
Astrocyte Ca2+ Waves and Subsequent Non-Synchronized Ca2+ Oscillations Coincide with Arteriole Diameter Changes in Response to Spreading Depolarization

 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
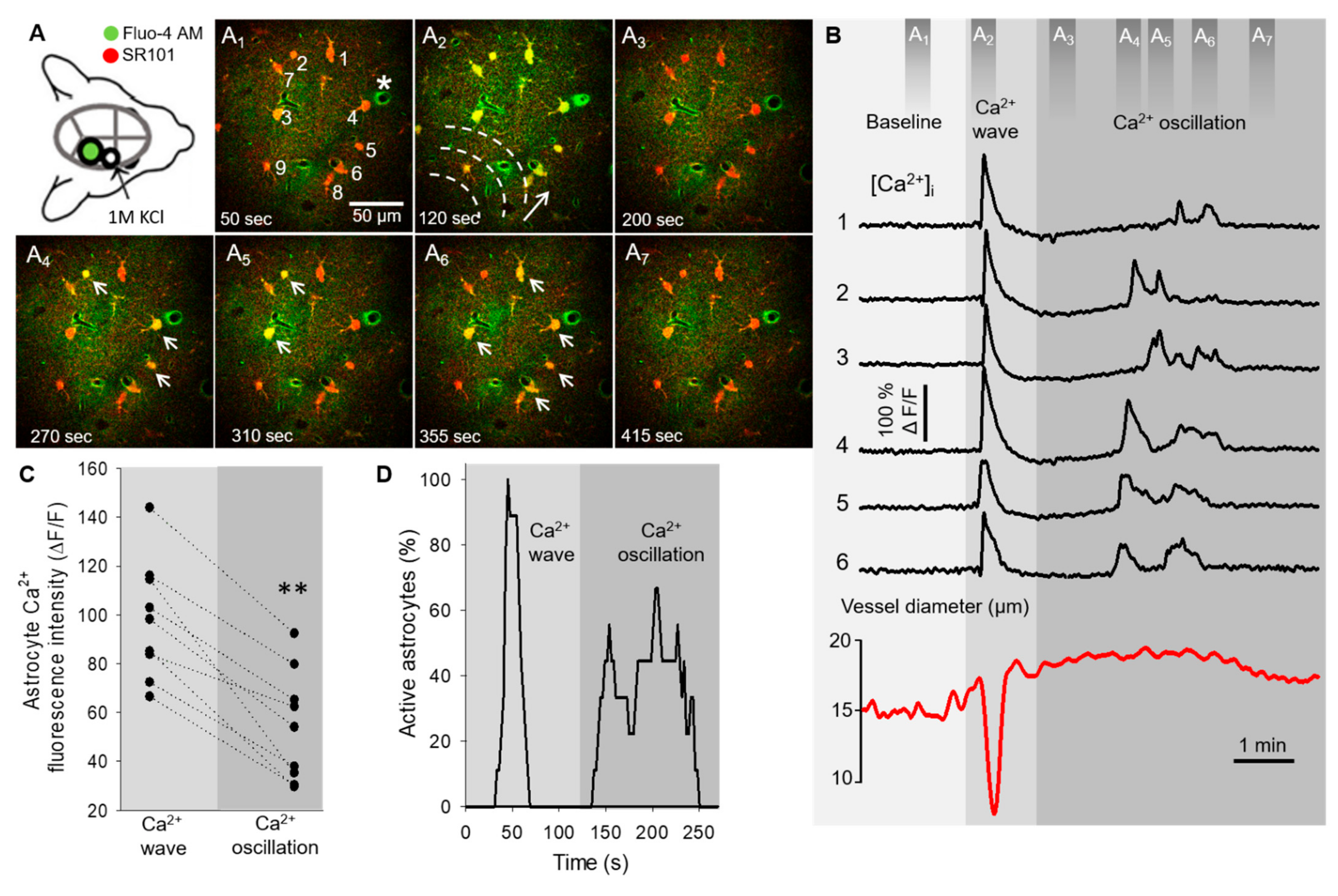
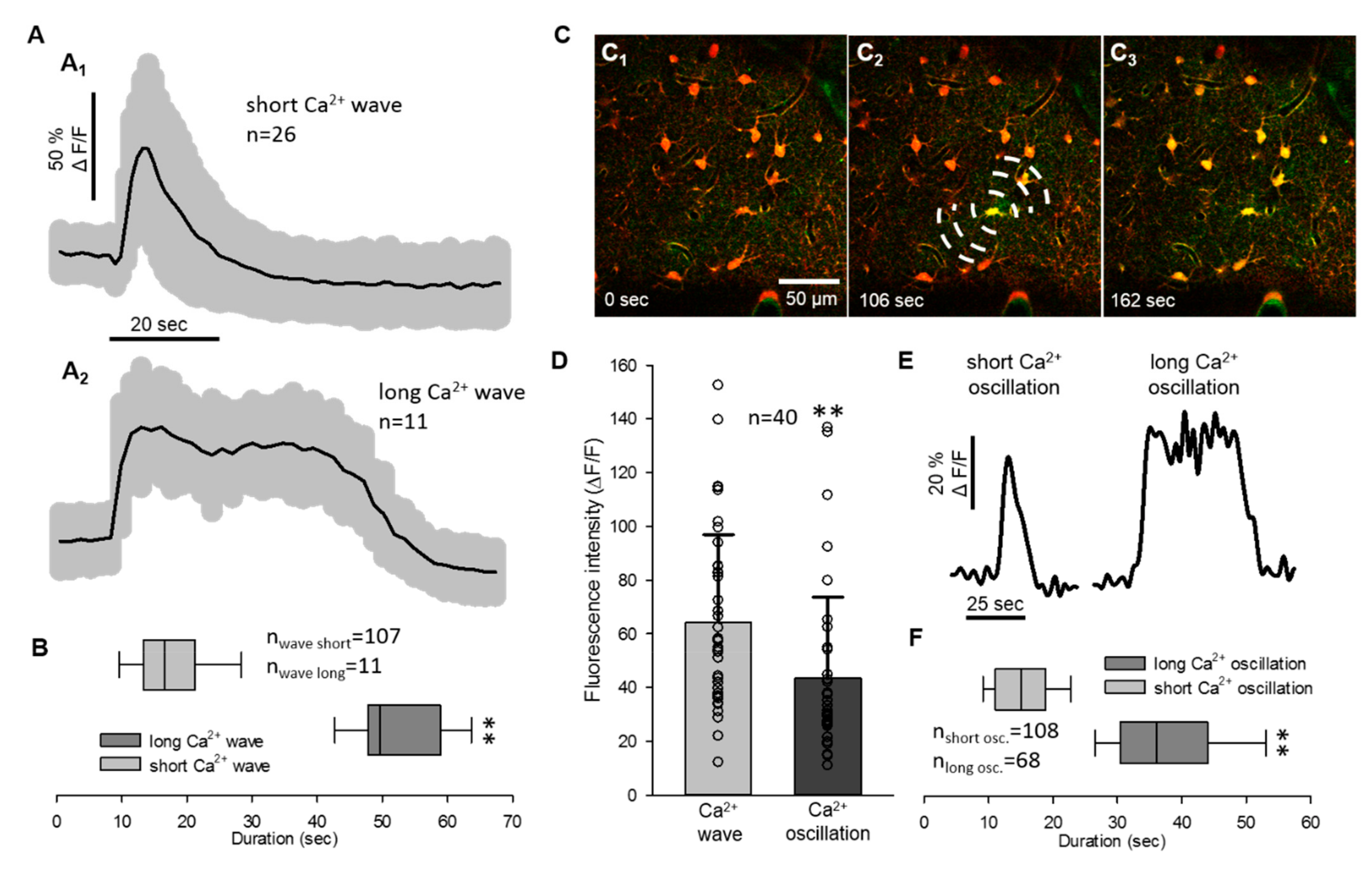
2.1. Non-Synchronized Astrocyte Ca2+ Oscillations Accompany the Concurrent Ca2+ Wave of Spreading Depolarization in the Mouse Somatosensory Cortex
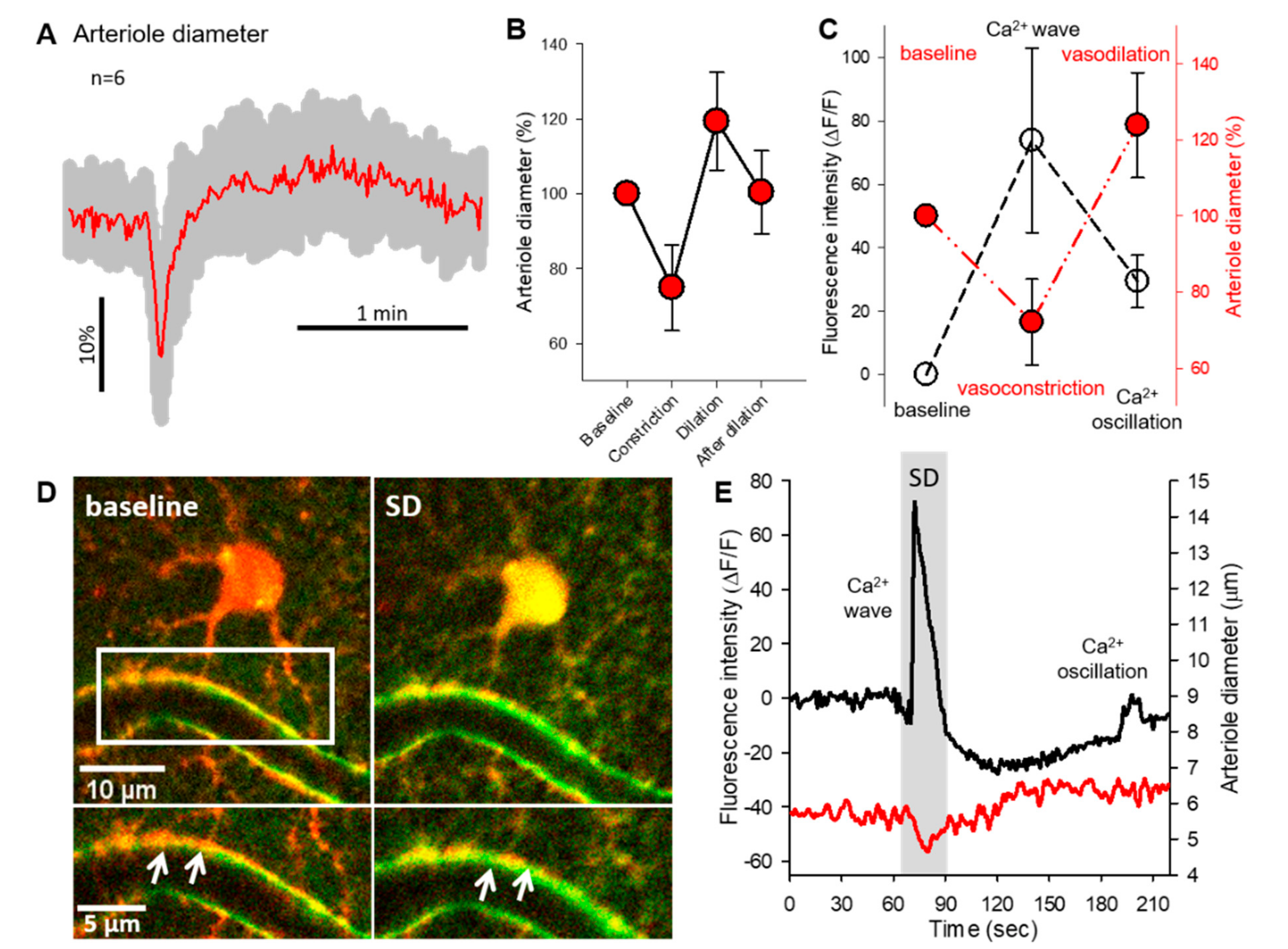
2.2. Astrocyte Ca2+ Waves Coincide with Arteriolar Constrictions While Ca2+ Oscillations Occur during Vasodilation
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Multiphoton Microscopy
4.3. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leao, A.A. Further observations on the spreading depression of activity in the cerebral cortex. J. Neurophysiol. 1947, 10, 409–414. [Google Scholar] [CrossRef]
- Somjen, G.G. Mechanisms of spreading depression and hypoxic spreading depression-like depolarization. Physiol. Rev. 2001, 81, 1065–1096. [Google Scholar] [CrossRef] [PubMed]
- von Bornstadt, D.; Houben, T.; Seidel, J.L.; Zheng, Y.; Dilekoz, E.; Qin, T.; Sandow, N.; Kura, S.; Eikermann-Haerter, K.; Endres, M.; et al. Supply-demand mismatch transients in susceptible peri-infarct hot zones explain the origins of spreading injury depolarizations. Neuron 2015, 85, 1117–1131. [Google Scholar] [CrossRef] [PubMed]
- Ayata, C.; Lauritzen, M. Spreading Depression, Spreading Depolarizations, and the Cerebral Vasculature. Physiol. Rev. 2015, 95, 953–993. [Google Scholar] [CrossRef] [PubMed]
- Dreier, J.P. The role of spreading depression, spreading depolarization and spreading ischemia in neurological disease. Nat. Med. 2011, 17, 439–447. [Google Scholar] [CrossRef]
- Hoffmann, U.; Ayata, C. Neurovascular coupling during spreading depolarizations. Acta Neurochir. Suppl. 2013, 115, 161–165. [Google Scholar] [CrossRef]
- Bere, Z.; Obrenovitch, T.P.; Bari, F.; Farkas, E. Ischemia-induced depolarizations and associated hemodynamic responses in incomplete global forebrain ischemia in rats. Neuroscience 2014, 260, 217–226. [Google Scholar] [CrossRef]
- Dreier, J.P.; Korner, K.; Ebert, N.; Gorner, A.; Rubin, I.; Back, T.; Lindauer, U.; Wolf, T.; Villringer, A.; Einhaupl, K.M.; et al. Nitric oxide scavenging by hemoglobin or nitric oxide synthase inhibition by N-nitro-L-arginine induces cortical spreading ischemia when K+ is increased in the subarachnoid space. J. Cereb. Blood. Flow. Metab. 1998, 18, 978–990. [Google Scholar] [CrossRef] [PubMed]
- Hartings, J.A.; Shuttleworth, C.W.; Kirov, S.A.; Ayata, C.; Hinzman, J.M.; Foreman, B.; Andrew, R.D.; Boutelle, M.G.; Brennan, K.C.; Carlson, A.P.; et al. The continuum of spreading depolarizations in acute cortical lesion development: Examining Leao’s legacy. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2016. [Google Scholar] [CrossRef]
- Fellin, T.; Pascual, O.; Gobbo, S.; Pozzan, T.; Haydon, P.G.; Carmignoto, G. Neuronal synchrony mediated by astrocytic glutamate through activation of extrasynaptic NMDA receptors. Neuron 2004, 43, 729–743. [Google Scholar] [CrossRef]
- Perea, G.; Araque, A. Astrocytes potentiate transmitter release at single hippocampal synapses. Science 2007, 317, 1083–1086. [Google Scholar] [CrossRef]
- Bazargani, N.; Attwell, D. Astrocyte calcium signaling: The third wave. Nat. Neurosci. 2016, 19, 182–189. [Google Scholar] [CrossRef]
- Filosa, J.A.; Morrison, H.W.; Iddings, J.A.; Du, W.; Kim, K.J. Beyond neurovascular coupling, role of astrocytes in the regulation of vascular tone. Neuroscience 2016, 323, 96–109. [Google Scholar] [CrossRef]
- Otsu, Y.; Couchman, K.; Lyons, D.G.; Collot, M.; Agarwal, A.; Mallet, J.M.; Pfrieger, F.W.; Bergles, D.E.; Charpak, S. Calcium dynamics in astrocyte processes during neurovascular coupling. Nat. Neurosci. 2015, 18, 210–218. [Google Scholar] [CrossRef]
- Lind, B.L.; Jessen, S.B.; Lonstrup, M.; Josephine, C.; Bonvento, G.; Lauritzen, M. Fast Ca(2+) responses in astrocyte end-feet and neurovascular coupling in mice. GLIA 2018, 66, 348–358. [Google Scholar] [CrossRef]
- Koide, M.; Bonev, A.D.; Nelson, M.T.; Wellman, G.C. Inversion of neurovascular coupling by subarachnoid blood depends on large-conductance Ca2+-activated K+ (BK) channels. Proc. Natl. Acad. Sci. USA 2012, 109, E1387–E1395. [Google Scholar] [CrossRef]
- Pappas, A.C.; Koide, M.; Wellman, G.C. Astrocyte Ca2+ Signaling Drives Inversion of Neurovascular Coupling after Subarachnoid Hemorrhage. J. Neurosci. 2015, 35, 13375–13384. [Google Scholar] [CrossRef]
- Gordon, G.R.; Choi, H.B.; Rungta, R.L.; Ellis-Davies, G.C.; MacVicar, B.A. Brain metabolism dictates the polarity of astrocyte control over arterioles. Nature 2008, 456, 745–749. [Google Scholar] [CrossRef] [PubMed]
- King, C.M.; Bohmbach, K.; Minge, D.; Delekate, A.; Zheng, K.; Reynolds, J.; Rakers, C.; Zeug, A.; Petzold, G.C.; Rusakov, D.A.; et al. Local Resting Ca(2+) Controls the Scale of Astroglial Ca(2+) Signals. Cell. Rep. 2020, 30, 3466–3477.e4. [Google Scholar] [CrossRef] [PubMed]
- Attwell, D.; Buchan, A.M.; Charpak, S.; Lauritzen, M.; Macvicar, B.A.; Newman, E.A. Glial and neuronal control of brain blood flow. Nature 2010, 468, 232–243. [Google Scholar] [CrossRef]
- Howarth, C. The contribution of astrocytes to the regulation of cerebral blood flow. Front. Neurosci. 2014, 8, 103. [Google Scholar] [CrossRef]
- Fordsmann, J.C.; Ko, R.W.; Choi, H.B.; Thomsen, K.; Witgen, B.M.; Mathiesen, C.; Lonstrup, M.; Piilgaard, H.; MacVicar, B.A.; Lauritzen, M. Increased 20-HETE synthesis explains reduced cerebral blood flow but not impaired neurovascular coupling after cortical spreading depression in rat cerebral cortex. J. Neurosci. 2013, 33, 2562–2570. [Google Scholar] [CrossRef]
- Varga, D.P.; Puskas, T.; Menyhart, A.; Hertelendy, P.; Zolei-Szenasi, D.; Toth, R.; Ivankovits-Kiss, O.; Bari, F.; Farkas, E. Contribution of prostanoid signaling to the evolution of spreading depolarization and the associated cerebral blood flow response. Sci. Rep. 2016, 6, 31402. [Google Scholar] [CrossRef]
- Menyhart, A.; Farkas, A.E.; Varga, D.P.; Frank, R.; Toth, R.; Balint, A.R.; Makra, P.; Dreier, J.P.; Bari, F.; Krizbai, I.A.; et al. Large-conductance Ca(2+)-activated potassium channels are potently involved in the inverse neurovascular response to spreading depolarization. Neurobiol. Dis. 2018, 119, 41–52. [Google Scholar] [CrossRef]
- Varga, D.P.; Szabo, I.; Varga, V.E.; Tóth, O.M.; Kozma, M.; Bálint, A.R.; Krizbai, I.A.; Bari, F.; Farakas, E. The antagonism of prostaglandin FP receptors inhibits the evolution of spreading depolarization in an experimental model of global forebrain ischemia. Neurobiol. Dis. 2020, 137, 104780. [Google Scholar] [CrossRef]
- Basarsky, T.A.; Duffy, S.N.; Andrew, R.D.; MacVicar, B.A. Imaging spreading depression and associated intracellular calcium waves in brain slices. J. Neurosci. 1998, 18, 7189–7199. [Google Scholar] [CrossRef]
- Peters, O.; Schipke, C.G.; Hashimoto, Y.; Kettenmann, H. Different mechanisms promote astrocyte Ca2+ waves and spreading depression in the mouse neocortex. J. Neurosci. 2003, 23, 9888–9896. [Google Scholar] [CrossRef]
- Chuquet, J.; Hollender, L.; Nimchinsky, E.A. High-resolution in vivo imaging of the neurovascular unit during spreading depression. J. Neurosci. 2007, 27, 4036–4044. [Google Scholar] [CrossRef]
- Enger, R.; Tang, W.; Vindedal, G.F.; Jensen, V.; Johannes Helm, P.; Sprengel, R.; Looger, L.L.; Nagelhus, E.A. Dynamics of Ionic Shifts in Cortical Spreading Depression. Cereb. Cortex 2015, 25, 4469–4476. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.C.; Chen, R.Y.; Cheng, T.C.; Chiang, Y.C.; Shen, M.L.; Hsu, L.L.; Zhou, N. Spreading Depression Promotes Astrocytic Calcium Oscillations and Enhances Gliotransmission to Hippocampal Neurons. Cereb. Cortex. 2018, 28, 3204–3216. [Google Scholar] [CrossRef]
- Hassinger, T.D.; Guthrie, P.B.; Atkinson, P.B.; Bennett, M.V.; Kater, S.B. An extracellular signaling component in propagation of astrocytic calcium waves. Proc. Natl. Acad. Sci. USA 1996, 93, 13268–13273. [Google Scholar] [CrossRef] [PubMed]
- Newman, E.A.; Zahs, K.R. Calcium waves in retinal glial cells. Science 1997, 275, 844–847. [Google Scholar] [CrossRef]
- Dani, J.W.; Chernjavsky, A.; Smith, S.J. Neuronal activity triggers calcium waves in hippocampal astrocyte networks. Neuron 1992, 8, 429–440. [Google Scholar] [CrossRef]
- Parri, H.R.; Gould, T.M.; Crunelli, V. Spontaneous astrocytic Ca2+ oscillations in situ drive NMDAR-mediated neuronal excitation. Nat. Neurosci. 2001, 4, 803–812. [Google Scholar] [CrossRef]
- Berger, M.; Speckmann, E.J.; Pape, H.C.; Gorji, A. Spreading depression enhances human neocortical excitability in vitro. Cephalalgia 2008, 28, 558–562. [Google Scholar] [CrossRef]
- Mulligan, S.J.; MacVicar, B.A. Calcium transients in astrocyte endfeet cause cerebrovascular constrictions. Nature 2004, 431, 195–199. [Google Scholar] [CrossRef]
- Seidel, J.L.; Escartin, C.; Ayata, C.; Bonvento, G.; Shuttleworth, C.W. Multifaceted roles for astrocytes in spreading depolarization: A target for limiting spreading depolarization in acute brain injury? GLIA 2016, 64, 5–20. [Google Scholar] [CrossRef]
- Nimmerjahn, A.; Helmchen, F. In vivo labeling of cortical astrocytes with sulforhodamine 101 (SR101). Cold. Spring. Harb. Protoc. 2012, 2012, 326–334. [Google Scholar] [CrossRef]
- Nimmerjahn, A.; Kirchhoff, F.; Kerr, J.N.; Helmchen, F. Sulforhodamine 101 as a specific marker of astroglia in the neocortex in vivo. Nat. Methods 2004, 1, 31–37. [Google Scholar] [CrossRef]
- Schnell, C.; Hagos, Y.; Hulsmann, S. Active sulforhodamine 101 uptake into hippocampal astrocytes. PLoS ONE 2012, 7, e49398. [Google Scholar] [CrossRef]
- Hulsmann, S.; Hagos, L.; Heuer, H.; Schnell, C. Limitations of Sulforhodamine 101 for Brain Imaging. Front. Cell. Neurosci. 2017, 11, 44. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.S.; Kalloniatis, M.; Truong, H.T.; Binder, M.D.; Cate, H.S.; Kilpatrick, T.J.; Hammond, V.E. Oligodendrocyte positioning in cerebral cortex is independent of projection neuron layering. GLIA. 2009, 57, 1024–1030. [Google Scholar] [CrossRef] [PubMed]
- Hotaling, N.A.; Bharti, K.; Kriel, H.; Simon, C.G., Jr. DiameterJ. A validated open source nanofiber diameter measurement tool. Biomaterials 2015, 61, 327–338. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tóth, R.; Farkas, A.E.; Krizbai, I.A.; Makra, P.; Bari, F.; Farkas, E.; Menyhárt, Á. Astrocyte Ca2+ Waves and Subsequent Non-Synchronized Ca2+ Oscillations Coincide with Arteriole Diameter Changes in Response to Spreading Depolarization. Int. J. Mol. Sci. 2021, 22, 3442. https://doi.org/10.3390/ijms22073442
Tóth R, Farkas AE, Krizbai IA, Makra P, Bari F, Farkas E, Menyhárt Á. Astrocyte Ca2+ Waves and Subsequent Non-Synchronized Ca2+ Oscillations Coincide with Arteriole Diameter Changes in Response to Spreading Depolarization. International Journal of Molecular Sciences. 2021; 22(7):3442. https://doi.org/10.3390/ijms22073442
Chicago/Turabian StyleTóth, Réka, Attila E. Farkas, István A. Krizbai, Péter Makra, Ferenc Bari, Eszter Farkas, and Ákos Menyhárt. 2021. "Astrocyte Ca2+ Waves and Subsequent Non-Synchronized Ca2+ Oscillations Coincide with Arteriole Diameter Changes in Response to Spreading Depolarization" International Journal of Molecular Sciences 22, no. 7: 3442. https://doi.org/10.3390/ijms22073442
APA StyleTóth, R., Farkas, A. E., Krizbai, I. A., Makra, P., Bari, F., Farkas, E., & Menyhárt, Á. (2021). Astrocyte Ca2+ Waves and Subsequent Non-Synchronized Ca2+ Oscillations Coincide with Arteriole Diameter Changes in Response to Spreading Depolarization. International Journal of Molecular Sciences, 22(7), 3442. https://doi.org/10.3390/ijms22073442