
Regulation of Flowering Timing by ABA-NnSnRK1 Signaling Pathway in Lotus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussions
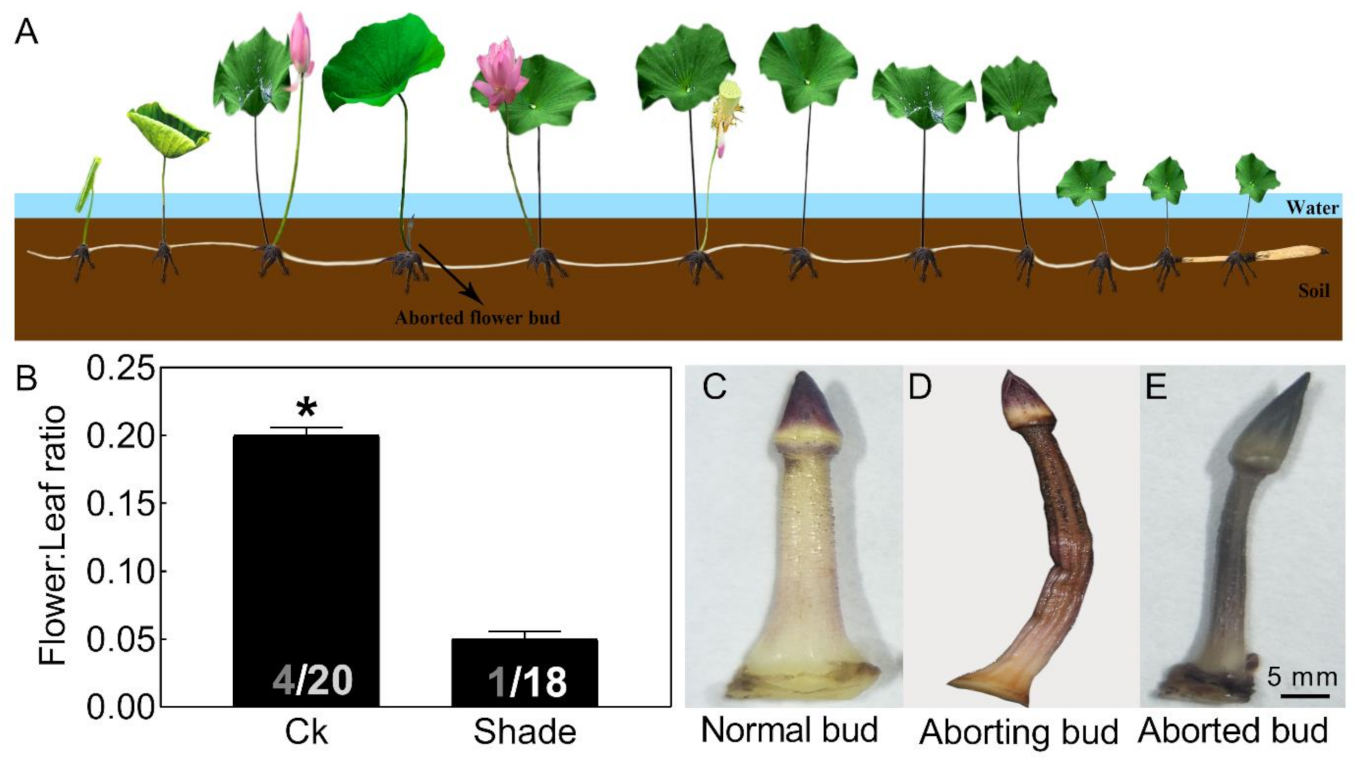
2.1. Most Lotus Flower Buds Aborted in Unfavorable Environments
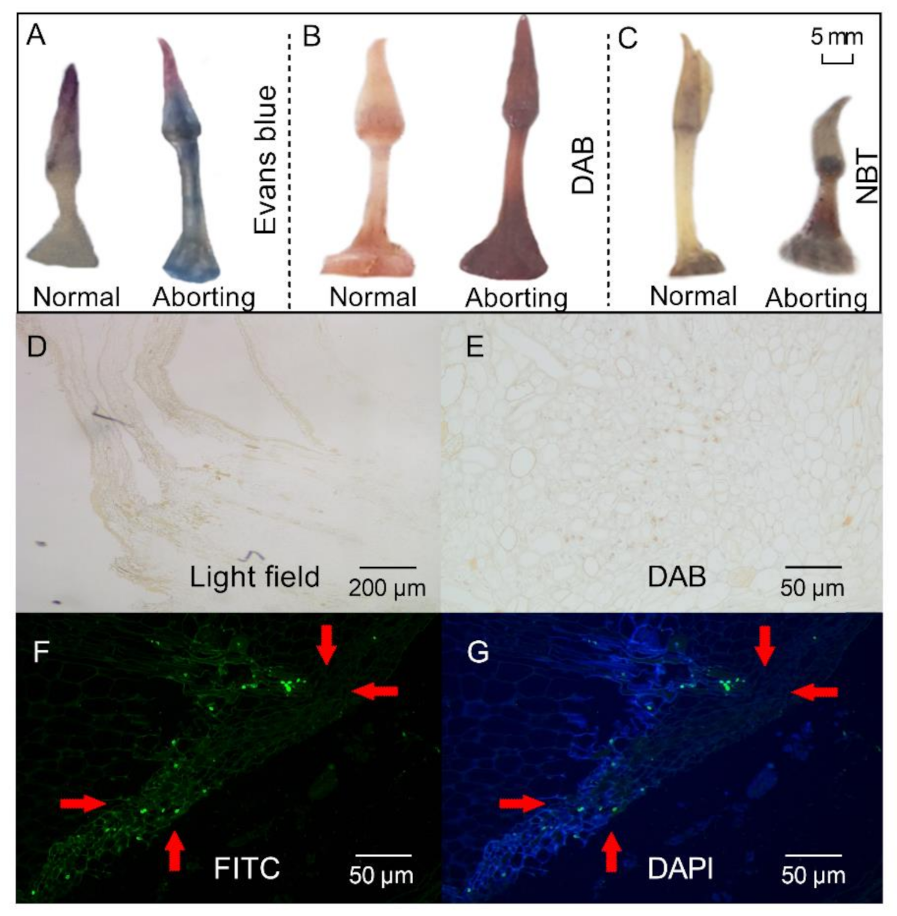
2.2. Programmed Cell Death (PCD) Might Lead to Flower Bud Abortion in the Lotus
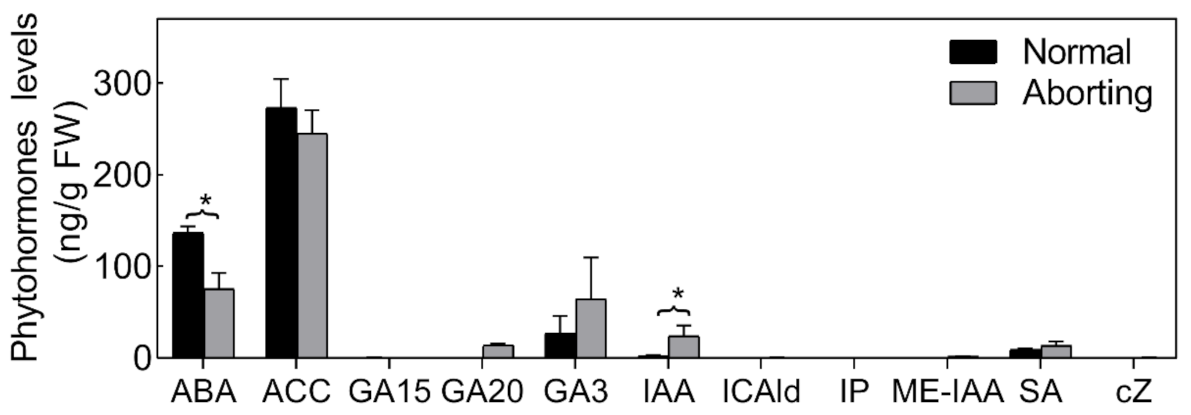
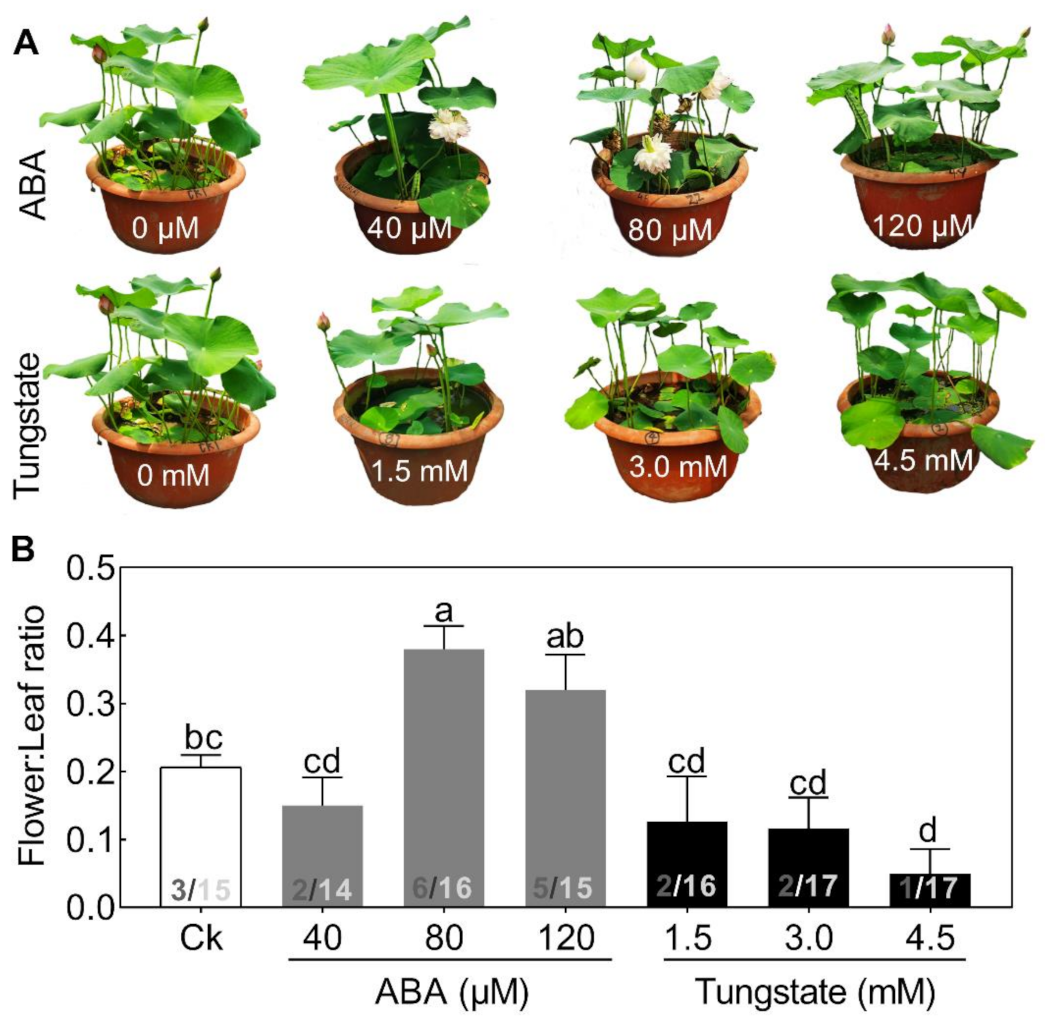
2.3. ABA Plays an Important Role in Regulating Lotus Flower Bud Abortion
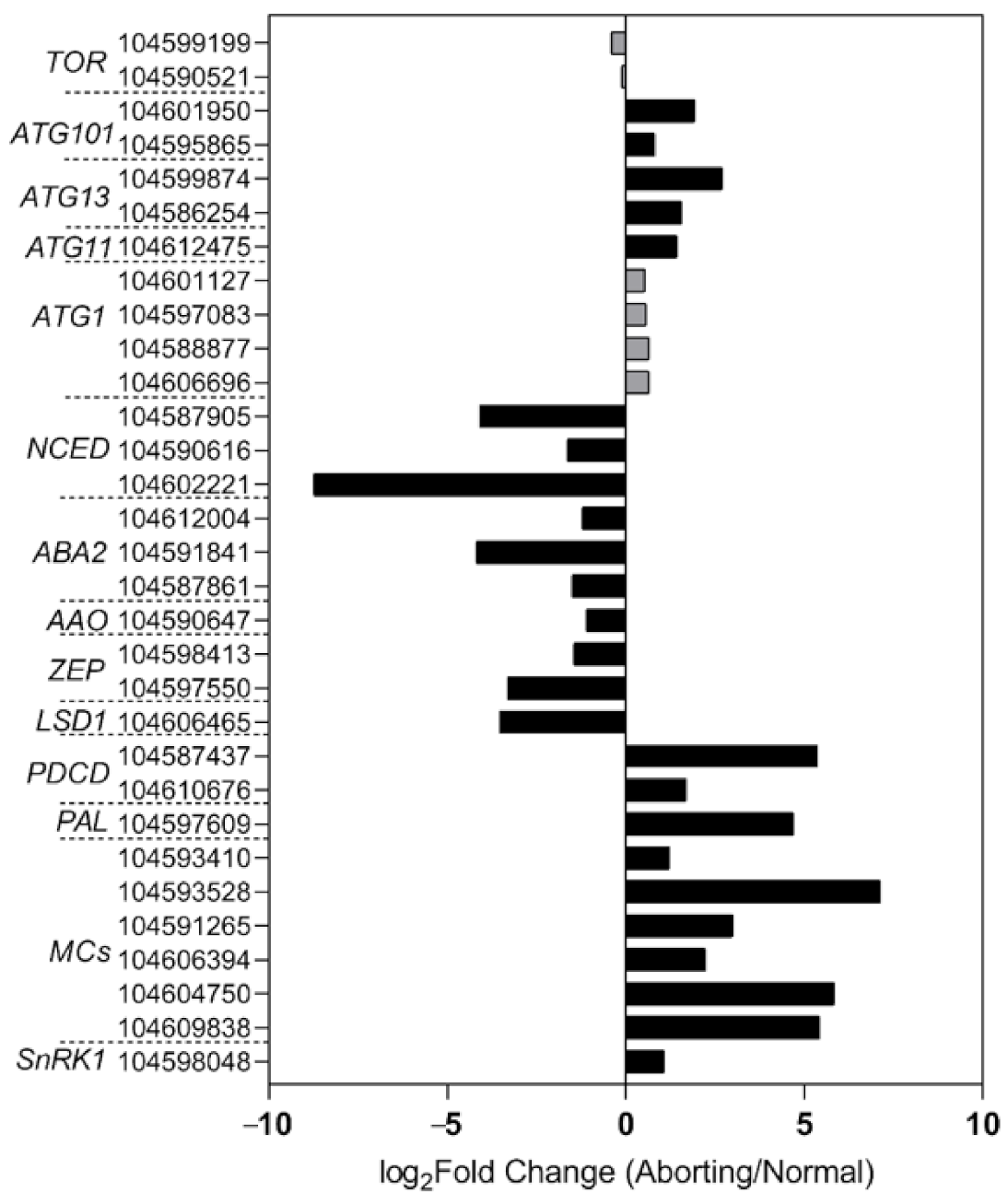
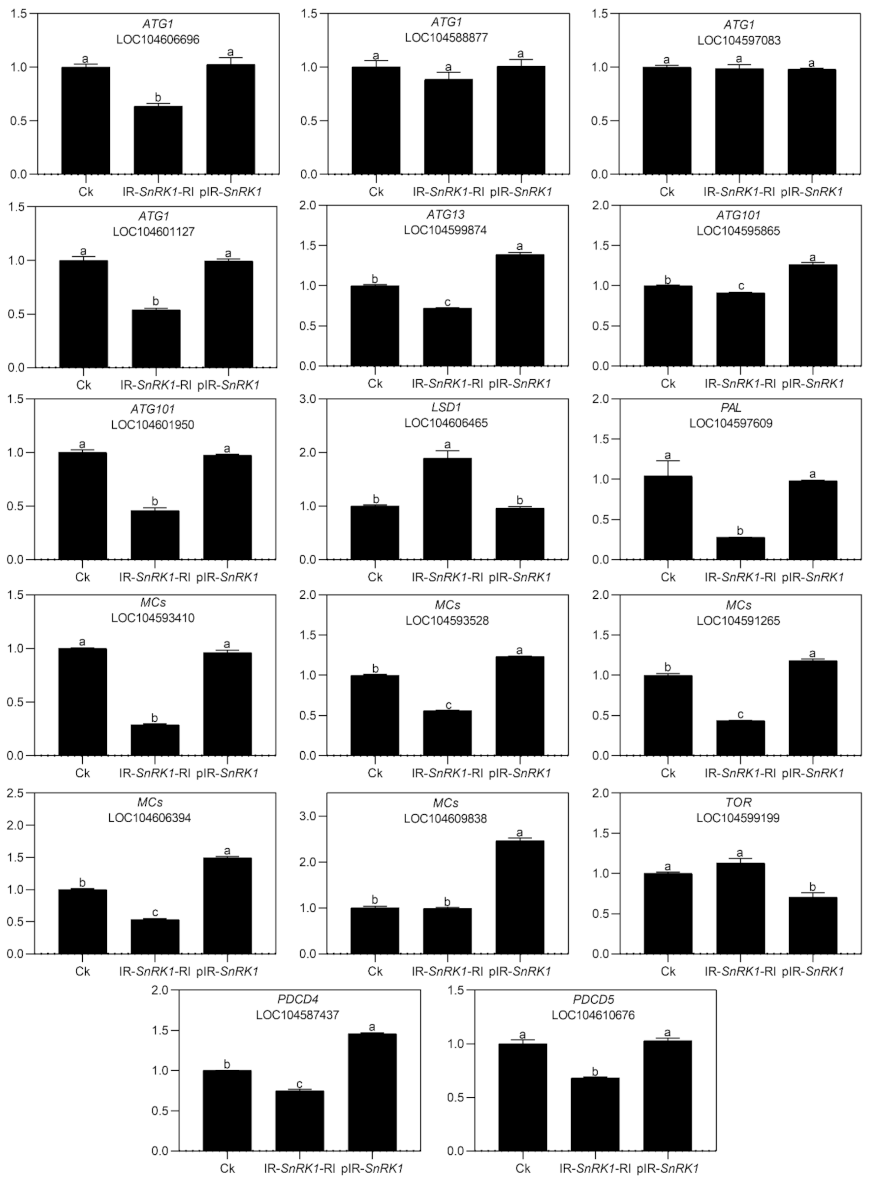
2.4. Transcriptome Analysis Confirmed Changes in ABA and the Occurrence of PCD
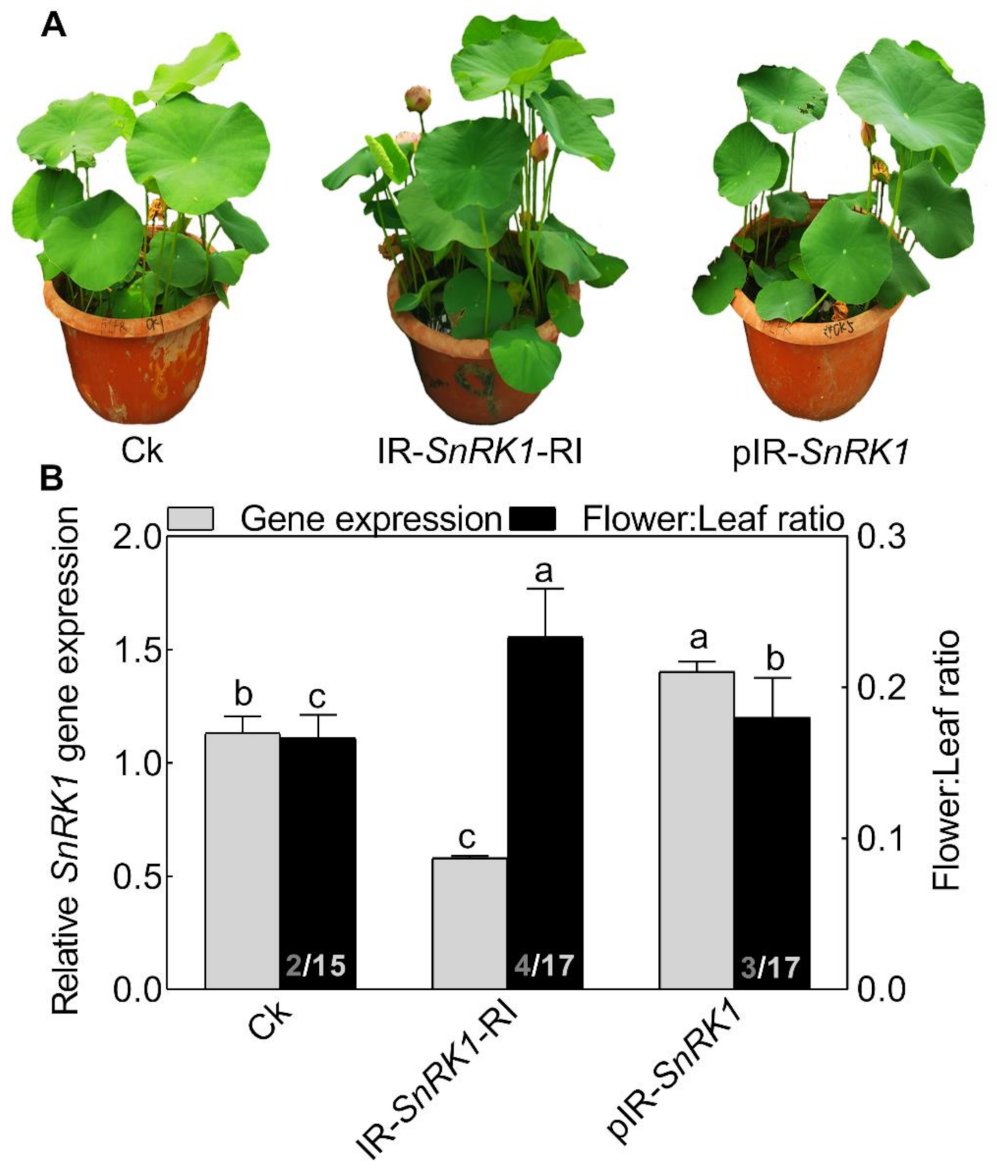
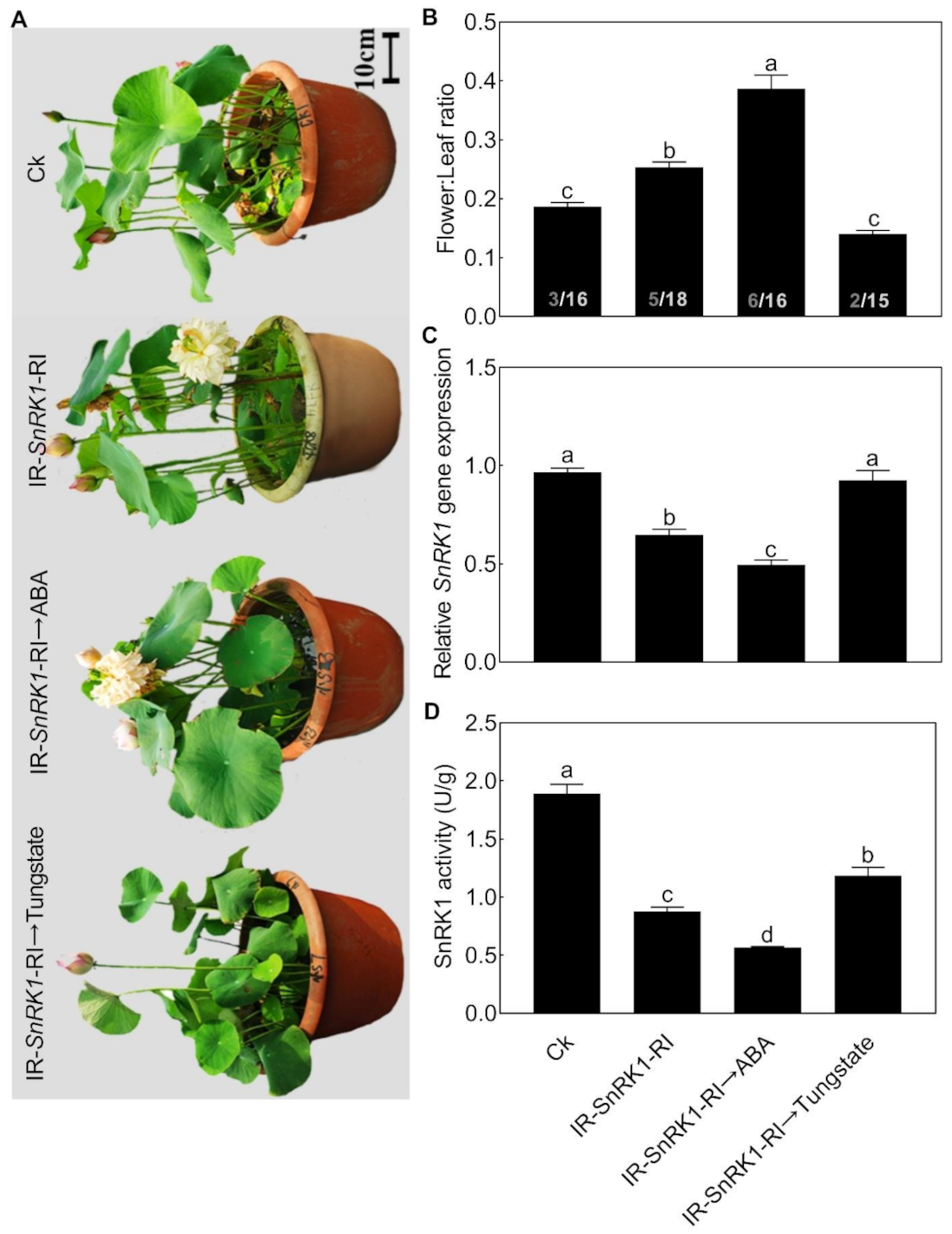
2.5. Silencing NnSnRK1 Could Enhance the Flowering Ability of Lotus
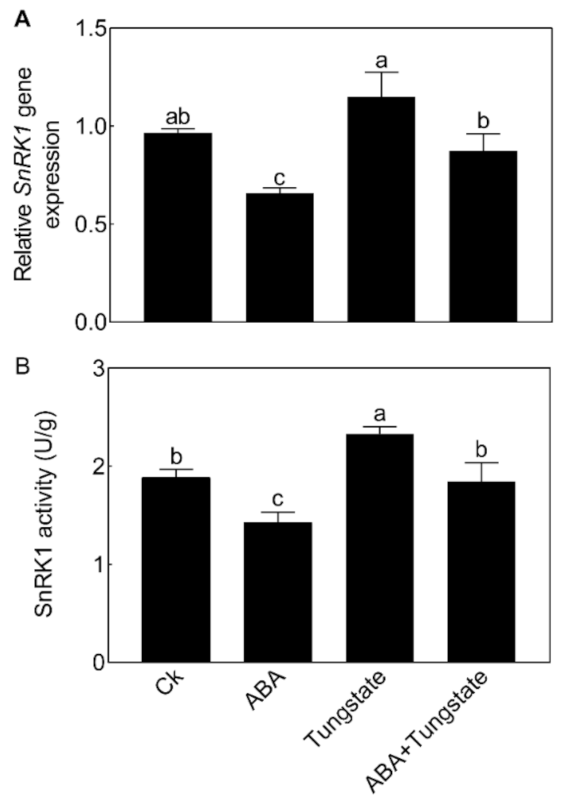
2.6. NnSnRK1 Was a Negative Flowering Regulator in ABA Signaling
3. Materials and Methods
3.1. Plant Material and Reagents
3.2. Histochemical Staining
3.3. TUNEL Analysis
3.4. Hormone Contents in Aborting and Normal Lotus Flower Buds
3.5. RNA-seq Library Preparation and Sequencing
3.6. qRT-PCR and Kinase Activity Analysis of NnSnRK1
3.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wahl, V.; Ponnu, J.; Schlereth, A.; Arrivault, S.; Langenecker, T.; Franke, A.; Feil, R.; Lunn, J.E.; Stitt, M.; Schmid, M. Regulation of flowering by trehalose-6-phosphate signaling in Arabidopsis thaliana. Science 2013, 339, 704–707. [Google Scholar] [CrossRef]
- Srikanth, S. Regulation of flowering time: All roads lead to Rome. Cell. Mol. Life Sci. 2011, 68, 2013–2037. [Google Scholar] [CrossRef]
- Wolters, H.; Juergens, G. Survival of the flexible: Hormonal growth control and adaptation in plant development. Nat. Rev. Genet. 2009, 10, 305–317. [Google Scholar] [CrossRef]
- Li, S.; Li, X.; Wei, Z.; Liu, F. ABA-mediated modulation of elevated CO2 on stomatal response to drought. Curr. Opin. Plant Biol. 2020, 56, 174–180. [Google Scholar] [CrossRef]
- Martignago, D.; Siemiatkowska, B.; Lombardi, A.; Conti, L. Abscisic acid and flowering regulation: Many targets, different places. Int. J. Mol. Sci. 2020, 21, 9700. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.; Susila, H.; Nasim, Z.; Jung, J.Y.; Ahn, J.H. Arabidopsis ABF3 and ABF4 transcription factors act with the NF-YC complex to regulate SOC1 expression and mediate drought-accelerated flowering. Mol. Plant 2019, 12, 489–505. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Huang, F.; Wu, N.; Li, X.; Hu, H.; Xiong, L. Integrative regulation of drought escape through ABA-dependent and -independent pathways in rice. Mol. Plant 2018, 11, 584–597. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Fujita, Y.; Maruyama, K.; Mogami, J.; Todaka, D.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Four Arabidopsis AREB/ABF transcription factors function predominantly in gene expression downstream of SnRK2 kinases in abscisic acid signalling in response to osmotic stress. Plant Cell Environ. 2015, 38, 35–49. [Google Scholar] [CrossRef]
- Coello, P.; Hirano, E.; Hey, S.J.; Muttucumaru, N.; Martinez-Barajas, E.; Parry, M.A.J.; Halford, N.G. Evidence that abscisic acid promotes degradation of SNF1-related protein kinase (SnRK) 1 in wheat and activation of a putative calcium-dependent SnRK2. J. Exp. Bot. 2012, 63, 913–924. [Google Scholar] [CrossRef]
- Jeong, E.-Y.; Seo, P.J.; Woo, J.C.; Park, C.-M. AKIN10 delays flowering by inactivating IDD8 transcription factor through protein phosphorylation in Arabidopsis. BMC Plant Biol. 2015, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Ishizuna, F.; Tsutsumi, N. Flower Bud Formation of Sacred Lotus (Nelumbo nucifera Gaertn.): A Case Study of ‘Gyozankouren’ Grown in a Container. HortScience 2014, 49, 516–518. [Google Scholar] [CrossRef]
- Wang, S.; Xue, J.; Ahmadi, N.; Holloway, P.; Zhu, F.; Ren, X.; Zhang, X. Molecular characterization and expression patterns of PsSVP genes reveal distinct roles in flower bud abortion and flowering in tree peony (Paeonia suffruticosa). Can. J. Plant Sci. 2017, 94, 1181–1193. [Google Scholar] [CrossRef]
- Pien, H.; Lemeur, R.; Van Labeke, M.C. Influence of PAR flux and temperature on the flower bud abortion of rose (Rosa hybrida cv. Frisco) and the carbon balance of the shoot. Acta Hortic. 2000, 515, 119–128. [Google Scholar] [CrossRef]
- Cui, Y.; Peng, Y.; Zhang, Q.; Xia, S.; Ruan, B.; Xu, Q.; Yu, X.; Zhou, T.; Liu, H.; Zeng, D.J.T.P.J. Disruption of EARLY LESION LEAF 1, encoding a cytochrome P450 monooxygenase, induces ROS accumulation and cell death in rice. Plant J. 2020, 105, 942–956. [Google Scholar] [CrossRef]
- Sychta, K.; Slomka, A.; Kuta, E. Insights into Plant Programmed Cell Death Induced by Heavy Metals-Discovering a Terra Incognita. Cells 2021, 10, 65. [Google Scholar] [CrossRef]
- Rogers, H.J.J.A. Programmed Cell Death in Floral Organs: How and Why do Flowers Die? Ann. Bot. 2006, 97, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Valandro, F.; Menguer, P.K.; Cabreira-Cagliari, C.; Margis-Pinheiro, M.; Cagliari, A. Programmed cell death (PCD) control in plants: New insights from the Arabidopsis thaliana deathosome. Plant Sci. 2020, 299, 110603. [Google Scholar] [CrossRef]
- Nambara, E.; Marion-Poll, A. ABA action and interactions in seeds. Trends Plant Sci. 2003, 8, 213–217. [Google Scholar] [CrossRef]
- Jiang, J.; Fan, X.; Zhang, Y.; Tang, X.; Li, X.; Liu, C.; Zhang, Z. Construction of a high-density genetic map and mapping of firmness in grapes (Vitis vinifera L.) based on whole-genome resequencing. Int. J. Mol. Sci. 2020, 21, 797. [Google Scholar] [CrossRef]
- Kim, M.-H.; Cho, J.-S.; Park, E.-J.; Lee, H.; Choi, Y.-I.; Bae, E.-K.; Han, K.-H.; Ko, J.-H. Overexpression of a Poplar RING-H2 Zinc Finger, Ptxerico, Confers Enhanced Drought Tolerance via Reduced Water Loss and Ion Leakage in Populus. Int. J. Mol. Sci. 2020, 21, 9454. [Google Scholar] [CrossRef] [PubMed]
- Grafi, G. Dead but not dead end: Multifunctional role of dead organs enclosing embryos in seed biology. Int. J. Mol. Sci. 2020, 21, 8024. [Google Scholar] [CrossRef]
- Belda-Palazon, B.; Adamo, M.; Valerio, C.; Ferreira, L.J.; Confraria, A.; Reis-Barata, D.; Rodrigues, A.; Meyer, C.; Rodriguez, P.L.; Baena-Gonzalez, E. A dual function of SnRK2 kinases in the regulation of SnRK1 and plant growth. Nat. Plants 2020, 6, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Bao, Y.; Yu, X.; Xia, X.; Liu, C.; Yin, W. Autophagy and its regulators in response to stress in plants. Int. J. Mol. Sci. 2020, 21, 8889. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Li, Z.; An, X.; Long, Y.; Xue, X.; Xie, K.; Ma, B.; Zhang, D.; Guan, Y.; Niu, C.; et al. Normal structure and function of endothecium chloroplasts maintained by ZmMs33-mediated lipid biosynthesis in tapetal cells are critical for anther development in maize. Mol. Plant 2020, 13, 1624–1643. [Google Scholar] [CrossRef] [PubMed]
- Mozes-Koch, R.; Gover, O.; Tanne, E.; Peretz, Y.; Maori, E.; Chernin, L.; Sela, I. Expression of an Entire Bacterial Operon in Plants. Plant Physiol. 2012, 158, 1883–1892. [Google Scholar] [CrossRef] [PubMed]
- Rensburg, H.C.J.V.; Ende, W.V.D.; Signorelli, S. Autophagy in Plants: Both a Puppet and a Puppet Master of Sugars. Other 2019, 10, 14. [Google Scholar] [CrossRef]
- Minina, E.A.; Bozhkov, P.V.; Hofius, D. Autophagy as initiator or executioner of cell death. Trends Plant Sci. 2014, 19, 692–697. [Google Scholar] [CrossRef]
- Kabbage, M.; Kessens, R.; Bartholomay, L.C.; Williams, B. The Life and Death of a Plant Cell. Annu. Rev. Plant Biol. 2017, 68, 375–404. [Google Scholar] [CrossRef]
- Dauphinee, A.N.; Warner, T.S.; Gunawardena, A.H. A comparison of induced and developmental cell death morphologies in lace plant (Aponogeton madagascariensis) leaves. BMC Plant Biol. 2014, 14, 389. [Google Scholar] [CrossRef]
- Cao, J.J.; Liu, C.X.; Shao, S.J.; Zhou, J. Molecular Mechanisms of Autophagy Regulation in Plants and Their Applications in Agriculture. Front. Plant Sci. 2020, 11, 618944. [Google Scholar] [CrossRef]
- Cao, L.; Wu, Y.; Wang, X.; Li, X.; Tan, Z.; Guan, F. Role of Site-Specific Glycosylation in the I-Like Domain of Integrin beta1 in Small Extracellular Vesicle-Mediated Malignant Behavior and FAK Activation. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.; Kalia, M.; Shorey, R. Trehalose-6-phosphate and SnRK1 kinases in plant development and signaling: The emerging picture. Front. Plant Sci. 2014, 5, 119. [Google Scholar] [CrossRef]
- Lee, J.-H.; Terzaghi, W.; Gusmaroli, G.; Charron, J.-B.F.; Yoon, H.-J.; Chen, H.; He, Y.J.; Xiong, Y.; Deng, X.W. Characterization of Arabidopsis and rice DWD proteins and their roles as substrate receptors for CUL4-RING E3 ubiquitin ligases. Plant Cell 2008, 20, 152–167. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.; Adamo, M.; Crozet, P.; Margalha, L.; Confraria, A.; Martinho, C.; Elias, A.; Rabissi, A.; Lumbreras, V.; Gonzalez-Guzman, M.; et al. ABI1 and PP2CA phosphatases are negative regulators of Snf1-related protein kinase1 signaling in Arabidopsis. Plant Cell 2013, 25, 3871–3884. [Google Scholar] [CrossRef]
- Li, X.S.; Song, L.L. The role of ABA in the responses of wild-type and abscisic acid mutants of Arabidopsis thaliana to excess zinc. Acta Physiol. Plant. 2020, 42, 74. [Google Scholar] [CrossRef]
- Bertocchi, M.; Rigillo, A.; Elmi, A.; Ventrella, D.; Aniballi, C.; Scorpio, D.G.; Scozzoli, M.; Bettini, G.; Forni, M.; Bacci, M.L. Preliminary assessment of the mucosal toxicity of tea tree (Melaleuca alternifolia) and rosemary (Rosmarinus officinalis) essential oils on novel porcine uterus models. Int. J. Mol. Sci. 2020, 21, 3350. [Google Scholar] [CrossRef]
- Muneer, S.; Wei, H.; Park, Y.G.; Jeong, H.K.; Jeong, B.R. Proteomic analysis reveals the dynamic role of silicon in alleviation of hyperhydricity in carnation grown in vitro. Int. J. Mol. Sci. 2018, 19, 50. [Google Scholar] [CrossRef]
- Wu, T.-M.; Huang, J.-Z.; Oung, H.-M.; Hsu, Y.-T.; Tsai, Y.-C.; Hong, C.-Y. H2O2-based method for rapid detection of transgene-free rice plants from segregating CRISPR/Cas9 genome-edited progenies. Int. J. Mol. Sci. 2019, 20, 3885. [Google Scholar] [CrossRef]
- Karna, K.K.; Choi, N.Y.; Kim, C.Y.; Kim, H.K.; Shin, Y.S.; Park, J.K. Gui-A-Gra attenuates testicular dysfunction in varicocele-induced rats via oxidative stress, ER stress and mitochondrial apoptosis pathway. Int. J. Mol. Sci. 2020, 21, 9231. [Google Scholar] [CrossRef]
- Šimura, J.; Antoniadi, I.; Široká, J.; Tarkowská, D.; Strnad, M.; Ljung, K.; Novák, O. Plant hormonomics: Multiple phytohormone profiling by targeted metabolomics. Plant Physiol. 2018, 177, 476–489. [Google Scholar] [CrossRef]
- Floková, K.; Tarkowská, D.; Miersch, O.; Strnad, M.; Wasternack, C.; Novák, O. UHPLC–MS/MS based target profiling of stress-induced phytohormones. Phytochemistry 2014, 105, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; Lin, Y.; Zhou, X.; Li, S.; Liu, H.; Zeng, F.; Zhu, F.; Ouyang, G.; Zeng, Z. Comparison of sample pretreatment methods for the determination of multiple phytohormones in plant samples by liquid chromatography–electrospray ionization-tandem mass spectrometry. Microchem. J. 2015, 121, 25–31. [Google Scholar] [CrossRef]
- Xiao, H.-M.; Cai, W.-J.; Ye, T.-T.; Ding, J.; Feng, Y.-Q. Spatio-temporal profiling of abscisic acid, indoleacetic acid and jasmonic acid in single rice seed during seed germination. Anal. Chim. Acta 2018, 1031, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Wang, J.; Li, B.; Meng, Y.; Ma, X.; Si, E.; Ren, P.; Yang, K.; Shang, X.; Wang, H. Transcriptome sequencing and comparative analysis of differentially-expressed isoforms in the roots of Halogeton glomeratus under salt stress. Gene 2018, 646, 159–168. [Google Scholar] [CrossRef]
- Gover, O.; Peretz, Y.; Mozes-Koch, R.; Maori, E.; Rabinowitch, H.D.; Sela, I. Only minimal regions of tomato yellow leaf curl virus (TYLCV) are required for replication, expression and movement. Arch. Virol. 2014, 159, 2263–2274. [Google Scholar] [CrossRef]
- Peretz, Y.; Mozes-Koch, R.; Akad, F.; Tanne, E.; Czosnek, H.; Sela, I. A universal expression/silencing vector in plants. Plant Physiol. 2007, 145, 1251–1263. [Google Scholar] [CrossRef]
- Zhang, B.; Li, X.; Li, X.; Lu, Z.; Cai, X.; Yang, Q.O.; Ma, P.; Dong, J. Lipopolysaccharide Enhances Tanshinone Biosynthesis via a Ca2+-Dependent Manner in Salvia miltiorrhiza Hairy Roots. Int. J. Mol. Sci. 2020, 21, 9576. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, J.; Jin, Q.; Kuang, J.; Wang, Y.; Xu, Y. Regulation of Flowering Timing by ABA-NnSnRK1 Signaling Pathway in Lotus. Int. J. Mol. Sci. 2021, 22, 3932. https://doi.org/10.3390/ijms22083932
Cao J, Jin Q, Kuang J, Wang Y, Xu Y. Regulation of Flowering Timing by ABA-NnSnRK1 Signaling Pathway in Lotus. International Journal of Molecular Sciences. 2021; 22(8):3932. https://doi.org/10.3390/ijms22083932
Chicago/Turabian StyleCao, Jing, Qijiang Jin, Jiaying Kuang, Yanjie Wang, and Yingchun Xu. 2021. "Regulation of Flowering Timing by ABA-NnSnRK1 Signaling Pathway in Lotus" International Journal of Molecular Sciences 22, no. 8: 3932. https://doi.org/10.3390/ijms22083932
APA StyleCao, J., Jin, Q., Kuang, J., Wang, Y., & Xu, Y. (2021). Regulation of Flowering Timing by ABA-NnSnRK1 Signaling Pathway in Lotus. International Journal of Molecular Sciences, 22(8), 3932. https://doi.org/10.3390/ijms22083932