
The Multifaceted Roles of USP15 in Signal Transduction


Abstract
1. Introduction
2. USP15 Gene and Its Paralogs
3. USP15 Regulates Signal Transduction Pathways
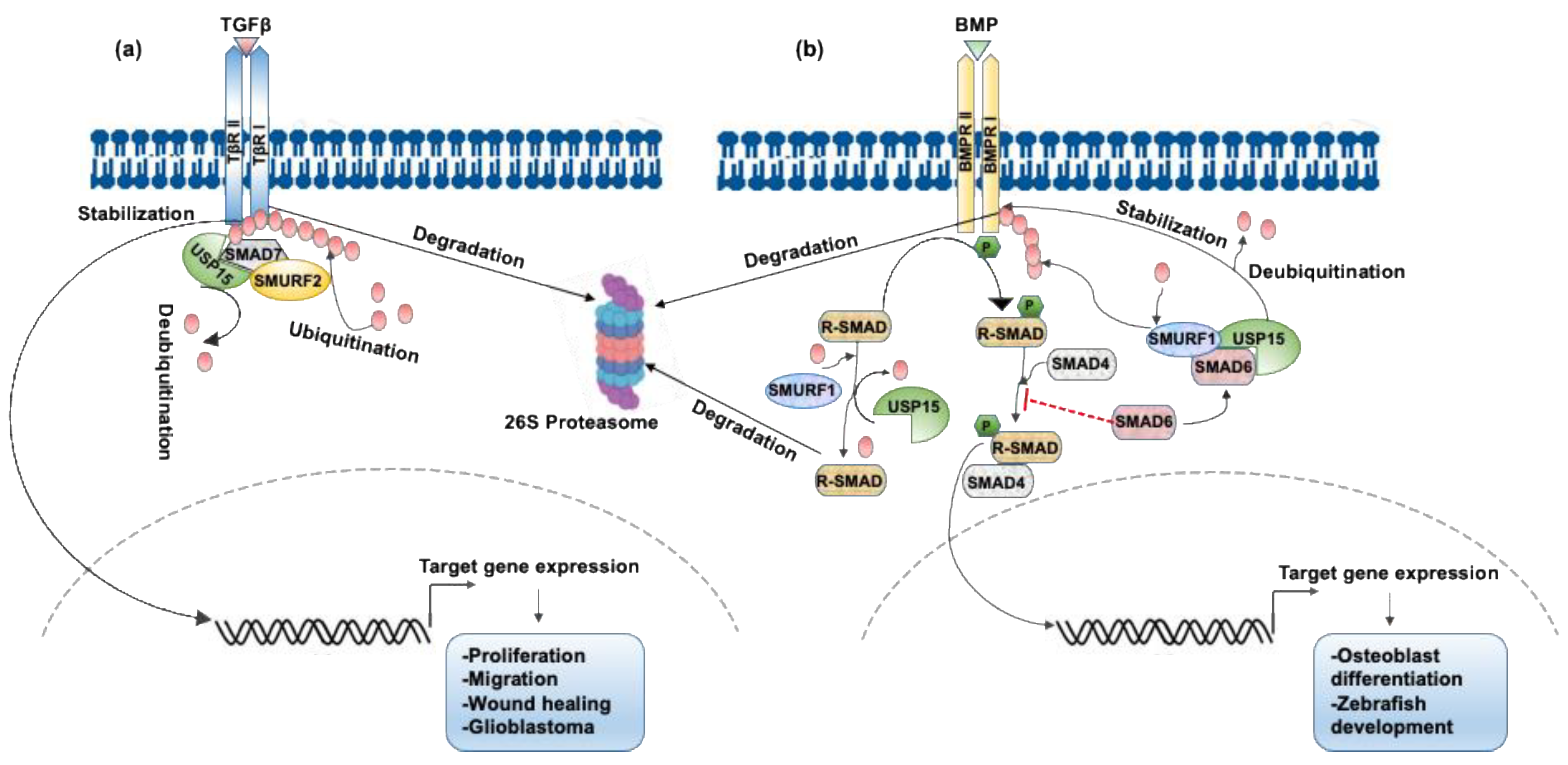
3.1. USP15 Enhances TGF-β Signaling by Deubiquitinating the Type I TGFβ Receptor ALK5
3.2. USP15 Deubiquitinates Type I BMP Receptor ALK3 and Enhances BMP Signaling
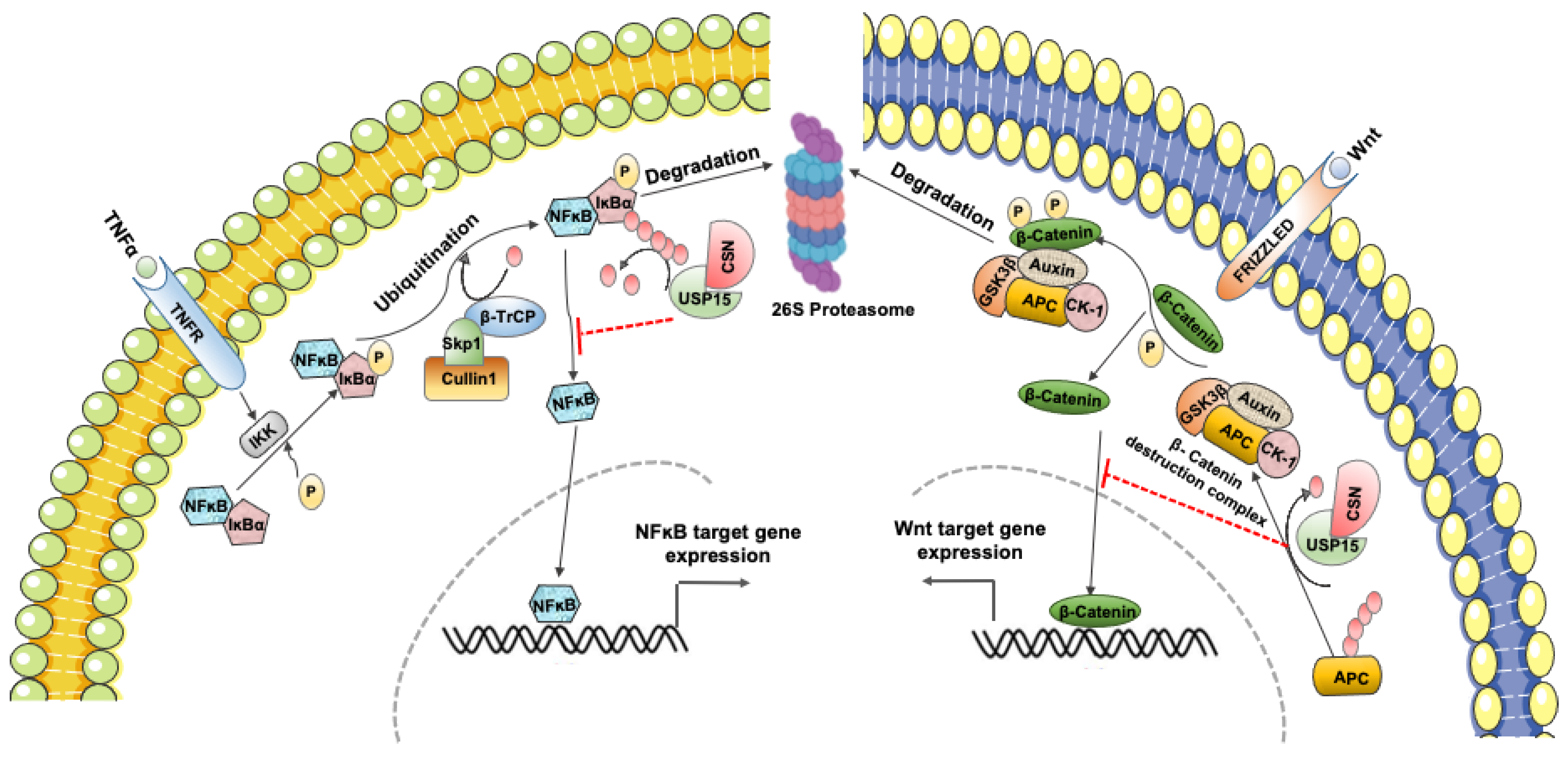
3.3. USP15 Regulates the Activity of the COP9 Signalosome toward CRLs
3.4. USP15 Can Either Suppress or Activate NF-κB Signaling
3.5. USP15 Shows Both Positive and Negative Effects on Wnt/β-Catenin Signaling
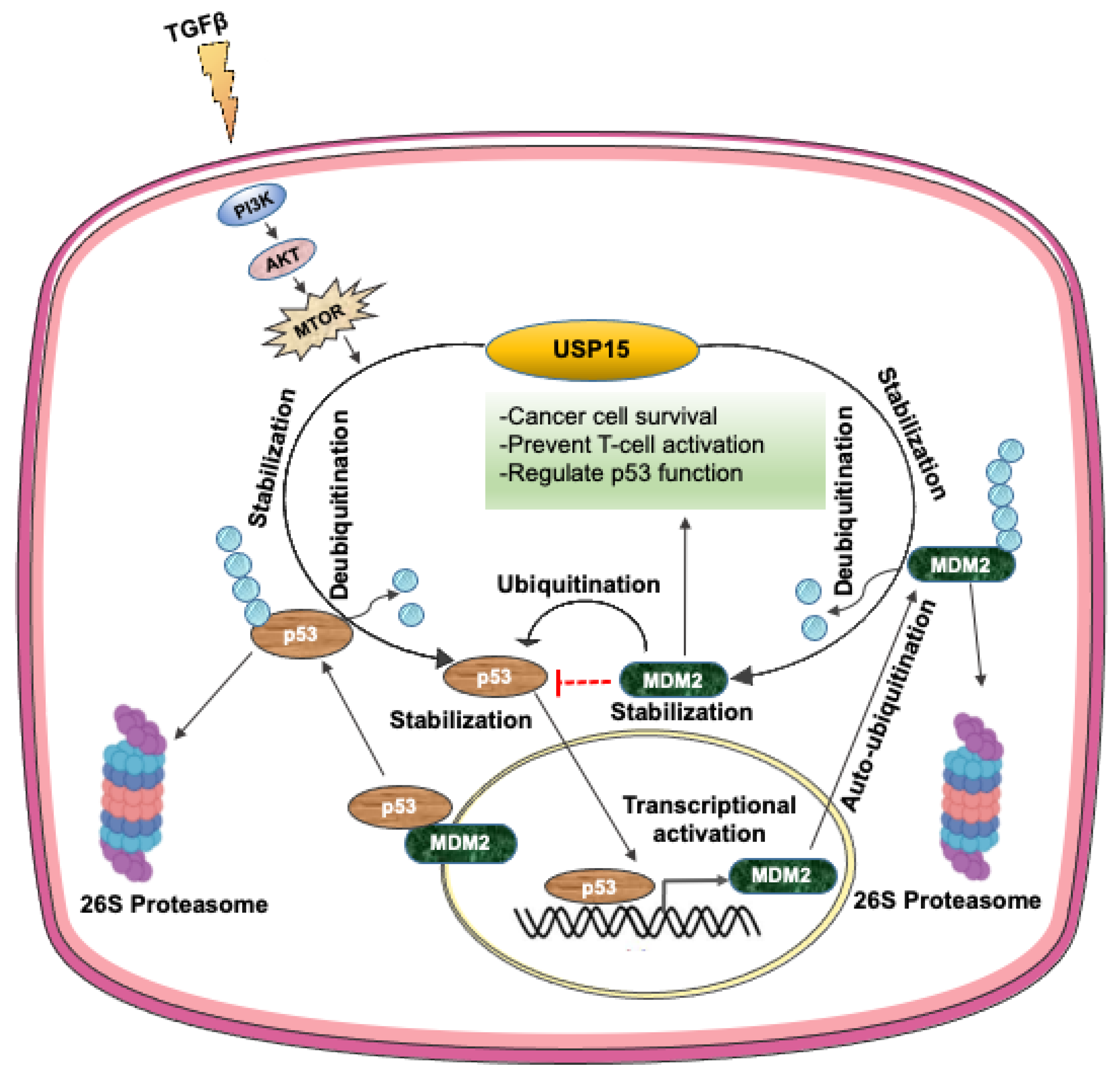
3.6. USP15 Regulates p53 Signaling by Controlling the Stability of MDM2 and p53
3.7. USP15 Attenuates IGF Signaling in Prostate Cancer
3.8. USP15 Promotes RIG-I-Mediated Antiviral Innate Immune Signaling
3.9. USP15 Downregulates Nrf2 Signaling and Target Gene Expression
4. Coupling of Signal Transduction to Pre-mRNA Splicing Regulated by USP15
5. Misregulation of USP15 in Diseases
6. Perspective
Author Contributions
Funding
Conflicts of Interest
References
- Hershko, A.; Ciechanover, A.; Varshavsky, A. Basic medical research award. The ubiquitin system. Nat. Med. 2000, 6, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Glickman, M.H.; Ciechanover, A. The ubiquitin-proteasome proteolytic pathway: Destruction for the sake of construction. Physiol. Rev. 2002, 82, 373–428. [Google Scholar] [CrossRef] [PubMed]
- Schnell, J.D.; Hicke, L. Non-traditional functions of ubiquitin and ubiquitin-binding proteins. J. Biol. Chem. 2003, 278, 35857–35860. [Google Scholar] [CrossRef]
- D’Andrea, A.; Pellman, D. Deubiquitinating enzymes: A new class of biological regulators. Crit. Rev. Biochem. Mol. Biol. 1998, 33, 337–352. [Google Scholar] [CrossRef]
- Reyes-Turcu, F.E.; Ventii, K.H.; Wilkinson, K.D. Regulation and cellular roles of ubiquitin-specific deubiquitinating enzymes. Annu. Rev. Biochem. 2009, 78, 363–397. [Google Scholar] [CrossRef]
- Clague, M.J.; Barsukov, I.; Coulson, J.M.; Liu, H.; Rigden, D.J.; Urbe, S. Deubiquitylases from genes to organism. Physiol. Rev. 2013, 93, 1289–1315. [Google Scholar] [CrossRef]
- Mevissen, T.E.T.; Komander, D. Mechanisms of deubiquitinase specificity and regulation. Annu. Rev. Biochem. 2017, 86, 159–192. [Google Scholar] [CrossRef]
- Clague, M.J.; Urbe, S.; Komander, D. Breaking the chains: Deubiquitylating enzyme specificity begets function. Nat. Rev. Mol. Cell Biol. 2019, 20, 338–352. [Google Scholar] [CrossRef]
- Grabbe, C.; Husnjak, K.; Dikic, I. The spatial and temporal organization of ubiquitin networks. Nat. Rev. Mol. Cell Biol. 2011, 12, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Pfoh, R.; Lacdao, I.K.; Saridakis, V. Deubiquitinases and the new therapeutic opportunities offered to cancer. Endocr. Relat. Cancer 2015, 22, T35–T54. [Google Scholar] [CrossRef] [PubMed]
- Komander, D.; Clague, M.J.; Urbe, S. Breaking the chains: Structure and function of the deubiquitinases. Nat. Rev. Mol. Cell Biol. 2009, 10, 550–563. [Google Scholar] [CrossRef]
- Fielding, A.B.; Concannon, M.; Darling, S.; Rusilowicz-Jones, E.V.; Sacco, J.J.; Prior, I.A.; Clague, M.J.; Urbe, S.; Coulson, J.M. The deubiquitylase USP15 regulates topoisomerase II alpha to maintain genome integrity. Oncogene 2018, 37, 2326–2342. [Google Scholar] [CrossRef]
- Eichhorn, P.J.; Rodon, L.; Gonzalez-Junca, A.; Dirac, A.; Gili, M.; Martinez-Saez, E.; Aura, C.; Barba, I.; Peg, V.; Prat, A.; et al. USP15 stabilizes TGF-beta receptor I and promotes oncogenesis through the activation of TGF-beta signaling in glioblastoma. Nat. Med. 2012, 18, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Srihari, S.; Ragan, M.A. Systematic tracking of dysregulated modules identifies novel genes in cancer. Bioinformatics 2013, 29, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Besche, H.C.; Sha, Z.; Kukushkin, N.V.; Peth, A.; Hock, E.M.; Kim, W.; Gygi, S.; Gutierrez, J.A.; Liao, H.; Dick, L.; et al. Autoubiquitination of the 26S proteasome on Rpn13 regulates breakdown of ubiquitin conjugates. EMBO J. 2014, 33, 1159–1176. [Google Scholar] [CrossRef] [PubMed]
- Park, K.C.; Choi, E.J.; Min, S.W.; Chung, S.S.; Kim, H.; Suzuki, T.; Tanaka, K.; Chung, C.H. Tissue-specificity, functional characterization and subcellular localization of a rat ubiquitin-specific processing protease, UBP109, whose mRNA expression is developmentally regulated. Biochem. J. 2000, 349, 443–453. [Google Scholar] [CrossRef]
- Kotani, Y.; Morito, D.; Sakata, K.; Ainuki, S.; Sugihara, M.; Hatta, T.; Iemura, S.I.; Takashima, S.; Natsume, T.; Nagata, K. Alternative exon skipping biases substrate preference of the deubiquitylase USP15 for mysterin/RNF213, the moyamoya disease susceptibility factor. Sci. Rep. 2017, 7, 44293. [Google Scholar] [CrossRef]
- Ward, S.J.; Gratton, H.E.; Indrayudha, P.; Michavila, C.; Mukhopadhyay, R.; Maurer, S.K.; Caulton, S.G.; Emsley, J.; Dreveny, I. The structure of the deubiquitinase USP15 reveals a misaligned catalytic triad and an open ubiquitin-binding channel. J. Biol. Chem. 2018, 293, 17362–17374. [Google Scholar] [CrossRef] [PubMed]
- Harper, S.; Besong, T.M.; Emsley, J.; Scott, D.J.; Dreveny, I. Structure of the USP15 N-terminal domains: A beta-hairpin mediates close association between the DUSP and UBL domains. Biochemistry 2011, 50, 7995–8004. [Google Scholar] [CrossRef]
- Elliott, P.R.; Liu, H.; Pastok, M.W.; Grossmann, G.J.; Rigden, D.J.; Clague, M.J.; Urbe, S.; Barsukov, I.L. Structural variability of the ubiquitin specific protease DUSP-UBL double domains. FEBS Lett. 2011, 585, 3385–3390. [Google Scholar] [CrossRef] [PubMed]
- Harper, S.; Gratton, H.E.; Cornaciu, I.; Oberer, M.; Scott, D.J.; Emsley, J.; Dreveny, I. Structure and catalytic regulatory function of ubiquitin specific protease 11 N-terminal and ubiquitin-like domains. Biochemistry 2014, 53, 2966–2978. [Google Scholar] [CrossRef] [PubMed]
- Park, J.K.; Das, T.; Song, E.J.; Kim, E.E. Structural basis for recruiting and shuttling of the spliceosomal deubiquitinase USP4 by SART3. Nucleic Acids Res. 2016, 44, 5424–5437. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Harding, R.; Hou, F.; Dong, A.; Walker, J.R.; Bteich, J.; Tong, Y. Structural basis of the recruitment of ubiquitin-specific protease USP15 by spliceosome recycling factor SART3. J. Biol. Chem. 2016, 291, 17283–17292. [Google Scholar] [CrossRef] [PubMed]
- Das, T.; Park, J.K.; Park, J.; Kim, E.; Rape, M.; Kim, E.E.; Song, E.J. USP15 regulates dynamic protein-protein interactions of the spliceosome through deubiquitination of PRP31. Nucleic Acids Res. 2017, 45, 5010–5011. [Google Scholar] [CrossRef][Green Version]
- Das, T.; Kim, E.E.; Song, E.J. Phosphorylation of USP15 and USP4 regulates localization and spliceosomal deubiquitination. J. Mol. Biol. 2019, 431, 3900–3912. [Google Scholar] [CrossRef] [PubMed]
- Vlasschaert, C.; Xia, X.; Coulombe, J.; Gray, D.A. Evolution of the highly networked deubiquitinating enzymes USP4, USP15, and USP11. BMC Evol. Biol. 2015, 15, 230. [Google Scholar] [CrossRef]
- Chou, C.K.; Chang, Y.T.; Korinek, M.; Chen, Y.T.; Yang, Y.T.; Leu, S.; Lin, I.L.; Tang, C.J.; Chiu, C.C. The regulations of deubiquitinase USP15 and its pathophysiological mechanisms in diseases. Int J. Mol. Sci. 2017, 18, 483. [Google Scholar] [CrossRef]
- Hu, B.; Zhang, D.; Zhao, K.; Wang, Y.; Pei, L.; Fu, Q.; Ma, X. Spotlight on USP4: Structure, function, and regulation. Front. Cell Dev. Biol. 2021, 9, 595159. [Google Scholar] [CrossRef]
- Clerici, M.; Luna-Vargas, M.P.; Faesen, A.C.; Sixma, T.K. The DUSP-Ubl domain of USP4 enhances its catalytic efficiency by promoting ubiquitin exchange. Nat. Commun. 2014, 5, 5399. [Google Scholar] [CrossRef]
- Al-Salihi, M.A.; Herhaus, L.; Macartney, T.; Sapkota, G.P. USP11 augments TGFbeta signalling by deubiquitylating ALK5. Open Biol. 2012, 2, 120063. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, F.; Drabsch, Y.; Gao, R.; Snaar-Jagalska, B.E.; Mickanin, C.; Huang, H.; Sheppard, K.A.; Porter, J.A.; Lu, C.X.; et al. USP4 is regulated by AKT phosphorylation and directly deubiquitylates TGF-beta type I receptor. Nat. Cell Biol. 2012, 14, 717–726. [Google Scholar] [CrossRef]
- Klein, W.L. Biochemistry and regulation of signal transduction by neuronal acetylcholine receptors. Curr. Top. Cell Regul. 1984, 24, 129–144. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, K.; Bozko, P.M.; Dubiel, W.; Naumann, M. CSN controls NF-kappaB by deubiquitinylation of IkappaBalpha. EMBO J. 2007, 26, 1532–1541. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q.; Jin, J.; Hu, H.; Li, H.S.; Romano, S.; Xiao, Y.; Nakaya, M.; Zhou, X.; Cheng, X.; Yang, P.; et al. USP15 stabilizes MDM2 to mediate cancer-cell survival and inhibit antitumor T cell responses. Nat. Immunol. 2014, 15, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Langelotz, C.; Hetfeld-Pechoc, B.K.; Schwenk, W.; Dubiel, W. The COP9 signalosome mediates beta-catenin degradation by deneddylation and blocks adenomatous polyposis coli destruction via USP15. J. Mol. Biol. 2009, 391, 691–702. [Google Scholar] [CrossRef]
- Torre, S.; Polyak, M.J.; Langlais, D.; Fodil, N.; Kennedy, J.M.; Radovanovic, I.; Berghout, J.; Leiva-Torres, G.A.; Krawczyk, C.M.; Ilangumaran, S.; et al. USP15 regulates type I interferon response and is required for pathogenesis of neuroinflammation. Nat. Immunol. 2017, 18, 54–63. [Google Scholar] [CrossRef]
- Yang, X.D.; Sun, S.C. Deubiquitinases as pivotal regulators of T cell functions. Front. Med. 2018, 12, 451–462. [Google Scholar] [CrossRef]
- Villeneuve, N.F.; Tian, W.; Wu, T.; Sun, Z.; Lau, A.; Chapman, E.; Fang, D.; Zhang, D.D. USP15 negatively regulates Nrf2 through deubiquitination of Keap1. Mol. Cell. 2013, 51, 68–79. [Google Scholar] [CrossRef]
- Herhaus, L.; Al-Salihi, M.A.; Dingwell, K.S.; Cummins, T.D.; Wasmus, L.; Vogt, J.; Ewan, R.; Bruce, D.; Macartney, T.; Weidlich, S.; et al. USP15 targets ALK3/BMPR1A for deubiquitylation to enhance bone morphogenetic protein signalling. Open Biol. 2014, 4, 140065. [Google Scholar] [CrossRef]
- Wrana, J.L.; Attisano, L.; Carcamo, J.; Zentella, A.; Doody, J.; Laiho, M.; Wang, X.F.; Massague, J. TGF beta signals through a heteromeric protein kinase receptor complex. Cell 1992, 71, 1003–1014. [Google Scholar] [CrossRef]
- Massague, J.; Seoane, J.; Wotton, D. Smad transcription factors. Genes Dev. 2005, 19, 2783–2810. [Google Scholar] [CrossRef] [PubMed]
- Wicks, S.J.; Grocott, T.; Haros, K.; Maillard, M.; ten Dijke, P.; Chantry, A. Reversible ubiquitination regulates the Smad/TGF-beta signalling pathway. Biochem. Soc. Trans. 2006, 34, 761–763. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Imamura, T. Regulation of TGF-beta family signaling by E3 ubiquitin ligases. Cancer Sci. 2008, 99, 2107–2112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, X.; Xie, F.; Zhang, Z.; van Dam, H.; Zhang, L.; Zhou, F. The regulation of TGF-beta/SMAD signaling by protein deubiquitination. Protein Cell. 2014, 5, 503–517. [Google Scholar] [CrossRef] [PubMed]
- Iyengar, P.V.; Jaynes, P.; Rodon, L.; Lama, D.; Law, K.P.; Lim, Y.P.; Verma, C.; Seoane, J.; Eichhorn, P.J. USP15 regulates SMURF2 kinetics through C-lobe mediated deubiquitination. Sci. Rep. 2015, 5, 14733. [Google Scholar] [CrossRef]
- Feng, A.P.; He, Y.M.; Liu, X.X.; Li, J.W.; Tu, Y.T.; Hu, F.; Chen, S.J. Expression of USP15, TbetaR-I and Smad7 in psoriasis. J. Huazhong Univ. Sci. Technol. Med. Sci. 2014, 34, 415–419. [Google Scholar] [CrossRef]
- Akhurst, R.J.; Derynck, R. TGF-beta signaling in cancer—A double-edged sword. Trends Cell Biol. 2001, 11, S44–S51. [Google Scholar] [CrossRef]
- Inui, M.; Manfrin, A.; Mamidi, A.; Martello, G.; Morsut, L.; Soligo, S.; Enzo, E.; Moro, S.; Polo, S.; Dupont, S.; et al. USP15 is a deubiquitylating enzyme for receptor-activated SMADs. Nat. Cell Biol. 2011, 13, 1368–1375. [Google Scholar] [CrossRef]
- Galant, C.; Marchandise, J.; Stoenoiu, M.S.; Ducreux, J.; de Groof, A.; Pirenne, S.; van den Eynde, B.; Houssiau, F.A.; Lauwerys, B.R. Overexpression of ubiquitin-specific peptidase 15 in systemic sclerosis fibroblasts increases response to transforming growth factor beta. Rheumatology 2019, 58, 708–718. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Z.; Ho, C.; Zhang, G.; Li, Q. Ubiquitin-specific protease 15 maintains transforming growth factor-beta pathway activity by deubiquitinating transforming growth factor-beta receptor i during wound healing. Am. J. Pathol. 2019, 189, 1351–1362. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, H.; Zhou, F.; Schimmel, J.; Pardo, C.G.; Zhang, T.; Barakat, T.S.; Sheppard, K.A.; Mickanin, C.; Porter, J.A.; et al. RNF12 controls embryonic stem cell fate and morphogenesis in zebrafish embryos by targeting Smad7 for degradation. Mol. Cell. 2012, 46, 650–661. [Google Scholar] [CrossRef] [PubMed]
- Xiao, N.; Li, H.; Luo, J.; Wang, R.; Chen, H.; Chen, J.; Wang, P. Ubiquitin-specific protease 4 (USP4) targets TRAF2 and TRAF6 for deubiquitination and inhibits TNFalpha-induced cancer cell migration. Biochem. J. 2012, 441, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Tse, W.K.; Jiang, Y.J.; Wong, C.K. Zebrafish transforming growth factor-beta-stimulated clone 22 domain 3 (TSC22D3) plays critical roles in Bmp-dependent dorsoventral patterning via two deubiquitylating enzymes Usp15 and Otud4. Biochim. Biophys. Acta 2013, 1830, 4584–4593. [Google Scholar] [CrossRef] [PubMed]
- Tse, W.K.; Eisenhaber, B.; Ho, S.H.; Ng, Q.; Eisenhaber, F.; Jiang, Y.J. Genome-wide loss-of-function analysis of deubiquitylating enzymes for zebrafish development. BMC Genomics 2009, 10, 637. [Google Scholar] [CrossRef] [PubMed]
- Schwechheimer, C.; Deng, X.W. COP9 signalosome revisited: A novel mediator of protein degradation. Trends Cell Biol. 2001, 11, 420–426. [Google Scholar] [CrossRef]
- Lee, M.H.; Zhao, R.; Phan, L.; Yeung, S.C. Roles of COP9 signalosome in cancer. Cell Cycle 2011, 10, 3057–3066. [Google Scholar] [CrossRef]
- Li, P.; Xie, L.; Gu, Y.; Li, J.; Xie, J. Roles of multifunctional COP9 signalosome complex in cell fate and implications for drug discovery. J. Cell Physiol. 2017, 232, 1246–1253. [Google Scholar] [CrossRef]
- Zhou, C.; Wee, S.; Rhee, E.; Naumann, M.; Dubiel, W.; Wolf, D.A. Fission yeast COP9/signalosome suppresses cullin activity through recruitment of the deubiquitylating enzyme Ubp12p. Mol. Cell. 2003, 11, 927–938. [Google Scholar] [CrossRef]
- Meister, C.; Thieme, K.G.; Thieme, S.; Kohler, A.M.; Schmitt, K.; Valerius, O.; Braus, G.H. COP9 signalosome interaction with UspA/Usp15 deubiquitinase controls VeA-Mediated fungal multicellular development. Biomolecules 2019, 9, 238. [Google Scholar] [CrossRef]
- Hetfeld, B.K.; Helfrich, A.; Kapelari, B.; Scheel, H.; Hofmann, K.; Guterman, A.; Glickman, M.; Schade, R.; Kloetzel, P.M.; Dubiel, W. The zinc finger of the CSN-associated deubiquitinating enzyme USP15 is essential to rescue the E3 ligase Rbx1. Curr. Biol. 2005, 15, 1217–1221. [Google Scholar] [CrossRef]
- Schweitzer, K.; Naumann, M. CSN-associated USP48 confers stability to nuclear NF-kappaB/RelA by trimming K48-linked Ub-chains. Biochim. Biophys. Acta 2015, 1853, 453–469. [Google Scholar] [CrossRef] [PubMed]
- Dubiel, W.; Chaithongyot, S.; Dubiel, D.; Naumann, M. The COP9 signalosome: A multi-DUB complex. Biomolecules 2020, 10, 1082. [Google Scholar] [CrossRef]
- Rozen, S.; Tieri, A.; Ridner, G.; Stark, A.K.; Schmaler, T.; Ben-Nissan, G.; Dubiel, W.; Sharon, M. Exposing the subunit diversity within protein complexes: A mass spectrometry approach. Methods 2013, 59, 270–277. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-kappaB signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Ben-Neriah, Y. Phosphorylation meets ubiquitination: The control of NF-[kappa]B activity. Annu. Rev. Immunol. 2000, 18, 621–663. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, K.; Naumann, M. Control of NF-kappaB activation by the COP9 signalosome. Biochem. Soc. Trans. 2010, 38, 156–161. [Google Scholar] [CrossRef]
- Zhou, Q.; Cheng, C.; Wei, Y.; Yang, J.; Zhou, W.; Song, Q.; Ke, M.; Yan, W.; Zheng, L.; Zhang, Y.; et al. USP15 potentiates NF-kappaB activation by differentially stabilizing TAB2 and TAB3. FEBS J. 2020, 287, 3165–3183. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Jiang, H.; Du, J.; Li, L.; Li, R.; Lu, J.; Fu, W.; Hou, J. USP15 inhibits multiple myeloma cell apoptosis through activating a feedback loop with the transcription factor NF-kappaBp65. Exp. Mol. Med. 2018, 50, 1–12. [Google Scholar] [CrossRef]
- Fan, Y.H.; Yu, Y.; Mao, R.F.; Tan, X.J.; Xu, G.F.; Zhang, H.; Lu, X.B.; Fu, S.B.; Yang, J. USP4 targets TAK1 to downregulate TNFalpha-induced NF-kappaB activation. Cell Death Differ. 2011, 18, 1547–1560. [Google Scholar] [CrossRef]
- Li, Z.; Hao, Q.; Luo, J.; Xiong, J.; Zhang, S.; Wang, T.; Bai, L.; Wang, W.; Chen, M.; Wang, W.; et al. USP4 inhibits p53 and NF-kappaB through deubiquitinating and stabilizing HDAC2. Oncogene 2016, 35, 2902–2912. [Google Scholar] [CrossRef]
- Sun, W.; Tan, X.; Shi, Y.; Xu, G.; Mao, R.; Gu, X.; Fan, Y.; Yu, Y.; Burlingame, S.; Zhang, H.; et al. USP11 negatively regulates TNFalpha-induced NF-kappaB activation by targeting on IkappaBalpha. Cell. Signal. 2010, 22, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef]
- Oikonomaki, M.; Bady, P.; Hegi, M.E. Ubiquitin specific peptidase 15 (USP15) suppresses glioblastoma cell growth via stabilization of HECTD1 E3 ligase attenuating WNT pathway activity. Oncotarget 2017, 8, 110490–110502. [Google Scholar] [CrossRef] [PubMed]
- Greenblatt, M.B.; Shin, D.Y.; Oh, H.; Lee, K.Y.; Zhai, B.; Gygi, S.P.; Lotinun, S.; Baron, R.; Liu, D.; Su, B.; et al. MEKK2 mediates an alternative beta-catenin pathway that promotes bone formation. Proc. Natl. Acad. Sci. USA 2016, 113, E1226–E1235. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.I.; Kim, H.H.; Yoon, J.H.; Park, W.S.; Hahn, M.J.; Kim, H.C.; Chung, C.H.; Kim, K.K. Ubiquitin specific protease 4 positively regulates the WNT/beta-catenin signaling in colorectal cancer. Mol. Oncol. 2015, 9, 1834–1851. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Kimura, J.; Miki, Y.; Yoshida, K. The deubiquitinating enzyme USP11 controls an IkappaB kinase alpha (IKKalpha)-p53 signaling pathway in response to tumor necrosis factor alpha (TNFalpha). J. Biol. Chem. 2007, 282, 33943–33948. [Google Scholar] [CrossRef] [PubMed]
- White, M.F. IRS proteins and the common path to diabetes. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E413–E422. [Google Scholar] [CrossRef]
- Li, S.; Hong, X.; Wei, Z.; Xie, M.; Li, W.; Liu, G.; Guo, H.; Yang, J.; Wei, W.; Zhang, S. Ubiquitination of the HPV oncoprotein E6 is critical for E6/E6AP-mediated p53 degradation. Front. Microbiol. 2019, 10, 2483. [Google Scholar] [CrossRef]
- Jones, J.I.; Clemmons, D.R. Insulin-like growth factors and their binding proteins: Biological actions. Endocr. Rev. 1995, 16, 3–34. [Google Scholar] [CrossRef]
- Niederkorn, M.; Hueneman, K.; Choi, K.; Varney, M.E.; Romano, L.; Pujato, M.A.; Greis, K.D.; Inoue, J.I.; Meetei, R.; Starczynowski, D.T. TIFAB regulates USP15-mediated p53 signaling during stressed and malignant hematopoiesis. Cell Rep. 2020, 30, 2776–2790.e6. [Google Scholar] [CrossRef]
- Padmanabhan, A.; Candelaria, N.; Wong, K.K.; Nikolai, B.C.; Lonard, D.M.; O’Malley, B.W.; Richards, J.S. USP15-dependent lysosomal pathway controls p53-R175H turnover in ovarian cancer cells. Nat. Commun. 2018, 9, 1270. [Google Scholar] [CrossRef]
- Vos, R.M.; Altreuter, J.; White, E.A.; Howley, P.M. The ubiquitin-specific peptidase USP15 regulates human papillomavirus type 16 E6 protein stability. J. Virol. 2009, 83, 8885–8892. [Google Scholar] [CrossRef]
- Koivusalo, R.; Mialon, A.; Pitkanen, H.; Westermarck, J.; Hietanen, S. Activation of p53 in cervical cancer cells by human papillomavirus E6 RNA interference is transient, but can be sustained by inhibiting endogenous nuclear export-dependent p53 antagonists. Cancer Res. 2006, 66, 11817–11824. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.T.; Huang, K.Y.; Lu, M.C.; Huang, H.L.; Chen, C.Y.; Cheng, Y.L.; Yu, H.C.; Liu, S.Q.; Lai, N.S.; Huang, H.B. TGF-beta upregulates the translation of USP15 via the PI3K/AKT pathway to promote p53 stability. Oncogene 2017, 36, 2715–2723. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Berger, F.G.; Yang, J.; Lu, X. USP4 inhibits p53 through deubiquitinating and stabilizing ARF-BP1. EMBO J. 2011, 30, 2177–2189. [Google Scholar] [CrossRef]
- Ke, J.Y.; Dai, C.J.; Wu, W.L.; Gao, J.H.; Xia, A.J.; Liu, G.P.; Lv, K.S.; Wu, C.L. USP11 regulates p53 stability by deubiquitinating p53. J. Zhejiang Univ. Sci. B 2014, 15, 1032–1038. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, T.; Yoshihara, H.; Furuta, H.; Hakuno, F.; Iemura, S.I.; Natsume, T.; Nakatsu, Y.; Kamata, H.; Asano, T.; Komada, M.; et al. USP15 attenuates IGF-I signaling by antagonizing Nedd4-induced IRS-2 ubiquitination. Biochem. Biophys. Res. Commun. 2017, 484, 522–528. [Google Scholar] [CrossRef]
- Pollak, M. Insulin and insulin-like growth factor signalling in neoplasia. Nat. Rev. Cancer. 2008, 8, 915–928. [Google Scholar] [CrossRef]
- Park, M.K.; Yao, Y.; Xia, W.; Setijono, S.R.; Kim, J.H.; Vila, I.K.; Chiu, H.H.; Wu, Y.; Billalabeitia, E.G.; Lee, M.G.; et al. PTEN self-regulates through USP11 via the PI3K-FOXO pathway to stabilize tumor suppression. Nat. Commun. 2019, 10, 636. [Google Scholar] [CrossRef]
- Inn, K.S.; Gack, M.U.; Tokunaga, F.; Shi, M.; Wong, L.Y.; Iwai, K.; Jung, J.U. Linear ubiquitin assembly complex negatively regulates RIG-I- and TRIM25-mediated type I interferon induction. Mol. Cell. 2011, 41, 354–365. [Google Scholar] [CrossRef]
- Pauli, E.K.; Chan, Y.K.; Davis, M.E.; Gableske, S.; Wang, M.K.; Feister, K.F.; Gack, M.U. The ubiquitin-specific protease USP15 promotes RIG-I-mediated antiviral signaling by deubiquitylating TRIM25. Sci. Signal. 2014, 7, ra3. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Zhang, X.; van Dam, H.; ten Dijke, P.; Huang, H.; Zhang, L. Ubiquitin-specific protease 4 mitigates Toll-like/interleukin-1 receptor signaling and regulates innate immune activation. J. Biol. Chem. 2012, 287, 11002–11010. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Nie, J.; Ren, J.; Liang, R.; Li, D.; Wang, P.; Gao, C.; Zhuo, C.; Yang, C.; Li, B. USP4 interacts and positively regulates IRF8 function via K48-linked deubiquitination in regulatory T cells. FEBS Lett. 2017, 591, 1677–1686. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Xu, P.; Ge, S.; Zhang, C.; Zheng, X.; Xu, J.; Liu, Z.; Li, B.; Ge, S. Ubiquitin specific peptidase 4 stabilizes interferon regulatory factor protein and promotes its function to facilitate interleukin-4 expression in T helper type 2 cells. Int. J. Mol. Med. 2017, 40, 979–986. [Google Scholar] [CrossRef]
- Zhang, D.D.; Lo, S.C.; Sun, Z.; Habib, G.M.; Lieberman, M.W.; Hannink, M. Ubiquitination of Keap1, a BTB-Kelch substrate adaptor protein for Cul3, targets Keap1 for degradation by a proteasome-independent pathway. J. Biol. Chem. 2005, 280, 30091–30099. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Bao, G.; Liu, F. Inhibition of USP15 prevent glutamate-induced oxidative damage by activating Nrf2/HO-1 signaling pathway in HT22 Cells. Cell Mol. Neurobiol. 2020, 40, 999–1010. [Google Scholar] [CrossRef]
- Lynch, K.W. Regulation of alternative splicing by signal transduction pathways. Adv. Exp. Med. Biol. 2007, 623, 161–174. [Google Scholar] [CrossRef]
- Song, E.J.; Werner, S.L.; Neubauer, J.; Stegmeier, F.; Aspden, J.; Rio, D.; Harper, J.W.; Elledge, S.J.; Kirschner, M.W.; Rape, M. The Prp19 complex and the Usp4Sart3 deubiquitinating enzyme control reversible ubiquitination at the spliceosome. Genes Dev. 2010, 24, 1434–1447. [Google Scholar] [CrossRef]
- Leeman, J.R.; Gilmore, T.D. Alternative splicing in the NF-kappaB signaling pathway. Gene 2008, 423, 97–107. [Google Scholar] [CrossRef]
- Das, T.; Shin, S.C.; Song, E.J.; Kim, E.E. Regulation of deubiquitinating enzymes by post-translational modifications. Int. J. Mol. Sci. 2020, 21, 4028. [Google Scholar] [CrossRef]
- Lai, K.P.; Chen, J.; Tse, W.K.F. Role of deubiquitinases in human cancers: Potential targeted therapy. Int. J. Mol. Sci. 2020, 21, 2548. [Google Scholar] [CrossRef] [PubMed]
- Heideker, J.; Wertz, I.E. DUBs, the regulation of cell identity and disease. Biochem. J. 2015, 467, 191. [Google Scholar] [CrossRef]
- Harrigan, J.A.; Jacq, X.; Martin, N.M.; Jackson, S.P. Deubiquitylating enzymes and drug discovery: Emerging opportunities. Nat. Rev. Drug Discov. 2018, 17, 57–78. [Google Scholar] [CrossRef]
- Xu, M.; Takanashi, M.; Oikawa, K.; Tanaka, M.; Nishi, H.; Isaka, K.; Kudo, M.; Kuroda, M. USP15 plays an essential role for caspase-3 activation during Paclitaxel-induced apoptosis. Biochem. Biophys. Res. Commun. 2009, 388, 366–371. [Google Scholar] [CrossRef]
- Xie, L.; Wei, J.; Qian, X.; Chen, G.; Yu, L.; Ding, Y.; Liu, B. CXCR4, a potential predictive marker for docetaxel sensitivity in gastric cancer. Anticancer Res. 2010, 30, 2209–2216. [Google Scholar]
- Peng, Y.; Liao, Q.; Tan, W.; Peng, C.; Hu, Z.; Chen, Y.; Li, Z.; Li, J.; Zhen, B.; Zhu, W.; et al. The deubiquitylating enzyme USP15 regulates homologous recombination repair and cancer cell response to PARP inhibitors. Nat. Commun. 2019, 10, 1224. [Google Scholar] [CrossRef]
- Cornelissen, T.; Haddad, D.; Wauters, F.; van Humbeeck, C.; Mandemakers, W.; Koentjoro, B.; Sue, C.; Gevaert, K.; de Strooper, B.; Verstreken, P.; et al. The deubiquitinase USP15 antagonizes Parkin-mediated mitochondrial ubiquitination and mitophagy. Hum. Mol. Genet. 2014, 23, 5227–5242. [Google Scholar] [CrossRef]
- Menzies, F.M.; Huebener, J.; Renna, M.; Bonin, M.; Riess, O.; Rubinsztein, D.C. Autophagy induction reduces mutant ataxin-3 levels and toxicity in a mouse model of spinocerebellar ataxia type 3. Brain 2010, 133, 93–104. [Google Scholar] [CrossRef]
- Bardehle, S.; Rafalski, V.; Akassoglou, K. Finding the ‘ubiquitous’ threads in infection and autoimmune neuroinflammation. Nat. Immunol. 2016, 18, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Poondla, N.; Chandrasekaran, A.P.; Kim, K.S.; Ramakrishna, S. Deubiquitinating enzymes as cancer biomarkers: New therapeutic opportunities? BMB Rep. 2019, 52, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Tanguturi, P.; Kim, K.S.; Ramakrishna, S. The role of deubiquitinating enzymes in cancer drug resistance. Cancer Chemother. Pharmacol. 2020, 85, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Schauer, N.J.; Magin, R.S.; Liu, X.; Doherty, L.M.; Buhrlage, S.J. Advances in discovering deubiquitinating enzyme (DUB) inhibitors. J. Med. Chem. 2020, 63, 2731–2750. [Google Scholar] [CrossRef] [PubMed]
- Teyra, J.; Singer, A.U.; Schmitges, F.W.; Jaynes, P.; Kit Leng Lui, S.; Polyak, M.J.; Fodil, N.; Krieger, J.R.; Tong, J.; Schwerdtfeger, C.; et al. Structural and functional characterization of ubiquitin variant inhibitors of USP15. Structure 2019, 27, 590–605.e5. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Pathway | Target Substrate | Mechanism of Action | Effect on Cells | Reference |
|---|---|---|---|---|
| TGF-β signaling | ALK5 | Deubiquitination and stabilization of TβR-I by making a complex with SMAD7 and SMURF2 | Enhances the activity of TGF-β signaling and promotes oncogenesis in glioblastoma | [13] |
| BMP signaling | ALK3 | Deubiquitination of type I BMP receptor ALK3 by interacting with BMP inhibitor SMAD6 | Enhances BMP target gene transcription in mammalian cells, osteoblastic differentiation in mouse, and embryogenesis in Xenopus | [39] |
| COP9 signalosome | CRLs | Deubiquitination and stabilization of CRL adaptor proteins and associated substrates | Regulation of cullin activity | [58] |
| Regulation of multicellular development and secondary metabolism | [59] | |||
| Protects and stabilizes the components of CRLs for rearrangement and adaptation to altered cellular requirements | [60] | |||
| NF-κB signaling | IκBα | Deubiquitination and stabilization of IκBα | Inhibition of NF-ĸB signaling and downstream gene expression | [33] |
| NF-κB p65 | Inhibition of NF-κBp65 ubiquitination | Induces cell proliferation and inhibition of cell apoptosis in multiple myeloma by activating NF-κB signaling | [68] | |
| TAB2 and TAB3 | Stabilization of TAB2 and TAB3 by inhibiting their proteolysis | Potentiates TNFα- or IL-1β-induced NF-ĸB activation and downstream gene transcription | [67] | |
| Wnt/β-catenin signaling | APC | Stabilization of APC in the β-catenin destruction complex | Suppresses Wnt/β-catenin signaling and target gene expression | [35] |
| HECTD1 | Deubiquitination and stabilization of HECTD1 | Tumor suppression in a subset of glioblastoma by attenuating the canonical Wnt pathway | [73] | |
| β-catenin | Stabilization of β-catenin by inhibiting degradation | Osteoblast differentiation and bone formation | [74] | |
| p53 signaling | MDM2 | Deubiquitination and stabilization of MDM2 | Downregulation of p53 along with its target genes and cancer cell survival | [34] |
| MDM2 via TIFAB | Increases the deubiquitination and stabilization of MDM2, which is boosted by TIFAB | Suppresses p53 activity in stressed and malignant hematopoietic cells | [80] | |
| E6 | Increases E6 protein stability | Upregulation of E6 oncoproteins may facilitate cancer progression in HPV-infected cells | [82] | |
| p53 | Binding and stabilization of p53 | TGF-β signaling-mediated stabilization of p53 through increased USP15 translation that suppresses the early stages of cancer progression | [84] | |
| IGF signaling | IRS-2 | Binding to and antagonizing of IRS-2 ubiquitination | Regulation of cancer cell progression by fine-tuning of IGF-induced growth-promoting activity | [87] |
| Immune signaling | TRIM25 | Deubiquitination and stabilization of TRIM25 | Promotes antiviral innate immune responses | [91] |
| Nrf2–Keap1 signaling | Keap1 | Deubiquitination of Keap1 and enhancement of the Keap1–Cul3–E3 ligase complex stability | Promotes Nrf2 protein degradation and reduces the Nrf2 target gene expression | [38] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Das, T.; Song, E.J.; Kim, E.E. The Multifaceted Roles of USP15 in Signal Transduction. Int. J. Mol. Sci. 2021, 22, 4728. https://doi.org/10.3390/ijms22094728
Das T, Song EJ, Kim EE. The Multifaceted Roles of USP15 in Signal Transduction. International Journal of Molecular Sciences. 2021; 22(9):4728. https://doi.org/10.3390/ijms22094728
Chicago/Turabian StyleDas, Tanuza, Eun Joo Song, and Eunice EunKyeong Kim. 2021. "The Multifaceted Roles of USP15 in Signal Transduction" International Journal of Molecular Sciences 22, no. 9: 4728. https://doi.org/10.3390/ijms22094728
APA StyleDas, T., Song, E. J., & Kim, E. E. (2021). The Multifaceted Roles of USP15 in Signal Transduction. International Journal of Molecular Sciences, 22(9), 4728. https://doi.org/10.3390/ijms22094728