Pathways in cancer | MAPK1, CASP3, HSP90AA1, FGF22, F2R | 13 | map05200 |
| Rap1 signaling pathway | MAPK1, FGF22, MAPK11, F2R | 13 | map04015 |
| TNF signaling pathway | MAPK1, CASP3, MAPK11 | 11 | map04668 |
| Proteoglycans in cancer | MAPK1, CASP3, MAPK11 | 8 | map05205 |
| Regulation of actin cytoskeleton | MAPK1, FGF22, F2R | 7 | map04810 |
| No. | Author | Key Findings | Year | References |
|---|---|---|---|---|
| (i) Specific signaling pathways involved in radio-resistant breast cancer cells | ||||
| 1. | Gray et al. | The radiation-resistant ER+ breast cancer cell line (MCF-7, ZR-751) showed increased migration and invasion compared to the radiation-resistant ER- breast cancer cell line (MDA-MB-231). ER+ cells also showed a shift from ER to EGFR signaling pathways with increased MAPK and PI3K activity. | 2019 | [26] |
| 2. | Ediriweera et al. | A phenolic lipid, 10-Gingerol, promotes apoptosis in radiation-resistant MDA-MB-231 cells through the PI3K/Akt signaling pathway. | 2020 | [27] |
| 3. | Jin et al. | The overexpression of ESM-1 plays a critical role in radiation-resistant MDA-MB-231 cells through the regulation of PDK, PKC, and ERK1/2 pathways, and the subsequent activation of transcription factors HIF-1α, NF-κB, and STAT-3 to regulate adhesion molecules, MMPs, and VEGF. | 2020 | [28] |
| 4. | lu et al. | The Wnt/β-catenin signaling pathway plays an important role in the development of radioresistance and Niclosamide, an FDA-approved anthelmintic drug that induces radiosensitivity in radiation-resistant MDA-MB-231 via inhibiting STAT3 and Bcl-2. | 2018 | [29] |
| 5. | Bravatà et al. | Gene expression profiles of the MDA-MB-231 radiation cell fraction show increased TNF signaling, Phagosome, NF-kappa B signaling, Jak-STAT signaling, and PI3K-Akt signaling. | 2019 | [30] |
| 6. | Choi et al. | DLX2 expression with irradiation incidence causes the increase in the EMT process and CSCs population through the Smad2/3 signaling pathway in MDA-MB-231 and A549 cells. | 2016 | [31] |
| (ii) Signaling pathways involved in radio sensitivity of MDA-MB-231 cells | ||||
| 7. | Yin et al. | Niclosamide, an antihelminthic drug, inhibited the Wnt/β-catenin signaling pathway and increased the radiation sensitivity to triple-negative breast cancer (TNBC) cells (MDA-MB-231, MDA-MB-468, and Hs578T cells). | 2016 | [32] |
| 8. | Lin et al. | COX-2 upregulation promotes radioresistance in MDA-MB-231 cells through the p38/MAPK-mediated alteration of apoptosis and metastasis. | 2013 | [33] |
| 9. | KO et al. | Radiation-resistant MDA-MB-231 cells showed an increased cell proliferation, cell adhesion, EMT process, and increased stem cell population. | 2018 | [8] |
| 10. | Koh et al. | Baicalein reduced the stem cell-like properties and metastasis in radiation-resistant MDA-MB-231 cells through the upregulation of IFIT2. | 2019 | [34] |
| (iii) Altered expression of gene/proteins involved in radio-resistant MDA-MB-231 cells. | ||||
| 11. | Lammering et al. | Irradiation increased the expression of the EGFR protein in MDA-MB-231 xenograft tumors. | 2004 | [35] |
| 12 | Kim et al. | Proteomic analysis revealed single and the fraction of radiation increased cathepsin D (CTSD), gelsolin (GSN), argininosuccinate synthase 1 (ASS1), and C-type mannose receptor 2 (MRC2) in MDA-MB-231 cells. | 2015 | [36] |
| 13. | Miao et al. | Radiation-resistant MDA-MB-231 and MCF-7 cells showed an altered expression of several members of the HSP70 and HSP40, subfamilies of HSPs, and an increased level of HSPB8, a target of NF-κB that could be responsible for the development of radioresistance. | 2019 | [37] |
| 14. | Lee et al. | Increased expression of TRIB3 in radiation-resistant MDA-MB-231 cells causes the resistance and knockdown of TRIB3 sensitized toward radiation. | 2019 | [38] |
| 15. | HOU et al. | Microarray analysis of radiation-resistant MDA-MB-231 cells showed increased cell adhesion and EMT factors. | 2019 | [39] |
| 16. | Yang et al. | Overexpression of a small RNA molecule miR-634 decreases the survival rate of radiation-resistant MCF-7 and MDA-MB-231 cells by direct interaction with STAT3. | 2020 | [40] |
| (iv) Studies that did not fall into any of the three above categories. | ||||
| 17. | Li et al. | A small molecule, ABT-787, induces radiosensitivity in radiation-resistant MDA-MB-231 by targeting Bcl-2 and Bcl-xL. | 2012 | [41] |
| 18. | Nguyen et al. | A phytochemical phenethyl isothiocyanate reduces the CSC population in radiation-resistant MDA-MB-231 cells through upregulating ROS levels and targeting Metadherin at the post-transcriptional levels. | 2020 | [42] |
| 19. | Oommen and Prise | A novel benzylidene lactam compound, KNK437, inhibits HIF-1α, HSF1, and AKT in hypoxia-induced MDA-MB-231 and T98G cells, which, in turn, induces radiosensitivity. | 2012 | [43] |
| 20. | Kuger et al. | PI3K/mTOR inhibitor NVP-BEZ235 showed a synergistic effect with irradiation (IR) in MCF-7 and MDA-MB-231 cells. | 2014 | [44] |
| 21. | Holler et al. | The molecular targeting of Akt by Akt inhibitor MK2206 or the knockdown of Akt1 led to a rapamycin-induced radiosensitization of SK-MES-1, HTB-182, or MDA-MB-231 cells by increasing DNA-double-stranded breaks. | 2016 | [45] |
| 22. | Chen et al. | The estrogen receptor mediates the radiosensitivity of TNBC cells. | 2017 | [46] |
| 23. | Liu et al. | Hypoxia due to a high cell density downregulated the EGFR expression and increased the sensitivity to ionizing radiation in MCF-7 and MDA-MB-231 cells. | 2018 | [47] |
| 24. | Arnold et al. | STAT3 inhibition combined with radiation reduces the cellular plasticity in MDA-MB-231 and SUM159PT cells. | 2019 | [48] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paramanantham, A.; Jung, E.J.; Go, S.-I.; Jeong, B.K.; Jung, J.-M.; Hong, S.C.; Kim, G.S.; Lee, W.S. Activated ERK Signaling Is One of the Major Hub Signals Related to the Acquisition of Radiotherapy-Resistant MDA-MB-231 Breast Cancer Cells. Int. J. Mol. Sci. 2021, 22, 4940. https://doi.org/10.3390/ijms22094940
Paramanantham A, Jung EJ, Go S-I, Jeong BK, Jung J-M, Hong SC, Kim GS, Lee WS. Activated ERK Signaling Is One of the Major Hub Signals Related to the Acquisition of Radiotherapy-Resistant MDA-MB-231 Breast Cancer Cells. International Journal of Molecular Sciences. 2021; 22(9):4940. https://doi.org/10.3390/ijms22094940
Chicago/Turabian StyleParamanantham, Anjugam, Eun Joo Jung, Se-IL Go, Bae Kwon Jeong, Jin-Myung Jung, Soon Chan Hong, Gon Sup Kim, and Won Sup Lee. 2021. "Activated ERK Signaling Is One of the Major Hub Signals Related to the Acquisition of Radiotherapy-Resistant MDA-MB-231 Breast Cancer Cells" International Journal of Molecular Sciences 22, no. 9: 4940. https://doi.org/10.3390/ijms22094940
APA StyleParamanantham, A., Jung, E. J., Go, S.-I., Jeong, B. K., Jung, J.-M., Hong, S. C., Kim, G. S., & Lee, W. S. (2021). Activated ERK Signaling Is One of the Major Hub Signals Related to the Acquisition of Radiotherapy-Resistant MDA-MB-231 Breast Cancer Cells. International Journal of Molecular Sciences, 22(9), 4940. https://doi.org/10.3390/ijms22094940