
The Main Alkaloids in Uncaria rhynchophylla and Their Anti-Alzheimer’s Disease Mechanism Determined by a Network Pharmacology Approach


Abstract
1. Introduction
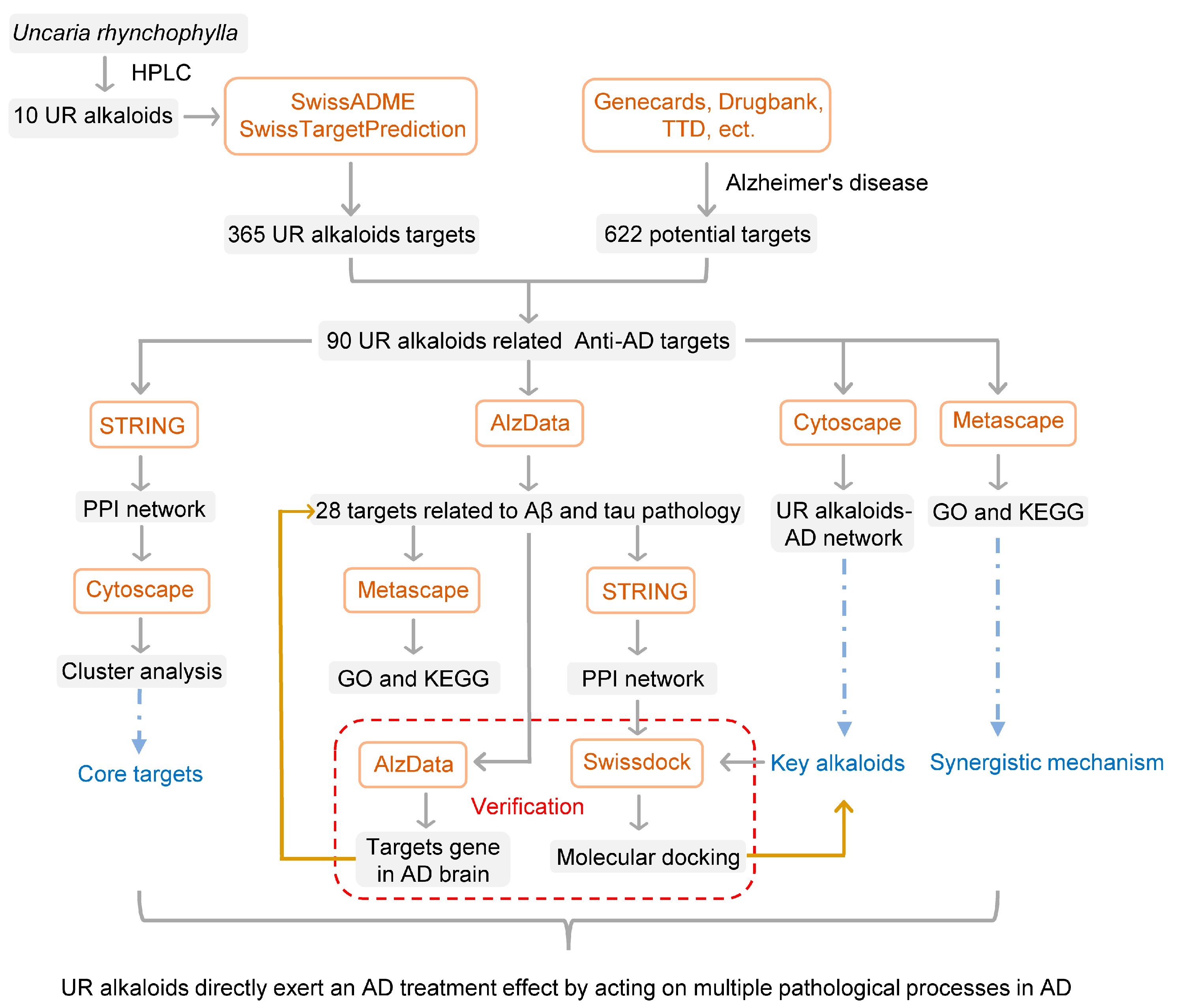
2. Results
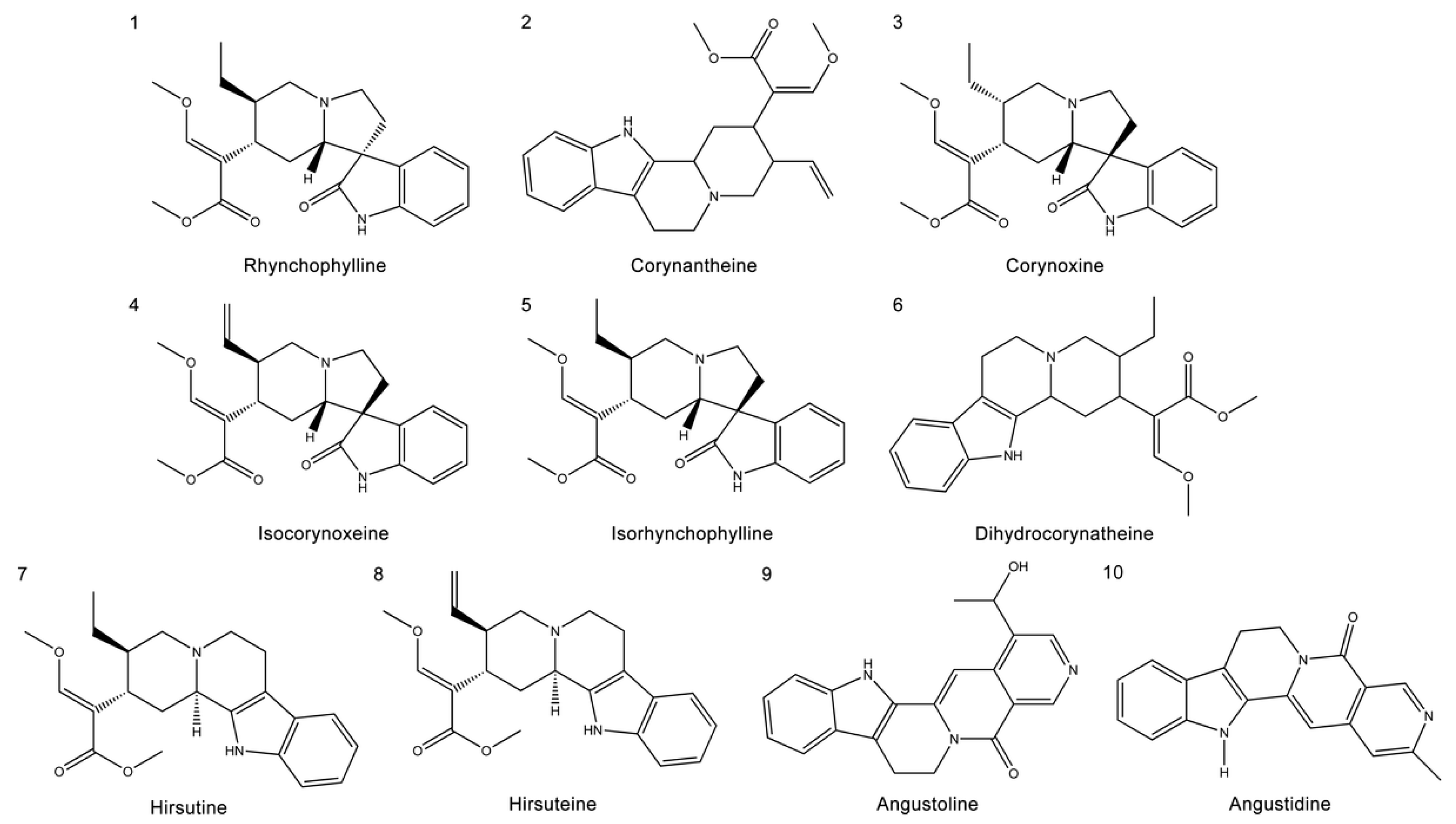
2.1. The Chemical Structure and ADME Properties of the Main Alkaloids from UR
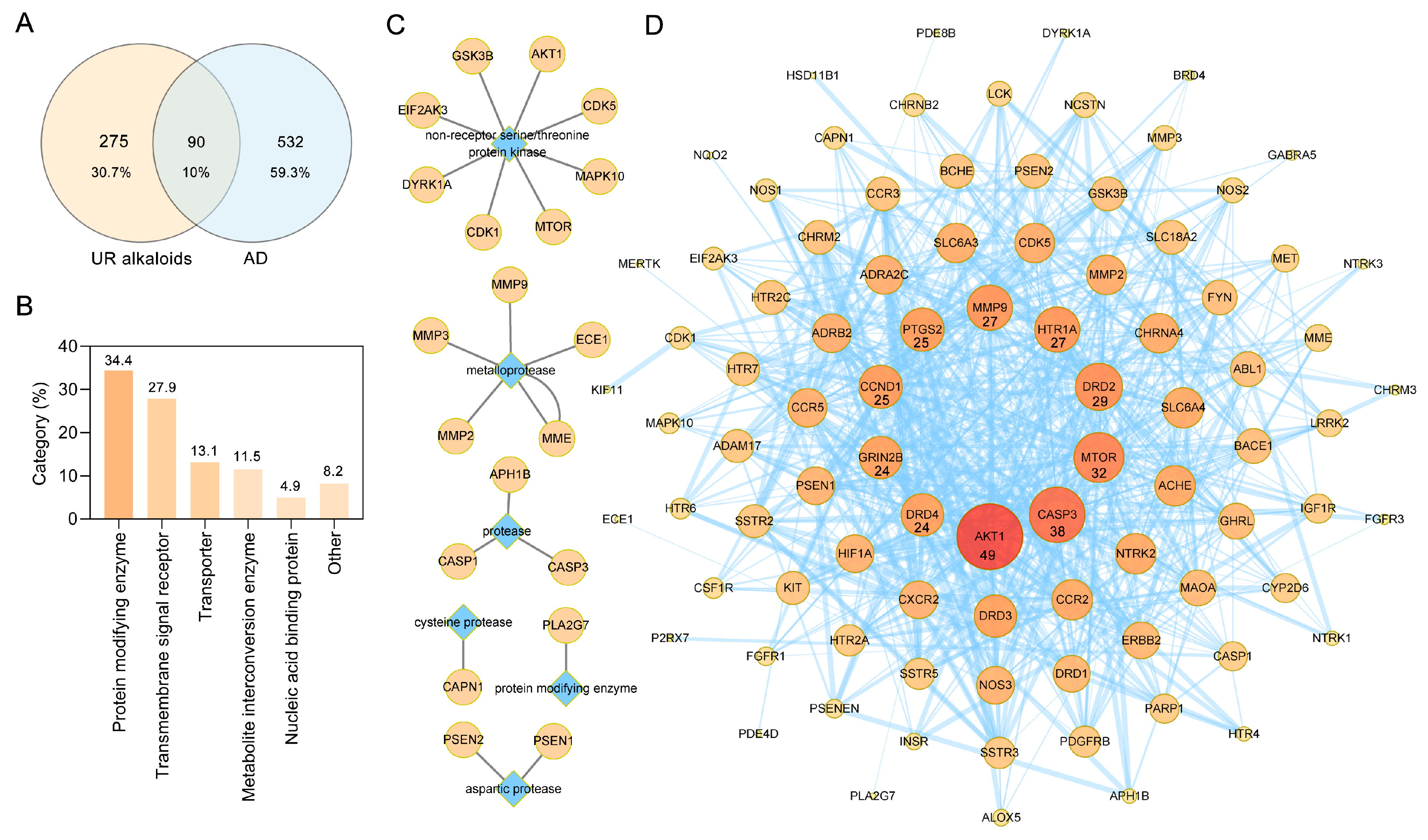
2.2. Screening of Targets of the Main Alkaloids from UR in AD
2.3. PPI Analysis of Targets of the Main Alkaloids against AD
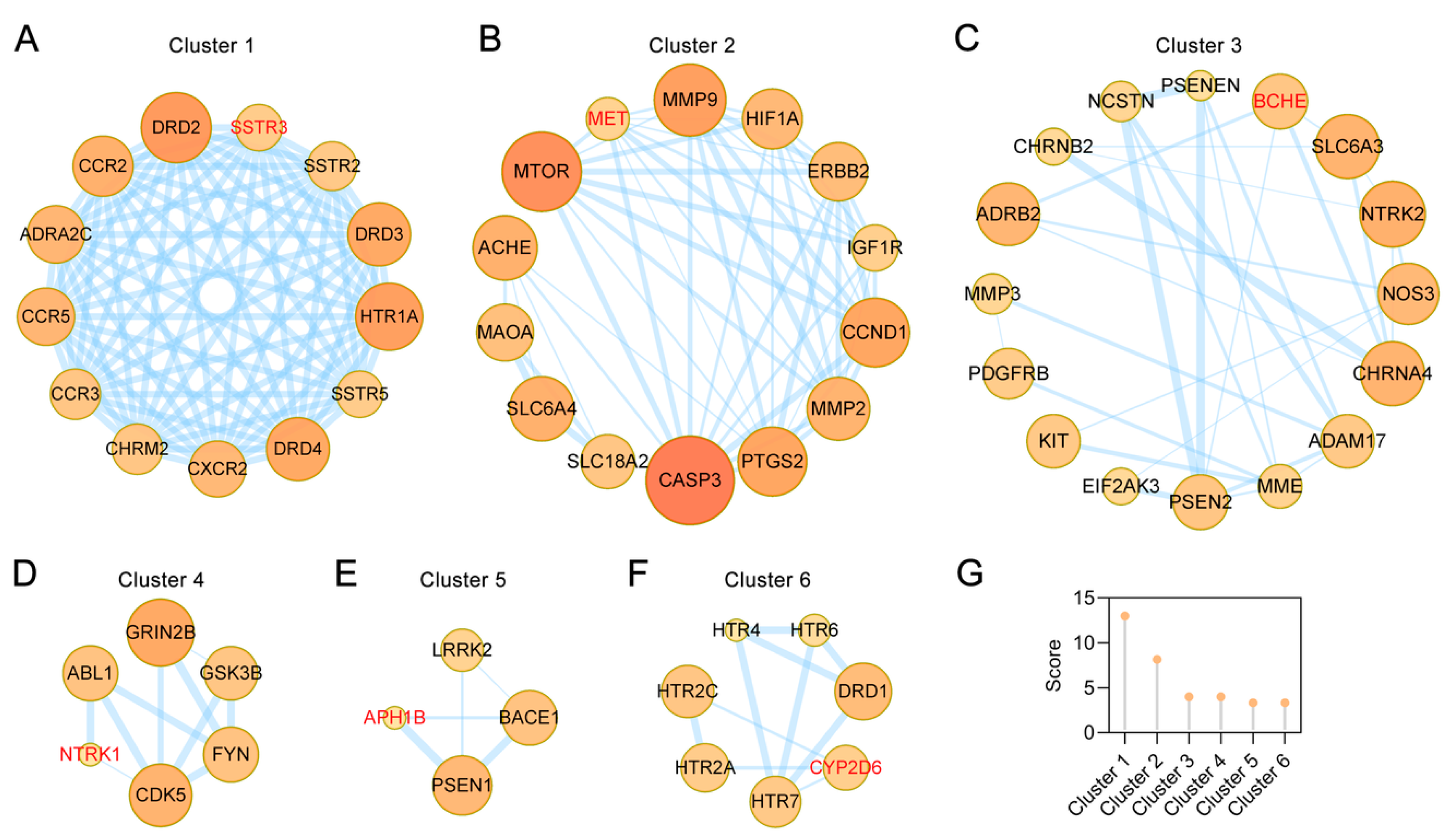
2.4. Clusters of Common Targets of the Main Alkaloids against AD
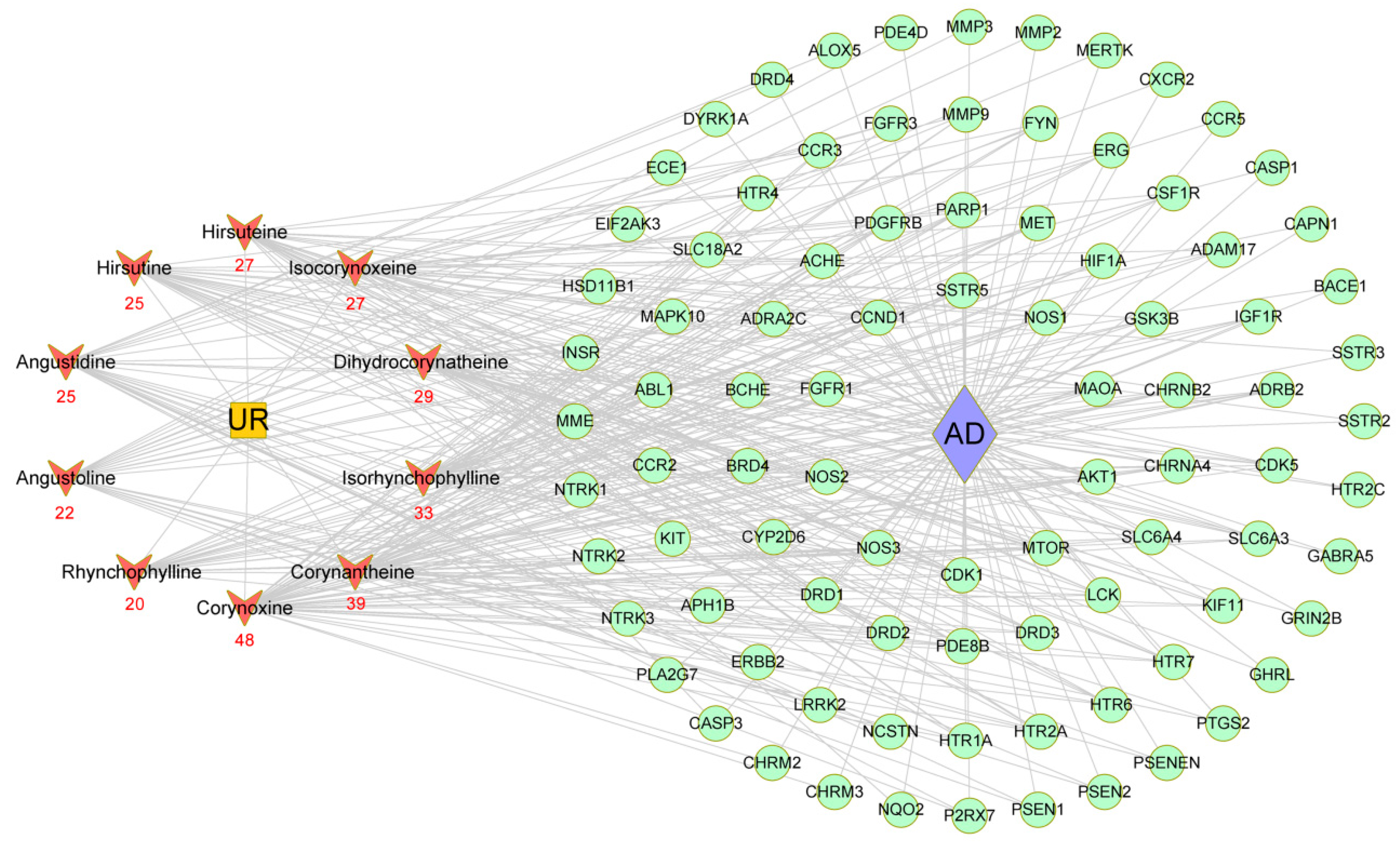
2.5. Construction of a Main Potential Alkaloid Target-AD Target Network
2.6. Potential Synergistic Mechanism of the 10 Alkaloids against AD
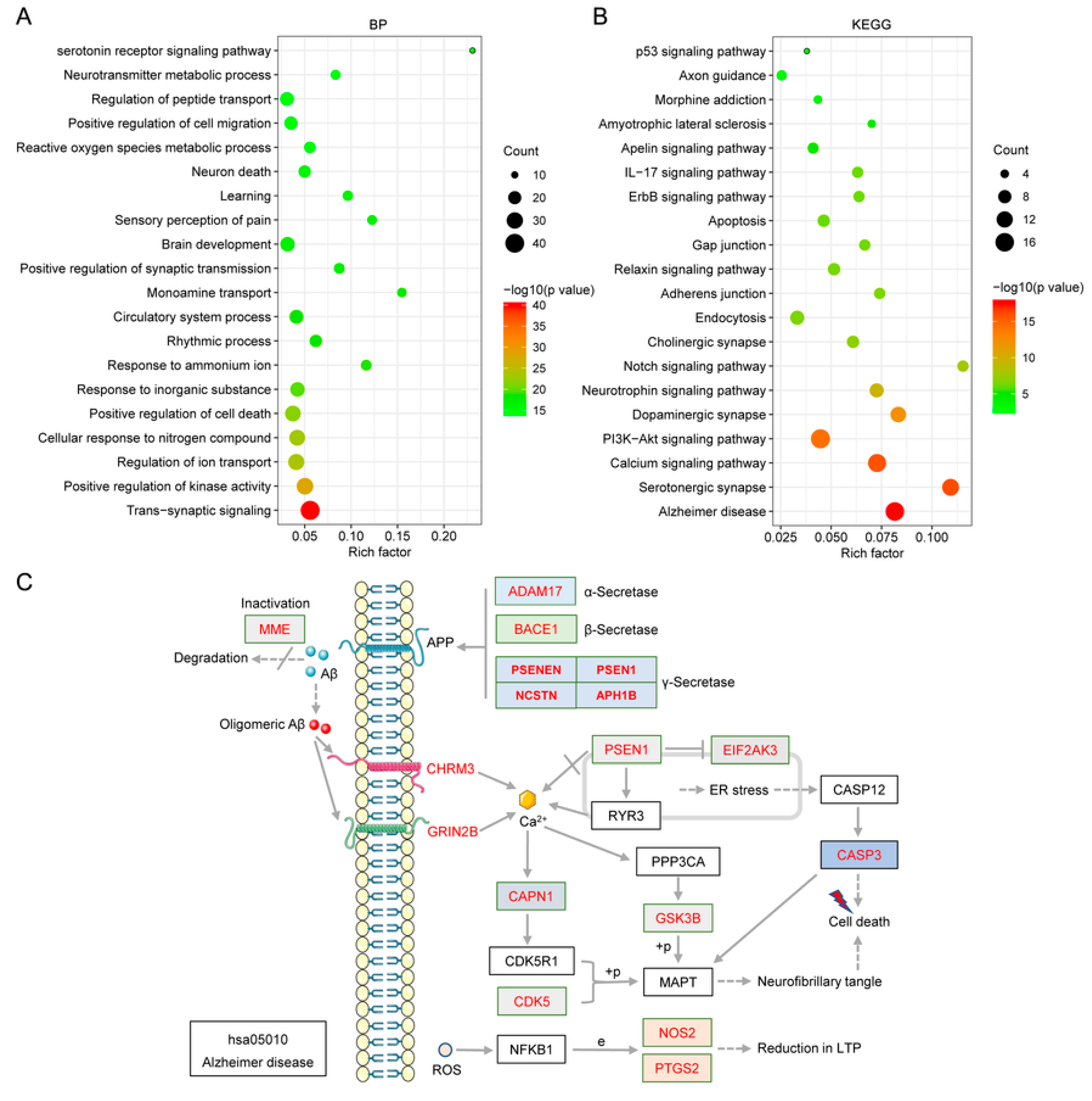
2.6.1. Gene Ontology (GO) Enrichment Analysis
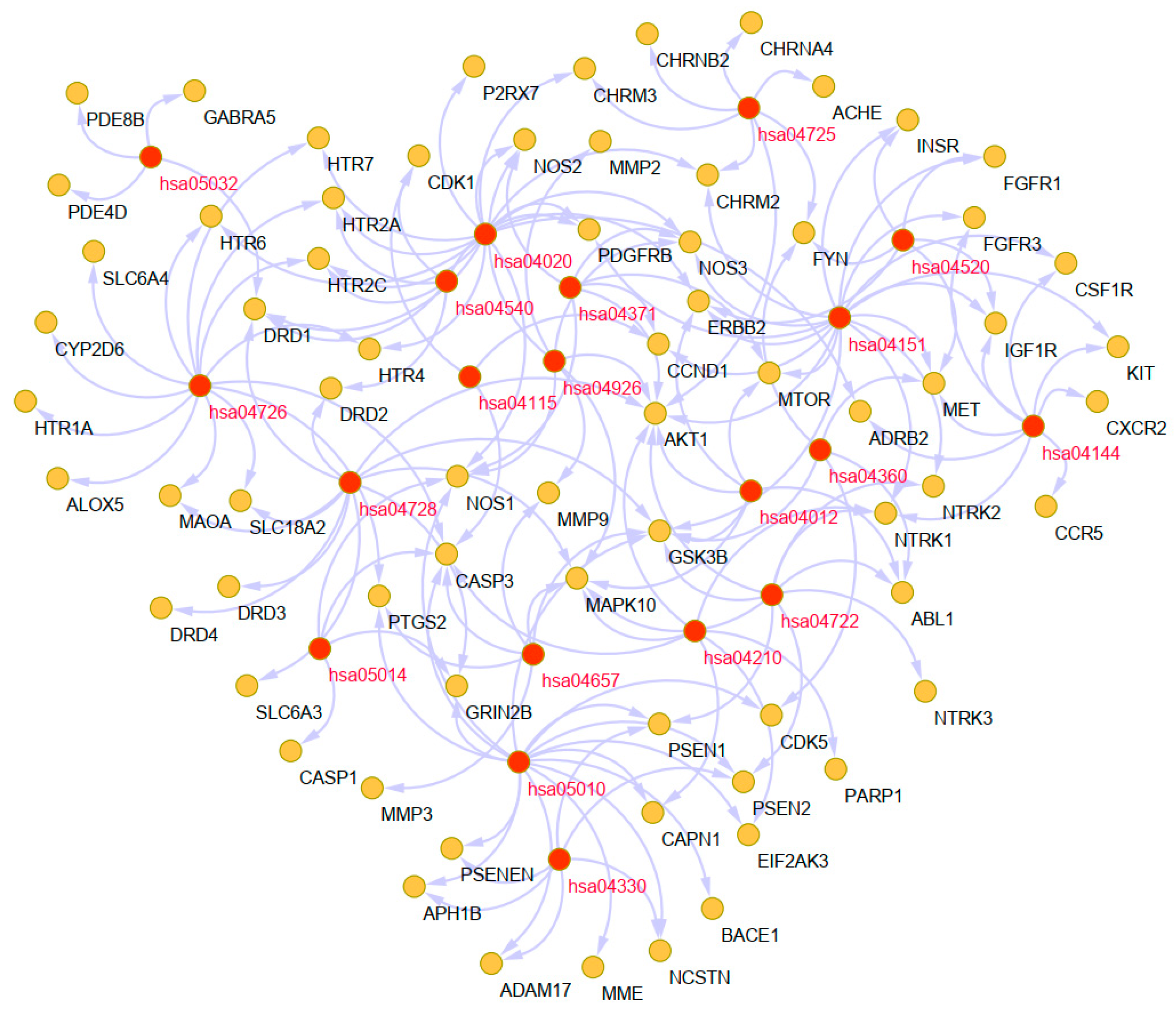
2.6.2. Pathway Analysis to Explore the Therapeutic Mechanisms of the Main Alkaloids in AD
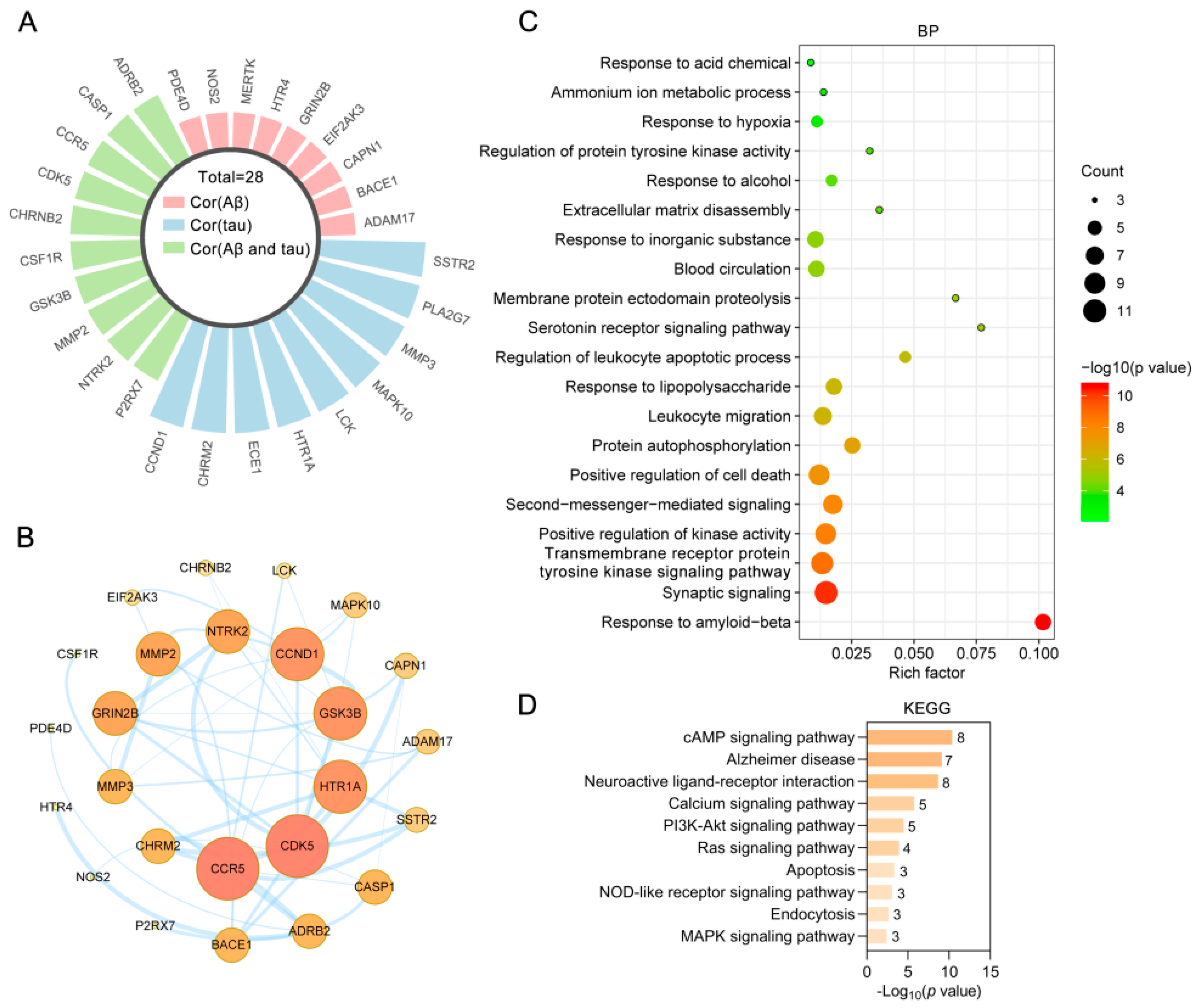
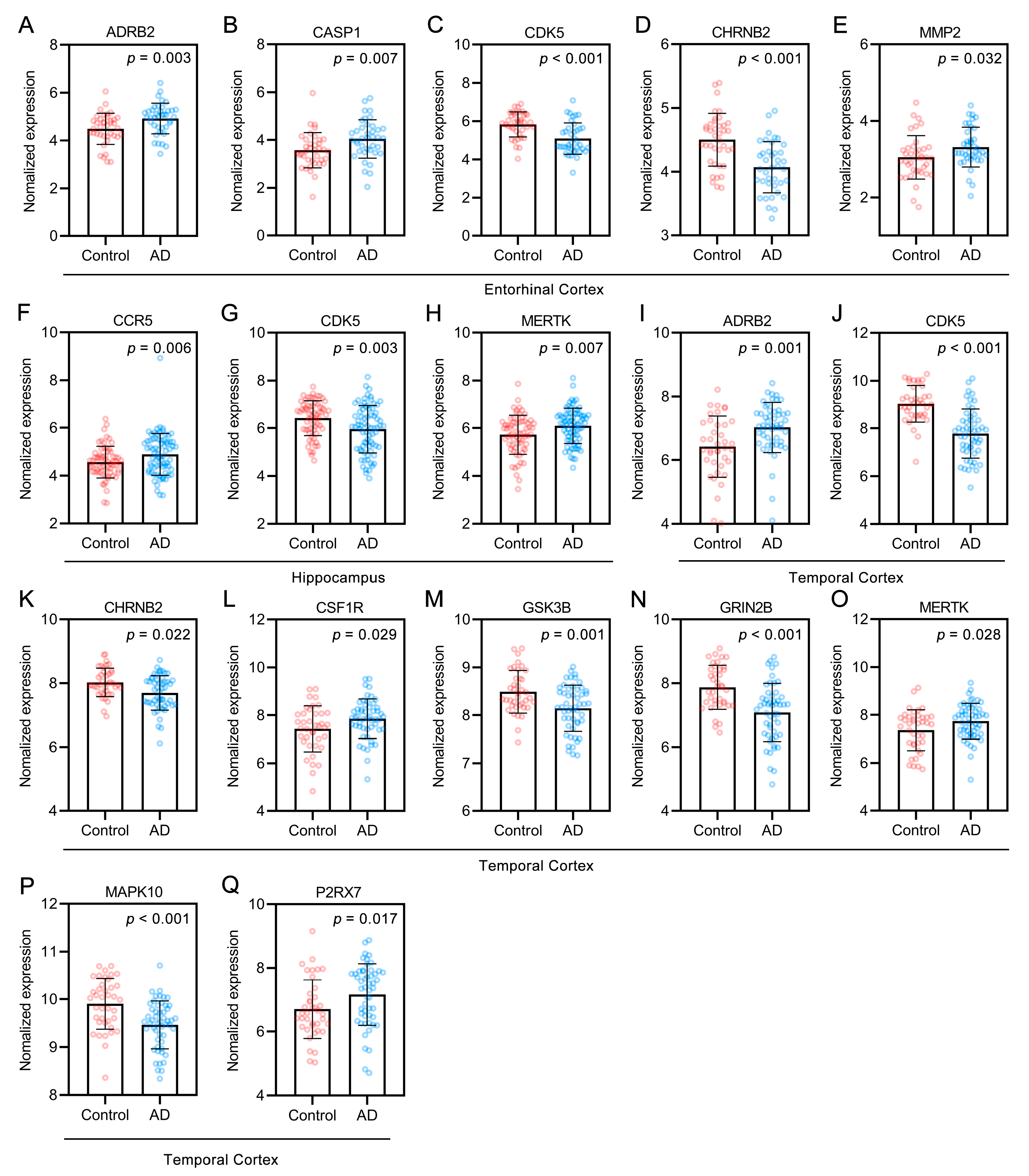
2.7. Bioinformatics Analysis of Targets of the Alkaloids Related to Aβ and Tau Pathology
2.8. Molecular Docking Simulation
3. Discussion
4. Materials and Methods
4.1. Collection of the Main Alkaloids from UR and ADME Evaluation
4.2. Target Fishing
4.2.1. Collection of the Main Alkaloid-Related Targets
4.2.2. Screening of Targets of the Main Alkaloids against AD
4.3. PPI Network Construction and Clustering Analysis
4.4. GO and KEGG Pathway Enrichment Analyses
4.5. Analysis of Alkaloid Targets Related to AD Pathology
4.6. Molecular Docking
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| APH1B | Aph-1 homolog b |
| APP | β-amyloid precursor protein |
| Aβ | amyloid beta |
| BCHE | Butyrylcholinesterase |
| BP | Biological process |
| CYP2D6 | Cytochrome p450 family 2 subfamily d member 6 |
| GEO | Gene Expression Omnibus |
| GO | Gene Ontology |
| Hacc | Hydrogen bond acceptors |
| Hdon | Hydrogen bond donors |
| HPLC | High-performance liquid chromatography |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| MCODE | Molecular complex detection |
| MET | Met proto-oncogene receptor tyrosine kinase |
| MW | Molecule weight |
| NCT | Nicastrin |
| NFT | Neurofibrillary tangles |
| N-methyl D-aspartate | Nmda |
| NTRK1 | Neurotrophic receptor tyrosine kinase 1 |
| PPI | Protein–protein interaction |
| Rbon | Rotatable bonds |
| SP | Senile plaques |
| SSTR3 | Somatostatin receptor 3 |
| TCMSP | Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform |
| TPSA | Topological polar surface area |
| TTD | Therapeutic target database |
| UR | Uncaria rhynchophylla |
References
- McKhann, G.; Drachman, D.; Folstein, M.; Katzman, R.; Price, D.; Stadlan, E.M. Clinical diagnosis of Alzheimer’s disease: Report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology 1984, 34, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhong, R.J.; Cheng, C.; Li, S.; Le, W.D. New therapeutics beyond amyloid-beta and tau for the treatment of Alzheimer’s disease. Acta Pharmacol. Sin. 2020, 1–8. [Google Scholar] [CrossRef]
- Guo, Y.; Yang, H.; Huang, Z.; Tian, S.; Li, Q.; Du, C.; Chen, T.; Liu, Y.; Sun, H.; Liu, Z. Design, Synthesis, and Evaluation of Acetylcholinesterase and Butyrylcholinesterase Dual-Target Inhibitors against Alzheimer’s Diseases. Molecules 2020, 25, 489. [Google Scholar] [CrossRef]
- Wallace, R.A.; Dalton, A.J. What can we learn from study of Alzheimer’s disease in patients with Down syndrome for early-onset Alzheimer’s disease in the general population? Alzheimers Res. Ther. 2011, 3, 13. [Google Scholar] [CrossRef][Green Version]
- Asaad, M.; Lee, J.H. A guide to using functional magnetic resonance imaging to study Alzheimer’s disease in animal models. Dis. Model. Mech. 2018, 11, dmm031724. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.G.; Zhou, X.Q.; Mrak, R.E.; Griffin, W.S. Progressive neuronal injury associated with amyloid plaque formation in Alzheimer disease. J. Neuropathol. Exp. Neurol. 1998, 57, 714–717. [Google Scholar] [CrossRef] [PubMed]
- Scheltens, P.; Blennow, K.; Breteler, M.M.; de Strooper, B.; Frisoni, G.B.; Salloway, S.; Van der Flier, W.M. Alzheimer’s disease. Lancet 2016, 388, 505–517. [Google Scholar] [CrossRef]
- Mehta, D.; Jackson, R.; Paul, G.; Shi, J.; Sabbagh, M. Why do trials for Alzheimer’s disease drugs keep failing? A discontinued drug perspective for 2010–2015. Expert Opin. Investig. Drugs 2017, 26, 735–739. [Google Scholar] [CrossRef]
- Melnikova, I. Therapies for Alzheimer’s disease. Nat. Rev. Drug Discov. 2007, 6, 341–342. [Google Scholar] [CrossRef]
- Inglis, F. The tolerability and safety of cholinesterase inhibitors in the treatment of dementia. Int. J. Clin. Pract. Suppl. 2002, 127, 45–63. [Google Scholar]
- Lee, J.; Jin, C.; Cho, S.Y.; Park, S.U.; Jung, W.S.; Moon, S.K.; Park, J.M.; Ko, C.N.; Cho, K.H.; Kwon, S. Herbal medicine treatment for Alzheimer disease: A protocol for a systematic review and meta-analysis. Medicine 2020, 99, e21745. [Google Scholar] [CrossRef]
- Zhou, J.; Zhou, S. Antihypertensive and neuroprotective activities of rhynchophylline: The role of rhynchophylline in neurotransmission and ion channel activity. J. Ethnopharmacol. 2010, 132, 15–27. [Google Scholar] [CrossRef]
- Hsieh, C.L.; Chen, M.F.; Li, T.C.; Li, S.C.; Tang, N.Y.; Hsieh, C.T.; Pon, C.Z.; Lin, J.G. Anticonvulsant effect of Uncaria rhynchophylla (Miq) Jack. in rats with kainic acid-induced epileptic seizure. Am. J. Chin. Med. 1999, 27, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.L.; Tang, N.Y.; Chiang, S.Y.; Hsieh, C.T.; Lin, J.G. Anticonvulsive and free radical scavenging actions of two herbs, Uncaria rhynchophylla (MIQ) Jack and Gastrodia elata Bl., in kainic acid-treated rats. Life Sci. 1999, 65, 2071–2082. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, Y.H.; Zeng, C.Q.; Zeng, Y. Research progress in pharmacological effects of Uncaria Hook on Alzheimer disease models. Acta Pharm. Sin. 2016, 51, 536–542. [Google Scholar]
- Yuan, D.; Ma, B.; Yang, J.Y.; Xie, Y.Y.; Wang, L.; Zhang, L.J.; Kano, Y.; Wu, C.F. Anti-inflammatory effects of rhynchophylline and isorhynchophylline in mouse N9 microglial cells and the molecular mechanism. Int. Immunopharmacol. 2009, 9, 1549–1554. [Google Scholar] [CrossRef]
- Zheng, M.; Chen, M.; Liu, C.; Fan, Y.; Shi, D. Alkaloids extracted from Uncaria rhynchophylla demonstrate neuroprotective effects in MPTP-induced experimental parkinsonism by regulating the PI3K/Akt/mTOR signaling pathway. J. Ethnopharmacol. 2021, 266, 113451. [Google Scholar] [CrossRef]
- Shi, J.S.; Yu, J.X.; Chen, X.P.; Xu, R.X. Pharmacological actions of Uncaria alkaloids, rhynchophylline and isorhynchophylline. Acta Pharmacol. Sin. 2003, 24, 97–101. [Google Scholar]
- Shin, S.J.; Jeong, Y.; Jeon, S.G.; Kim, S.; Lee, S.K.; Choi, H.S.; Im, C.S.; Kim, S.H.; Kim, S.H.; Park, J.H.; et al. Uncaria rhynchophylla ameliorates amyloid beta deposition and amyloid beta-mediated pathology in 5XFAD mice. Neurochem. Int. 2018, 121, 114–124. [Google Scholar] [CrossRef]
- Yang, Y.; Ji, W.G.; Zhu, Z.R.; Wu, Y.L.; Zhang, Z.Y.; Qu, S.C. Rhynchophylline suppresses soluble Abeta1-42-induced impairment of spatial cognition function via inhibiting excessive activation of extrasynaptic NR2B-containing NMDA receptors. Neuropharmacology 2018, 135, 100–112. [Google Scholar] [CrossRef]
- Xian, Y.F.; Mao, Q.Q.; Wu, J.C.; Su, Z.R.; Chen, J.N.; Lai, X.P.; Ip, S.P.; Lin, Z.X. Isorhynchophylline treatment improves the amyloid-beta-induced cognitive impairment in rats via inhibition of neuronal apoptosis and tau protein hyperphosphorylation. J. Alzheimers Dis. 2014, 39, 331–346. [Google Scholar] [CrossRef]
- Li, H.Q.; Ip, S.P.; Yuan, Q.J.; Zheng, G.Q.; Tsim, K.; Dong, T.; Lin, G.; Han, Y.; Liu, Y.; Xian, Y.F.; et al. Isorhynchophylline ameliorates cognitive impairment via modulating amyloid pathology, tau hyperphosphorylation and neuroinflammation: Studies in a transgenic mouse model of Alzheimer’s disease. Brain Behav. Immun. 2019, 82, 264–278. [Google Scholar] [CrossRef]
- Xian, Y.F.; Su, Z.R.; Chen, J.N.; Lai, X.P.; Mao, Q.Q.; Cheng, C.H.; Ip, S.P.; Lin, Z.X. Isorhynchophylline improves learning and memory impairments induced by D-galactose in mice. Neurochem. Int. 2014, 76, 42–49. [Google Scholar] [CrossRef]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zhang, D.F.; Luo, R.; Wu, Y.; Zhou, H.; Kong, L.L.; Bi, R.; Yao, Y.G. A systematic integrated analysis of brain expression profiles reveals YAP1 and other prioritized hub genes as important upstream regulators in Alzheimer’s disease. Alzheimers Dement. 2018, 14, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.N.; Yang, Y.F.; Xu, W.; Yang, X.W. The Blood-Brain Barrier Permeability of Six Indole Alkaloids from Uncariae Ramulus Cum Uncis in the MDCK-pHaMDR Cell Monolayer Model. Molecules 2017, 22, 1944. [Google Scholar] [CrossRef]
- Chen, L.L.; Song, J.X.; Lu, J.H.; Yuan, Z.W.; Liu, L.F.; Durairajan, S.S.; Li, M. Corynoxine, a natural autophagy enhancer, promotes the clearance of alpha-synuclein via Akt/mTOR pathway. J. Neuroimmune Pharmacol. 2014, 9, 380–387. [Google Scholar] [CrossRef]
- Chen, L.L.; Wang, Y.B.; Song, J.X.; Deng, W.K.; Lu, J.H.; Ma, L.L.; Yang, C.B.; Li, M.; Xue, Y. Phosphoproteome-based kinase activity profiling reveals the critical role of MAP2K2 and PLK1 in neuronal autophagy. Autophagy 2017, 13, 1969–1980. [Google Scholar] [CrossRef]
- Xian, Y.F.; Lin, Z.X.; Mao, Q.Q.; Hu, Z.; Zhao, M.; Che, C.T.; Ip, S.P. Bioassay-Guided Isolation of Neuroprotective Compounds from Uncaria rhynchophylla against Beta-Amyloid-Induced Neurotoxicity. Evid. Based Complement. Alternat. Med. 2012, 2012, 802625. [Google Scholar] [CrossRef]
- Frey, H.J.; Mattila, K.M.; Korolainen, M.A.; Pirttila, T. Problems associated with biological markers of Alzheimer’s disease. Neurochem. Res. 2005, 30, 1501–1510. [Google Scholar] [CrossRef]
- Skovronsky, D.M.; Fath, S.; Lee, V.M.; Milla, M.E. Neuronal localization of the TNFalpha converting enzyme (TACE) in brain tissue and its correlation to amyloid plaques. J. Neurobiol. 2001, 49, 40–46. [Google Scholar] [CrossRef]
- Qian, M.; Shen, X.; Wang, H. The Distinct Role of ADAM17 in APP Proteolysis and Microglial Activation Related to Alzheimer’s Disease. Cell. Mol. Neurobiol. 2016, 36, 471–482. [Google Scholar] [CrossRef]
- Koelsch, G. BACE1 Function and Inhibition: Implications of Intervention in the Amyloid Pathway of Alzheimer’s Disease Pathology. Molecules 2017, 22, 1723. [Google Scholar] [CrossRef] [PubMed]
- De Strooper, B. Aph-1, Pen-2, and Nicastrin with Presenilin generate an active gamma-Secretase complex. Neuron 2003, 38, 9–12. [Google Scholar] [CrossRef]
- Ryan, N.S.; Nicholas, J.M.; Weston, P.; Liang, Y.; Lashley, T.; Guerreiro, R.; Adamson, G.; Kenny, J.; Beck, J.; Chavez-Gutierrez, L.; et al. Clinical phenotype and genetic associations in autosomal dominant familial Alzheimer’s disease: A case series. Lancet Neurol. 2016, 15, 1326–1335. [Google Scholar] [CrossRef]
- Ly, P.T.; Wu, Y.; Zou, H.; Wang, R.; Zhou, W.; Kinoshita, A.; Zhang, M.; Yang, Y.; Cai, F.; Woodgett, J.; et al. Inhibition of GSK3beta-mediated BACE1 expression reduces Alzheimer-associated phenotypes. J. Clin. Investig. 2013, 123, 224–235. [Google Scholar] [CrossRef]
- Ochalek, A.; Mihalik, B.; Avci, H.X.; Chandrasekaran, A.; Teglasi, A.; Bock, I.; Giudice, M.L.; Tancos, Z.; Molnar, K.; Laszlo, L.; et al. Neurons derived from sporadic Alzheimer’s disease iPSCs reveal elevated TAU hyperphosphorylation, increased amyloid levels, and GSK3B activation. Alzheimers Res. Ther. 2017, 9, 90. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhu, J. Entanglement of CCR5 and Alzheimer’s Disease. Front. Aging Neurosci. 2019, 11, 209. [Google Scholar] [CrossRef] [PubMed]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef] [PubMed]
- Gabbouj, S.; Ryhanen, S.; Marttinen, M.; Wittrahm, R.; Takalo, M.; Kemppainen, S.; Martiskainen, H.; Tanila, H.; Haapasalo, A.; Hiltunen, M.; et al. Altered Insulin Signaling in Alzheimer’s Disease Brain–Special Emphasis on PI3K-Akt Pathway. Front. Neurosci. 2019, 13, 629. [Google Scholar] [CrossRef] [PubMed]
- Arvanitakis, Z.; Wang, H.Y.; Capuano, A.W.; Khan, A.; Taib, B.; Anokye-Danso, F.; Schneider, J.A.; Bennett, D.A.; Ahima, R.S.; Arnold, S.E. Brain Insulin Signaling, Alzheimer Disease Pathology, and Cognitive Function. Ann. Neurol. 2020, 88, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Grimes, C.A.; Jope, R.S. The multifaceted roles of glycogen synthase kinase 3beta in cellular signaling. Prog. Neurobiol. 2001, 65, 391–426. [Google Scholar] [CrossRef]
- Cross, D.A.; Alessi, D.R.; Cohen, P.; Andjelkovich, M.; Hemmings, B.A. Inhibition of glycogen synthase kinase-3 by insulin mediated by protein kinase B. Nature 1995, 378, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Stadelmann, C.; Deckwerth, T.L.; Srinivasan, A.; Bancher, C.; Bruck, W.; Jellinger, K.; Lassmann, H. Activation of caspase-3 in single neurons and autophagic granules of granulovacuolar degeneration in Alzheimer’s disease. Evidence for apoptotic cell death. Am. J. Pathol. 1999, 155, 1459–1466. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Lipinski, C.A. Lead- and drug-like compounds: The rule-of-five revolution. Drug Discov. Today Technol. 2004, 1, 337–341. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Huang, J.; Liang, Y.; Tian, W.; Ma, J.; Huang, L.; Li, B.; Chen, R.; Li, D. Antitumor Activity and Mechanism of Robustic Acid from Dalbergia benthami Prain via Computational Target Fishing. Molecules 2020, 25, 3919. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.A.; Cheng, T.; Yu, B.; Shoemaker, B.A.; Wang, J.; Bolton, E.E.; Wang, Y.; Bryant, S.H. Literature information in PubChem: Associations between PubChem records and scientific articles. J. Cheminform. 2016, 8, 32. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Knox, C.; Guo, A.C.; Cheng, D.; Shrivastava, S.; Tzur, D.; Gautam, B.; Hassanali, M. DrugBank: A knowledgebase for drugs, drug actions and drug targets. Nucleic Acids Res. 2008, 36, D901–D906. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, S.; Li, F.; Zhou, Y.; Zhang, Y.; Wang, Z.; Zhang, R.; Zhu, J.; Ren, Y.; Tan, Y.; et al. Therapeutic target database 2020: Enriched resource for facilitating research and early development of targeted therapeutics. Nucleic Acids Res. 2020, 48, D1031–D1041. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Guo, N.; Kejariwal, A.; Thomas, P.D. PANTHER version 6: Protein sequence and function evolution data with expanded representation of biological pathways. Nucleic Acids Res. 2007, 35, D247–D252. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Bader, G.D.; Hogue, C.W. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinform. 2003, 4, 2. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Westbrook, J.; Feng, Z.; Jain, S.; Bhat, T.N.; Thanki, N.; Ravichandran, V.; Gilliland, G.L.; Bluhm, W.; Weissig, H.; Greer, D.S.; et al. The Protein Data Bank: Unifying the archive. Nucleic Acids Res. 2002, 30, 245–248. [Google Scholar] [CrossRef]
- Grosdidier, A.; Zoete, V.; Michielin, O. SwissDock, a protein-small molecule docking web service based on EADock DSS. Nucleic Acids Res. 2011, 39, W270–W277. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Formula | MW (g/mol) | Hdon | Hacc | Rbon | TPSA (Å) | LogP | LogS | Log Kp (cm/s) |
|---|---|---|---|---|---|---|---|---|---|
| Rhynchophylline | C22H28N2O4 | 384.47 | 1 | 5 | 5 | 67.87 | 2.43 | −3.51 | −7.01 |
| Corynantheine | C22H26N2O3 | 366.45 | 1 | 4 | 5 | 54.56 | 3.08 | −4.03 | −6.28 |
| Corynoxine | C22H28N2O4 | 384.47 | 1 | 5 | 5 | 67.87 | 2.38 | −3.51 | −7.01 |
| Isocorynoxeine | C22H26N2O4 | 382.45 | 1 | 5 | 5 | 67.87 | 2.26 | −3.34 | −7.17 |
| Isorhynchophylline | C22H28N2O4 | 384.47 | 1 | 5 | 5 | 67.87 | 2.4 | −3.51 | −7.01 |
| Dihydrocorynatheine | C22H28N2O3 | 368.47 | 1 | 4 | 5 | 54.56 | 3.22 | −4.21 | −6.11 |
| Hirsutine | C22H28N2O3 | 368.47 | 1 | 4 | 5 | 54.56 | 3.22 | −4.21 | −6.11 |
| Hirsuteine | C22H26N2O3 | 366.45 | 1 | 4 | 5 | 54.56 | 3.1 | −4.03 | −6.28 |
| Angustoline | C20H17N3O2 | 331.37 | 2 | 3 | 1 | 70.91 | 2.52 | −3.52 | −7.05 |
| Angustidine | C19H15N3O | 301.34 | 1 | 2 | 0 | 50.68 | 3.02 | −4.01 | −6.24 |
| Number | Gene ID | Gene Symbol | Description |
|---|---|---|---|
| 1 | 25 | ABL1 | ABL proto-oncogene 1, non-receptor tyrosine kinase |
| 2 | 43 | ACHE | acetylcholinesterase (Cartwright blood group) |
| 3 | 6868 | ADAM17 | ADAM metallopeptidase domain 17 |
| 4 | 152 | ADRA2C | adrenoceptor alpha 2C |
| 5 | 154 | ADRB2 | adrenoceptor beta 2 |
| 6 | 207 | AKT1 | AKT serine/threonine kinase 1 |
| 7 | 240 | ALOX5 | arachidonate 5-lipoxygenase |
| 8 | 83464 | APH1B | aph-1 homolog B, gamma-secretase subunit |
| 9 | 23621 | BACE1 | beta-secretase 1 |
| 10 | 590 | BCHE | butyrylcholinesterase |
| 11 | 23476 | BRD4 | bromodomain containing 4 |
| 12 | 823 | CAPN1 | calpain 1 |
| 13 | 834 | CASP1 | caspase 1 |
| 14 | 836 | CASP3 | caspase 3 |
| 15 | 595 | CCND1 | cyclin D1 |
| 16 | 729230 | CCR2 | C-C motif chemokine receptor 2 |
| 17 | 1232 | CCR3 | C-C motif chemokine receptor 3 |
| 18 | 1234 | CCR5 | C-C motif chemokine receptor 5 |
| 19 | 983 | CDK1 | cyclin dependent kinase 1 |
| 20 | 1020 | CDK5 | cyclin dependent kinase 5 |
| 21 | 1129 | CHRM2 | cholinergic receptor muscarinic 2 |
| 22 | 1131 | CHRM3 | cholinergic receptor muscarinic 3 |
| 23 | 1137 | CHRNA4 | cholinergic receptor nicotinic alpha 4 subunit |
| 24 | 1141 | CHRNB2 | cholinergic receptor nicotinic beta 2 subunit |
| 25 | 1436 | CSF1R | colony stimulating factor 1 receptor |
| 26 | 3579 | CXCR2 | C-X-C motif chemokine receptor 2 |
| 27 | 1565 | CYP2D6 | cytochrome P450 family 2 subfamily D member 6 |
| 28 | 1812 | DRD1 | dopamine receptor D1 |
| 29 | 1813 | DRD2 | dopamine receptor D2 |
| 30 | 1814 | DRD3 | dopamine receptor D3 |
| 31 | 1815 | DRD4 | dopamine receptor D4 |
| 32 | 1859 | DYRK1A | dual specificity tyrosine phosphorylation regulated kinase 1A |
| 33 | 1889 | ECE1 | endothelin converting enzyme 1 |
| 34 | 9451 | EIF2AK3 | eukaryotic translation initiation factor 2 alpha kinase 3 |
| 35 | 2064 | ERBB2 | erb-b2 receptor tyrosine kinase 2 |
| 36 | 2078 | ERG | ETS transcription factor ERG |
| 37 | 2260 | FGFR1 | fibroblast growth factor receptor 1 |
| 38 | 2261 | FGFR3 | fibroblast growth factor receptor 3 |
| 39 | 2534 | FYN | FYN proto-oncogene, Src family tyrosine kinase |
| 40 | 2558 | GABRA5 | gamma-aminobutyric acid type A receptor subunit alpha5 |
| 41 | 51738 | GHRL | ghrelin and obestatin prepropeptide |
| 42 | 2904 | GRIN2B | glutamate ionotropic receptor NMDA type subunit 2B |
| 43 | 2932 | GSK3B | glycogen synthase kinase 3 beta |
| 44 | 3091 | HIF1A | hypoxia inducible factor 1 subunit alpha |
| 45 | 3290 | HSD11B1 | hydroxysteroid 11-beta dehydrogenase 1 |
| 46 | 3350 | HTR1A | 5-hydroxytryptamine receptor 1A |
| 47 | 3356 | HTR2A | 5-hydroxytryptamine receptor 2A |
| 48 | 3358 | HTR2C | 5-hydroxytryptamine receptor 2C |
| 49 | 3360 | HTR4 | 5-hydroxytryptamine receptor 4 |
| 50 | 3362 | HTR6 | 5-hydroxytryptamine receptor 6 |
| 51 | 3363 | HTR7 | 5-hydroxytryptamine receptor 7 |
| 52 | 3480 | IGF1R | insulin like growth factor 1 receptor |
| 53 | 3643 | INSR | insulin receptor |
| 54 | 3832 | KIF11 | kinesin family member 11 |
| 55 | 3815 | KIT | KIT proto-oncogene, receptor tyrosine kinase |
| 56 | 3932 | LCK | LCK proto-oncogene, Src family tyrosine kinase |
| 57 | 120892 | LRRK2 | leucine rich repeat kinase 2 |
| 58 | 4128 | MAOA | monoamine oxidase A |
| 59 | 5602 | MAPK10 | mitogen-activated protein kinase 10 |
| 60 | 10461 | MERTK | MER proto-oncogene, tyrosine kinase |
| 61 | 4233 | MET | MET proto-oncogene, receptor tyrosine kinase |
| 62 | 4311 | MME | membrane metalloendopeptidase |
| 63 | 4313 | MMP2 | matrix metallopeptidase 2 |
| 64 | 4314 | MMP3 | matrix metallopeptidase 3 |
| 65 | 4318 | MMP9 | matrix metallopeptidase 9 |
| 66 | 2475 | MTOR | mechanistic target of rapamycin kinase |
| 67 | 23385 | NCSTN | nicastrin |
| 68 | 4842 | NOS1 | nitric oxide synthase 1 |
| 69 | 4843 | NOS2 | nitric oxide synthase 2 |
| 70 | 4846 | NOS3 | nitric oxide synthase 3 |
| 71 | 4835 | NQO2 | N-ribosyldihydronicotinamide:quinone reductase 2 |
| 72 | 4914 | NTRK1 | neurotrophic receptor tyrosine kinase 1 |
| 73 | 4915 | NTRK2 | neurotrophic receptor tyrosine kinase 2 |
| 74 | 4916 | NTRK3 | neurotrophic receptor tyrosine kinase 3 |
| 75 | 5027 | P2RX7 | purinergic receptor P2X 7 |
| 76 | 142 | PARP1 | poly(ADP-ribose) polymerase 1 |
| 77 | 5144 | PDE4D | phosphodiesterase 4D |
| 78 | 8622 | PDE8B | phosphodiesterase 8B |
| 79 | 5159 | PDGFRB | platelet derived growth factor receptor beta |
| 80 | 7941 | PLA2G7 | phospholipase A2 group VII |
| 81 | 5663 | PSEN1 | presenilin 1 |
| 82 | 5664 | PSEN2 | presenilin 2 |
| 83 | 55851 | PSENEN | presenilin enhancer, gamma-secretase subunit |
| 84 | 5743 | PTGS2 | prostaglandin-endoperoxide synthase 2 |
| 85 | 6571 | SLC18A2 | solute carrier family 18 member A2 |
| 86 | 6531 | SLC6A3 | solute carrier family 6 member 3 |
| 87 | 6532 | SLC6A4 | solute carrier family 6 member 4 |
| 88 | 6752 | SSTR2 | somatostatin receptor 2 |
| 89 | 6753 | SSTR3 | somatostatin receptor 3 |
| 90 | 6755 | SSTR5 | somatostatin receptor 5 |
| Term | Pathway | Rich Factor | p-Value | Count | Symbols |
|---|---|---|---|---|---|
| hsa05010 | Alzheimer disease | 0.08 | 2.445 × 10−18 | 16 | CAPN1,CASP3,CDK5,GRIN2B,GSK3B,MME,NOS1,PSEN1,PSEN2,PTGS2,ADAM17,EIF2AK3,NCSTN,BACE1,PSENEN,APH1B |
| hsa04726 | Serotonergic synapse | 0.11 | 9.421 × 10−17 | 13 | ALOX5,CASP3,CYP2D6,HTR1A,HTR2A,HTR2C,HTR4,HTR6,HTR7,MAOA,PTGS2,SLC6A4,SLC18A2 |
| hsa04020 | Calcium signaling pathway | 0.07 | 1.693 × 10−16 | 15 | ADRB2,CHRM2,CHRM3,DRD1,ERBB2,HTR2A,HTR2C,HTR4,HTR6,HTR7,NOS1,NOS2,NOS3,P2RX7,PDGFRB |
| hsa04151 | PI3K-Akt signaling pathway | 0.04 | 4.79 × 10−15 | 17 | AKT1,CCND1,CHRM2,CSF1R,ERBB2,FGFR1,FGFR3,MTOR,GSK3B,IGF1R,INSR,KIT,MET,NOS3,NTRK1,NTRK2,PDGFRB |
| hsa04728 | Dopaminergic synapse | 0.08 | 5.086 × 10−13 | 11 | AKT1,DRD1,DRD2,DRD3,DRD4,GRIN2B,GSK3B,MAOA,MAPK10,SLC6A3,SLC18A2 |
| hsa04722 | Neurotrophin signaling pathway | 0.07 | 2.555 × 10−10 | 9 | ABL1,AKT1,GSK3B,NTRK1,NTRK2,NTRK3,MAPK10,PSEN1,PSEN2 |
| hsa04330 | Notch signaling pathway | 0.12 | 1.685 × 10−8 | 6 | PSEN1,PSEN2,ADAM17,NCSTN,PSENEN,APH1B |
| hsa04725 | Cholinergic synapse | 0.06 | 9.258 × 10−8 | 7 | ACHE,AKT1,CHRM2,CHRM3,CHRNA4,CHRNB2,FYN |
| hsa04144 | Endocytosis | 0.03 | 2.33 × 10−7 | 9 | ADRB2,CCR5,CSF1R,FGFR3,IGF1R,CXCR2,KIT,MET,NTRK1 |
| hsa04520 | Adherens junction | 0.07 | 2.494 × 10−7 | 6 | ERBB2,FGFR1,FYN,IGF1R,INSR,MET |
| hsa04926 | Relaxin signaling pathway | 0.05 | 2.92 × 10−7 | 7 | AKT1,MMP2,MMP9,NOS1,NOS2,NOS3,MAPK10 |
| hsa04540 | Gap junction | 0.07 | 4.675 × 10−7 | 6 | CDK1,DRD1,DRD2,HTR2A,HTR2C,PDGFRB |
| hsa04210 | Apoptosis | 0.05 | 5.935 × 10−7 | 7 | PARP1,AKT1,CAPN1,CASP3,NTRK1,MAPK10,EIF2AK3 |
| hsa04012 | ErbB signaling pathway | 0.06 | 6.051 × 10−7 | 6 | ABL1,AKT1,ERBB2,MTOR,GSK3B,MAPK10 |
| hsa04657 | IL-17 signaling pathway | 0.06 | 6.442 × 10−7 | 6 | CASP3,GSK3B,MMP3,MMP9,MAPK10,PTGS2 |
| hsa04371 | Apelin signaling pathway | 0.04 | 7.882 × 10−6 | 6 | AKT1,CCND1,MTOR,NOS1,NOS2,NOS3 |
| hsa05014 | Amyotrophic lateral sclerosis | 0.07 | 3.464 × 10−5 | 4 | CASP1,CASP3,GRIN2B,NOS1 |
| hsa05032 | Morphine addiction | 0.04 | 0.0002249 | 4 | DRD1,GABRA5,PDE4D,PDE8B |
| hsa04360 | Axon guidance | 0.03 | 0.0004339 | 5 | ABL1,CDK5,FYN,GSK3B,MET |
| hsa04115 | p53 signaling pathway | 0.04 | 0.00213 | 3 | CCND1,CASP3,CDK1 |
| Ligands | Target | PDB | deltaG (kcal/mol) | deltaGvdw | FullFitness (kcal/mol) | Energy (kcal/mol) |
|---|---|---|---|---|---|---|
| Corynoxine | CCR5 | 6AKY | −9.28 | −83.41 | −1260.02 | 16.61 |
| Corynoxine | MMP3 | 2D1O | −8.52 | −70.64 | −866.94 | 16.06 |
| Corynoxine | CCND1 | 2W9F | −8.17 | −66.74 | −1571.13 | 21.40 |
| Corynoxine | NTRK2 | 4AT4 | −8.10 | −56.35 | −1692.15 | 1.58 |
| Corynoxine | GRIN2B | 7KL4 | −7.95 | −76.52 | −1938.77 | 29.92 |
| Corynoxine | CHRM2 | 4MQT | −7.92 | −51.24 | −874.59 | 14.59 |
| Corynoxine | CDK5 | 1UNH | −7.90 | −54.59 | −3964.05 | 20.41 |
| Corynoxine | MMP2 | 1QIB | −7.67 | −54.18 | −1258.85 | 16.61 |
| Corynoxine | GSK3B | 2O5K | −7.66 | −62.37 | −1986.98 | 17.42 |
| Corynantheine | CHRM2 | 4MQT | −9.38 | −82.53 | −860.06 | −5.88 |
| Corynantheine | MMP3 | 2D1O | −9.30 | −82.62 | −857.66 | −0.88 |
| Corynantheine | MMP2 | 1QIB | −9.24 | −94.70 | −1246.61 | −5.34 |
| Corynantheine | GRIN2B | 7KL4 | −9.02 | −87.46 | −1941.76 | −5.64 |
| Corynantheine | CCR5 | 6AKY | −8.92 | −75.97 | −1250.69 | 1.62 |
| Corynantheine | CCND1 | 2W9F | −8.77 | −83.19 | −1556.05 | 8.76 |
| Corynantheine | NTRK2 | 4AT4 | −8.10 | −53.43 | −1676.61 | −3.78 |
| Corynantheine | CDK5 | 1UNH | −7.95 | −61.20 | −3952.57 | −1.90 |
| Corynantheine | GSK3B | 2O5K | −7.38 | −56.22 | −1952.87 | 24.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, P.; Wang, X.-M.; Ye, C.-Y.; Su, H.-F.; Tian, Q. The Main Alkaloids in Uncaria rhynchophylla and Their Anti-Alzheimer’s Disease Mechanism Determined by a Network Pharmacology Approach. Int. J. Mol. Sci. 2021, 22, 3612. https://doi.org/10.3390/ijms22073612
Zeng P, Wang X-M, Ye C-Y, Su H-F, Tian Q. The Main Alkaloids in Uncaria rhynchophylla and Their Anti-Alzheimer’s Disease Mechanism Determined by a Network Pharmacology Approach. International Journal of Molecular Sciences. 2021; 22(7):3612. https://doi.org/10.3390/ijms22073612
Chicago/Turabian StyleZeng, Peng, Xiao-Ming Wang, Chao-Yuan Ye, Hong-Fei Su, and Qing Tian. 2021. "The Main Alkaloids in Uncaria rhynchophylla and Their Anti-Alzheimer’s Disease Mechanism Determined by a Network Pharmacology Approach" International Journal of Molecular Sciences 22, no. 7: 3612. https://doi.org/10.3390/ijms22073612
APA StyleZeng, P., Wang, X.-M., Ye, C.-Y., Su, H.-F., & Tian, Q. (2021). The Main Alkaloids in Uncaria rhynchophylla and Their Anti-Alzheimer’s Disease Mechanism Determined by a Network Pharmacology Approach. International Journal of Molecular Sciences, 22(7), 3612. https://doi.org/10.3390/ijms22073612