
Induction of PLXNA4 Gene during Neural Differentiation in Human Umbilical-Cord-Derived Mesenchymal Stem Cells by Low-Intensity Sub-Sonic Vibration


Abstract
:1. Introduction
2. Results
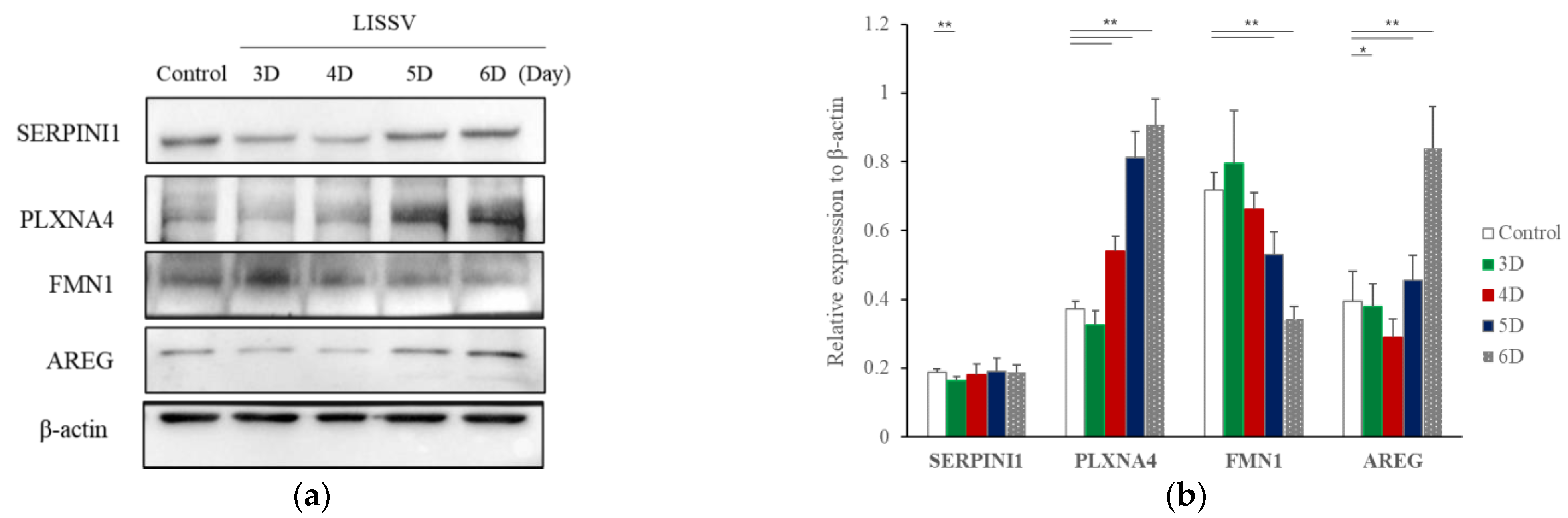
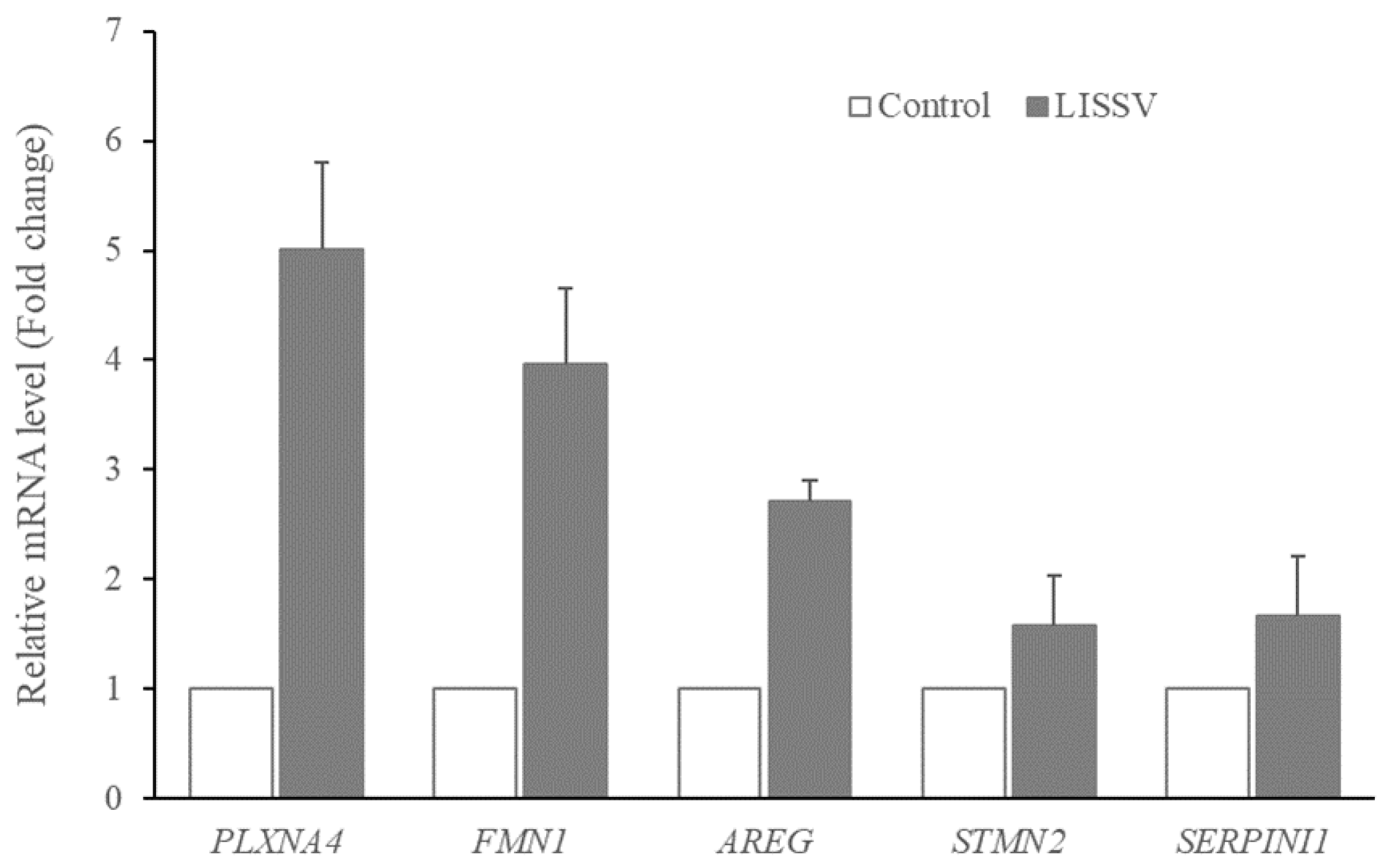
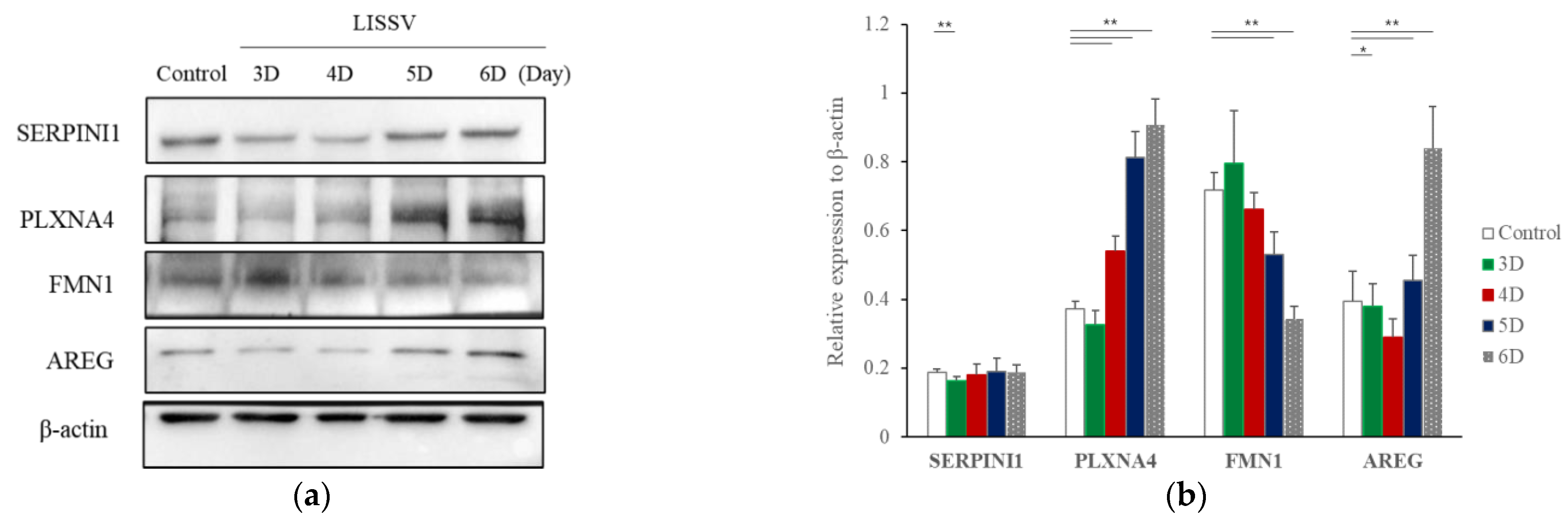
2.1. RNA Sequencing Confirmed That Six Genes Were Increased by LISSV
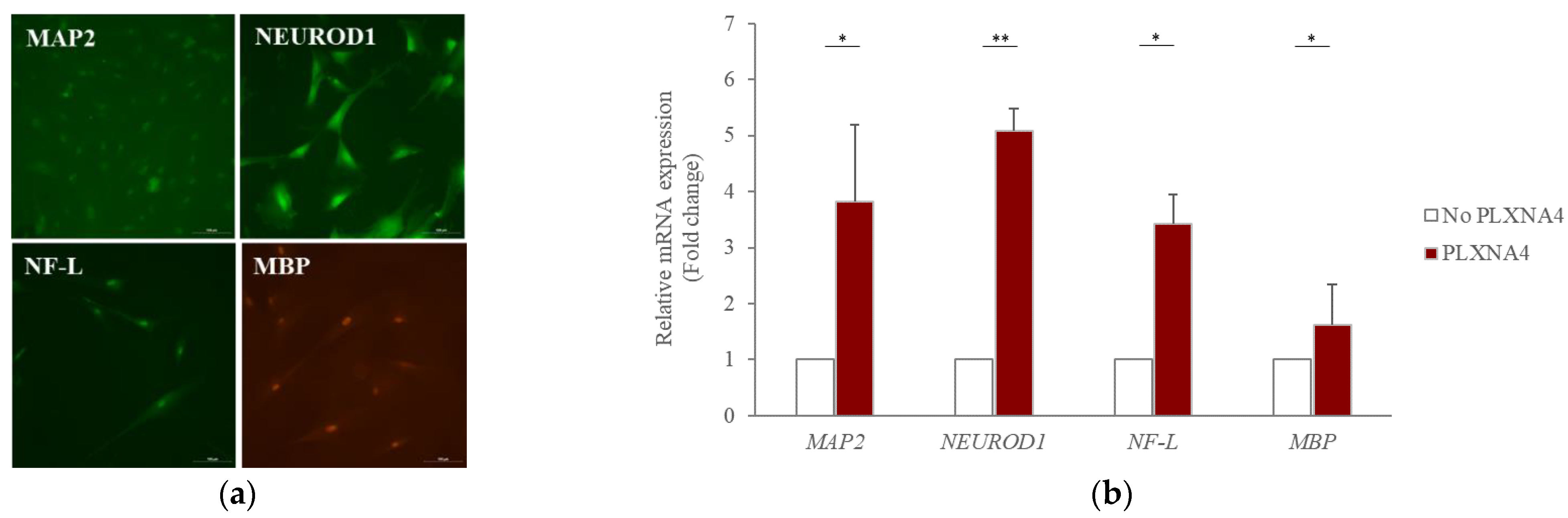
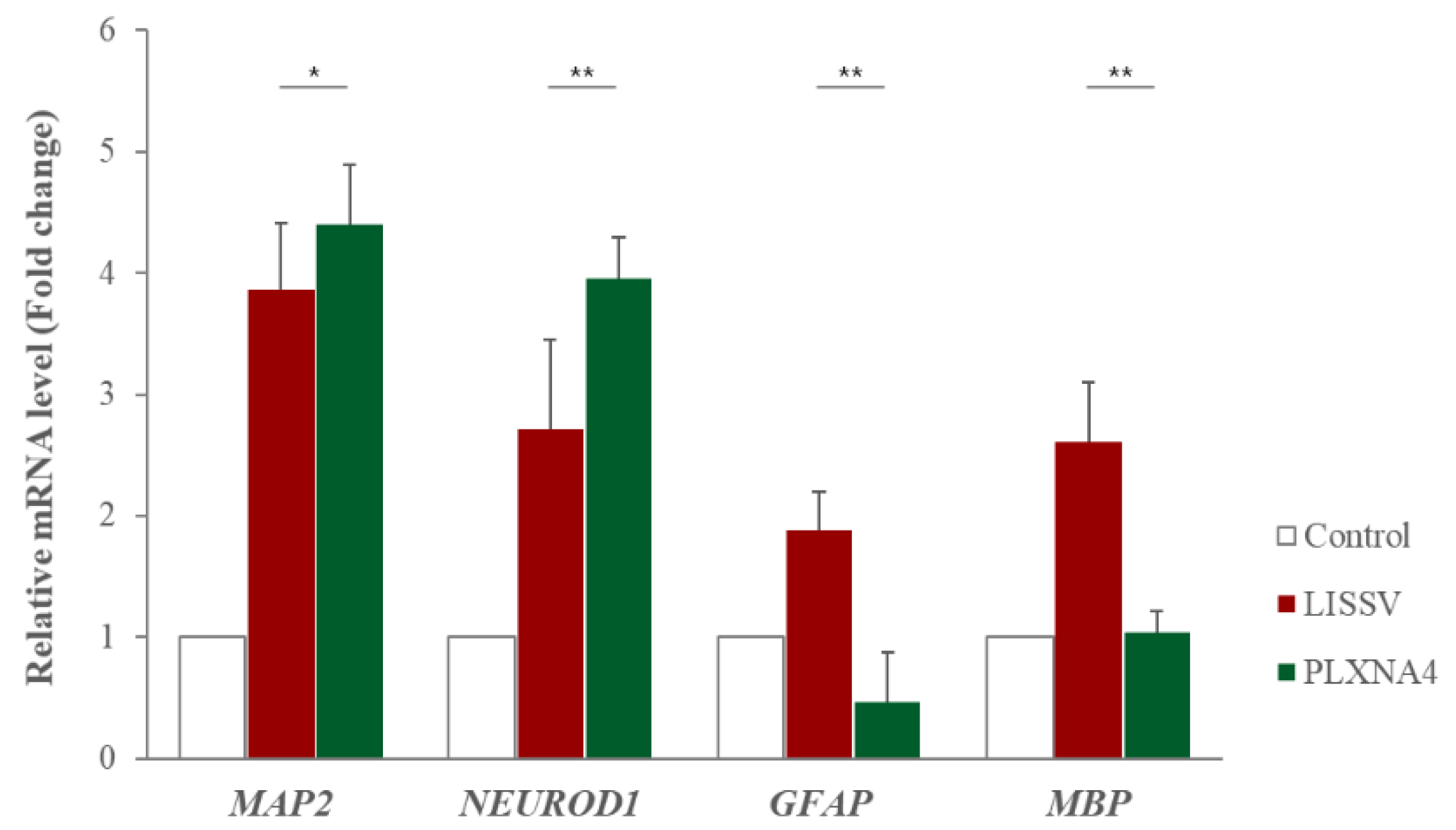
2.2. Recombinant PLXNA4 Protein Affected Neural Differentiation of hUC-MSCs
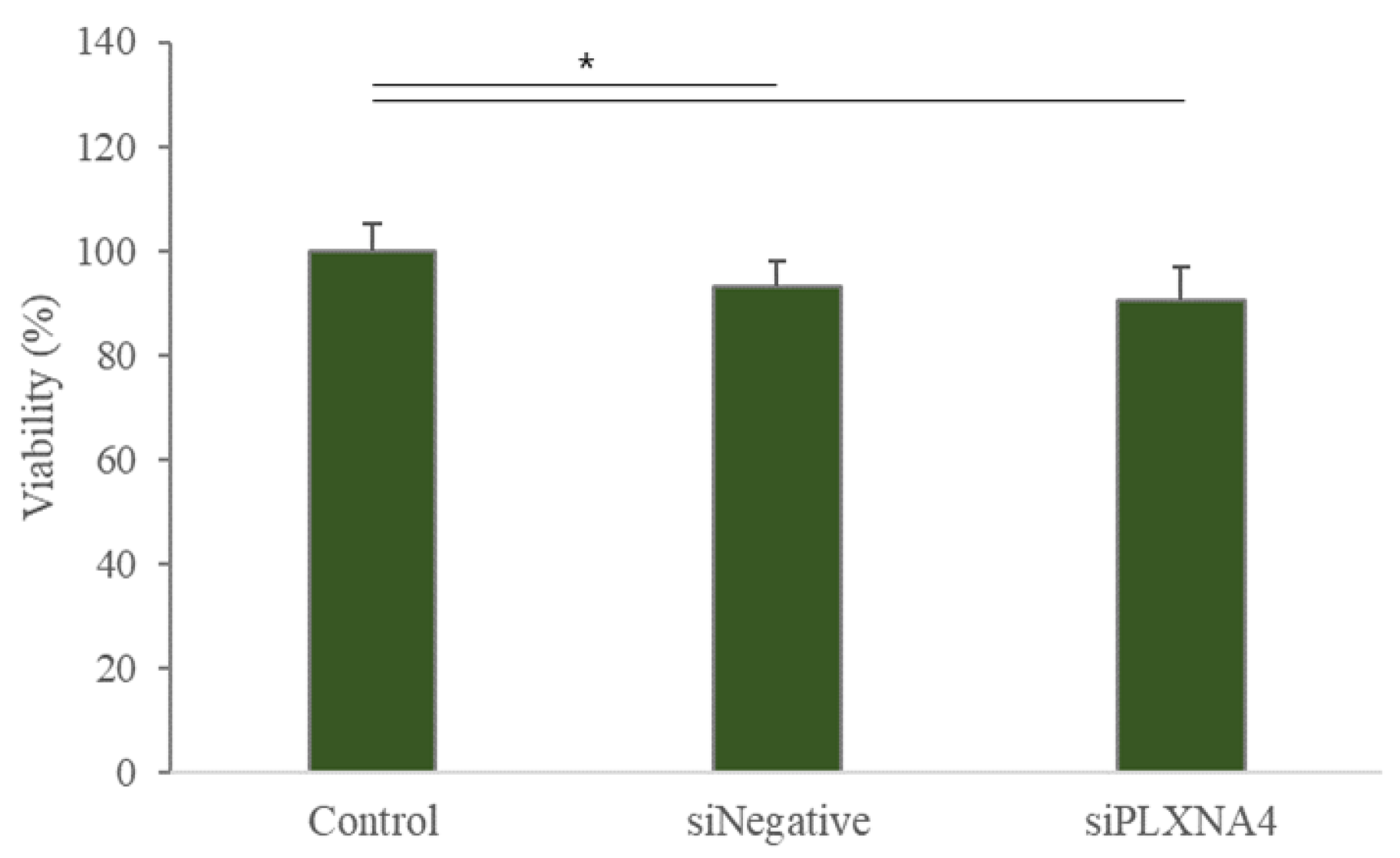
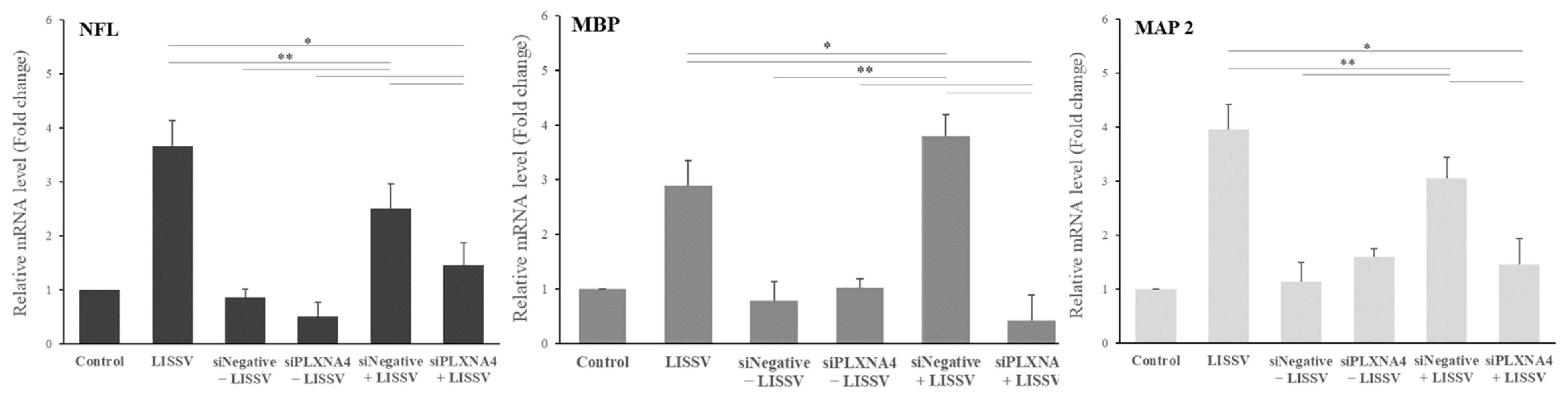
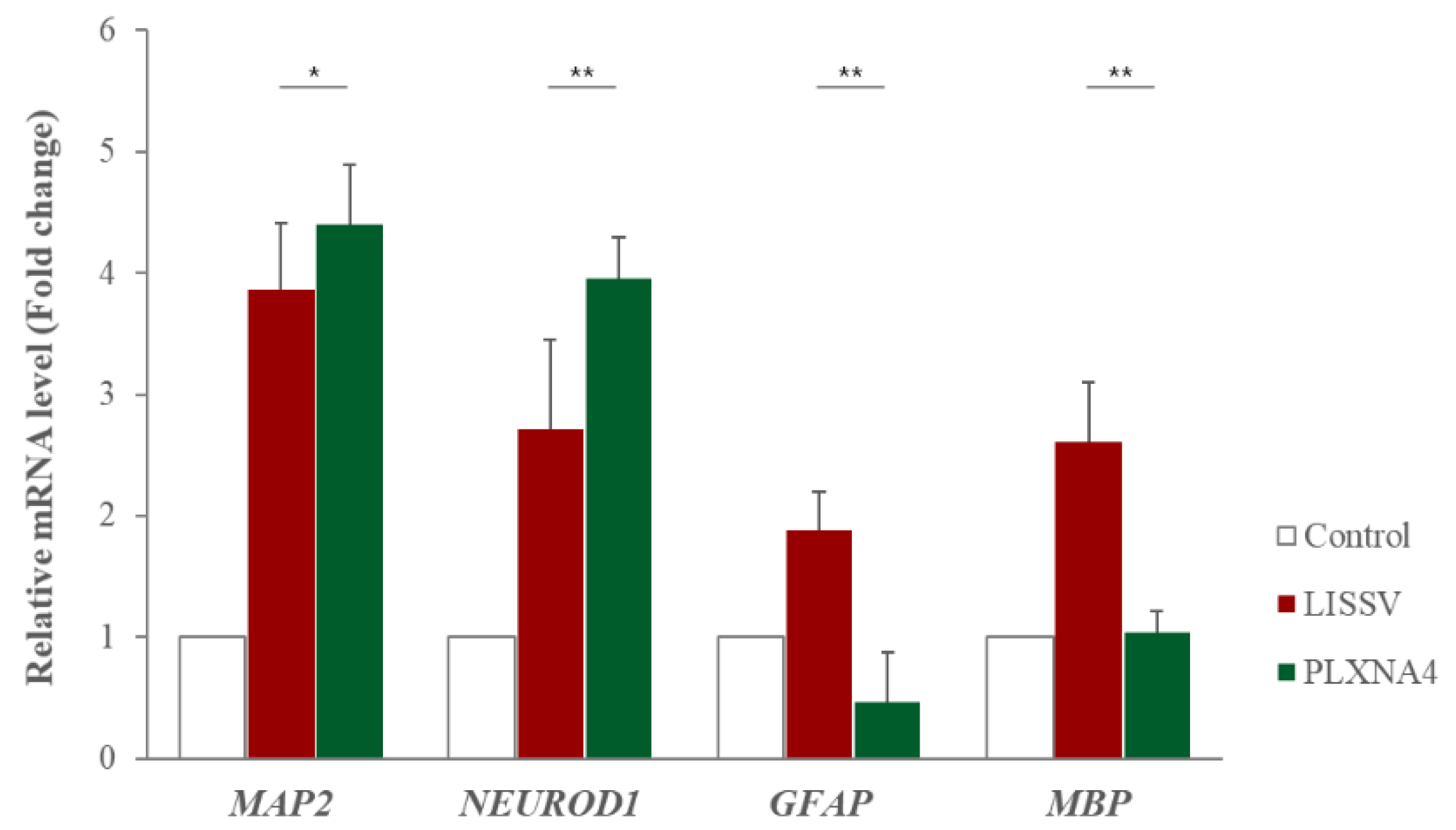
2.3. The Neural Differentiation of hUC-MSCs by LISSV Was Not Induced upon PLXNA4 Gene Silencing
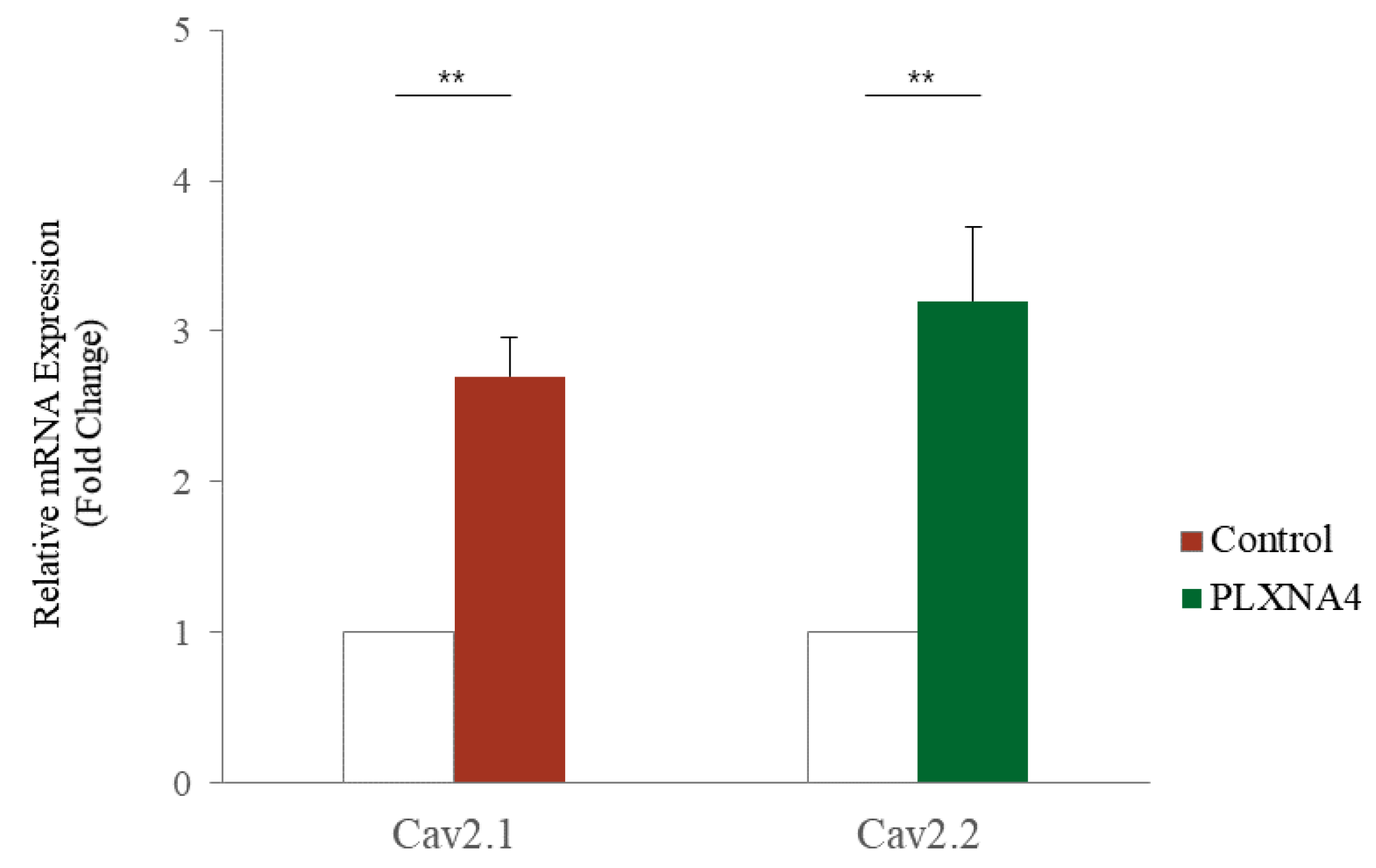
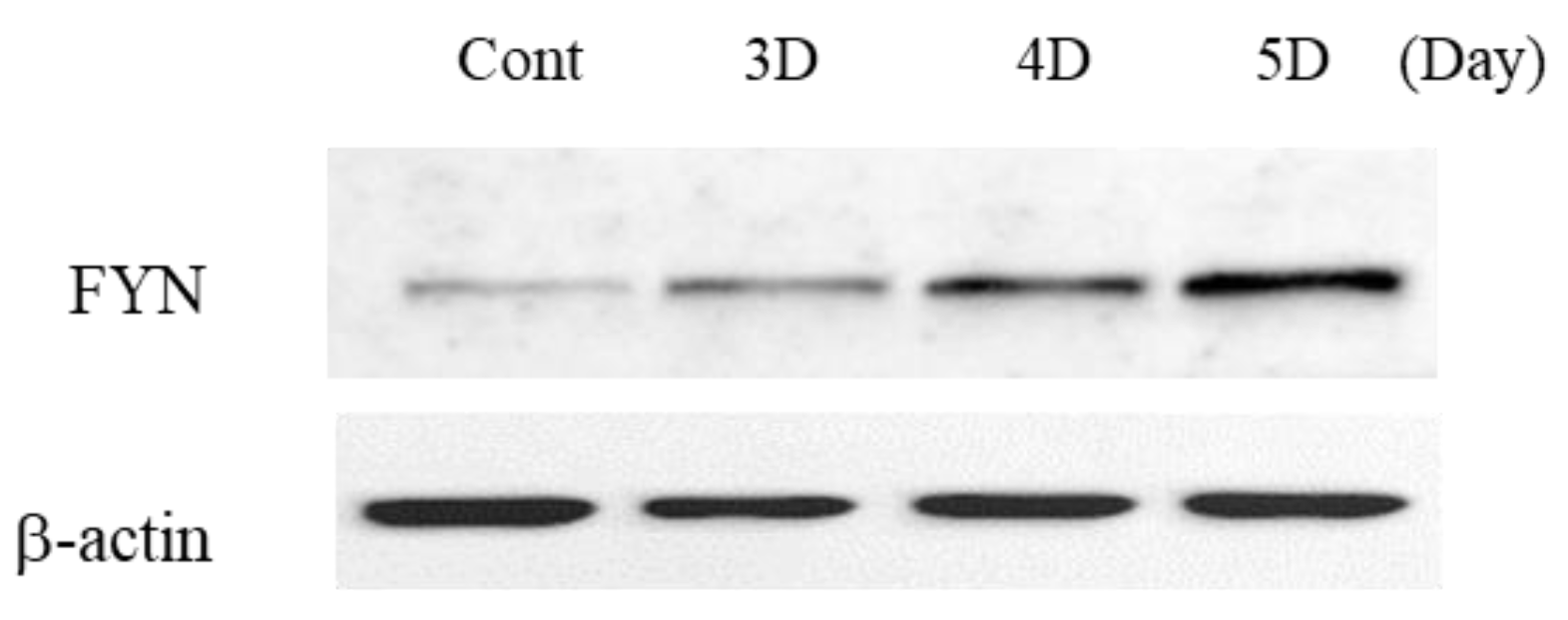
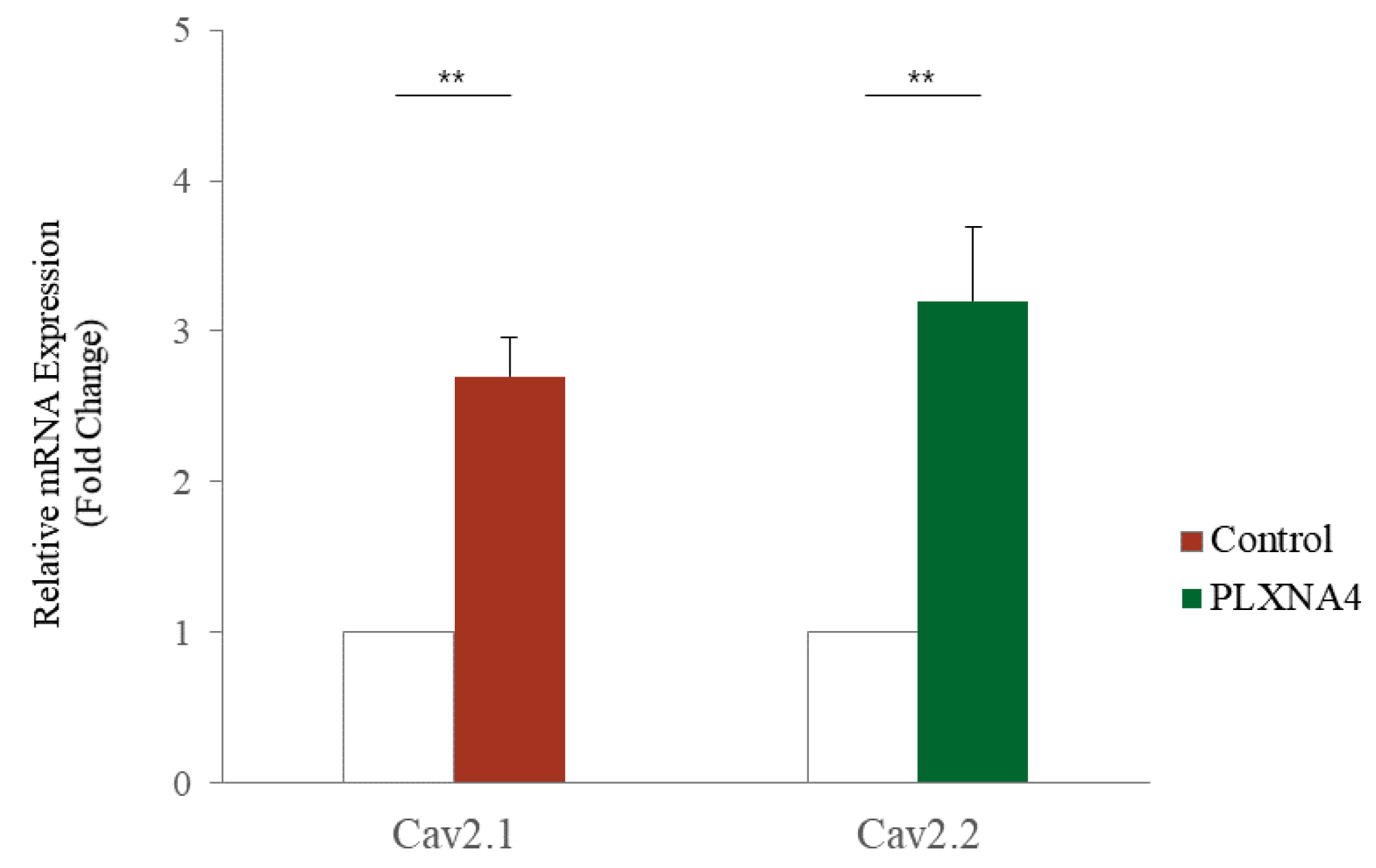
2.4. Changes with LISSV versus PLXNA4 in Sema-Dependent Signaling
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. LISSV Exposure
4.3. Cell Growth Assay
4.4. RNA-Sequencing Assay
4.5. Treatment with Recombinant PLXN4 Protein
4.6. Small Interfering RNA (siRNA) Transfection
4.7. Polymerase Chain Reaction (PCR) and Real-Time qPCR
4.8. Western Blot Analysis
4.9. Immunofluorescence Imaging
4.10. Fluorescence-Activated Cell Sorting (FACS) Analysis
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, Y.; Jia, X.; Bai, K.; Gong, X.; Fan, Y. Effect of Fluid Shear Stress on Cardiomyogenic Differentiation of Rat Bone Marrow Mesenchymal Stem Cells. Arch. Med. Res. 2010, 41, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Maul, T.M.; Chew, D.W.; Nieponice, A.; Vorp, D.A. Mechanical stimuli differentially control stem cell behavior: Morphology, proliferation, and differentiation. Biomech. Model. Mechanobiol. 2011, 10, 939–953. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Heo, S.J.; Kim, S.H.; Shin, J.W.; Park, S.H.; Shin, J.W. Shear stress magnitude is critical in regulating the differentiation of mesenchymal stem cells even with endothelial growth medium. Biotechnol. Lett. 2011, 33, 2351–2359. [Google Scholar] [CrossRef] [PubMed]
- Dan, P.; Velot, É.; Decot, V.; Menu, P. The role of mechanical stimuli in the vascular differentiation of mesenchymal stem cells. J. Cell Sci. 2015, 128, 2415–2422. [Google Scholar] [CrossRef] [Green Version]
- Fahy, N.; Alini, M.; Stoddart, M.J. Mechanical stimulation of mesenchymal stem cells: Implications for cartilage tissue engineering. J. Orthop. Res. 2018, 36, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Lee, W.Y.W.; Feng, Q.; Xu, L.; Wang, B.; Man, G.C.W.; Chen, Y.; Jiang, X.; Bian, L.; Cui, L.; et al. Synergistic effects on mesenchymal stem cell-based cartilage regeneration by chondrogenic preconditioning and mechanical stimulation. Stem. Cell. Res. Ther. 2017, 8, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, C.D.; Zhang, N.; Tong, W.X.; Zhang, Y.F.; Shan, S.Z.; Zhang, X.L.; Li, Q.F. Mechanical stimulation orchestrates the osteogenic differentiation of human bone marrow stromal cells by regulating HDAC1. Cell Death Dis. 2016, 7, e2221. [Google Scholar] [CrossRef] [Green Version]
- Pagnotti, G.M.; Styner, M.; Uzer, G.; Patel, V.S.; Wright, L.E.; Ness, K.K.; Guise, T.A.; Rubin, J.; Rubin, C.T. Combating osteoporosis and obesity with exercise: Leveraging cell mechanosensitivity. Nat. Rev. Endocrinol. 2019, 15, 339–355. [Google Scholar] [CrossRef]
- Rubin, C.; Turner, A.S.; Bain, S.; Mallinckrodt, C.; McLeod, K. Anabolism. Low mechanical signals strengthen long bones. Nature 2001, 412, 603–604. [Google Scholar] [CrossRef]
- Rubin, C.; Turner, A.S.; Müller, R.; Mittra, E.; McLeod, K.; Lin, W.; Qin, Y.X. Quantity and quality of trabecular bone in the femur are enhanced by a strongly anabolic, noninvasive mechanical intervention. J. Bone Min. Res. 2002, 17, 349–357. [Google Scholar] [CrossRef]
- Pagnotti, G.M.; Kurihara, N.; Roodman, G.D. Low Intensity Vibration Treatment Reduces Myeloma Cell-Induced Osteoclast Formation By Inhibiting IL-6 and GFI-1 Production By MM Cells. Blood 2017, 130, 1775. [Google Scholar]
- Pongkitwitoona, S.; Weinheimer-Hausb, E.M.; Koh, T.J.; Judex, S. Low-intensity vibrations accelerate proliferation and alter macrophage phenotype in vitro. J. Biomech. 2016, 49, 793–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagnotti, G.M.; Chan, M.E.; Adler, B.J.; Shroyer, K.R.; Rubin, J.; Bain, S.D.; Rubin, C.T. Low Intensity Vibration Mitigates Tumor Progression and Protect Bone Quantity and Quality in a Murine Model of Myeloma. Bone 2016, 90, 69–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suto, F.; Murakami, Y.; Nakamura, F.; Goshima, Y.; Fujisawa, H. Identification and characterization of a novel mouse plexin, plexin-A4. Mech. Dev. 2003, 120, 385–396. [Google Scholar] [CrossRef]
- Suto, F.; Ito, K.; Uemura, M.; Shimizu, M.; Shinkawa, Y.; Sanbo, M.; Shinoda, T.; Tsuboi, M.; Takashima, S.; Yagi, T.; et al. Plexin-A4 Mediates Axon-Repulsive Activities of Both Secreted and Transmembrane Semaphorins and Plays Roles in Nerve Fiber Guidance. J. Neurosci. 2005, 25, 3628–3637. [Google Scholar] [CrossRef]
- Liu, R.; Tian, B.; Gearing, M.; Hunter, S.; Ye, K.; Mao, Z. Cdk5-mediated regulation of the PIKE-A-Akt pathway and glioblastoma cell invasion. Proc. Natl. Acad. Sci. USA 2008, 105, 7570–7575. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Janssen, B.J.; Malinauskas, T.; Vangoor, V.R.; Coles, C.H.; Kaufmann, R.; Ni, T.; Gilbert, R.J.; Padilla-Parra, S.; Pasterkamp, R.J.; et al. Structural Basis for Plexin Activation and Regulation. Neuron 2016, 91, 548–560. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.Y.; Rong, R.; Kroll, T.G.; Meir, E.G.V.; Snyder, S.H.; Ye, K. PIKE (Phosphatidylinositol 3-Kinase Enhancer)-A GTPase Stimulates Akt Activity and Mediates Cellular Invasion. J. Biol. Chem. 2004, 279, 16441–16451. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.Y.; Hu, Y.; Kroll, T.G.; Allard, P.; Ye, K. PIKE-A is amplified in human cancers and prevents apoptosis by up-regulating Akt. Proc. Natl. Acad. Sci. USA 2004, 101, 6993–6998. [Google Scholar] [CrossRef] [Green Version]
- Han, Q.; Sun, Y.A.; Zong, Y.; Chen, C.; Wang, H.F.; Tan, L. Alzheimer’s Disease Neuroimaging Initiative Common Variants in PLXNA4 and Correlation to CSF-related Phenotypes in Alzheimer’s Disease. Front. Neurosci. 2018, 12, 946. [Google Scholar] [CrossRef]
- Cho, H.; Seo, Y.K.; Jeon, S.; Yoon, H.H.; Choi, Y.K.; Park, J.K. Neural differentiation of umbilical cord mesenchymal stem cells by sub-sonic vibration. Life Sci. 2012, 9, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.K.; Cho, H.; Seo, Y.K.; Yoon, H.H.; Park, J.K. Stimulation of sub-sonic vibration promotes the differentiation of adipose tissue-derived mesenchymal stem cells into neural cells. Life Sci. 2012, 91, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.K.; Lee, D.H.; Seo, Y.K.; Jung, H.; Park, J.K.; Cho, H. Stimulation of neural differentiation in human bone marrow mesenchymal stem cells by extremely low-frequency electromagnetic fields incorporated with MNPs. Appl. Biochem. Biotechnol. 2014, 174, 1233–1245. [Google Scholar] [CrossRef] [PubMed]
- Urnukhsaikhan, E.; Cho, T.; Mishig-Ochir, H.; Seo, Y.K.; Park, J.K. Pulsed lectromagnetic fields promote survival and neuronal differentiation of human BM-MSCs. Life Sci. 2016, 15, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Choi, Y.K.; Lee, D.H.; Park, H.J.; Seo, Y.K.; Jung, H.; Kim, S.C.; Kim, S.M.; Park, J.K. Effects of magnetic nanoparticle-incorporated human bone marrow—Derived mesenchymal stem cells exposed to pulsed electromagnetic fields on injured rat spinal cord. Biotechnol. Appl. Biochem. 2013, 60, 596–602. [Google Scholar] [CrossRef]
- Choi, Y.K.; Urnukhsaikhan, E.; Yoon, H.H.; Seo, Y.K.; Cho, H.; Jeong, J.S.; Kim, S.C.; Park, J.K. Combined effect of pulsed electromagnetic field and sound wave on In vitro and In vivo neural differentiation of human mesenchymal stem cells. Biotechnol. Prog. 2017, 33, 201–211. [Google Scholar] [CrossRef]
- Akiyama, H.; Fkuda, T.; Tojima, T.; Nikolaev, V.O.; Kamiguchi, H. Cyclic Nucleotide Control of Microtubule Dynamics for Axon Guidance. J. Neurosci. 2016, 36, 5636–5649. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, D.B.; Randall, A.; Tsien, R.W. Roles of N-type and Q-type Ca2+ channels in supporting hippocampal synaptic transmission. Science 1994, 264, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Pham, H.; Tonai, R.; Wu, M.; Birtolo, C.; Chen, M. CD73, CD90, CD105 and Cadherin-11 RT-PCR Screening for Mesenchymal Stem Cells from Cryopreserved Human Cord Tissue. Int. J. Stem. Cells 2018, 11, 26–38. [Google Scholar] [CrossRef] [Green Version]
- Law, J.W.S.; Lee, A.Y.W. The Role of Semaphorins and Their Receptors in Gliomas. J. Signal Transd. 2012, 2012, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Winberg, M.L.; Noordermeer, J.N.; Tamagnone, L.; Comoglio, P.M.; Spriggs, M.K.; Tessier-Lavigne, M.; Goodman, C.S. Plexin A Is a Neuronal Semaphorin Receptor that Controls Axon Guidance. Cell 1998, 95, 903–916. [Google Scholar] [CrossRef] [Green Version]
- Falk, J.; Castellani, V. Cellular Migration and Formation of Neuronal Connections, 1st ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 87–108. [Google Scholar]
- Bashaw, G.J.; Klein, R. Signaling from axon guidance receptors. Cold Spring Harb. Perspect. Biol. 2010, 2, a001941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yogev, S.; Shen, K. Cellular and Molecular Mechanisms of Synaptic Specificity. Annu. Rev. Cell Dev. Biol. 2014, 30, 417–437. [Google Scholar] [CrossRef] [PubMed]
- Pasterkamp, R.J. Getting neural circuits into shape with semaphorins. Nat. Rev. Neurosci. 2012, 13, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Benarroch, E.E. Neuronal voltage-gated calcium channels: Brief overview of their function and clinical implications in neurology. Neurology 2010, 74, 1310–1315. [Google Scholar] [CrossRef]
- Malgieri, A.; Kantzari, E.; Patrizi, M.P.; Gambardella, S. Bone marrow and umbilical cord blood human mesenchymal stem cells: State of the art. Int. J. Clin. Exp. Med. 2010, 3, 248–269. [Google Scholar]
- Baksh, D.; Yao, R.; Tuan, R.S. Comparison of Proliferative and Multilineage Differentiation Potential of Human Mesenchymal Stem Cells Derived from Umbilical Cord and Bone Marrow. Stem Cells 2007, 25, 1384–1392. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Maleki, M.; Ghanbarvand, F.; Behvarz, M.R.; Ejtemaei, M.; Ghadirkhomi, E. Comparison of Mesenchymal Stem Cell Markers in Multiple Human Adult Stem Cells. Int. J. Stem Cells. 2014, 7, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Limoni, G.; Niquille, M. Semaphorins and Plexins in central nervous system patterning: The key to it all? Curr. Opin. Neurobiol. 2021, 66, 224–232. [Google Scholar] [CrossRef]
- Kigel, B.; Varshavsky, A.; Kessler, O.; Neufeld, G. Successful inhibition of tumor development by specific class-3 semaphorins is associated with expression of appropriate semaphorin receptors by tumor cells. PLoS ONE 2008, 3, e3287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maione, F.; Molla, F.; Meda, C. Semaphorin 3A is an endogenous angiogenesis inhibitor that blocks tumor growth and normalizes tumor vasculature in transgenic mouse models. J. Clin. Investig. 2009, 119, 3356–3372. [Google Scholar] [CrossRef] [PubMed]
- Kigel, B.; Rabinowicz, N.; Varshavsky, A.; Kessler, O.; Neufeld, G. Plexin-A4 promotes tumor progression and tumor angiogenesis by enhancement of VEGF and bFGF signaling. Blood 2011, 118, 4285–4296. [Google Scholar] [CrossRef] [PubMed]
- Haklai-Topper, L.; Mlechkovich, G.; Savariego, D.; Gokhman, I.; Yaron, A. Cis interaction between Semaphorin6A and Plexin-A4 modulates the repulsive response to Sema6A. EMBO J. 2010, 29, 2635–2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suto, F.; Tsuboi, M.; Kamiya, H.; Mizuno, H.; Kiyama, Y.; Komai, S.; Shimizu, M.; Sanbo, M.; Yagi, T.; Hiromi, Y.; et al. Interactions between plexin-A2, plexin-A4, and semaphorin 6A control lamina-restricted projection of hippocampal mossy fibers. Neuron 2007, 53, 535–547. [Google Scholar] [CrossRef]
- Nasarre, P.; Gemmill, R.M.; Drabkin, H.A. The emerging role of class-3 semaphorins and their neuropilin receptors in oncology. Onco. Targets Ther. 2014, 7, 1663–1687. [Google Scholar]
- Uchida, Y.; Ohshima, T.; Sasaki, Y.; Suzuki, H.; Yanai, S.; Yamashita, N.; Nakamura, F.; Takei, K.; Ihara, Y.; Mikoshiba, K.; et al. Semaphorin-3A signaling is mediated via sequential Cdk5 and GSK3β phosphorylation of CRMP2: Implication of common phosphorylating mechanism underlying axon guidance and Alzheimer’s disease. Genes Cells 2005, 10, 165–179. [Google Scholar] [CrossRef]
- Sasaki, Y.; Cheng, C.; Uchida, Y.; Nakajima, O.; Ohshima, T.; Yagi, T.; Taniguchi, M.; Nakayama, T.; Kishida, R.; Kudo, Y.; et al. Fyn and Cdk5 mediate Semaphorin-3A signaling, which is involved in regulation of dendrite orientation in cerebral cortex. Neuron 2002, 35, 907–920. [Google Scholar] [CrossRef] [Green Version]
- Holahan, M.R. A Shift from a Pivotal to Supporting Role for the Growth-Associated Protein (GAP-43) in the Coordination of Axonal Structural and Functional Plasticity. Front. Cell. Neurosci. 2017, 11, 266. [Google Scholar] [CrossRef] [Green Version]
- Pasterkamp, R.J.; Giger, R.J. Semaphorin Function in Neural Plasticity and Disease. Curr. Opin. Neurobiol. 2009, 19, 263–274. [Google Scholar] [CrossRef] [Green Version]
- Morita, A.; Yamashita, N.; Sasaki, Y.; Uchida, Y.; Nakajima, O.; Nakamura, F.; Yagi, T.; Taniguchi, M.; Usui, H.; Katoh-Semba, R. Regulation of dendritic branching and spine maturation by semaphorin3AFyn signaling. J. Neurosci. 2006, 26, 2971–2980. [Google Scholar] [CrossRef] [PubMed]
- Kruminis-Kaszkiel, E.; Osowski, A.; Bejer-Ole´nska, E.; Dzieko´nski, M.; Wojtkiewicz, J. Differentiation of Human Mesenchymal Stem Cells from Wharton’s Jelly Towards Neural Stem Cells Using a Feasible and Repeatable Protocol. Cells 2020, 9, 739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, S.; Hamblin, M.R.; Abrahamse, H. Differentiation of Mesenchymal Stem Cells to Neuroglia: In the Context of Cell Signalling. Stem Cell Rev. Rep. 2019, 15, 814–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alizadeha, R.; Baghera, Z.; Kamravaa, S.K.; Falaha, M.; Hamidabadib, H.G.; Boroujenic, M.E.; Khodavedie, F.S.; Zare-Sadeghif, A.; Olyac, A.; Komeilig, A. Differentiation of human mesenchymal stem cells (MSC) to dopaminergic neurons: A comparison between Wharton’s Jelly and olfactory mucosa as sources of MSCs. J. Chem. Neuroanat. 2019, 96, 126–133. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Upstream Primer Sequence | Downstream Primer Sequence |
| PLXNA4 | 5′-ATC TCC GTC TCT CAG TAC AA-3′ | 5′-GTG ATA GGC TTG ATC ACC TC-3′ |
| FMN1 | 5′-CCA TCA CCG TTT TCT TCT TC-3′ | 5′-AGT TAC AGT GCC CTT GTA TG-3′ |
| AREG | 5′-TTC TAG TAG TGA ACC GTC CT-3′ | 5′-AGA CAT AAA GGC AGC TAT GG-3′ |
| STMN2 | 5′-CAG AGG GAA GGA GAG AAG CAA T-3′ | 5′-TCA TTA GGC AAT GGT GGG TT-3′ |
| SERPINI1 | 5′-AAA ACC TCT CGG GTG AAA G-3′ | 5′-GCT GTC ATA TCC CAT TGA GT-3′ |
| MAP2 | 5′-CTC AAC AGT TCT ATC TCT TCT TCA-3′ | 5′-TCT TCT TGT TTA AAA TCC TAA CCT-3′ |
| NFL | 5′-CAA GAA CAT GCA GAA CGC TG-3′ | 5′-GCC TTC CAA GAG TTT CCT GT-3′ |
| NEUROD1 | 5′-ACA GTC ACC AGT GTG GTG GA-3′ | 5′-CGT AGC CTC TGG AGA ACC TG-3′ |
| GFAP | 5′-TCATCGCTCAGGAGGTCCTT-3′ | 5′-CTGTTGCCAGAGATGGAGGTT-3′ |
| MBP | 5′-CGG CAA CTA CGT GCT CTT CA-3′ | 5′-GTG ACT TCA TCT CGT GGG CC-3′ |
| SEMA3A | 5′-TAA GGA GAA AGG AGG AGA GGT G-3′ | 5′-GTG CTG GTT TGA ACT AGA GG-3′ |
| SEMA6A | 5′-TTA CAA CAC AGT GTA TGG GC-3′ | 5′-CTT TGA GGT AAC TTT CCC GA-3′ |
| SYN1 | 5′-GCT CAA CAA ATC CCA GTC TC-3′ | 5′-GAG GAG TCA GGT TTC TCA AG-3′ |
| Synaptophysin | 5′-CCT ATA CCC TAG GTC TCC AC-3′ | 5′-CCT GTC CTC CTT TTA GAT CC-3′ |
| GAP43 | 5′-CCA TGC TGT GCT GTA TGA GAA G-3′ | 5′-TAA GGA CTA GGT CGA ACT GC-3′ |
| Cav2.1 | 5′-CCG TGT GAT AAG AAC TCT GG-3′ | 5′-GAC ATG TGT CTC AGC ATC-3′ |
| Cav2.2 | 5′-CCA TCT TCT ACG TGG TCT AC-3′ | 5′-CAT CAG CTC GTA CTC ATA GG-3′ |
| GAPDH | 5′-ACC ACA GTC CAT GCC ATC AC-3′ | 5′-TCC ACC ACC CTG TTG CTG TA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, H.; Park, H.-J.; Seo, Y.-K. Induction of PLXNA4 Gene during Neural Differentiation in Human Umbilical-Cord-Derived Mesenchymal Stem Cells by Low-Intensity Sub-Sonic Vibration. Int. J. Mol. Sci. 2022, 23, 1522. https://doi.org/10.3390/ijms23031522
Cho H, Park H-J, Seo Y-K. Induction of PLXNA4 Gene during Neural Differentiation in Human Umbilical-Cord-Derived Mesenchymal Stem Cells by Low-Intensity Sub-Sonic Vibration. International Journal of Molecular Sciences. 2022; 23(3):1522. https://doi.org/10.3390/ijms23031522
Chicago/Turabian StyleCho, Hyunjin, Hee-Jung Park, and Young-Kwon Seo. 2022. "Induction of PLXNA4 Gene during Neural Differentiation in Human Umbilical-Cord-Derived Mesenchymal Stem Cells by Low-Intensity Sub-Sonic Vibration" International Journal of Molecular Sciences 23, no. 3: 1522. https://doi.org/10.3390/ijms23031522
APA StyleCho, H., Park, H.-J., & Seo, Y.-K. (2022). Induction of PLXNA4 Gene during Neural Differentiation in Human Umbilical-Cord-Derived Mesenchymal Stem Cells by Low-Intensity Sub-Sonic Vibration. International Journal of Molecular Sciences, 23(3), 1522. https://doi.org/10.3390/ijms23031522