
The Role of B Cells in PE Pathophysiology: A Potential Target for Perinatal Cell-Based Therapy?


 ,
, 
Abstract
1. Introduction
2. Preeclampsia: Pathogenic Mechanisms
2.1. Definition and Pathogenesis
2.2. Hypoxia and Oxidative Stress
2.3. Angiogenic Factors
2.4. Heme Oxygenase, Hydrogen Sulfide, and Nitric Oxide Pathways
3. Role of Immune Cells in Preeclampsia
3.1. Immune Cells in Physiological Pregnancy
3.2. Immune Cells in PE
4. Focus on B Cells
4.1. B Cell Main Features
4.2. B Cells in Physiological Pregnancy
4.3. B Cells in PE
5. B Cells as Target of Perinatal Cells
5.1. In Vitro
5.2. In Vivo

{kind=link}
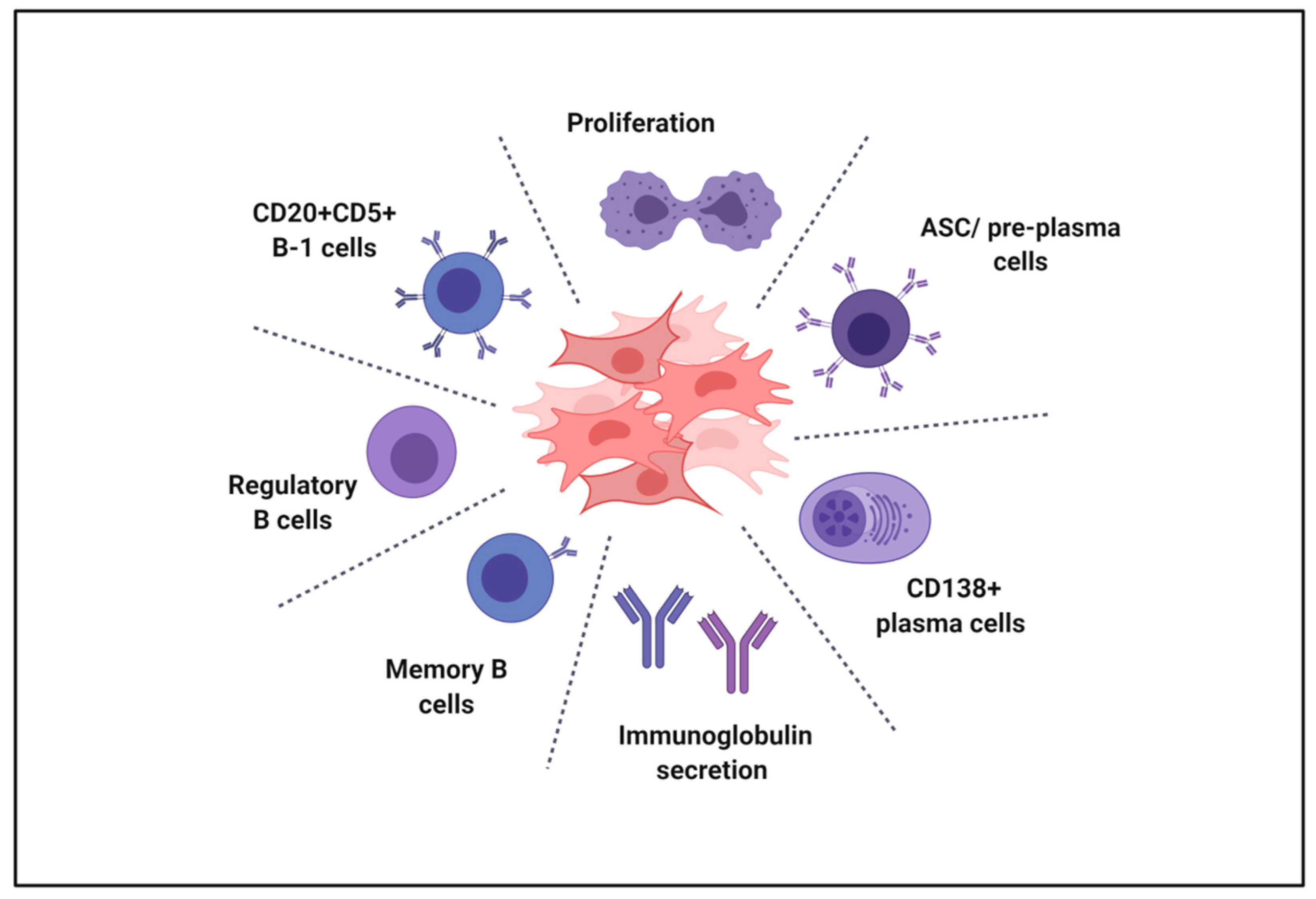{kind=link}
| Perinatal Cells | Experimental Model | Effects on B Cells | Mechanisms Involved | Ref |
|---|---|---|---|---|
| Human umbilical cord mesenchymal stromal cells | In vitro co-cultured with purified mouse splenic B cells stimulated by CpG+ IgM + CD40L + IL-4 in a contact-independent setting | - Abrogation of B220+ B cell proliferation - Blockage of the terminal differentiation into CD138+ plasma cells - Reduction in immunoglobulin secretion | - Increase of PAX-5 and suppression of Blimp-1 mRNA expression - Inhibition of Akt and p38 MAPK phosphorylation | [88] |
| In vitro co-cultured with the Burkitt’s lymphoma cell line, even in the absence of cell–cell contact | - Abrogation of lymphoma cells proliferation | - Arrest of lymphoma cells division in the S phase | [89] | |
| In vitro co-cultured with autoreactive B lymphocytes from peripheral blood of immune thrombocytopenic patients | - Abrogation of CD19+ B cell proliferation | - Not investigated | [90] | |
| In vitro co-cultured with human PBMC stimulated by PHA | - Not inhibition of CD19+ B cell proliferation | - Not investigated | [93] | |
| In vitro co-cultured with human purified B cells stimulated by CpG+ IgA + IgG + IgM + CD40L + IL-2, even in the absence of cell–cell contact | - Stimulation of CD19+ B cell proliferation - Increase of CD138+ cell terminal differentiation, and immunoglobulin production | - Prostaglandin E2 | [94] | |
| Clinical trial of cGVHD prophylaxis in patients after HLA-haplo HSCT | - Increase of CD27+ memory B cells | - Not investigated | [121] | |
| Human amniotic membrane mesenchymal stromal cells | In vitro co-cultured of human PBMC and purified B cells stimulated by CpG with conditioned medium from hAMSC | - Suppression of CD19+ B cell proliferation - Blockage of ASC/pre-plasma cells CD27+CD38+ formation - Blockage of the terminal differentiation into CD138+ plasma cells - Increase CD19+CD27+CD38− memory B cells | - Inhibition of IRF-4, PRDM1, XBP1 mRNA expression - Suppression of TLR9-MyD88-IRAK1/4 and TLR9-PI3K-AKT signaling pathways - Reduction of CpG sensors (CD205, TLR9, and CD14) - Inhibition of IRAK-4, and phosphorylated MAPK (JNK, p38 MAPK, ERK), NF-kB, and AKT - Prostanoids may be involved | [85] |
| Experimental mouse model of idiopathic pulmonary fibrosis | - Reduction of B220 B cells in alveolar spaces - Reduction of CD138+ antibody-secreting cells in lung tissues | - Down-modulation of CXCL13 and BAFF lung expression | [86] | |
| Human amniotic fluid stromal cells | In vitro co-culture of human purified B cells stimulated by CpG+ Immunoglobulin + CD40L +IL-2 + IL-4 | - Inhibition of CD19+ B cell proliferation and activation (CD80/CD86 expression) - Reduction of the proportion of CD19+CD20+CD27+ memory B cells - Blockage of CD19+CD20−CD27+ plasma cells - Reduction in immunoglobulin secretion - Inhibition of IL-10+ cells Breg formation - Downregulation of the proportion of CD5+ B1 cells - Inhibition of apoptosis of activated B cell | - Decrease in the percentage of B cells in S phase cycle - Downregulation of the expression of CD80/CD86 costimulatory molecules on activated B lymphocytes - Decrease in the expression of B7H4 and PD-L1 in activated B cells may be involved | [91] |
| Human amniotic epithelial cells | In vitro co-culture with human purified B cells from peripheral blood stimulated by CpG+ CD40L + IL-4 | - Increase in B cell proliferation - Diminished spontaneous apoptosis - Expansion of CD19+CD24hiCD38hi Bregs | - Adenosine may be involved | [92] |
| Experimental mouse model of autoimmune thyroiditis | - Upregulation of IL-10+ Bregs | - Not investigated | [118] | |
| Experimental mouse model of systemic lupus erythematosus | - Reduction of autoantibody production | - Not investigated | [118] |
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PE | Preeclampsia |
| HIF | Hypoxia-Inducible Factor |
| ROS | Reactive Oxygen Species |
| sFlt-1 | soluble Fms-like tyrosine kinase-1 |
| VEGF | Vascular-Endothelial Growth Factor |
| PlGF | Placental Growth Factor |
| sEng | Soluble Endoglin |
| TGF | Transforming Growth Factor |
| HO | Heme Oxygenase |
| CO | Carbon Monoxide |
| H2S | Hydrogen Sulfide |
| NO | Nitric Oxide |
| NK | Natural Killer |
| Th | T helper |
| Tregs | Regulatory T cells |
| TNF | Tumor Necrosis Factor |
| IFN | Interferon |
| MPO | Myeloperoxidase |
| hUC-MSC | Human Umbilical Cord Mesenchymal Stromal Cells |
| PBMC | Peripheral Blood Mononuclear Cells |
| hAMSC | Human Amniotic Mesenchymal Stromal Sells |
| CM | Conditioned Medium |
| B7H4 | B7 homolog 4 |
| PD-L1 | Programmed Death-Ligand 1 |
| hAEC | Human Amniotic Epithelial Cells |
| ASC | Antibody-Secreting Cells |
| PAX-5 | Paired Box 5 |
| BCL-6 | B-cell CLL/Lymphoma 6 |
| IRF4 | Interferon Regulatory Factor 4 |
| PRDM1 | PR/SET Domain 1 |
| BLIMP-1 | B Lymphocyte Induced Maturation Protein-1 |
| XBP-1 | X-box Binding Protein 1 |
| CpG ODN | CpG Oligodeoxynucleotides |
| TLR9 | Toll-Like Receptor 9 |
| MyD88 | Myeloid Differentiation Primary Response 88 |
| IRAK1 | Interleukin-1 Receptor-Associated Kinase 1 |
| IRAK4 | Interleukin-1 Receptor-Associated Kinase 4 |
| PI3K | Phosphatidylinositol 3-Kinase |
| AKT | Protein Kinase B |
| MAPK | Mitogen-Activated Protein Kinases |
| JNK | c-Jun N-terminal Kinase |
| p38 MAPK | p38 Mitogen-Activated Protein Kinases |
| ERK | Extracellular Signal-Regulated Kinase |
| NF-kB | Nuclear Factor Kappa-Light-Chain-Enhancer Of Activated B Cells |
| BCR | B Cell Receptor |
| PGE2 | Prostaglandin E2 |
| IPF | Idiopathic Pulmonary Fibrosis |
| CXCL13 | Chemokine (C-X-C motif) Ligand 13 |
| BAFF | B-Cell Activating Factor |
| HT | Hashimoto’s Thyroiditis |
| SLE | Systemic Lupus Erythematosus |
| Bregs | Regulatory B cells |
| B10 | Regulatory B cells that produce IL-10 |
| cGVHD | Chronic Graft-Versus-Host Disease |
| HLA | Human Leukocyte Antigen |
| HLA-haplo HSCT | HLA-Haploidentical Hematopoietic Stem-Cell Transplantation |
References
- Ives, C.W.; Sinkey, R.; Rajapreyar, I.; Tita, A.T.; Oparil, S. Preeclampsia—Pathophysiology and Clinical Presentations. J. Am. Coll. Cardiol. 2020, 76, 1690–1702. [Google Scholar] [CrossRef]
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia: Pathophysiology, Challenges, and Perspectives. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef]
- Silini, A.R.; Di Pietro, R.; Lang-Olip, I.; Alviano, F.; Banerjee, A.; Basile, M.; Borutinskaite, V.; Eissner, G.; Gellhaus, A.; Giebel, B.; et al. Perinatal Derivatives: Where Do We Stand? A Roadmap of the Human Placenta and Consensus for Tissue and Cell Nomenclature. Front. Bioeng. Biotechnol. 2020, 8, 610544. [Google Scholar] [CrossRef]
- American College of Obstetricians and Gynecologists. ACOG Practice Bulletin No. 202 Summary: Gestational Hypertension and Preeclampsia. Obstet. Gynecol. 2019, 133, 211–214. [Google Scholar]
- Jim, B.; Karumanchi, S.A. Preeclampsia: Pathogenesis, Prevention, and Long-Term Complications. Semin. Nephrol. 2017, 37, 386–397. [Google Scholar] [CrossRef]
- Soleymanlou, N.; Jurisica, I.; Nevo, O.; Ietta, F.; Zhang, X.; Zamudio, S.; Post, M.; Caniggia, I. Molecular Evidence of Placental Hypoxia in Preeclampsia. J. Clin. Endocrinol. Metab. 2005, 90, 4299–4308. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Placental Oxidative Stress: From Miscarriage to Preeclampsia. J. Soc. Gynecol. Investig. 2004, 11, 342–352. [Google Scholar] [CrossRef]
- Vaughan, J.E.; Walsh, S.W. Oxidative stress reproduces placental abnormalities of preeclampsia. Hypertens. Pregnancy 2002, 21, 205–223. [Google Scholar] [CrossRef]
- Nagamatsu, T.; Fujii, T.; Kusumi, M.; Zou, L.; Yamashita, T.; Osuga, Y.; Momoeda, M.; Kozuma, S.; Taketani, Y. Cytotrophoblasts Up-Regulate Soluble Fms-Like Tyrosine Kinase-1 Expression under Reduced Oxygen: An Implication for the Placental Vascular Development and the Pathophysiology of Preeclampsia. Endocrinology 2004, 145, 4838–4845. [Google Scholar] [CrossRef]
- Maynard, S.E.; Min, J.-Y.; Merchan, J.; Lim, K.-H.; Li, J.; Mondal, S.; Libermann, T.A.; Morgan, J.P.; Sellke, F.W.; Stillman, I.E.; et al. Excess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsia. J. Clin. Investig. 2003, 111, 649–658. [Google Scholar] [CrossRef]
- Venkatesha, S.; Toporsian, M.; Lam, C.; Hanai, J.-I.; Mammoto, T.; Kim, Y.M.; Bdolah, Y.; Lim, K.-H.; Yuan, H.-T.; Libermann, T.A.; et al. Soluble endoglin contributes to the pathogenesis of preeclampsia. Nat. Med. 2006, 12, 642–649. [Google Scholar] [CrossRef]
- Rana, S.; Schnettler, W.T.; Powe, C.; Wenger, J.; Salahuddin, S.; Cerdeira, A.S.; Verlohren, S.; Perschel, F.H.; Arany, Z.; Lim, K.-H.; et al. Clinical characterization and outcomes of preeclampsia with normal angiogenic profile. Hypertens. Pregnancy 2013, 32, 189–201. [Google Scholar] [CrossRef]
- McCaig, D.; Lyall, F. Inhibitors of Heme Oxygenase Reduce Invasion of Human Primary Cytotrophoblast Cells In Vitro. Placenta 2009, 30, 536–538. [Google Scholar] [CrossRef] [PubMed]
- Lyall, F.; Barber, A.; Myatt, L.; Bulmer, J.N.; Robson, S.C. Hemeoxygenase expression in human placenta and placental bed implies a role in regulation of trophoblast invasion and placental function. FASEB J. 2000, 14, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Barber, A.; Robson, S.C.; Myatt, L.; Bulmer, J.N.; Lyall, F. Heme oxygenase expression in human placenta and placental bed: Reduced expression of placenta endothelial HO-2 in preeclampsia and fetal growth restriction. FASEB J. 2001, 15, 1158–1168. [Google Scholar] [CrossRef] [PubMed]
- Cudmore, M.; Ahmad, S.; Al-Ani, B.; Fujisawa, T.; Coxall, H.; Chudasama, K.; Devey, L.R.; Wigmore, S.J.; Abbas, A.; Hewett, P.W.; et al. Negative Regulation of Soluble Flt-1 and Soluble Endoglin Release by Heme Oxygenase-1. Circulation 2007, 115, 1789–1797. [Google Scholar] [CrossRef] [PubMed]
- Lowe, D.T. Nitric Oxide Dysfunction in the Pathophysiology of Preeclampsia. Nitric Oxide 2000, 4, 441–458. [Google Scholar] [CrossRef]
- Holwerda, K.M.; Burke, S.D.; Faas, M.M.; Zsengeller, Z.; Stillman, I.E.; Kang, P.M.; Van Goor, H.; McCurley, A.; Jaffe, I.Z.; Karumanchi, S.A.; et al. Hydrogen Sulfide Attenuates sFlt1-Induced Hypertension and Renal Damage by Upregulating Vascular Endothelial Growth Factor. J. Am. Soc. Nephrol. 2013, 25, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Aneman, I.; Pienaar, D.; Suvakov, S.; Simic, T.P.; Garovic, V.D.; McClements, L. Mechanisms of Key Innate Immune Cells in Early- and Late-Onset Preeclampsia. Front. Immunol. 2020, 11, 1864. [Google Scholar] [CrossRef]
- Laresgoiti-Servitje, E. A leading role for the immune system in the pathophysiology of preeclampsia. J. Leukoc. Biol. 2013, 94, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Bulmer, J.N.; Pace, D.; Ritson, A. Immunoregulatory cells in human decidua: Morphology, immunohistochemistry and function. Reprod. Nutr. Dev. 1988, 28, 1599–1614. [Google Scholar] [CrossRef]
- Ozenci, C.C.; Korgun, E.T.; Demir, R. Immunohistochemical detection of CD45+, CD56+, and CD14+ cells in human decidua during early pregnancy. Early Pregnancy 2001, 5, 164–175. [Google Scholar]
- Muzzio, D.O.; Soldati, R.; Ehrhardt, J.; Utpatel, K.; Evert, M.; Zenclussen, A.C.; Zygmunt, M.; Jensen, F. B Cell Development Undergoes Profound Modifications and Adaptations During Pregnancy in Mice1. Biol. Reprod. 2014, 91, 115. [Google Scholar] [CrossRef]
- Saito, S.; Nakashima, A.; Shima, T.; Ito, M. Th1/Th2/Th17 and regulatory T-cell paradigm in pregnancy. Am. J. Reprod. Immunol. 2010, 63, 601–610. [Google Scholar] [CrossRef]
- Mjösberg, J.; Berg, G.; Jenmalm, M.C.; Ernerudh, J. FOXP3+ Regulatory T Cells and T Helper 1, T Helper 2, and T Helper 17 Cells in Human Early Pregnancy Decidua1. Biol. Reprod. 2010, 82, 698–705. [Google Scholar] [CrossRef] [PubMed]
- La Rocca, C.; Carbone, F.; Longobardi, S.; Matarese, G. The immunology of pregnancy: Regulatory T cells control maternal immune tolerance toward the fetus. Immunol. Lett. 2014, 162, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W.; Sargent, I.L. Immunology of Pre-Eclampsia. Am. J. Reprod. Immunol. 2010, 63, 534–543. [Google Scholar] [CrossRef]
- Croy, B.A.; Ashkar, A.A.; Minhas, K.; Greenwood, J.D. Can Murine Uterine Natural Killer Cells Give Insights Into the Pathogenesis of Preeclampsia? J. Soc. Gynecol. Investig. 2000, 7, 12–20. [Google Scholar] [CrossRef]
- Wallace, A.E.; Host, A.J.; Whitley, G.S.; Cartwright, J.E. Decidual Natural Killer Cell Interactions with Trophoblasts Are Impaired in Pregnancies at Increased Risk of Preeclampsia. Am. J. Pathol. 2013, 183, 1853–1861. [Google Scholar] [CrossRef]
- Nagamatsu, T.; Schust, D.J. The Contribution of Macrophages to Normal and Pathological Pregnancies. Am. J. Reprod. Immunol. 2010, 63, 460–471. [Google Scholar] [CrossRef]
- Renaud, S.J.; Graham, C.H. The Role of Macrophages in Utero-placental Interactions during Normal and Pathological Pregnancy. Immunol. Investig. 2008, 37, 535–564. [Google Scholar] [CrossRef]
- Melgert, B.N.; Spaans, F.; Borghuis, T.; Klok, P.A.; Groen, B.; Bolt, A.; De Vos, P.; Van Pampus, M.G.; Wong, T.Y.; Van Goor, H.; et al. Pregnancy and Preeclampsia Affect Monocyte Subsets in Humans and Rats. PLoS ONE 2012, 7, e45229. [Google Scholar] [CrossRef]
- Al-Ofi, E.; Coffelt, S.B.; Anumba, D.O. Monocyte Subpopulations from Pre-Eclamptic Patients Are Abnormally Skewed and Exhibit Exaggerated Responses to Toll-Like Receptor Ligands. PLoS ONE 2012, 7, e42217. [Google Scholar] [CrossRef]
- Faas, M.M.; Spaans, F.; De Vos, P. Monocytes and Macrophages in Pregnancy and Pre-Eclampsia. Front. Immunol. 2014, 5, 298. [Google Scholar] [CrossRef]
- Dong, M.; Wang, Z.; He, J. Serum T helper 1- and 2-type cytokines in preeclampsia. Int. J. Gynecol. Obstet. 2005, 89, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Borzychowski, A.M.; Croy, B.A.; Chan, W.L.; Redman, C.W.G.; Sargent, I.L. Changes in systemic type 1 and type 2 immunity in normal pregnancy and pre-eclampsia may be mediated by natural killer cells. Eur. J. Immunol. 2005, 35, 3054–3063. [Google Scholar] [CrossRef]
- Dong, M.; He, J.; Wang, Z.; Xie, X.; Wang, H. Placental imbalance of Th1- and Th2-type cytokines in preeclampsia. Acta Obstet. Gynecol. Scand. 2005, 84, 788–793. [Google Scholar] [CrossRef] [PubMed]
- Darmochwal-Kolarz, D.; Rolinski, J.; Leszczynska-Gorzelak, B.; Oleszczuk, J. The Expressions of Intracellular Cytokines in the Lymphocytes of Preeclamptic Patients. Am. J. Reprod. Immunol. 2002, 48, 381–386. [Google Scholar] [CrossRef]
- Van Nieuwenhoven, A.L.V.; Moes, H.; Heineman, M.J.; Santema, J.; Faas, M.M. Cytokine Production by Monocytes, NK Cells, and Lymphocytes Is Different in Preeclamptic Patients as Compared with Normal Pregnant Women. Hypertens. Pregnancy 2008, 27, 207–224. [Google Scholar] [CrossRef]
- Saito, S. Th17 cells and regulatory T cells: New light on pathophysiology of preeclampsia. Immunol. Cell Biol. 2010, 88, 615–617. [Google Scholar] [CrossRef]
- Darmochwal-Kolarz, D.; Kludka-Sternik, M.; Tabarkiewicz, J.; Kolarz, B.; Rolinski, J.; Leszczynska-Gorzelak, B.; Oleszczuk, J. The predominance of Th17 lymphocytes and decreased number and function of Treg cells in preeclampsia. J. Reprod. Immunol. 2012, 93, 75–81. [Google Scholar] [CrossRef]
- Wu, L.; Li, J.; Xu, H.L.; Xu, B.; Tong, X.H.; Kwak-Kim, J.; Liu, Y.S. IL-7/IL-7R signaling pathway might play a role in recurrent pregnancy losses by increasing inflammatory Th17 cells and decreasing Treg cells. Am. J. Reprod. Immunol. 2016, 76, 454–464. [Google Scholar] [CrossRef]
- Toldi, G.; Vásárhelyi, Z.E.; Rigó, J.; Orbán, C.; Tamássy, Z.; Bajnok, A.; Shima, T.; Saito, S.; Molvarec, A. Prevalence of Regulatory T-Cell Subtypes in Preeclampsia. Am. J. Reprod. Immunol. 2015, 74, 110–115. [Google Scholar] [CrossRef]
- Prins, J.R.; Boelens, H.M.; Heimweg, J.; Van Der Heide, S.; Dubois, A.E.; Van Oosterhout, A.J.; Erwich, J.J.H. Preeclampsia is Associated with Lower Percentages of Regulatory T Cells in Maternal Blood. Hypertens. Pregnancy 2009, 28, 300–311. [Google Scholar] [CrossRef]
- Sasaki, Y.; Darmochwal-Kolarz, D.; Suzuki, D.; Sakai, M.; Ito, M.; Shima, T.; Shiozaki, A.; Rolinski, J.; Saito, S. Proportion of peripheral blood and decidual CD4+ CD25bright regulatory T cells in pre-eclampsia. Clin. Exp. Immunol. 2007, 149, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Tilburgs, T.; Roelen, D.L.; Van Der Mast, B.J.; Van Schip, J.J.; Kleijburg, C.; De Groot-Swings, G.M.; Kanhai, H.H.H.; Claas, F.H.J.; Scherjon, S.A. Differential Distribution of CD4+CD25bright and CD8+CD28− T-cells in Decidua and Maternal Blood During Human Pregnancy. Placenta 2006, 27, 47–53. [Google Scholar] [CrossRef]
- Cua, D.J.; Tato, C.M. Innate IL-17-producing cells: The sentinels of the immune system. Nat. Rev. Immunol. 2010, 10, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Moodley, M.; Moodley, J.; Naicker, T. The Role of Neutrophils and Their Extracellular Traps in the Synergy of Pre-eclampsia and HIV Infection. Curr. Hypertens. Rep. 2020, 22, 1–9. [Google Scholar] [CrossRef]
- Greer, I.A.; Haddad, N.G.; Dawes, J.; Johnstone, F.D.; Calder, A.A. Neutrophil activation in pregnancy-induced hypertension. BJOG Int. J. Obstet. Gynaecol. 1989, 96, 978–982. [Google Scholar] [CrossRef] [PubMed]
- Shukla, J.; Walsh, S.W. Neutrophil Release of Myeloperoxidase in Systemic Vasculature of Obese Women May Put Them at Risk for Preeclampsia. Reprod. Sci. 2015, 22, 300–307. [Google Scholar] [CrossRef]
- Alijotas-Reig, J.; Esteve-Valverde, E.; Ferrer-Oliveras, R.; Llurba, E.; Gris, J.M. Tumor Necrosis Factor-Alpha and Pregnancy: Focus on Biologics. An Updated and Comprehensive Review. Clin. Rev. Allergy Immunol. 2017, 53, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Cadden, K.A.; Walsh, S.W. Neutrophils, but Not Lymphocytes or Monocytes, Infiltrate Maternal Systemic Vasculature in Women with Preeclampsia. Hypertens. Pregnancy 2008, 27, 396–405. [Google Scholar] [CrossRef]
- Xia, Y.; Kellems, R.E. Is preeclampsia an autoimmune disease? Clin. Immunol. 2009, 133, 1–12. [Google Scholar] [CrossRef]
- Stall, A.M.; Fariñas, M.C.; Tarlinton, D.M.; Lalor, P.A.; Herzenberg, L.A.; Strober, S. Ly-1 B-cell clones similar to human chronic lymphocytic leukemias routinely develop in older normal mice and young autoimmune (New Zealand Black-related) animals. Proc. Natl. Acad. Sci. USA 1988, 85, 7312–7316. [Google Scholar] [CrossRef]
- Hayakawa, K.; Hardy, R.R.; Herzenberg, L.A. Progenitors for Ly-1 B cells are distinct from progenitors for other B cells. J. Exp. Med. 1985, 161, 1554–1568. [Google Scholar] [CrossRef]
- Griffin, D.O.; Holodick, N.E.; Rothstein, T.L. Human B1 cells in umbilical cord and adult peripheral blood express the novel phenotype CD20+CD27+CD43+CD70. J. Exp. Med. 2011, 208, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Montecino-Rodriguez, E.; Dorshkind, K. New perspectives in B-1 B cell development and function. Trends Immunol. 2006, 27, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Berland, R.; Wortis, H.H. Origins and functions of B-1 cells with notes on the role of CD5. Annu. Rev. Immunol. 2002, 20, 253–300. [Google Scholar] [CrossRef]
- Monroe, J.G.; Dorshkind, K. Fate Decisions Regulating Bone Marrow and Peripheral B Lymphocyte Development. Adv. Immunol. 2007, 95, 1–50. [Google Scholar] [CrossRef]
- Martin, F.; Kearney, J.F. Marginal-zone B cells. Nat. Rev. Immunol. 2002, 2, 323–335. [Google Scholar] [CrossRef]
- Pillai, S.; Cariappa, A.; Moran, S.T. Marginal Zone B Cells. Annu. Rev. Immunol. 2005, 23, 161–196. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.V.; Hancock, K.W. Antigenicity of trophoblast and possible antigen-masking effects during pregnancy. Immunology 1975, 28, 973–982. [Google Scholar] [PubMed]
- Kolb, J.P.; Chaouat, G.; Chassoux, D. Immunoactive products of placenta. III. Suppression of natural killing activity. J. Immunol. 1984, 132, 2305–2310. [Google Scholar]
- Margni, R.A.; Paz, C.B.; Cordal, M.E. Immunochemical behavior of sheep non-precipitating antibodies isolated by immunoadsorption. Immunochemistry 1976, 13, 209–214. [Google Scholar] [CrossRef]
- Canellada, A.; Farber, A.; Zenclussen, A.; Gentile, T.; Dokmetjian, J.; Keil, A.; Blois, S.; Miranda, S.; Berod, L.; Gutierrez, G.; et al. Interleukin Regulation of Asymmetric Antibody Synthesized by Isolated Placental B Cells. Am. J. Reprod. Immunol. 2002, 48, 275–282. [Google Scholar] [CrossRef]
- Kaneko, Y.; Hirose, S.; Abe, M.; Yagita, H.; Okumura, K.; Shirai, T. CD40-mediated stimulation of B1 and B2 cells: Implication in autoantibody production in murine lupus. Eur. J. Immunol. 1996, 26, 3061–3065. [Google Scholar] [CrossRef]
- Rolle, L.; Tehran, M.M.; Morell-García, A.; Raeva, Y.; Schumacher, A.; Hartig, R.; Costa, S.-D.; Jensen, F.; Zenclussen, A.C. Cutting Edge: IL-10-Producing Regulatory B Cells in Early Human Pregnancy. Am. J. Reprod. Immunol. 2013, 70, 448–453. [Google Scholar] [CrossRef]
- Wallukat, G.; Homuth, V.; Fischer, T.; Lindschau, C.; Horstkamp, B.; Jüpner, A.; Baur, E.; Nissen, E.; Vetter, K.; Neichel, D.; et al. Patients with preeclampsia develop agonistic autoantibodies against the angiotensin AT1 receptor. J. Clin. Investig. 1999, 103, 945–952. [Google Scholar] [CrossRef]
- Lamarca, B.; Wallukat, G.; Llinas, M.; Herse, F.; Dechend, R.; Granger, J.P. Autoantibodies to the Angiotensin Type I Receptor in Response to Placental Ischemia and Tumor Necrosis Factor α in Pregnant Rats. Hypertension 2008, 52, 1168–1172. [Google Scholar] [CrossRef] [PubMed]
- Parrish, M.R.; Murphy, S.R.; Rutland, S.; Wallace, K.; Wenzel, K.; Wallukat, G.; Keiser, S.; Ray, L.F.; Dechend, R.; Martin, J.N.; et al. The Effect of Immune Factors, Tumor Necrosis Factor-, and Agonistic Autoantibodies to the Angiotensin II Type I Receptor on Soluble fms-Like Tyrosine-1 and Soluble Endoglin Production in Response to Hypertension during Pregnancy. Am. J. Hypertens. 2010, 23, 911–916. [Google Scholar] [CrossRef]
- Zhou, C.C.; Zhang, Y.; Irani, R.A.; Zhang, H.; Mi, T.; Popek, E.J.; Hicks, M.J.; Ramin, S.M.; Kellems, R.E.; Xia, Y. Angiotensin receptor agonistic autoantibodies induce pre-eclampsia in pregnant mice. Nat. Med. 2008, 14, 855–862. [Google Scholar] [CrossRef] [PubMed]
- LaMarca, B.; Parrish, M.; Ray, L.F.; Murphy, S.R.; Roberts, L.; Glover, P.; Wallukat, G.; Wenzel, K.; Cockrell, K.; Martin, J.N., Jr.; et al. Hypertension in response to autoantibodies to the angiotensin II type I receptor (AT1-AA) in pregnant rats: Role of endothelin-1. Hypertension 2009, 54, 905–909. [Google Scholar] [CrossRef]
- Yang, X.; Wang, F.; Lau, W.B.; Zhang, S.; Zhang, S.; Liu, H.; Ma, X.-L. Autoantibodies Isolated from Preeclamptic Patients Induce Endothelial Dysfunction via Interaction with the Angiotensin II AT1 Receptor. Cardiovasc. Toxicol. 2013, 14, 21–29. [Google Scholar] [CrossRef]
- Yia, Y.; Wne, H.; Bobst, S.; Day, M.-C.; Kellems, R.E. Maternal Autoantibodies from Preeclamptic Patients Active Angiotensin Receptors on Human Trophoblast Cells. J. Soc. Gynecol. Investig. 2003, 10, 82–93. [Google Scholar] [CrossRef]
- Dechend, R.; Viedt, C.; Müller, D.N.; Ugele, B.; Brandes, R.P.; Wallukat, G.; Park, J.-K.; Janke, J.; Barta, P.; Theuer, J.; et al. AT 1 Receptor Agonistic Antibodies From Preeclamptic Patients Stimulate NADPH Oxidase. Circulation 2003, 107, 1632–1639. [Google Scholar] [CrossRef]
- Torricelli, M.; Sabatini, L.; Florio, P.; Scaccia, V.; Voltolini, C.; Biliotti, G.; De Bonis, M.; Petraglia, F. Levels of antibodies against protein C and protein S in pregnancy and in preeclampsia. J. Matern. Neonatal Med. 2009, 22, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Azlin, M.I.N.; Bakin, Y.D.; Mustafa, N.; Wahab, N.A.; Johari, M.J.M.; Kamarudin, N.A.; Jamil, M.A. Thyroid autoantibodies and associated complications during pregnancy. J. Obstet. Gynaecol. 2010, 30, 675–678. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P.; Haque, N. Reproductive immunomodulatory functions of B cells in pregnancy. Int. Rev. Immunol. 2019, 39, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Muzzio, D.; Zenclussen, A.C.; Jensen, F. The Role of B Cells in Pregnancy: The Good and the Bad. Am. J. Reprod. Immunol. 2013, 69, 408–412. [Google Scholar] [CrossRef]
- Fettke, F.; Schumacher, A.; Costa, S.-D.; Zenclussen, A.C. B Cells: The Old New Players in Reproductive Immunology. Front. Immunol. 2014, 5, 285. [Google Scholar] [CrossRef] [PubMed]
- Bhat, N.M.; Mithal, A.; Bieber, M.M.; Herzenberg, L.A.; Teng, N.N. Human CD5+ B lymphocytes (B-1 cells) decrease in peripheral blood during pregnancy. J. Reprod. Immunol. 1995, 28, 53–60. [Google Scholar] [CrossRef]
- Jensen, F.; Wallukat, G.; Herse, F.; Budner, O.; El-Mousleh, T.; Costa, S.-D.; Dechend, R.; Zenclussen, A.C. CD19+CD5+Cells as Indicators of Preeclampsia. Hypertension 2012, 59, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Eldeeb, S.M.; ElEdel, R.H.; Bassuoni, M.A.; Radwan, W.M.; Masoud, A. CD19+CD5+ B-cell expansion and risk of pre-eclampsia. Menoufia Med. J. 2016, 29, 319. [Google Scholar] [CrossRef]
- Liao, A.-H.; Liu, L.-P.; Ding, W.-P.; Zhang, L. ORIGINAL ARTICLE: Functional Changes of Human Peripheral B-Lymphocytes in Pre-Eclampsia. Am. J. Reprod. Immunol. 2009, 61, 313–321. [Google Scholar] [CrossRef]
- Magatti, M.; Masserdotti, A.; Signoroni, P.B.; Vertua, E.; Stefani, F.R.; Silini, A.R.; Parolini, O. B Lymphocytes as Targets of the Immunomodulatory Properties of Human Amniotic Mesenchymal Stromal Cells. Front. Immunol. 2020, 11, 1156. [Google Scholar] [CrossRef]
- Cargnoni, A.; Romele, P.; Signoroni, P.B.; Farigu, S.; Magatti, M.; Vertua, E.; Toschi, I.; Cesari, V.; Silini, A.R.; Stefani, F.R.; et al. Amniotic MSCs reduce pulmonary fibrosis by hampering lung B-cell recruitment, retention, and maturation. Stem Cells Transl. Med. 2020, 9, 1023–1035. [Google Scholar] [CrossRef]
- Magatti, M.; Vertua, E.; Cargnoni, A.; Silini, A.; Parolini, O. The Immunomodulatory Properties of Amniotic Cells: The Two Sides of the Coin. Cell Transplant. 2018, 27, 31–44. [Google Scholar] [CrossRef]
- Che, N.; Li, X.; Zhou, S.; Liu, R.; Shi, D.; Lu, L.; Sun, L. Umbilical cord mesenchymal stem cells suppress B-cell proliferation and differentiation. Cell. Immunol. 2012, 274, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.D.; Fong, C.Y.; Biswas, A.; Choolani, M.; Bongso, A. Human Wharton’s Jelly Stem Cells, its Conditioned Medium and Cell-Free Lysate Inhibit the Growth of Human Lymphoma Cells. Stem Cell Rev. Rep. 2014, 10, 573–586. [Google Scholar] [CrossRef]
- Ma, L.; Zhou, Z.; Zhang, D.; Yang, S.; Wang, J.; Xue, F.; Yang, Y.; Yang, R. Immunosuppressive function of mesenchymal stem cells from human umbilical cord matrix in immune thrombocytopenia patients. Thromb. Haemost. 2012, 107, 937–950. [Google Scholar] [CrossRef]
- Xue, Q.; Yin, Z.; Varshithreddy, N.; Liang, H.-S.; Wang, M.-Y.; Dong, W.-L.; Zhang, X.; Gu, Y.; Fang, Q. The immunomodulatory function of human amniotic fluid stromal cells on B lymphocytes. J. Neurorestoratol. 2018, 1, 122–133. [Google Scholar] [CrossRef]
- Morandi, F.; Horenstein, A.L.; Quarona, V.; Faini, A.C.; Castella, B.; Srinivasan, R.C.; Strom, S.C.; Malavasi, F.; Gramignoli, R. Ectonucleotidase Expression on Human Amnion Epithelial Cells: Adenosinergic Pathways and Dichotomic Effects on Immune Effector Cell Populations. J. Immunol. 2019, 202, 724–735. [Google Scholar] [CrossRef]
- Ribeiro, A.; Laranjeira, P.; Mendes, S.; Velada, I.; Leite, C.; Andrade, P.; Santos, F.; Henriques, A.; Grãos, M.; Cardoso, C.M.P.; et al. Mesenchymal stem cells from umbilical cord matrix, adipose tissue and bone marrow exhibit different capability to suppress peripheral blood B, natural killer and T cells. Stem Cell Res. Ther. 2013, 4, 125. [Google Scholar] [CrossRef]
- Ji, Y.R.; Yang, Z.X.; Han, Z.-B.; Meng, L.; Liang, L.; Feng, X.M.; Yang, S.G.; Chi, Y.; Chen, D.D.; Wang, Y.W.; et al. Mesenchymal Stem Cells Support Proliferation and Terminal Differentiation of B Cells. Cell. Physiol. Biochem. 2012, 30, 1526–1537. [Google Scholar] [CrossRef]
- LeBien, T.W.; Tedder, T.F. B lymphocytes: How they develop and function. Blood 2008, 112, 1570–1580. [Google Scholar] [CrossRef] [PubMed]
- Montecino-Rodriguez, E.; Leathers, H.; Dorshkind, K. Identification of a B-1 B cell–specified progenitor. Nat. Immunol. 2006, 7, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, D.A.; Wei, C.; Qian, Y.; Rosenberg, A.F.; Sanz, I. Advances in Human B Cell Phenotypic Profiling. Front. Immunol. 2012, 3, 302. [Google Scholar] [CrossRef] [PubMed]
- Shapiro-Shelef, M.; Calame, K. Regulation of plasma-cell development. Nat. Rev. Immunol. 2005, 5, 230–242. [Google Scholar] [CrossRef]
- Mittrücker, H.-W.; Matsuyama, T.; Grossman, A.; Kündig, T.M.; Potter, J.; Shahinian, A.; Wakeham, A.; Patterson, B.; Ohashi, P.S.; Mak, T.W. Requirement for the Transcription Factor LSIRF/IRF4 for Mature B and T Lymphocyte Function. Science 1997, 275, 540–543. [Google Scholar] [CrossRef]
- Reimold, A.M.; Iwakoshi, N.N.; Manis, J.; Vallabhajosyula, P.; Szomolanyi-Tsuda, E.; Gravallese, E.M.; Friend, D.; Grusby, M.J.; Alt, F.; Glimcher, L.H. Plasma cell differentiation requires the transcription factor XBP-1. Nature 2001, 412, 300–307. [Google Scholar] [CrossRef]
- Suthers, A.N.; Sarantopoulos, S. TLR7/TLR9- and B Cell Receptor-Signaling Crosstalk: Promotion of Potentially Dangerous B Cells. Front. Immunol. 2017, 8, 775. [Google Scholar] [CrossRef]
- De Nardo, D. Toll-like receptors: Activation, signalling and transcriptional modulation. Cytokine 2015, 74, 181–189. [Google Scholar] [CrossRef]
- Morbach, H.; Schickel, J.-N.; Cunningham-Rundles, C.; Conley, M.E.; Reisli, I.; Franco, J.L.; Meffre, E. CD19 controls Toll-like receptor 9 responses in human B cells. J. Allergy Clin. Immunol. 2016, 137, 889–898.e6. [Google Scholar] [CrossRef] [PubMed]
- Silini, A.R.; Masserdotti, A.; Papait, A.; Parolini, O. Shaping the Future of Perinatal Cells: Lessons From the Past and Interpretations of the Present. Front. Bioeng. Biotechnol. 2019, 7, 75. [Google Scholar] [CrossRef] [PubMed]
- Rossi, D.; Pianta, S.; Magatti, M.; Sedlmayr, P.; Parolini, O. Characterization of the Conditioned Medium from Amniotic Membrane Cells: Prostaglandins as Key Effectors of Its Immunomodulatory Activity. PLoS ONE 2012, 7, e46956. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, D.; Du, W.T.; Han, Z.B.; Ren, H.; Chi, Y.; Yang, S.G.; Zhu, D.; Bayard, F.; Han, Z.C. Human umbilical cord mesenchymal stem cells hUC-MSCs exert immunosuppressive activities through a PGE2-dependent mechanism. Clin. Immunol. 2010, 135, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Magatti, M.; Vertua, E.; De Munari, S.; Caro, M.; Caruso, M.; Silini, A.; Delgado, M.; Parolini, O. Human amnion favours tissue repair by inducing the M1-to-M2 switch and enhancing M2 macrophage features. J. Tissue Eng. Regen. Med. 2017, 11, 2895–2911. [Google Scholar] [CrossRef] [PubMed]
- Cargnoni, A.; Ressel, L.; Rossi, D.; Poli, A.; Arienti, D.; Lombardi, G.; Parolini, O. Conditioned medium from amniotic mesenchymal tissue cells reduces progression of bleomycin-induced lung fibrosis. Cytotherapy 2012, 14, 153–161. [Google Scholar] [CrossRef]
- Carbone, A.; Castellani, S.; Favia, M.; Diana, A.; Paracchini, V.; Di Gioia, S.; Seia, M.; Casavola, V.; Colombo, C.; Conese, M. Correction of defective CFTR/ENaC function and tightness of cystic fibrosis airway epithelium by amniotic mesenchymal stromal (stem) cells. J. Cell. Mol. Med. 2014, 18, 1631–1643. [Google Scholar] [CrossRef]
- Lee, P.-H.; Tu, C.-T.; Hsiao, C.-C.; Tsai, M.-S.; Ho, C.-M.; Cheng, N.-C.; Hung, T.-M.; Shih, D.T.-B. Antifibrotic Activity of Human Placental Amnion Membrane-Derived CD34+ Mesenchymal Stem/Progenitor Cell Transplantation in Mice with Thioacetamide-Induced Liver Injury. Stem Cells Transl. Med. 2016, 5, 1473–1484. [Google Scholar] [CrossRef]
- Parolini, O.; Souza-Moreira, L.; O’Valle, F.; Magatti, M.; Hernandez-Cortes, P.; Gonzalez-Rey, E.; Delgado, M. Therapeutic Effect of Human Amniotic Membrane-Derived Cells on Experimental Arthritis and Other Inflammatory Disorders. Arthritis Rheumatol. 2014, 66, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Tuca, A.-C.; Ertl, J.; Hingerl, K.; Pichlsberger, M.; Fuchs, J.; Wurzer, P.; Pfeiffer, D.; Bubalo, V.; Parvizi, D.; Kamolz, L.-P.; et al. Comparison of Matrigel and Matriderm as a carrier for human amnion-derived mesenchymal stem cells in wound healing. Placenta 2016, 48, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Pischiutta, F.; Brunelli, L.; Romele, P.; Silini, A.; Sammali, E.; Paracchini, L.; Marchini, S.; Talamini, L.; Bigini, P.; Boncoraglio, G.B.; et al. Protection of Brain Injury by Amniotic Mesenchymal Stromal Cell-Secreted Metabolites. Crit. Care Med. 2016, 44, e1118–e1131. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Ko, T.-L.; Shih, Y.-H.; Lin, M.-Y.A.; Fu, T.-W.; Hsiao, H.-S.; Hsu, J.-Y.C.; Fu, Y.-S. Human Umbilical Mesenchymal Stem Cells Promote Recovery After Ischemic Stroke. Stroke 2011, 42, 2045–2053. [Google Scholar] [CrossRef] [PubMed]
- Giampà, C.; Alvino, A.; Magatti, M.; Silini, A.R.; Cardinale, A.; Paldino, E.; Fusco, F.R.; Parolini, O. Conditioned medium from amniotic cells protects striatal degeneration and ameliorates motor deficits in the R6/2 mouse model of Huntington’s disease. J. Cell. Mol. Med. 2019, 23, 1581–1592. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Qiu, X.; NI, P.; Qiu, X.; Lin, X.; Wu, W.; Xie, L.; Lin, L.; Min, J.; Lai, X.; et al. Immunological characteristics of human umbilical cord mesenchymal stem cells and the therapeutic effects of their transplantion on hyperglycemia in diabetic rats. Int. J. Mol. Med. 2014, 33, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Hoyne, G.F.; Elliott, H.; Mutsaers, S.E.; Prêle, C.M. Idiopathic pulmonary fibrosis and a role for autoimmunity. Immunol. Cell Biol. 2017, 95, 577–583. [Google Scholar] [CrossRef]
- Tan, B.; Yuan, W.; Li, J.; Yang, P.; Ge, Z.; Liu, J.; Qiu, C.; Zhu, X.; Qiu, C.; Lai, D.; et al. Therapeutic effect of human amniotic epithelial cells in murine models of Hashimoto’s thyroiditis and Systemic lupus erythematosus. Cytotherapy 2018, 20, 1247–1258. [Google Scholar] [CrossRef] [PubMed]
- Hasham, A.; Tomer, Y. Genetic and epigenetic mechanisms in thyroid autoimmunity. Immunol. Res. 2012, 54, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Grammatikos, A.P.; Tsokos, G.C. Immunodeficiency and autoimmunity: Lessons from systemic lupus erythematosus. Trends Mol. Med. 2012, 18, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Zhang, Y.; Hu, B.; Liu, J.; Kong, P.; Lou, S.; Su, Y.; Yang, T.; Li, H.; Liu, Y.; et al. Phase II Multicenter, Randomized, Double-Blind Controlled Study of Efficacy and Safety of Umbilical Cord–Derived Mesenchymal Stromal Cells in the Prophylaxis of Chronic Graft-Versus-Host Disease After HLA-Haploidentical Stem-Cell Transplantation. J. Clin. Oncol. 2016, 34, 2843–2850. [Google Scholar] [CrossRef]
- Greinix, H.T.; Pohlreich, D.; Kouba, M.; Körmöczi, U.; Lohmann, I.; Feldmann, K.; Zielinski, C.C.; Pickl, W.F. Elevated Numbers of Immature/Transitional CD21− B Lymphocytes and Deficiency of Memory CD27+ B Cells Identify Patients with Active Chronic Graft-versus-Host Disease. Biol. Blood Marrow Transplant. 2008, 14, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Pallier, A.; Hillion, S.; Danger, R.; Giral, M.; Racapé, M.; Degauque, N.; Dugast, E.; Ashton-Chess, J.; Pettré, S.; Jos, J.; et al. Patients with drug-free long-term graft function display increased numbers of peripheral B cells with a memory and inhibitory phenotype. Kidney Int. 2010, 78, 503–513. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magatti, M.; Masserdotti, A.; Cargnoni, A.; Papait, A.; Stefani, F.R.; Silini, A.R.; Parolini, O. The Role of B Cells in PE Pathophysiology: A Potential Target for Perinatal Cell-Based Therapy? Int. J. Mol. Sci. 2021, 22, 3405. https://doi.org/10.3390/ijms22073405
Magatti M, Masserdotti A, Cargnoni A, Papait A, Stefani FR, Silini AR, Parolini O. The Role of B Cells in PE Pathophysiology: A Potential Target for Perinatal Cell-Based Therapy? International Journal of Molecular Sciences. 2021; 22(7):3405. https://doi.org/10.3390/ijms22073405
Chicago/Turabian StyleMagatti, Marta, Alice Masserdotti, Anna Cargnoni, Andrea Papait, Francesca Romana Stefani, Antonietta Rosa Silini, and Ornella Parolini. 2021. "The Role of B Cells in PE Pathophysiology: A Potential Target for Perinatal Cell-Based Therapy?" International Journal of Molecular Sciences 22, no. 7: 3405. https://doi.org/10.3390/ijms22073405
APA StyleMagatti, M., Masserdotti, A., Cargnoni, A., Papait, A., Stefani, F. R., Silini, A. R., & Parolini, O. (2021). The Role of B Cells in PE Pathophysiology: A Potential Target for Perinatal Cell-Based Therapy? International Journal of Molecular Sciences, 22(7), 3405. https://doi.org/10.3390/ijms22073405