
Chemokine-Cytokine Networks in the Head and Neck Tumor Microenvironment

 , , , and
, , , and
Abstract
1. Introduction
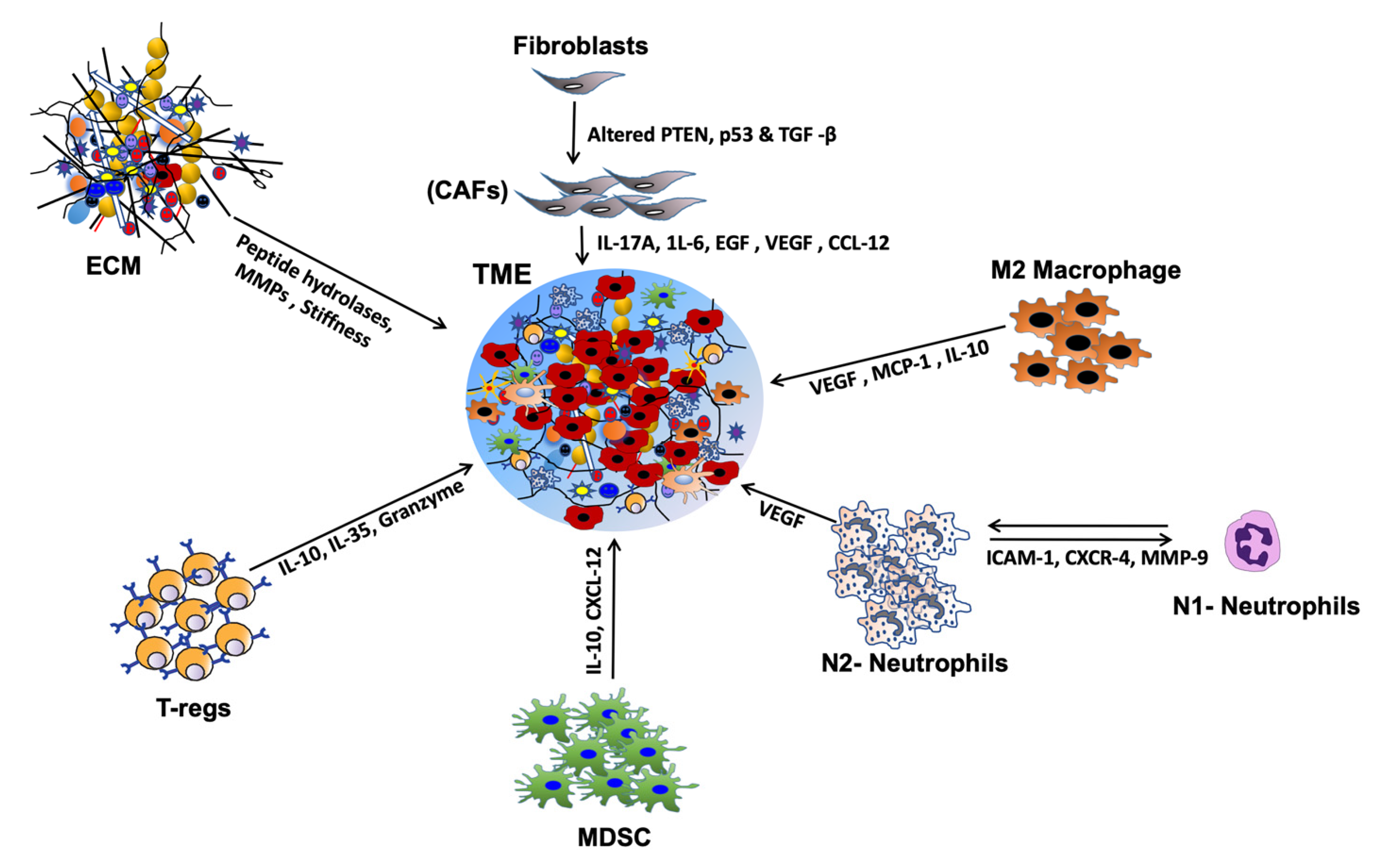
2. HNSCC Tumor Microenvironment
Extracellular Matrix and Chemokine/Cytokine Activation
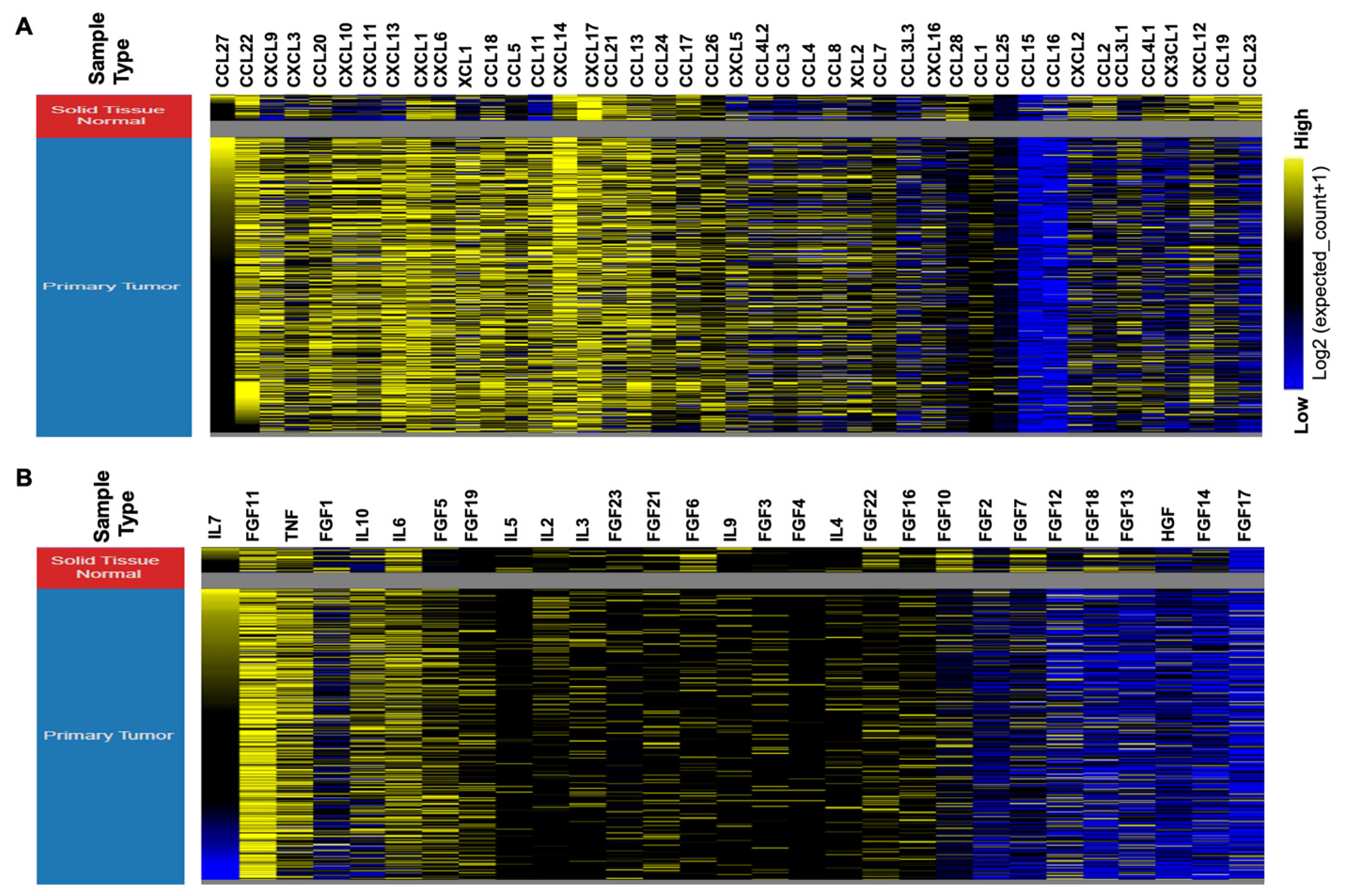
3. Deregulated Chemokine and Cytokine Expression in HNSCC
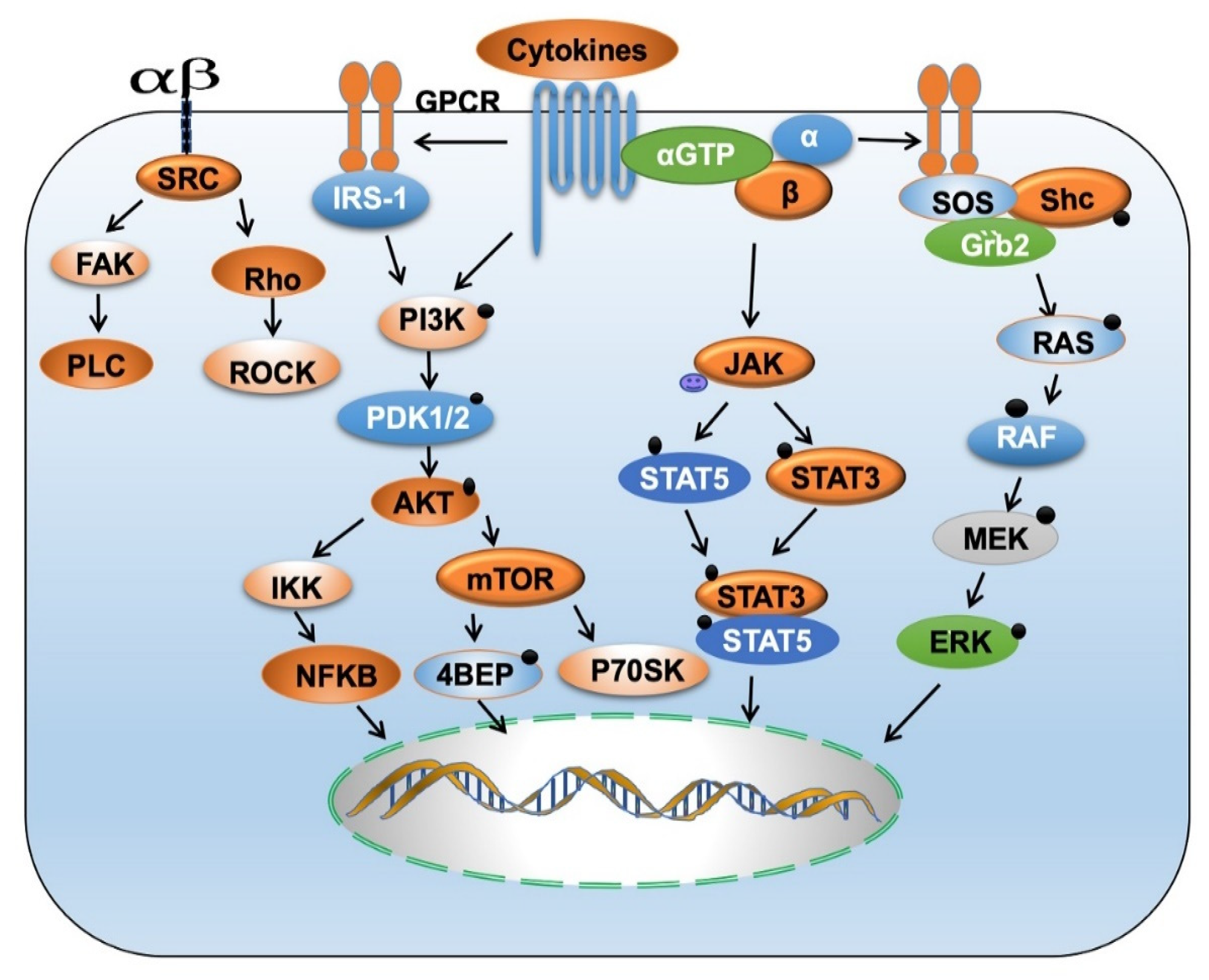
4. Chemokine and Cytokine Mediated Signaling Pathways in HNSCC
5. Chemokines and Cytokines Promote Aggressive HNSCC Phenotype
6. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| TAM | tumor associated macrophages |
| DC | dendritic cells |
| MDSC | myeloid derived suppressor cells |
| CAF | cancer associated fibroblasts |
| NK cells | natural killer cells |
| IL | interleukins |
| Tregs | T regulatory cells |
| CSC | cancer stem cells |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Leemans, C.R.; Braakhuis, B.J.; Brakenhoff, R.H. The molecular biology of head and neck cancer. Nat. Rev. Cancer 2011, 11, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Blot, W.J.; McLaughlin, J.K.; Winn, D.M.; Austin, D.F.; Greenberg, R.S.; Preston-Martin, S.; Bernstein, L.; Schoenberg, J.B.; Stemhagen, A.; Fraumeni, J.F., Jr. Smoking and drinking in relation to oral and pharyngeal cancer. Cancer Res. 1988, 48, 3282–3287. [Google Scholar]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef]
- Commins, S.P.; Borish, L.; Steinke, J.W. Immunologic messenger molecules: Cytokines, interferons, and chemokines. J. Allergy Clin. Immunol. 2010, 125 (Suppl. 2), S53–S72. [Google Scholar] [CrossRef]
- Hromas, R.; Broxmeyer, H.E.; Kim, C.; Nakshatri, H.; Christopherson, K., 2nd; Azam, M.; Hou, Y.H. Cloning of BRAK, a novel divergent CXC chemokine preferentially expressed in normal versus malignant cells. Biochem. Biophys. Res. Commun. 1999, 255, 703–706. [Google Scholar] [CrossRef]
- Bacon, K.; Baggiolini, M.; Broxmeyer, H.; Horuk, R.; Lindley, I.; Mantovani, A.; Maysushima, K.; Murphy, P.; Nomiyama, H.; Oppenheim, J.; et al. Chemokine/chemokine receptor nomenclature. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 2002, 22, 1067–1068. [Google Scholar]
- Cameron, M.J.; Kelvin, D.J. Cytokines and chemokines--their receptors and their genes: An overview. Adv. Exp. Med. Biol. 2003, 520, 8–32. [Google Scholar]
- Strieter, R.M.; Burdick, M.D.; Mestas, J.; Gomperts, B.; Keane, M.P.; Belperio, J.A. Cancer CXC chemokine networks and tumour angiogenesis. Eur. J. Cancer 2006, 42, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Luster, A.D. Chemokines—Chemotactic cytokines that mediate inflammation. N. Engl. J. Med. 1998, 338, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Belperio, J.A.; Keane, M.P.; Arenberg, D.A.; Addison, C.L.; Ehlert, J.E.; Burdick, M.D.; Strieter, R.M. CXC chemokines in angiogenesis. J. Leukoc. Biol. 2000, 68, 1–8. [Google Scholar]
- Borish, L.C.; Steinke, J.W. 2. Cytokines and chemokines. J. Allergy Clin. Immunol. 2003, 111 (Suppl. 2), S460–S475. [Google Scholar] [CrossRef] [PubMed]
- Akdis, M.; Burgler, S.; Crameri, R.; Eiwegger, T.; Fujita, H.; Gomez, E.; Klunker, S.; Meyer, N.; O’Mahony, L.; Palomares, O.; et al. Interleukins, from 1 to 37, and interferon-γ: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 2011, 127, 701–721.e70. [Google Scholar] [CrossRef] [PubMed]
- Quezada, S.A.; Peggs, K.S.; Simpson, T.R.; Allison, J.P. Shifting the equilibrium in cancer immunoediting: From tumor tolerance to eradication. Immunol. Rev. 2011, 241, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Hathaway, B.; Landsittel, D.P.; Gooding, W.; Whiteside, T.L.; Grandis, J.R.; Siegfried, J.M.; Bigbee, W.L.; Ferris, R.L. Multiplexed analysis of serum cytokines as biomarkers in squamous cell carcinoma of the head and neck patients. Laryngoscope 2005, 115, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Linkov, F.; Lisovich, A.; Yurkovetsky, Z.; Marrangoni, A.; Velikokhatnaya, L.; Nolen, B.; Winans, M.; Bigbee, W.; Siegfried, J.; Lokshin, A.; et al. Early detection of head and neck cancer: Development of a novel screening tool using multiplexed immunobead-based biomarker profiling. Cancer Epidemiol. Biomark. Prev. A Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2007, 16, 102–107. [Google Scholar] [CrossRef]
- Druzgal, C.H.; Chen, Z.; Yeh, N.T.; Thomas, G.R.; Ondrey, F.G.; Duffey, D.C.; Vilela, R.J.; Ende, K.; McCullagh, L.; Rudy, S.F.; et al. A pilot study of longitudinal serum cytokine and angiogenesis factor levels as markers of therapeutic response and survival in patients with head and neck squamous cell carcinoma. Head Neck 2005, 27, 771–784. [Google Scholar] [CrossRef]
- Johnson, S.D.; De Costa, A.M.; Young, M.R. Effect of the premalignant and tumor microenvironment on immune cell cytokine production in head and neck cancer. Cancers 2014, 6, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Mehanna, H.; Robinson, M.; Hartley, A.; Kong, A.; Foran, B.; Fulton-Lieuw, T.; Dalby, M.; Mistry, P.; Sen, M.; O’Toole, L.; et al. Radiotherapy plus cisplatin or cetuximab in low-risk human papillomavirus-positive oropharyngeal cancer (De-ESCALaTE HPV): An open-label randomised controlled phase 3 trial. Lancet 2019, 393, 51–60. [Google Scholar] [CrossRef]
- Dhawan, P.; Richmond, A. Role of CXCL1 in tumorigenesis of melanoma. J. Leukoc. Biol. 2002, 72, 9–18. [Google Scholar]
- Wilson, J.; Balkwill, F. The role of cytokines in the epithelial cancer microenvironment. Semin. Cancer Biol. 2002, 12, 113–120. [Google Scholar] [CrossRef]
- Giancotti, F.G. Deregulation of cell signaling in cancer. Febs Lett. 2014, 88, 2558–2570. [Google Scholar] [CrossRef]
- Chen, F.; Zhuang, X.; Lin, L.; Yu, P.; Wang, Y.; Shi, Y.; Hu, G.; Sun, Y. New horizons in tumor microenvironment biology: Challenges and opportunities. BMC Med. 2015, 13, 45. [Google Scholar] [CrossRef]
- Bhat, A.A.; Yousuf, P.; Wani, N.A.; Rizwan, A.; Chauhan, S.S.; Siddiqi, M.A.; Bedognetti, D.; El-Rifai, W.; Frenneaux, M.P.; Batra, S.K.; et al. Tumor microenvironment: An evil nexus promoting aggressive head and neck squamous cell carcinoma and avenue for targeted therapy. Signal. Transduct Target. 2021, 6, 12. [Google Scholar] [CrossRef]
- Cheng, L.; Huang, Z.; Zhou, W.; Wu, Q.; Donnola, S.; Liu, J.K.; Fang, X.; Sloan, A.E.; Mao, Y.; Lathia, J.D.; et al. Glioblastoma stem cells generate vascular pericytes to support vessel function and tumor growth. Cell 2013, 153, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Tlsty, T.D.; Hein, P.W. Know thy neighbor: Stromal cells can contribute oncogenic signals. Curr. Opin. Genet. Dev. 2001, 11, 54–59. [Google Scholar] [CrossRef]
- Canning, M.; Guo, G.; Yu, M.; Myint, C.; Groves, M.W.; Byrd, J.K.; Cui, Y. Heterogeneity of the Head and Neck Squamous Cell Carcinoma Immune Landscape and Its Impact on Immunotherapy. Front. Cell Dev. Biol. 2019, 7, 52. [Google Scholar] [CrossRef]
- Sazeides, C.; Le, A. Metabolic Relationship between Cancer-Associated Fibroblasts and Cancer Cells. Adv. Exp. Med. Biol. 2018, 1063, 149–165. [Google Scholar] [PubMed]
- Xing, F.; Saidou, J.; Watabe, K. Cancer associated fibroblasts (CAFs) in tumor microenvironment. Front. Biosci 2010, 15, 166–179. [Google Scholar] [CrossRef]
- Kalluri, R. The biology and function of fibroblasts in cancer. Nat. Rev. Cancer 2016, 16, 582–598. [Google Scholar] [CrossRef]
- Augsten, M.; Sjöberg, E.; Frings, O.; Vorrink, S.U.; Frijhoff, J.; Olsson, E.; Borg, Å.; Östman, A. Cancer-associated fibroblasts expressing CXCL14 rely upon NOS1-derived nitric oxide signaling for their tumor-supporting properties. Cancer Res. 2014, 74, 2999–3010. [Google Scholar] [CrossRef]
- Jia, C.C.; Wang, T.T.; Liu, W.; Fu, B.S.; Hua, X.; Wang, G.Y.; Li, T.J.; Li, X.; Wu, X.Y.; Tai, Y.; et al. Cancer-associated fibroblasts from hepatocellular carcinoma promote malignant cell proliferation by HGF secretion. PLoS ONE 2013, 8, e63243. [Google Scholar] [CrossRef]
- Glentis, A.; Oertle, P.; Mariani, P.; Chikina, A.; El Marjou, F.; Attieh, Y.; Zaccarini, F.; Lae, M.; Loew, D.; Dingli, F.; et al. Cancer-associated fibroblasts induce metalloprotease-independent cancer cell invasion of the basement membrane. Nat. Commun. 2017, 8, 924. [Google Scholar] [CrossRef] [PubMed]
- Hinsley, E.E.; Hunt, S.; Hunter, K.D.; Whawell, S.A.; Lambert, D.W. Endothelin-1 stimulates motility of head and neck squamous carcinoma cells by promoting stromal-epithelial interactions. Int. J. Cancer 2012, 130, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci 2018, 19, 1801. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Xu, J.Y.; Shi, X.Y.; Huang, W.; Ruan, T.Y.; Xie, P.; Ding, J.L. M2-polarized tumor-associated macrophages promoted epithelial-mesenchymal transition in pancreatic cancer cells, partially through TLR4/IL-10 signaling pathway. Lab. Investig. A J. Tech. Methods Pathol. 2013, 93, 844–854. [Google Scholar] [CrossRef]
- Peltanova, B.; Raudenska, M.; Masarik, M. Effect of tumor microenvironment on pathogenesis of the head and neck squamous cell carcinoma: A systematic review. Mol. Cancer 2019, 18, 63. [Google Scholar] [CrossRef]
- Wolf, G.T.; Chepeha, D.B.; Bellile, E.; Nguyen, A.; Thomas, D.; McHugh, J. Tumor infiltrating lymphocytes (TIL) and prognosis in oral cavity squamous carcinoma: A preliminary study. Oral Oncol. 2015, 51, 90–95. [Google Scholar] [CrossRef]
- Galdiero, M.R.; Garlanda, C.; Jaillon, S.; Marone, G.; Mantovani, A. Tumor associated macrophages and neutrophils in tumor progression. J. Cell. Physiol. 2013, 228, 1404–1412. [Google Scholar] [CrossRef] [PubMed]
- Liew, P.X.; Kubes, P. The Neutrophil’s Role During Health and Disease. Physiol. Rev. 2019, 99, 1223–1248. [Google Scholar] [CrossRef] [PubMed]
- Paneesha, S.; McManus, A.; Arya, R.; Scriven, N.; Farren, T.; Nokes, T.; Bacon, S.; Nieland, A.; Cooper, D.; Smith, H.; et al. Frequency, demographics and risk (according to tumour type or site) of cancer-associated thrombosis among patients seen at outpatient DVT clinics. Thromb. Haemost. 2010, 103, 338–343. [Google Scholar] [CrossRef]
- Stabile, H.; Fionda, C.; Gismondi, A.; Santoni, A. Role of Distinct Natural Killer Cell Subsets in Anticancer Response. Front. Immunol. 2017, 8, 293. [Google Scholar] [CrossRef]
- Teng, T.S.; Ji, A.L.; Ji, X.Y.; Li, Y.Z. Neutrophils and Immunity: From Bactericidal Action to Being Conquered. J. Immunol. Res. 2017, 2017, 9671604. [Google Scholar] [CrossRef] [PubMed]
- Korrer, M.J.; Kim, Y. Phenotypical and functional analysis of natural killer cells from primary human head and neck squamous cell carcinomas. J. Immunol. 2019, 202 (Suppl. 1), 134.19. [Google Scholar]
- Chan, C.J.; Smyth, M.J.; Martinet, L. Molecular mechanisms of natural killer cell activation in response to cellular stress. Cell death and differentiation 2014, 21, 5–14. [Google Scholar] [CrossRef]
- Horinaka, A.; Sakurai, D.; Ihara, F.; Makita, Y.; Kunii, N.; Motohashi, S.; Nakayama, T.; Okamoto, Y. Invariant NKT cells are resistant to circulating CD15+ myeloid-derived suppressor cells in patients with head and neck cancer. Cancer Sci. 2016, 107, 207–216. [Google Scholar] [CrossRef]
- Takami, M.; Ihara, F.; Motohashi, S. Clinical Application of iNKT Cell-mediated Anti-tumor Activity Against Lung Cancer and Head and Neck Cancer. Front. Immunol. 2018, 9, 2021. [Google Scholar] [CrossRef]
- Youn, J.I.; Nagaraj, S.; Collazo, M.; Gabrilovich, D.I. Subsets of myeloid-derived suppressor cells in tumor-bearing mice. J. Immunol. 2008, 181, 5791–5802. [Google Scholar] [CrossRef]
- Garrity, T.; Pandit, R.; Wright, M.A.; Benefield, J.; Keni, S.; Young, M.R. Increased presence of CD34+ cells in the peripheral blood of head and neck cancer patients and their differentiation into dendritic cells. Int. J. Cancer 1997, 73, 663–669. [Google Scholar] [CrossRef]
- Pak, A.S.; Wright, M.A.; Matthews, J.P.; Collins, S.L.; Petruzzelli, G.J.; Young, M.R. Mechanisms of immune suppression in patients with head and neck cancer: Presence of CD34(+) cells which suppress immune functions within cancers that secrete granulocyte-macrophage colony-stimulating factor. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 1995, 1, 95–103. [Google Scholar]
- Butt, A.Q.; Mills, K.H. Immunosuppressive networks and checkpoints controlling antitumor immunity and their blockade in the development of cancer immunotherapeutics and vaccines. Oncogene 2014, 33, 4623–4631. [Google Scholar] [CrossRef] [PubMed]
- Chikamatsu, K.; Sakakura, K.; Toyoda, M.; Takahashi, K.; Yamamoto, T.; Masuyama, K. Immunosuppressive activity of CD14+ HLA-DR- cells in squamous cell carcinoma of the head and neck. Cancer Sci. 2012, 103, 976–983. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Lu, K.V.; Petritsch, C.; Liu, P.; Ganss, R.; Passegué, E.; Song, H.; Vandenberg, S.; Johnson, R.S.; Werb, Z.; et al. HIF1alpha induces the recruitment of bone marrow-derived vascular modulatory cells to regulate tumor angiogenesis and invasion. Cancer Cell 2008, 13, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.G.; Wang, J.; Wang, P.; Gray, J.D.; Horwitz, D.A. IL-2 is essential for TGF-beta to convert naive CD4+CD25- cells to CD25+Foxp3+ regulatory T cells and for expansion of these cells. J. Immunol. 2007, 178, 2018–2027. [Google Scholar] [CrossRef] [PubMed]
- Vignali, D.A.; Collison, L.W.; Workman, C.J. How regulatory T cells work. Nat. Rev. Immunol. 2008, 8, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Hanna, G.J.; Liu, H.; Jones, R.E.; Bacay, A.F.; Lizotte, P.H.; Ivanova, E.V.; Bittinger, M.A.; Cavanaugh, M.E.; Rode, A.J.; Schoenfeld, J.D.; et al. Defining an inflamed tumor immunophenotype in recurrent, metastatic squamous cell carcinoma of the head and neck. Oral Oncol. 2017, 67, 61–69. [Google Scholar] [CrossRef]
- Badoual, C.; Hans, S.; Merillon, N.; Van Ryswick, C.; Ravel, P.; Benhamouda, N.; Levionnois, E.; Nizard, M.; Si-Mohamed, A.; Besnier, N.; et al. PD-1-expressing tumor-infiltrating T cells are a favorable prognostic biomarker in HPV-associated head and neck cancer. Cancer Res. 2013, 73, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Shah, B.H.; Rasheed, H.; Rahman, I.H.; Shariff, A.H.; Khan, F.L.; Rahman, H.B.; Hanif, S.; Saeed, S.A. Molecular mechanisms involved in human platelet aggregation by synergistic interaction of platelet-activating factor and 5-hydroxytryptamine. Exp. Mol. Med. 2001, 33, 226–233. [Google Scholar] [CrossRef]
- Ruiz, F.A.; Lea, C.R.; Oldfield, E.; Docampo, R. Human platelet dense granules contain polyphosphate and are similar to acidocalcisomes of bacteria and unicellular eukaryotes. J. Biol. Chem. 2004, 279, 44250–44257. [Google Scholar] [CrossRef]
- Metzelaar, M.J.; Wijngaard, P.L.; Peters, P.J.; Sixma, J.J.; Nieuwenhuis, H.K.; Clevers, H.C. CD63 antigen. A novel lysosomal membrane glycoprotein, cloned by a screening procedure for intracellular antigens in eukaryotic cells. J. Biol. Chem. 1991, 266, 3239–3245. [Google Scholar] [CrossRef]
- Gleissner, C.A.; von Hundelshausen, P.; Ley, K. Platelet chemokines in vascular disease. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1920–1927. [Google Scholar] [CrossRef] [PubMed]
- Prussin, C.; Metcalfe, D.D. 4. IgE, mast cells, basophils, and eosinophils. J. Allergy Clin. Immunol. 2003, 111 (Suppl. 2), S486–S494. [Google Scholar] [CrossRef]
- Saleem, S.J.; Martin, R.K.; Morales, J.K.; Sturgill, J.L.; Gibb, D.R.; Graham, L.; Bear, H.D.; Manjili, M.H.; Ryan, J.J.; Conrad, D.H. Cutting edge: Mast cells critically augment myeloid-derived suppressor cell activity. J. Immunol. 2012, 189, 511–515. [Google Scholar] [CrossRef]
- Stoyanov, E.; Uddin, M.; Mankuta, D.; Dubinett, S.M.; Levi-Schaffer, F. Mast cells and histamine enhance the proliferation of non-small cell lung cancer cells. Lung Cancer 2012, 75, 38–44. [Google Scholar] [CrossRef]
- Ciurea, R.; Mărgăritescu, C.; Simionescu, C.; Stepan, A.; Ciurea, M. VEGF and his R1 and R2 receptors expression in mast cells of oral squamous cells carcinomas and their involvement in tumoral angiogenesis. Rom. J. Morphol. Embryol. Rev. Roum. Morphol. Embryol. 2011, 52, 1227–1232. [Google Scholar]
- Barth, P.J.; Schenck zu Schweinsberg, T.; Ramaswamy, A.; Moll, R. CD34+ fibrocytes, alpha-smooth muscle antigen-positive myofibroblasts, and CD117 expression in the stroma of invasive squamous cell carcinomas of the oral cavity, pharynx, and larynx. Virchows Arch. Int. J. Pathol. 2004, 444, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Levite, M. Nerve-driven immunity. The direct effects of neurotransmitters on T-cell function. Ann. N. Y. Acad. Sci. 2000, 917, 307–321. [Google Scholar] [CrossRef]
- Muñoz, M.; González-Ortega, A.; Rosso, M.; Robles-Frias, M.J.; Carranza, A.; Salinas-Martín, M.V.; Coveñas, R. The substance P/neurokinin-1 receptor system in lung cancer: Focus on the antitumor action of neurokinin-1 receptor antagonists. Peptides 2012, 38, 318–325. [Google Scholar] [CrossRef]
- Koon, H.W.; Zhao, D.; Na, X.; Moyer, M.P.; Pothoulakis, C. Metalloproteinases and transforming growth factor-alpha mediate substance P-induced mitogen-activated protein kinase activation and proliferation in human colonocytes. J. Biol. Chem. 2004, 279, 45519–45527. [Google Scholar] [CrossRef]
- Brener, S.; González-Moles, M.A.; Tostes, D.; Esteban, F.; Gil-Montoya, J.A.; Ruiz-Avila, I.; Bravo, M.; Muñoz, M. A role for the substance P/NK-1 receptor complex in cell proliferation in oral squamous cell carcinoma. Anticancer Res. 2009, 29, 2323–2329. [Google Scholar]
- Mehboob, R.; Tanvir, I.; Warraich, R.A.; Perveen, S.; Yasmeen, S.; Ahmad, F.J. Role of neurotransmitter Substance P in progression of oral squamous cell carcinoma. Pathol. Res. Pract. 2015, 211, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Lala, P.K.; Parhar, R.S.; Singh, P. Indomethacin therapy abrogates the prostaglandin-mediated suppression of natural killer activity in tumor-bearing mice and prevents tumor metastasis. Cell. Immunol. 1986, 99, 108–118. [Google Scholar] [CrossRef]
- Bernabé, D.G.; Tamae, A.C.; Biasoli, É.R.; Oliveira, S.H. Stress hormones increase cell proliferation and regulates interleukin-6 secretion in human oral squamous cell carcinoma cells. Brain Behav. Immunol. 2011, 25, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Shang, Z.J.; Liu, K.; Liang, D.F. Expression of beta2-adrenergic receptor in oral squamous cell carcinoma. J. Oral Pathol. Med. Off. Publ. Int. Assoc. Oral Pathol. Am. Acad. Oral Pathol. 2009, 38, 371–376. [Google Scholar]
- Liu, H.; Wang, C.; Xie, N.; Zhuang, Z.; Liu, X.; Hou, J.; Huang, H. Activation of adrenergic receptor β2 promotes tumor progression and epithelial mesenchymal transition in tongue squamous cell carcinoma. Int. J. Mol. Med. 2018, 41, 147–154. [Google Scholar] [CrossRef]
- Shurin, M.R. Dendritic cells presenting tumor antigen. Cancer Immunol. Immunother. 1996, 43, 158–164. [Google Scholar] [CrossRef]
- Lutz, M.B.; Schuler, G. Immature, semi-mature and fully mature dendritic cells: Which signals induce tolerance or immunity? Trends Immunol. 2002, 23, 445–449. [Google Scholar] [CrossRef]
- Chaux, P.; Moutet, M.; Faivre, J.; Martin, F.; Martin, M. Inflammatory cells infiltrating human colorectal carcinomas express HLA class II but not B7–1 and B7–2 costimulatory molecules of the T-cell activation. Lab. Investig. A J. Tech. Methods Pathol. 1996, 74, 975–983. [Google Scholar]
- Shurin, M.R.; Shurin, G.V.; Lokshin, A.; Yurkovetsky, Z.R.; Gutkin, D.W.; Chatta, G.; Zhong, H.; Han, B.; Ferris, R.L. Intratumoral cytokines/chemokines/growth factors and tumor infiltrating dendritic cells: Friends or enemies? Cancer Metastasis Rev. 2006, 25, 333–356. [Google Scholar] [CrossRef] [PubMed]
- Makarenkova, V.P.; Shurin, G.V.; Tourkova, I.L.; Balkir, L.; Pirtskhalaishvili, G.; Perez, L.; Gerein, V.; Siegfried, J.M.; Shurin, M.R. Lung cancer-derived bombesin-like peptides down-regulate the generation and function of human dendritic cells. J. Neuroimmunol. 2003, 145, 55–67. [Google Scholar] [CrossRef]
- Lizée, G.; Radvanyi, L.G.; Overwijk, W.W.; Hwu, P. Improving antitumor immune responses by circumventing immunoregulatory cells and mechanisms. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2006, 12, 4794–4803. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mahnke, K.; Schmitt, E.; Bonifaz, L.; Enk, A.H.; Jonuleit, H. Immature, but not inactive: The tolerogenic function of immature dendritic cells. Immunol. Cell Biol. 2002, 80, 477–483. [Google Scholar] [CrossRef]
- Hawiger, D.; Inaba, K.; Dorsett, Y.; Guo, M.; Mahnke, K.; Rivera, M.; Ravetch, J.V.; Steinman, R.M.; Nussenzweig, M.C. Dendritic cells induce peripheral T cell unresponsiveness under steady state conditions in vivo. J. Exp. Med. 2001, 194, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Role of angiogenesis in tumor growth and metastasis. Semin. Oncol. 2002, 29 (Suppl. 16), 15–18. [Google Scholar] [CrossRef]
- Eisma, R.J.; Spiro, J.D.; Kreutzer, D.L. Vascular endothelial growth factor expression in head and neck squamous cell carcinoma. Am. J. Surg. 1997, 174, 513–517. [Google Scholar] [CrossRef]
- Kaneko, T.; Zhang, Z.; Mantellini, M.G.; Karl, E.; Zeitlin, B.; Verhaegen, M.; Soengas, M.S.; Lingen, M.; Strieter, R.M.; Nunez, G.; et al. Bcl-2 orchestrates a cross-talk between endothelial and tumor cells that promotes tumor growth. Cancer Res. 2007, 67, 9685–9693. [Google Scholar] [CrossRef]
- Nör, J.E.; Christensen, J.; Mooney, D.J.; Polverini, P.J. Vascular endothelial growth factor (VEGF)-mediated angiogenesis is associated with enhanced endothelial cell survival and induction of Bcl-2 expression. Am. J. Pathol. 1999, 154, 375–384. [Google Scholar] [CrossRef]
- Nör, J.E.; Christensen, J.; Liu, J.; Peters, M.; Mooney, D.J.; Strieter, R.M.; Polverini, P.J. Up-Regulation of Bcl-2 in microvascular endothelial cells enhances intratumoral angiogenesis and accelerates tumor growth. Cancer Res. 2001, 61, 2183–2188. [Google Scholar]
- Karl, E.; Warner, K.; Zeitlin, B.; Kaneko, T.; Wurtzel, L.; Jin, T.; Chang, J.; Wang, S.; Wang, C.Y.; Strieter, R.M.; et al. Bcl-2 acts in a proangiogenic signaling pathway through nuclear factor-kappaB and CXC chemokines. Cancer Res. 2005, 65, 5063–5069. [Google Scholar] [CrossRef]
- Warner, K.A.; Miyazawa, M.; Cordeiro, M.M.; Love, W.J.; Pinsky, M.S.; Neiva, K.G.; Spalding, A.C.; Nör, J.E. Endothelial cells enhance tumor cell invasion through a crosstalk mediated by CXC chemokine signaling. Neoplasia 2008, 10, 131–139. [Google Scholar] [CrossRef]
- Zeng, Q.; Li, S.; Chepeha, D.B.; Giordano, T.J.; Li, J.; Zhang, H.; Polverini, P.J.; Nor, J.; Kitajewski, J.; Wang, C.Y. Crosstalk between tumor and endothelial cells promotes tumor angiogenesis by MAPK activation of Notch signaling. Cancer Cell 2005, 8, 13–23. [Google Scholar] [CrossRef]
- Pietras, K.; Ostman, A. Hallmarks of cancer: Interactions with the tumor stroma. Exp. Cell Res. 2010, 316, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Durham, J.T.; Surks, H.K.; Dulmovits, B.M.; Herman, I.M. Pericyte contractility controls endothelial cell cycle progression and sprouting: Insights into angiogenic switch mechanics. Am. J. Physiol. Cell Physiol. 2014, 307, C878–C892. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Nico, B.; Crivellato, E. The role of pericytes in angiogenesis. Int. J. Dev. Biol. 2011, 55, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, H.; Betsholtz, C. Endothelial-pericyte interactions in angiogenesis. Cell Tissue Res. 2003, 314, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Bergers, G.; Song, S. The role of pericytes in blood-vessel formation and maintenance. Neuro Oncol. 2005, 7, 452–464. [Google Scholar] [CrossRef]
- Mărgăritescu, C.; Simionescu, C.; Pirici, D.; Mogoantă, L.; Ciurea, R.; Stepan, A. Immunohistochemical characterization of tumoral vessels in oral squamous cell carcinoma. Rom. J. Morphol. Embryol. Rev. Roum. Morphol. Embryol. 2008, 49, 447–458. [Google Scholar]
- Valle, I.B.; Schuch, L.F.; da Silva, J.M.; Gala-García, A.; Diniz, I.M.A.; Birbrair, A.; Abreu, L.G.; Silva, T.A. Pericyte in Oral Squamous Cell Carcinoma: A Systematic Review. Head Neck Pathol. 2020, 14, 1080–1091. [Google Scholar] [CrossRef]
- Hollemann, D.; Yanagida, G.; Rüger, B.M.; Neuchrist, C.; Fischer, M.B. New vessel formation in peritumoral area of squamous cell carcinoma of the head and neck. Head Neck 2012, 34, 813–820. [Google Scholar] [CrossRef]
- Muncie, J.M.; Weaver, V.M. The Physical and Biochemical Properties of the Extracellular Matrix Regulate Cell Fate. Curr. Top. Dev. Biol. 2018, 130, 1–37. [Google Scholar]
- Urbanczyk, M.; Layland, S.L.; Schenke-Layland, K. The role of extracellular matrix in biomechanics and its impact on bioengineering of cells and 3D tissues. Matrix Biol. 2020, 85–86, 1–14. [Google Scholar] [CrossRef]
- Zhu, J.; Liang, L.; Jiao, Y.; Liu, L. Enhanced invasion of metastatic cancer cells via extracellular matrix interface. PLoS ONE 2015, 10, e0118058. [Google Scholar] [CrossRef]
- Schminke, B.; Frese, J.; Bode, C.; Goldring, M.B.; Miosge, N. Laminins and Nidogens in the Pericellular Matrix of Chondrocytes: Their Role in Osteoarthritis and Chondrogenic Differentiation. Am. J. Pathol. 2016, 186, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.L.; Xu, R.J.; Zheng, Y.Y.; Liu, J.; Teng, Y.S.; Li, Y.; Zhu, J. Expression of type IV collagen, metalloproteinase-2, metalloproteinase-9 and tissue inhibitor of metalloproteinase-1 in laryngeal squamous cell carcinomas. Asian Pac. J. Cancer Prev. 2011, 12, 3245–3249. [Google Scholar] [PubMed]
- Parikka, M.; Nissinen, L.; Kainulainen, T.; Bruckner-Tuderman, L.; Salo, T.; Heino, J.; Tasanen, K. Collagen XVII promotes integrin-mediated squamous cell carcinoma transmigration--a novel role for alphaIIb integrin and tirofiban. Exp. Cell Res. 2006, 312, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Koontongkaew, S.; Amornphimoltham, P.; Yapong, B. Tumor-stroma interactions influence cytokine expression and matrix metalloproteinase activities in paired primary and metastatic head and neck cancer cells. Cell Biol. Int. 2009, 33, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.M.; Onodera, Y.; Bissell, M.J.; Park, C.C. Breast cancer cells in three-dimensional culture display an enhanced radioresponse after coordinate targeting of integrin alpha5beta1 and fibronectin. Cancer Res. 2010, 70, 5238–5248. [Google Scholar] [CrossRef]
- Levental, K.R.; Yu, H.; Kass, L.; Lakins, J.N.; Egeblad, M.; Erler, J.T.; Fong, S.F.; Csiszar, K.; Giaccia, A.; Weninger, W.; et al. Matrix crosslinking forces tumor progression by enhancing integrin signaling. Cell 2009, 139, 891–906. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, P.P.; Eliceiri, K.W.; Campbell, J.M.; Inman, D.R.; White, J.G.; Keely, P.J. Collagen reorganization at the tumor-stromal interface facilitates local invasion. BMC Med. 2006, 4, 38. [Google Scholar] [CrossRef] [PubMed]
- Ziober, A.F.; Falls, E.M.; Ziober, B.L. The extracellular matrix in oral squamous cell carcinoma: Friend or foe? Head Neck 2006, 28, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Shinohara, M.; Nakamura, S.; Oka, M. An immunohistochemical study of the extracellular matrix in oral squamous cell carcinoma and its association with invasive and metastatic potential. Virchows Arch. Int. J. Pathol. 1994, 424, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Ballabh, R. Expression of type IV collagen in different histological grades of oral squamous cell carcinoma: An immunohistochemical study. J. Cancer Res. 2013, 9, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Shruthy, R.; Sharada, P.; Swaminathan, U.; Nagamalini, B. Immunohistochemical expression of basement membrane laminin in histological grades of oral squamous cell carcinoma: A semiquantitative analysis. J. Oral Maxillofac. Pathol. 2013, 17, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Firth, N.A.; Reade, P.C. The prognosis of oral mucosal squamous cell carcinomas: A comparison of clinical and histopathological grading and of laminin and type IV collagen staining. Aust Dent. J. 1996, 41, 83–86. [Google Scholar] [CrossRef]
- Fabricius, E.M.; Wildner, G.P.; Kruse-Boitschenko, U.; Hoffmeister, B.; Goodman, S.L.; Raguse, J.D. Immunohistochemical analysis of integrins alphavbeta3, alphavbeta5 and alpha5beta1, and their ligands, fibrinogen, fibronectin, osteopontin and vitronectin, in frozen sections of human oral head and neck squamous cell carcinomas. Exp. Ther. Med. 2011, 2, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Popovic, Z.V.; Sandhoff, R.; Sijmonsma, T.P.; Kaden, S.; Jennemann, R.; Kiss, E.; Tone, E.; Autschbach, F.; Platt, N.; Malle, E.; et al. Sulfated glycosphingolipid as mediator of phagocytosis: SM4s enhances apoptotic cell clearance and modulates macrophage activity. J. Immunol. 2007, 179, 6770–6782. [Google Scholar] [CrossRef] [PubMed]
- Ohara, K.; Ohkuri, T.; Kumai, T.; Nagato, T.; Nozaki, Y.; Ishibashi, K.; Kosaka, A.; Nagata, M.; Harabuchi, S.; Ohara, M.; et al. Targeting phosphorylated p53 to elicit tumor-reactive T helper responses against head and neck squamous cell carcinoma. Oncoimmunology 2018, 7, e1466771. [Google Scholar] [CrossRef] [PubMed]
- Maggioni, D.; Pignataro, L.; Garavello, W. T-helper and T-regulatory cells modulation in head and neck squamous cell carcinoma. Oncoimmunology 2017, 6, e1325066. [Google Scholar] [CrossRef]
- Mironska, A.; Łukaszewicz-Zajac, M.; Mroczko, B. Clinical Significance of Selected Chemokines in Thyroid Cancer. Anticancer Res. 2019, 39, 2715–2720. [Google Scholar] [CrossRef]
- Khurshid, Z.; Zafar, M.S.; Khan, R.S.; Najeeb, S.; Slowey, P.D.; Rehman, I.U. Role of Salivary Biomarkers in Oral Cancer Detection. Adv. Clin. Chem. 2018, 86, 23–70. [Google Scholar]
- Riley, P.; Glenny, A.M.; Worthington, H.V.; Littlewood, A.; Fernandez Mauleffinch, L.M.; Clarkson, J.E.; McCabe, M.G. Interventions for preventing oral mucositis in patients with cancer receiving treatment: Cytokines and growth factors. Cochrane Database Syst. Rev. 2017, 11, Cd011990. [Google Scholar] [CrossRef]
- Muderris, T.K.; Gul, F.; Doblan, A.; Ergin, M.; Muderris, T. Role of T-helper 17 cell related cytokines in laryngeal cancer. J. Laryngol. Otol. 2019, 133, 394–398. [Google Scholar] [CrossRef]
- Sotirović, J.; Perić, A.; Vojvodić, D.; Baletić, N.; Zaletel, I.; Stanojević, I.; Erdoglija, M.; Milojević, M. Serum cytokine profile of laryngeal squamous cell carcinoma patients. J. Laryngol. Otol. 2017, 131, 455–461. [Google Scholar] [CrossRef]
- Miller, J.S.; Morishima, C.; McNeel, D.G.; Patel, M.R.; Kohrt, H.E.K.; Thompson, J.A.; Sondel, P.M.; Wakelee, H.A.; Disis, M.L.; Kaiser, J.C.; et al. A First-in-Human Phase I Study of Subcutaneous Outpatient Recombinant Human IL15 (rhIL15) in Adults with Advanced Solid Tumors. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2018, 24, 1525–1535. [Google Scholar] [CrossRef]
- Ishikawa, K.; Yagi-Nakanishi, S.; Nakanishi, Y.; Kondo, S.; Tsuji, A.; Endo, K.; Wakisaka, N.; Murono, S.; Yoshizaki, T. Expression of interleukin-33 is correlated with poor prognosis of patients with squamous cell carcinoma of the tongue. AurisNasusLarynx 2014, 41, 552–557. [Google Scholar] [CrossRef]
- Wen, Y.H.; Lin, H.Q.; Li, H.; Zhao, Y.; Lui, V.W.Y.; Chen, L.; Wu, X.M.; Sun, W.; Wen, W.P. Stromal interleukin-33 promotes regulatory T cell-mediated immunosuppression in head and neck squamous cell carcinoma and correlates with poor prognosis. Cancer Immunol. Immunother. 2019, 68, 221–232. [Google Scholar] [CrossRef]
- St John, M.A. Inflammatory mediators drive metastasis and drug resistance in head and neck squamous cell carcinoma. Laryngoscope 2015, 125 (Suppl. 3), S1–S11. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, C.; Zhang, C.; Li, Z.; Zhu, T.; Chen, J.; Ren, Y.; Wang, X.; Zhang, L.; Zhou, X. TGF-β-induced STAT3 overexpression promotes human head and neck squamous cell carcinoma invasion and metastasis through malat1/miR-30a interactions. Cancer Lett. 2018, 436, 52–62. [Google Scholar] [CrossRef]
- Li, X.; Xu, Q.; Wu, Y.; Li, J.; Tang, D.; Han, L.; Fan, Q. A CCL2/ROS autoregulation loop is critical for cancer-associated fibroblasts-enhanced tumor growth of oral squamous cell carcinoma. Carcinogenesis 2014, 35, 1362–1370. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.A.; Ribeiro, F.L.; Oliveira-Neto, H.H.; Watanabe, S.; Alencar Rde, C.; Fukada, S.Y.; Cunha, F.Q.; Leles, C.R.; Mendonça, E.F.; Batista, A.C. Dual role of CCL3/CCR1 in oral squamous cell carcinoma: Implications in tumor metastasis and local host defense. Oncol. Rep. 2007, 18, 1107–1113. [Google Scholar] [PubMed]
- Chuang, J.Y.; Yang, W.H.; Chen, H.T.; Huang, C.Y.; Tan, T.W.; Lin, Y.T.; Hsu, C.J.; Fong, Y.C.; Tang, C.H. CCL5/CCR5 axis promotes the motility of human oral cancer cells. J. Cell. Physiol. 2009, 220, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Jung, D.W.; Che, Z.M.; Kim, J.; Kim, K.; Kim, K.Y.; Williams, D.; Kim, J. Tumor-stromal crosstalk in invasion of oral squamous cell carcinoma: A pivotal role of CCL7. Int. J. Cancer 2010, 127, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.P.; Kao, H.K.; Yen, T.C.; Chang, Y.L.; Liang, Y.; Liu, S.C.; Lee, L.Y.; Chang, Y.L.; Kang, C.J.; Chen, I.H.; et al. Overexpression of macrophage inflammatory protein-3α in oral cavity squamous cell carcinoma is associated with nodal metastasis. Oral Oncol. 2011, 47, 108–113. [Google Scholar] [CrossRef]
- Shang, Z.J.; Liu, K.; Shao, Z. Expression of chemokine receptor CCR7 is associated with cervical lymph node metastasis of oral squamous cell carcinoma. Oral Oncol. 2009, 45, 480–485. [Google Scholar] [CrossRef]
- Li, P.; Zhao, Z.J.; Liu, F.Y.; Sun, L.Y.; Ding, X.; Zhang, W.Z.; Shang, D.H.; Sun, C.F. The chemokine receptor 7 regulates cell adhesion and migration via beta1 integrin in metastatic squamous cell carcinoma of the head and neck. Oncol. Rep. 2010, 24, 989–995. [Google Scholar]
- Guo, N.; Liu, F.; Yang, L.; Huang, J.; Ding, X.; Sun, C. Chemokine receptor 7 enhances cell chemotaxis and migration of metastatic squamous cell carcinoma of head and neck through activation of matrix metalloproteinase-9. Oncol. Rep. 2014, 32, 794–800. [Google Scholar] [CrossRef][Green Version]
- Liu, F.Y.; Safdar, J.; Li, Z.N.; Fang, Q.G.; Zhang, X.; Xu, Z.F.; Sun, C.F. CCR7 regulates cell migration and invasion through MAPKs in metastatic squamous cell carcinoma of head and neck. Int. J. Oncol. 2014, 45, 2502–2510. [Google Scholar] [CrossRef] [PubMed]
- Irino, T.; Takeuchi, H.; Matsuda, S.; Saikawa, Y.; Kawakubo, H.; Wada, N.; Takahashi, T.; Nakamura, R.; Fukuda, K.; Omori, T.; et al. CC-Chemokine receptor CCR7: A key molecule for lymph node metastasis in esophageal squamous cell carcinoma. BMC Cancer 2014, 14, 291. [Google Scholar] [CrossRef]
- Tsuzuki, H.; Takahashi, N.; Kojima, A.; Narita, N.; Sunaga, H.; Takabayashi, T.; Fujieda, S. Oral and oropharyngeal squamous cell carcinomas expressing CCR7 have poor prognoses. AurisNasusLarynx 2006, 33, 37–42. [Google Scholar] [CrossRef]
- Lee, C.H.; Syu, S.H.; Liu, K.J.; Chu, P.Y.; Yang, W.C.; Lin, P.; Shieh, W.Y. Interleukin-1 beta transactivates epidermal growth factor receptor via the CXCL1-CXCR2 axis in oral cancer. Oncotarget 2015, 6, 38866–38880. [Google Scholar] [CrossRef]
- Campbell, L.M.; Maxwell, P.J.; Waugh, D.J. Rationale and Means to Target Pro-Inflammatory Interleukin-8 (CXCL8) Signaling in Cancer. Pharm 2013, 6, 929–959. [Google Scholar] [CrossRef] [PubMed]
- Khurram, S.A.; Bingle, L.; McCabe, B.M.; Farthing, P.M.; Whawell, S.A. The chemokine receptors CXCR1 and CXCR2 regulate oral cancer cell behaviour. J. Oral Pathol. Med. Off. Publ. Int. Assoc. Oral Pathol. Am. Acad. Oral Pathol. 2014, 43, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Yeh, C.J.; Yu, C.C.; Chou, M.Y.; Lin, C.H.; Wei, L.H.; Lin, C.W.; Yang, S.F.; Chien, M.H. Impact of interleukin-8 gene polymorphisms and environmental factors on oral cancer susceptibility in Taiwan. Oral Dis. 2012, 18, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.P.; Wu, C.C.; Fang, K.H.; Tsai, C.Y.; Chang, Y.L.; Liu, S.C.; Kao, H.K. Serum levels of chemokine (C-X-C motif) ligand 9 (CXCL9) are associated with tumor progression and treatment outcome in patients with oral cavity squamous cell carcinoma. Oral Oncol. 2013, 49, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Kucia, M.; Jankowski, K.; Reca, R.; Wysoczynski, M.; Bandura, L.; Allendorf, D.J.; Zhang, J.; Ratajczak, J.; Ratajczak, M.Z. CXCR4-SDF-1 signalling, locomotion, chemotaxis and adhesion. J. Mol. Histol. 2004, 35, 233–245. [Google Scholar] [CrossRef]
- Xia, J.; Chen, N.; Hong, Y.; Chen, X.; Tao, X.; Cheng, B.; Huang, Y. Expressions of CXCL12/CXCR4 in oral premalignant and malignant lesions. Mediat. Inflamm. 2012, 2012, 516395. [Google Scholar] [CrossRef]
- Uchida, D.; Begum, N.M.; Almofti, A.; Nakashiro, K.; Kawamata, H.; Tateishi, Y.; Hamakawa, H.; Yoshida, H.; Sato, M. Possible role of stromal-cell-derived factor-1/CXCR4 signaling on lymph node metastasis of oral squamous cell carcinoma. Exp. Cell Res. 2003, 290, 289–302. [Google Scholar] [CrossRef]
- León, X.; Diez, S.; García, J.; Lop, J.; Sumarroca, A.; Quer, M.; Camacho, M. Expression of the CXCL12/CXCR4 chemokine axis predicts regional control in head and neck squamous cell carcinoma. Eur. Arch. Oto-Rhino-Laryngol. Off. J. Eur. Fed. Oto-Rhino-Laryngol. Soc. (Eufos) Affil. Ger. Soc. Oto-Rhino-Laryngol. Head Neck Surg. 2016, 273, 4525–4533. [Google Scholar] [CrossRef]
- Teng, Y.H.; Liu, T.H.; Tseng, H.C.; Chung, T.T.; Yeh, C.M.; Li, Y.C.; Ou, Y.H.; Lin, L.Y.; Tsai, H.T.; Yang, S.F. Contribution of genetic polymorphisms of stromal cell-derived factor-1 and its receptor, CXCR4, to the susceptibility and clinicopathologic development of oral cancer. Head Neck 2009, 31, 1282–1288. [Google Scholar] [CrossRef]
- Khurram, S.A.; Whawell, S.A.; Bingle, L.; Murdoch, C.; McCabe, B.M.; Farthing, P.M. Functional expression of the chemokine receptor XCR1 on oral epithelial cells. J. Pathol. 2010, 221, 153–163. [Google Scholar] [CrossRef]
- Luangdilok, S.; Box, C.; Harrington, K.; Rhŷs-Evans, P.; Eccles, S. MAPK and PI3K signalling differentially regulate angiogenic and lymphangiogenic cytokine secretion in squamous cell carcinoma of the head and neck. Eur. J. Cancer 2011, 47, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Stanam, A.; Gibson-Corley, K.N.; Love-Homan, L.; Ihejirika, N.; Simons, A.L. Interleukin-1 blockade overcomes erlotinib resistance in head and neck squamous cell carcinoma. Oncotarget 2016, 7, 76087–76100. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, N.; Kurata, M.; Yamamoto, T.; Morikawa, S.; Masumoto, J. The role of interleukin-1 in general pathology. Inflamm. Regen. 2019, 39, 12. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Chang, C.Y.; Kuo, Y.Z.; Fang, W.Y.; Kao, H.Y.; Tsai, S.T.; Wu, L.W. Cancer-associated fibroblast-derived interleukin-1beta activates protumor C-C motif chemokine ligand 22 signaling in head and neck cancer. Cancer Sci. 2019, 110, 2783–2793. [Google Scholar] [CrossRef]
- Yang, H.T.; Cohen, P.; Rousseau, S. IL-1beta-stimulated activation of ERK1/2 and p38alpha MAPK mediates the transcriptional up-regulation of IL-6, IL-8 and GRO-alpha in HeLa cells. Cell Signal. 2008, 20, 375–380. [Google Scholar] [CrossRef]
- Lee, M.K.; Park, J.H.; Gi, S.H.; Hwang, Y.S. IL-1β Induces Fascin Expression and Increases Cancer Invasion. Anticancer Res. 2018, 38, 6127–6132. [Google Scholar] [CrossRef]
- Wu, T.; Hong, Y.; Jia, L.; Wu, J.; Xia, J.; Wang, J.; Hu, Q.; Cheng, B. Modulation of IL-1β reprogrammes the tumor microenvironment to interrupt oral carcinogenesis. Sci. Rep. 2016, 6, 20208. [Google Scholar] [CrossRef]
- Suzuki, A.; Leland, P.; Joshi, B.H.; Puri, R.K. Targeting of IL-4 and IL-13 receptors for cancer therapy. Cytokine 2015, 75, 79–88. [Google Scholar] [CrossRef]
- Chonov, D.C.; Ignatova, M.M.K.; Ananiev, J.R.; Gulubova, M.V. IL-6 Activities in the Tumour Microenvironment. Part 1. Open Access Maced. J. Med. Sci. 2019, 7, 2391–2398. [Google Scholar] [CrossRef]
- Shinriki, S.; Jono, H.; Ota, K.; Ueda, M.; Kudo, M.; Ota, T.; Oike, Y.; Endo, M.; Ibusuki, M.; Hiraki, A.; et al. Humanized anti-interleukin-6 receptor antibody suppresses tumor angiogenesis and in vivo growth of human oral squamous cell carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2009, 15, 5426–5434. [Google Scholar] [CrossRef]
- Chan, L.P.; Wang, L.F.; Chiang, F.Y.; Lee, K.W.; Kuo, P.L.; Liang, C.H. IL-8 promotes HNSCC progression on CXCR1/2-meidated NOD1/RIP2 signaling pathway. Oncotarget 2016, 7, 61820–61831. [Google Scholar] [CrossRef]
- Bilusic, M.; Heery, C.R.; Collins, J.M.; Donahue, R.N.; Palena, C.; Madan, R.A.; Karzai, F.; Marté, J.L.; Strauss, J.; Gatti-Mays, M.E.; et al. Phase I trial of HuMax-IL8 (BMS-986253), an anti-IL-8 monoclonal antibody, in patients with metastatic or unresectable solid tumors. J. Immunother. Cancer 2019, 7, 240. [Google Scholar] [CrossRef] [PubMed]
- Aziz, S.; Ahmed, S.S.; Ali, A.; Khan, F.A.; Zulfiqar, G.; Iqbal, J.; Khan, A.A.; Shoaib, M. Salivary Immunosuppressive Cytokines IL-10 and IL-13 Are Significantly Elevated in Oral Squamous Cell Carcinoma Patients. Cancer Investig. 2015, 33, 318–328. [Google Scholar] [CrossRef]
- Takahashi, H.; Sakakura, K.; Kudo, T.; Toyoda, M.; Kaira, K.; Oyama, T.; Chikamatsu, K. Cancer-associated fibroblasts promote an immunosuppressive microenvironment through the induction and accumulation of protumoral macrophages. Oncotarget 2017, 8, 8633–8647. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.Y.; Tsai, T.H.; Yang, J.L.; Li, L.C. Therapeutic Strategies for Targeting IL-33/ST2 Signalling for the Treatment of Inflammatory Diseases. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 49, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, T.; Pollard, J.W. Therapeutic potential of chemokine signal inhibition for metastatic breast cancer. Pharmacol. Res. 2015, 100, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Thompson, W.L.; Van Eldik, L.J. Inflammatory cytokines stimulate the chemokines CCL2/MCP-1 and CCL7/MCP-3 through NFkB and MAPK dependent pathways in rat astrocytes [corrected]. Brain Res. 2009, 1287, 47–57. [Google Scholar] [CrossRef]
- Ranasinghe, R.; Eri, R. Modulation of the CCR6-CCL20 Axis: A Potential Therapeutic Target in Inflammation and Cancer. Medicina 2018, 54, 88. [Google Scholar] [CrossRef]
- Dufies, M.; Grytsai, O.; Ronco, C.; Camara, O.; Ambrosetti, D.; Hagege, A.; Parola, J.; Mateo, L.; Ayrault, M.; Giuliano, S.; et al. New CXCR1/CXCR2 inhibitors represent an effective treatment for kidney or head and neck cancers sensitive or refractory to reference treatments. Theranostics 2019, 9, 5332–5346. [Google Scholar] [CrossRef]
- Liu, Q.; Li, A.; Tian, Y.; Wu, J.D.; Liu, Y.; Li, T.; Chen, Y.; Han, X.; Wu, K. The CXCL8-CXCR1/2 pathways in cancer. Cytokine Growth Factor Rev. 2016, 31, 61–71. [Google Scholar] [CrossRef]
- Gladue, R.P.; Brown, M.F.; Zwillich, S.H. CCR1 antagonists: What have we learned from clinical trials. Curr. Top. Med. Chem. 2010, 10, 1268–1277. [Google Scholar] [CrossRef]
- Huang, S.; Mills, L.; Mian, B.; Tellez, C.; McCarty, M.; Yang, X.D.; Gudas, J.M.; Bar-Eli, M. Fully humanized neutralizing antibodies to interleukin-8 (ABX-IL8) inhibit angiogenesis, tumor growth, and metastasis of human melanoma. Am. J. Pathol. 2002, 161, 125–134. [Google Scholar] [CrossRef]
- Yan, W.L.; Shen, K.Y.; Tien, C.Y.; Chen, Y.A.; Liu, S.J. Recent progress in GM-CSF-based cancer immunotherapy. Immunotherapy 2017, 9, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Akl, M.R.; Nagpal, P.; Ayoub, N.M.; Tai, B.; Prabhu, S.A.; Capac, C.M.; Gliksman, M.; Goy, A.; Suh, K.S. Molecular and clinical significance of fibroblast growth factor 2 (FGF2 /bFGF) in malignancies of solid and hematological cancers for personalized therapies. Oncotarget 2016, 7, 44735–44762. [Google Scholar] [CrossRef]
- Bussu, F.; Graziani, C.; Gallus, R.; Cittadini, A.; Galli, J.; De Corso, E.; di Cintio, G.; Corbi, M.; Almadori, G.; Boninsegna, A.; et al. IFN-γ and other serum cytokines in head and neck squamous cell carcinomas. Acta Otorhinolaryngol. Ital. Organo Uff. Della Soc. Ital. Di Otorinolaringol. E Chir. Cervico-Facciale 2018, 38, 94–102. [Google Scholar]
- Jorgovanovic, D.; Song, M.; Wang, L.; Zhang, Y. Roles of IFN-γ in tumor progression and regression: A review. Biomark. Res. 2020, 8, 49. [Google Scholar] [CrossRef]
- Pang, X.; Tang, Y.L.; Liang, X.H. Transforming growth factor-β signaling in head and neck squamous cell carcinoma: Insights into cellular responses. Oncol. Lett. 2018, 16, 4799–4806. [Google Scholar] [CrossRef]
- Bedi, A.; Chang, X.; Noonan, K.; Pham, V.; Bedi, R.; Fertig, E.J.; Considine, M.; Califano, J.A.; Borrello, I.; Chung, C.H.; et al. Inhibition of TGF-β enhances the in vivo antitumor efficacy of EGF receptor-targeted therapy. Mol. Cancer Ther. 2012, 11, 2429–2439. [Google Scholar] [CrossRef]
- Selimovic, D.; Wahl, R.U.; Ruiz, E.; Aslam, R.; Flanagan, T.W.; Hassan, S.Y.; Santourlidis, S.; Haikel, Y.; Friedlander, P.; Megahed, M.; et al. Tumor necrosis factor-α triggers opposing signals in head and neck squamous cell carcinoma and induces apoptosis via mitochondrial- and non-mitochondrial-dependent pathways. Int. J. Oncol. 2019, 55, 1324–1338. [Google Scholar] [CrossRef]
- Wang, X.; Lin, Y. Tumor necrosis factor and cancer, buddies or foes? Acta Pharmacol. Sin. 2008, 29, 1275–1288. [Google Scholar] [CrossRef]
- Micaily, I.; Johnson, J.; Argiris, A. An update on angiogenesis targeting in head and neck squamous cell carcinoma. Cancers Head Neck 2020, 5, 5. [Google Scholar] [CrossRef]
- Umemura, N.; Zhu, J.; Mburu, Y.K.; Forero, A.; Hsieh, P.N.; Muthuswamy, R.; Kalinski, P.; Ferris, R.L.; Sarkar, S.N. Defective NF-κB signaling in metastatic head and neck cancer cells leads to enhanced apoptosis by double-stranded RNA. Cancer Res. 2012, 7, 45–55. [Google Scholar] [CrossRef]
- Brown, M.; Cohen, J.; Arun, P.; Chen, Z.; Van Waes, C. NF-kappaB in carcinoma therapy and prevention. Expert Opin. Ther. Targets 2008, 12, 1109–1122. [Google Scholar] [CrossRef]
- Sen, B.; Peng, S.; Woods, D.M.; Wistuba, I.; Bell, D.; El-Naggar, A.K.; Lai, S.Y.; Johnson, F.M. STAT5A-mediated SOCS2 expression regulates Jak2 and STAT3 activity following c-Src inhibition in head and neck squamous carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 127–139. [Google Scholar] [CrossRef]
- Lee, C.H.; Chang, J.S.; Syu, S.H.; Wong, T.S.; Chan, J.Y.; Tang, Y.C.; Yang, Z.P.; Yang, W.C.; Chen, C.T.; Lu, S.C.; et al. IL-1β promotes malignant transformation and tumor aggressiveness in oral cancer. J. Cell. Physiol. 2015, 230, 875–884. [Google Scholar] [CrossRef]
- León, X.; Bothe, C.; García, J.; Parreño, M.; Alcolea, S.; Quer, M.; Vila, L.; Camacho, M. Expression of IL-1α correlates with distant metastasis in patients with head and neck squamous cell carcinoma. Oncotarget 2015, 6, 37398–37409. [Google Scholar] [CrossRef]
- Kim, N.; Ryu, H.; Kim, S.; Joo, M.; Jeon, H.J.; Lee, M.W.; Song, I.C.; Kim, M.N.; Kim, J.M.; Lee, H.J. CXCR7 promotes migration and invasion in head and neck squamous cell carcinoma by upregulating TGF-beta1/Smad2/3 signaling. Sci. Rep. 2019, 9, 18100. [Google Scholar] [CrossRef] [PubMed]
- Freudlsperger, C.; Bian, Y.; Contag Wise, S.; Burnett, J.; Coupar, J.; Yang, X.; Chen, Z.; Van Waes, C. TGF-β and NF-κB signal pathway cross-talk is mediated through TAK1 and SMAD7 in a subset of head and neck cancers. Oncogene 2013, 32, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Miao, C.; Liu, W.; Qiao, X.; Yang, W.; Li, L.; Li, C. TGF-β1/TβRII/Smad3 signaling pathway promotes VEGF expression in oral squamous cell carcinoma tumor-associated macrophages. Biochem. Biophys. Res. Commun. 2018, 497, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liang, L.; Liu, X.; Wu, J.N.; Chen, J.; Su, K.; Zheng, Q.; Huang, H.; Liao, G.Q. TGFβ1-Smad3-Jagged1-Notch1-Slug signaling pathway takes part in tumorigenesis and progress of tongue squamous cell carcinoma. J. Oral Pathol. Med. Off. Publ. Int. Assoc. Oral Pathol. Am. Acad. Oral Pathol. 2016, 45, 486–493. [Google Scholar] [CrossRef]
- Wei, T.; Cong, X.; Wang, X.T.; Xu, X.J.; Min, S.N.; Ye, P.; Peng, X.; Wu, L.L.; Yu, G.Y. Interleukin-17A promotes tongue squamous cell carcinoma metastasis through activating miR-23b/versican pathway. Oncotarget 2017, 8, 6663–6680. [Google Scholar] [CrossRef] [PubMed]
- Bragado, P.; Estrada, Y.; Parikh, F.; Krause, S.; Capobianco, C.; Farina, H.G.; Schewe, D.M.; Aguirre-Ghiso, J.A. TGF-β2 dictates disseminated tumour cell fate in target organs through TGF-β-RIII and p38α/β signalling. Nat. Cell Biol. 2013, 15, 1351–1361. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.M.; France, T.J.; Teknos, T.N.; Kumar, P. Interleukin-6 role in head and neck squamous cell carcinoma progression. World J. Otorhinolaryngol. Head Neck Surg. 2016, 2, 90–97. [Google Scholar] [CrossRef]
- Kim, M.S.; Lee, W.S.; Jeong, J.; Kim, S.J.; Jin, W. Induction of metastatic potential by TrkB via activation of IL6/JAK2/STAT3 and PI3K/AKT signaling in breast cancer. Oncotarget 2015, 6, 40158–40171. [Google Scholar] [CrossRef] [PubMed]
- Linnskog, R.; Jönsson, G.; Axelsson, L.; Prasad, C.P.; Andersson, T. Interleukin-6 drives melanoma cell motility through p38α-MAPK-dependent up-regulation of WNT5A expression. Mol. Oncol. 2014, 8, 1365–1378. [Google Scholar] [CrossRef]
- Yadav, A.; Kumar, B.; Datta, J.; Teknos, T.N.; Kumar, P. IL-6 promotes head and neck tumor metastasis by inducing epithelial-mesenchymal transition via the JAK-STAT3-SNAIL signaling pathway. Mol. Cancer Res. 2011, 9, 1658–1667. [Google Scholar] [CrossRef]
- Lesinski, G.B.; Nannapaneni, S.; Griffith, C.C.; Patel, M.; Chen, W.; Chen, Z.; Ahmed, R.; Wieland, A.; Shin, D.M.; Chen, Z.G.; et al. Interleukin-6/STAT3 Signaling is Prominent and Associated with Reduced Overall Survival in p16 Negative Oropharyngeal Squamous Cell Carcinoma. Head Neck Pathol. 2019, 13, 304–312. [Google Scholar] [CrossRef]
- Bu, L.L.; Yu, G.T.; Wu, L.; Mao, L.; Deng, W.W.; Liu, J.F.; Kulkarni, A.B.; Zhang, W.F.; Zhang, L.; Sun, Z.J. STAT3 Induces Immunosuppression by Upregulating PD-1/PD-L1 in HNSCC. J. Dent. Res. 2017, 96, 1027–1034. [Google Scholar] [CrossRef]
- Karakasheva, T.A.; Lin, E.W.; Tang, Q.; Qiao, E.; Waldron, T.J.; Soni, M.; Klein-Szanto, A.J.; Sahu, V.; Basu, D.; Ohashi, S.; et al. IL-6 Mediates Cross-Talk between Tumor Cells and Activated Fibroblasts in the Tumor Microenvironment. Cancer Res. 2018, 78, 4957–4970. [Google Scholar] [CrossRef]
- Ma, H.; Yang, W.; Zhang, L.; Liu, S.; Zhao, M.; Zhou, G.; Wang, L.; Jin, S.; Zhang, Z.; Hu, J. Interferon-alpha promotes immunosuppression through IFNAR1/STAT1 signalling in head and neck squamous cell carcinoma. Br. J. Cancer 2019, 120, 317–330. [Google Scholar] [CrossRef]
- Ma, H.; Jin, S.; Yang, W.; Zhou, G.; Zhao, M.; Fang, S.; Zhang, Z.; Hu, J. Interferon-alpha enhances the antitumour activity of EGFR-targeted therapies by upregulating RIG-I in head and neck squamous cell carcinoma. Br. J. Cancer 2018, 118, 509–521. [Google Scholar] [CrossRef]
- Koul, H.K.; Pal, M.; Koul, S. Role of p38 MAP Kinase Signal Transduction in Solid Tumors. Genes Cancer 2013, 4, 342–359. [Google Scholar] [CrossRef]
- Chan, L.P.; Liu, C.; Chiang, F.Y.; Wang, L.F.; Lee, K.W.; Chen, W.T.; Kuo, P.L.; Liang, C.H. IL-8 promotes inflammatory mediators and stimulates activation of p38 MAPK/ERK-NF-κB pathway and reduction of JNK in HNSCC. Oncotarget 2017, 8, 56375–56388. [Google Scholar] [CrossRef]
- Muller, A.; Sonkoly, E.; Eulert, C.; Gerber, P.A.; Kubitza, R.; Schirlau, K.; Franken-Kunkel, P.; Poremba, C.; Snyderman, C.; Klotz, L.O.; et al. Chemokine receptors in head and neck cancer: Association with metastatic spread and regulation during chemotherapy. Int. J. Cancer 2006, 118, 2147–2157. [Google Scholar] [CrossRef]
- Yoshimura, T. The chemokine MCP-1 (CCL2) in the host interaction with cancer: A foe or ally? Cell. Mol. Immunol. 2018, 15, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Ji, W.T.; Chen, H.R.; Lin, C.H.; Lee, J.W.; Lee, C.C. Monocyte chemotactic protein 1 (MCP-1) modulates pro-survival signaling to promote progression of head and neck squamous cell carcinoma. PLoS ONE 2014, 9, e88952. [Google Scholar] [CrossRef] [PubMed]
- Marcuzzi, E.; Angioni, R.; Molon, B.; Calì, B. Chemokines and Chemokine Receptors: Orchestrating Tumor Metastasization. Int. J. Mol. Sci. 2018, 20, 96. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.W.; Kim, N.; Kim, S.; Joo, M.; Ryu, H.; Lee, H.J. Chemokine receptor CXCR7 expression, function and clinical implications in head and neck squamous cell carcinoma. Ann. Oncol. 2018, 29, viii680–viii681. [Google Scholar] [CrossRef]
- Mburu, Y.K.; Egloff, A.M.; Walker, W.H.; Wang, L.; Seethala, R.R.; van Waes, C.; Ferris, R.L. Chemokine receptor 7 (CCR7) gene expression is regulated by NF-κB and activator protein 1 (AP1) in metastatic squamous cell carcinoma of head and neck (SCCHN). J. Biol. Chem. 2012, 287, 3581–3590. [Google Scholar] [CrossRef]
- Gao, L.; Wang, F.-Q.; Li, H.-M.; Yang, J.-G.; Ren, J.-G.; He, K.-F.; Liu, B.; Zhang, W.; Zhao, Y.-F. CCL2/EGF positive feedback loop between cancer cells and macrophages promotes cell migration and invasion in head and neck squamous cell carcinoma. Oncotarget 2016, 7, 87037–87051. [Google Scholar] [CrossRef]
- Samara, G.J.; Lawrence, D.M.; Chiarelli, C.J.; Valentino, M.D.; Lyubsky, S.; Zucker, S.; Vaday, G.G. CXCR4-mediated adhesion and MMP-9 secretion in head and neck squamous cell carcinoma. Cancer Lett. 2004, 214, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.-T.; Chu, C.-Y.; Lu, Y.-C.; Chang, C.-C.; Lin, B.-R.; Wu, H.-H.; Liu, H.-L.; Cha, S.-T.; Prakash, E.; Ko, J.-Y.; et al. CXCL12/CXCR4 promotes laryngeal and hypopharyngeal squamous cell carcinoma metastasis through MMP-13-dependent invasion via the ERK1/2/AP-1 pathway. Carcinogenesis 2008, 29, 1519–1527. [Google Scholar] [CrossRef] [PubMed]
- Faber, A.; Goessler, U.R.; Hoermann, K.; Schultz, J.D.; Umbreit, C.; Stern-Straeter, J. SDF-1-CXCR4 axis: Cell trafficking in the cancer stem cell niche of head and neck squamous cell carcinoma. Oncol. Rep. 2013, 29, 2325–2331. [Google Scholar] [CrossRef] [PubMed]
- De-Colle, C.; Mönnich, D.; Welz, S.; Boeke, S.; Sipos, B.; Fend, F.; Mauz, P.-S.; Tinhofer, I.; Budach, V.; Jawad, J.A.; et al. SDF-1/CXCR4 expression in head and neck cancer and outcome after postoperative radiochemotherapy. Clin. Transl Radiat Oncol. 2017, 5, 28–36. [Google Scholar] [CrossRef]
- Miyazaki, H.; Patel, V.; Wang, H.; Edmunds, R.K.; Gutkind, J.S.; Yeudall, W.A. Down-regulation of CXCL5 Inhibits Squamous Carcinogenesis. Cancer Res. 2006, 66, 4279. [Google Scholar] [CrossRef] [PubMed]
- Hsin, L.-J.; Kao, H.-K.; Chen, I.H.; Tsang, N.-M.; Hsu, C.-L.; Liu, S.-C.; Chang, Y.-S.; Chang, K.-P. Serum CXCL9 Levels Are Associated with Tumor Progression and Treatment Outcome in Patients with Nasopharyngeal Carcinoma. PLoS ONE 2013, 8, e80052. [Google Scholar] [CrossRef]
- Lin, Y.-C. Cancer-associated fibroblast induced chemokine (C-C motif) ligand 11 contribute to the progression of head and neck cancer. In Joint Event on 33rd International Conference on Oncology Nursing and Cancer Care and 16th Asia Pacific Pathology Congress, Tokyo, Japan, 2018; e-Century Publishing Corporatio: Tokyo, Japan, 2018. [Google Scholar]
- Chen, S.-F.; Nieh, S.; Jao, S.-W.; Wu, M.-Z.; Liu, C.-L.; Chang, Y.-C.; Lin, Y.-S. The paracrine effect of cancer-associated fibroblast-induced interleukin-33 regulates the invasiveness of head and neck squamous cell carcinoma. J. Pathol. 2013, 231, 180–189. [Google Scholar] [CrossRef]
- Xu, Q.; Ma, H.; Chang, H.; Feng, Z.; Zhang, C.; Yang, X. The interaction of interleukin-8 and PTEN inactivation promotes the malignant progression of head and neck squamous cell carcinoma via the STAT3 pathway. Cell Death Dis 2020, 11, 405. [Google Scholar] [CrossRef]
- Dumitru, C.A.; Gholaman, H.; Trellakis, S.; Bruderek, K.; Dominas, N.; Gu, X.; Bankfalvi, A.; Whiteside, T.L.; Lang, S.; Brandau, S. Tumor-derived macrophage migration inhibitory factor modulates the biology of head and neck cancer cells via neutrophil activation. Int. J. Cancer 2011, 129, 859–869. [Google Scholar] [CrossRef]
- DiNatale, B.C.; Smith, K.; John, K.; Krishnegowda, G.; Amin, S.G.; Perdew, G.H. Ah Receptor Antagonism Represses Head and Neck Tumor Cell Aggressive Phenotype. Mol. Cancer Res. 2012, 10, 1369–1379. [Google Scholar] [CrossRef]
- White, R.A.; Malkoski, S.P.; Wang, X.J. TGFβ signaling in head and neck squamous cell carcinoma. Oncogene 2010, 29, 5437–5446. [Google Scholar] [CrossRef] [PubMed]
- Muro-Cacho, C.A.; Anderson, M.; Cordero, J.; Munoz-Antonia, T. Expression of transforming growth factor beta type II receptors in head and neck squamous cell carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 1999, 5, 1243–1248. [Google Scholar]
- Principe, S.; Dikova, V.; Bagán, J. Salivary Cytokines in patients with Head and Neck Cancer (HNC) treated with Radiotherapy. J. Clin. Exp. Dent. 2019, 11, e1072–e1077. [Google Scholar] [CrossRef] [PubMed]
- Bigbee, W.L.; Grandis, J.R.; Siegfried, J.M. Multiple cytokine and growth factor serum biomarkers predict therapeutic response and survival in advanced-stage head and neck cancer patients. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2007, 13, 3107–3108. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wolf, G.T.; Moyer, J.S.; Kaplan, M.J.; Newman, J.G.; Egan, J.E.; Berinstein, N.L.; Whiteside, T.L. IRX-2 natural cytokine biologic for immunotherapy in patients with head and neck cancers. Onco Targets 2018, 11, 3731–3746. [Google Scholar] [CrossRef]
- Chulpanova, D.S.; Kitaeva, K.V.; Green, A.R.; Rizvanov, A.A.; Solovyeva, V.V. Molecular Aspects and Future Perspectives of Cytokine-Based Anti-cancer Immunotherapy. Front. Cell Dev. Biol. 2020, 8, 402. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| S.No | Chemokine/Cytokine | Expression | Signaling Pathway(s) Activated | Interacts with Cells | Involved in | Targeted OR Can Be Targeted by | References |
|---|---|---|---|---|---|---|---|
| 1 | IL-1 | Up | MAPK/ERK1/2, NF-KB, PI3K/AKT, & JAK/STAT | Tumor, T-cells, TAMs & macrophages | Recruitment of TAMs, MDSCs and Tregs | Recombinant IL-1R antagonist (anakinra) | [151,152] |
| 2 | IL-1β | Up | NF-kB, ERK1/2, JNK, CREB | Endothelial & leukocytes | Increase integrin expression | Lenti virus mediated shRNA | [153,154,155,156,157] |
| 3 | IL-4 | Up | MAPK | Tumor, TAMs & endothelial cells | Promote tumor growth, angiogenesis & immuno-suppression | rIL-4 with Pseudomonas exotoxin (PE) targeting IL-4R | [158] |
| 4 | IL-6 | Up | JAK/STAT, PI3K/AKT, RAS/RAF/MEK/ERK1/2 & Wnt | Tumor, Th17 cells & CAFs | Support tumor growth, immune evasion & CAF activation | Humanized antiIL-6R antibody (Tocilizumab) | [159,160] |
| 5 | IL-8 | Up | NF-kB, MAPK, CXCR1/2/NOD1/RIP2 | Tumor, endothelial cells & neutrophils | Promotes tumor invasion, angiogenesis & recruit neutrophils to the TME | Humanized anti- IL-8 antibody (HuMax-IL8) | [161,162] |
| 6 | IL-10 | Up | JAK/STAT3 | T cells, TAMs | Suppress T-cell proliferation and promote immuno-suppression | - | [163,164] |
| 7 | IL-15 | Up | JAK/STAT3, PI3K/AKT | CD8+ T cell & NK cell | Stimulate NK and CD8+ T cell function | Recombinant human IL-15 (rhIL15) | [124] |
| 8 | IL-33 | Up | RAS/RAF//MEK/ERK1/2 and JNK | Tregs | Enrich FOXP3+ Tregs | Anti-IL-33 antibodies | [125,129,165] |
| 9 | CCL-2 | Up | MAPK, PI3K/AKT | Monocytes, TAMs | Recruits TAMs and monocytes | CCL-2 inhibitor mNOX-E36 or neutralizing antibody CNTO88 | [166] |
| 10 | CCL-7 | Up | NF-kB and MAPK | DCs, NK and T cells | Recruitment of TAMs and CAFs proliferation | CCL-7 neutralizing antibodies | [132,167] |
| 11 | CCL-20 | Up | NF-kB, RAS/RAF/MEK/ERK1/2 | NK, TAMs and Tregs | Recruit TAMs & Tregs | Anti-CCL20 (WO2017011559A1) or CCR6 antibodies | [133,168] |
| 12 | CXCL-1 | Up | PI3K/AKT/mTOR, RAS/RAF/MEK/ERK, and NF-ĸB | MDSCs, TAMs, CAFs | Recruit MDSCs to TME and promote metastasis | Small molecule inhibitor (C29) against CXCR1 | [140,169] |
| 13 | CXCL-8 | Up | PI3K/AKT, FAK/Src, Rho/GTPase and MAPK | CSC, endothelial cells | Promote angiogenesis, and CSC proliferation | Antibodies (ABX-CXCL8, HuMax CXCL8) or CXCR1/2 inhibitor Reparixin | [142,170] |
| 14 | CCR-1 | Up | RAS/RAF/MEK/ERK, AKT-mTOR, JAK/STAT3 | MDSCs, TAMs, T cells | Tumor infiltration of MDSCs and Treg cells | CCR-1 inhibitor MLN3897 | [169,170,171] |
| 15 | CXCR-2 | Up | MAPK/MEK/ERK1/2, NF-kB, PI3K/AKT and JAK/STAT3 | Cancer cells, TAMs, Monocytes, T cells | Increase cell proliferation | Antibodies (ABX-CXCL8, HuMax CXCL8) or CXCR-2 inhibitor Reparixin | [162,172] |
| 16 | GM-CSF | Up | JAK/STAT, SRC kinase, MAPK/MEK/ERK1/2, and PI3K/AKT | Tumor cells, DC | DC differentiation, TAM & Treg function and favour IL9-producing Th (Th9) cells | Recombinant GM-CSF or GM-CSF-based DNA vaccines | [18,19,173] |
| 17 | βFGF | Up | FGFR, JAK/STAT, RAS/RAF/MEK/ERK | Tumor cells, CAFs and endothelial cells | Promote cell proliferation and tumor growth | FGFR inhibitors or antibodies (MFGR1877S, BAY 1179470) | [174] |
| 18 | IFN-γ | Down | IFN-γR1/2/JAK-STAT, PI3K/AKT/mTOR, IFN-γ/ICAM1-PI3K-Akt-Notch1 | Tumor cells, DC, T cells, Tregs, Macrophages, | Recruits NK cells to the TME & modulates their activity | rIFN-γ | [122,123,175,176] |
| 19 | TGF-β | Up | TGFβRII/SMAD3, TGFβ/TAK1, NF-κB | Tumor cells, Macrophages, Tregs, NK cells, MDSCs | DC dysfunction, TAMs formation, suppression of NK cells, MDSC recruitment | Anti TGF-β antibody | [18,19,128,177,178] |
| 20 | TNF-α | Up | MAPK, PI3K/AKT, NF-kB, JAK/STAT, FAK/Src | Tumor cells, CAFs | Increase angiogenesis, invasion, and metastasis | - | [18,19,179,180] |
| 21 | VEGF | Up | PI3K/AKT/mTOR, RAS/RAF/MEK/ERK | Endothelial cells | Controls vascular permeability & angiogenesis | Anti-VEGF (Bevacizumab) or anti-VEGFR (Ramucirumab) antibodies | [181] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nisar, S.; Yousuf, P.; Masoodi, T.; Wani, N.A.; Hashem, S.; Singh, M.; Sageena, G.; Mishra, D.; Kumar, R.; Haris, M.; et al. Chemokine-Cytokine Networks in the Head and Neck Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 4584. https://doi.org/10.3390/ijms22094584
Nisar S, Yousuf P, Masoodi T, Wani NA, Hashem S, Singh M, Sageena G, Mishra D, Kumar R, Haris M, et al. Chemokine-Cytokine Networks in the Head and Neck Tumor Microenvironment. International Journal of Molecular Sciences. 2021; 22(9):4584. https://doi.org/10.3390/ijms22094584
Chicago/Turabian StyleNisar, Sabah, Parvaiz Yousuf, Tariq Masoodi, Nissar A. Wani, Sheema Hashem, Mayank Singh, Geetanjali Sageena, Deepika Mishra, Rakesh Kumar, Mohammad Haris, and et al. 2021. "Chemokine-Cytokine Networks in the Head and Neck Tumor Microenvironment" International Journal of Molecular Sciences 22, no. 9: 4584. https://doi.org/10.3390/ijms22094584
APA StyleNisar, S., Yousuf, P., Masoodi, T., Wani, N. A., Hashem, S., Singh, M., Sageena, G., Mishra, D., Kumar, R., Haris, M., Bhat, A. A., & Macha, M. A. (2021). Chemokine-Cytokine Networks in the Head and Neck Tumor Microenvironment. International Journal of Molecular Sciences, 22(9), 4584. https://doi.org/10.3390/ijms22094584