
Megalocytivirus Induces Complicated Fish Immune Response at Multiple RNA Levels Involving mRNA, miRNA, and circRNA

Abstract
1. Introduction
2. Results
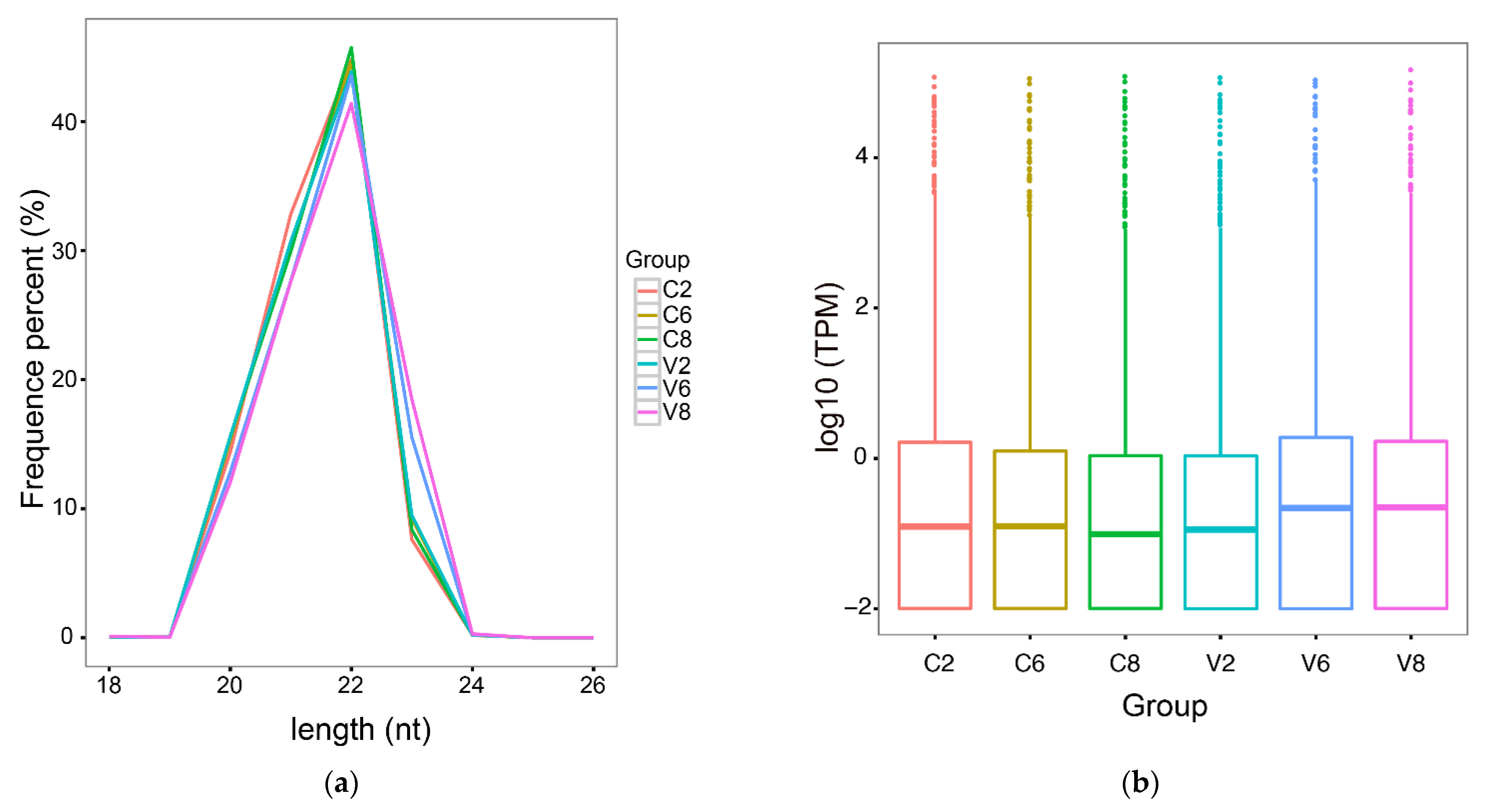
2.1. High-Throughput Sequencing and Quality Assessment
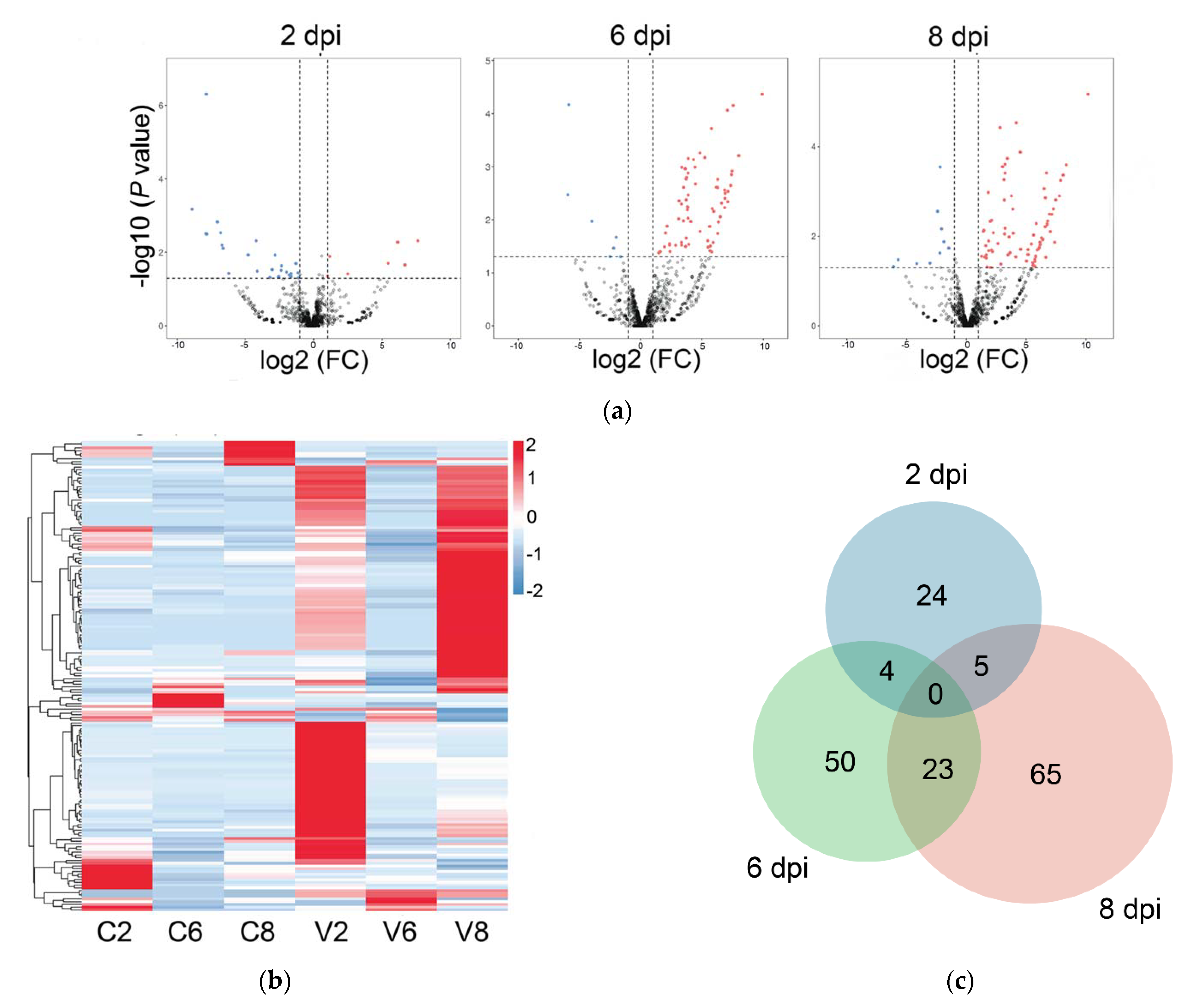
2.2. Differentially Expressed miRNAs (DEmiRs) Induced by Megalocytivirus
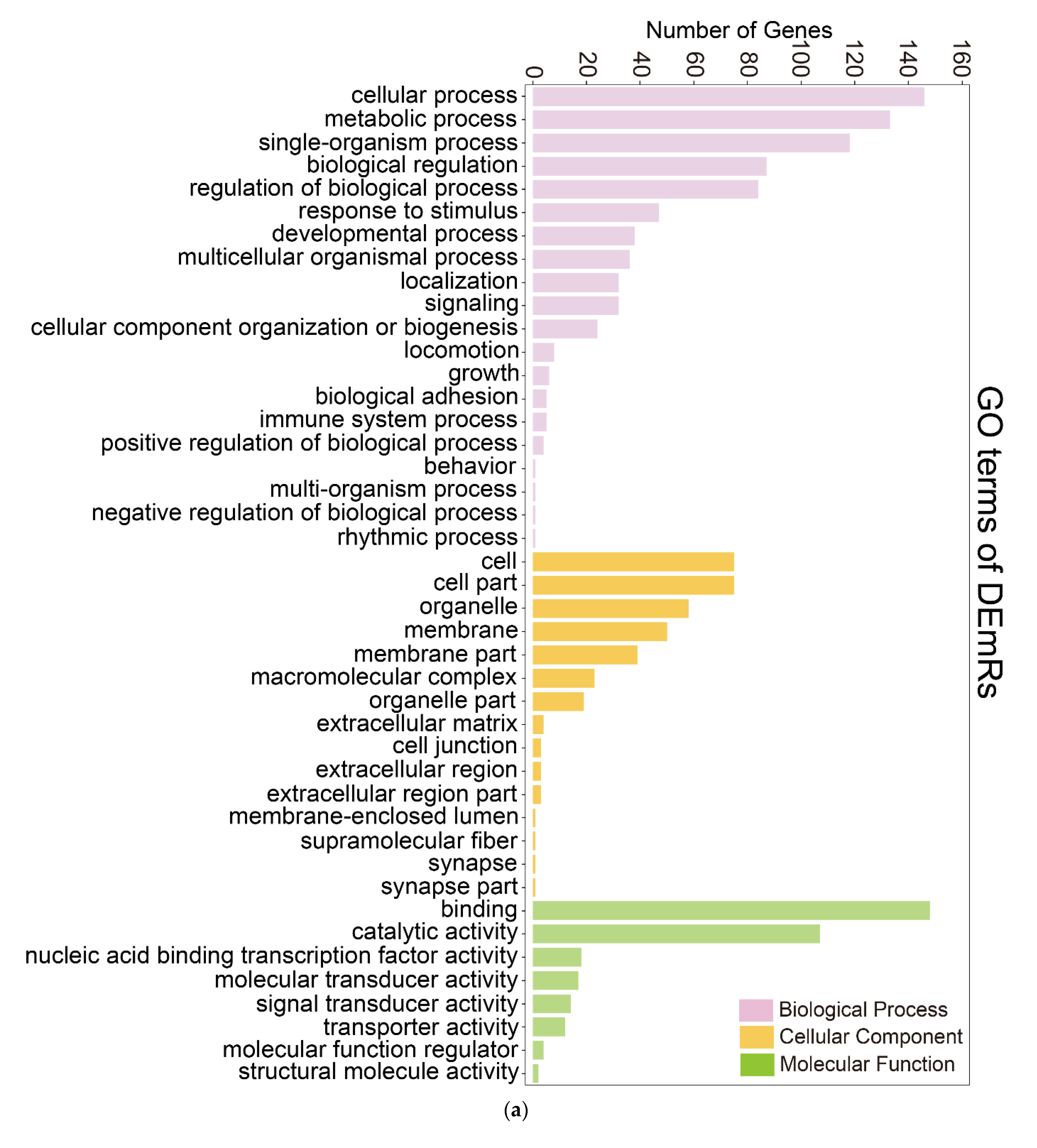
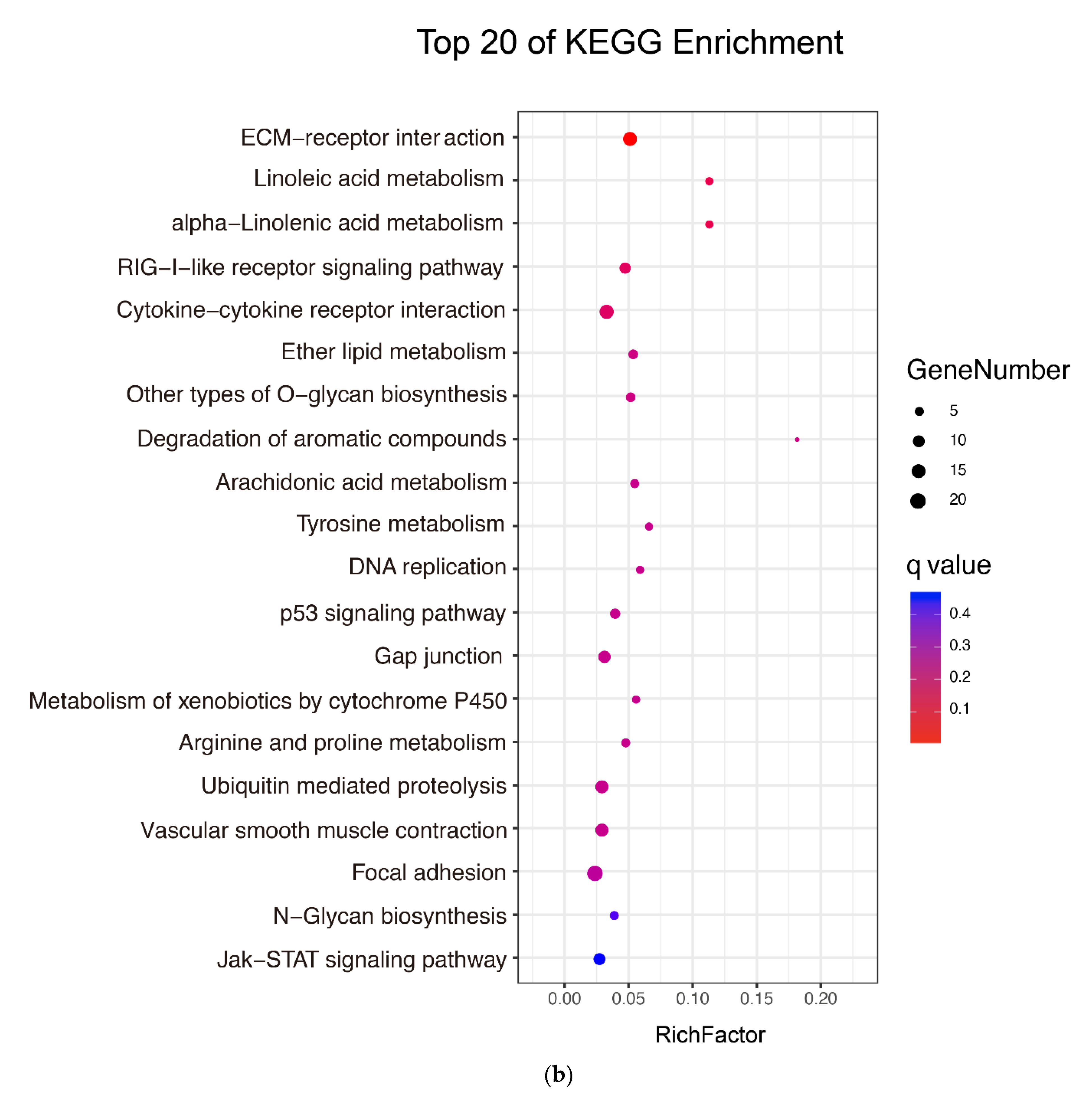
2.3. Identification and Analysis of the Targets of DEmiRs
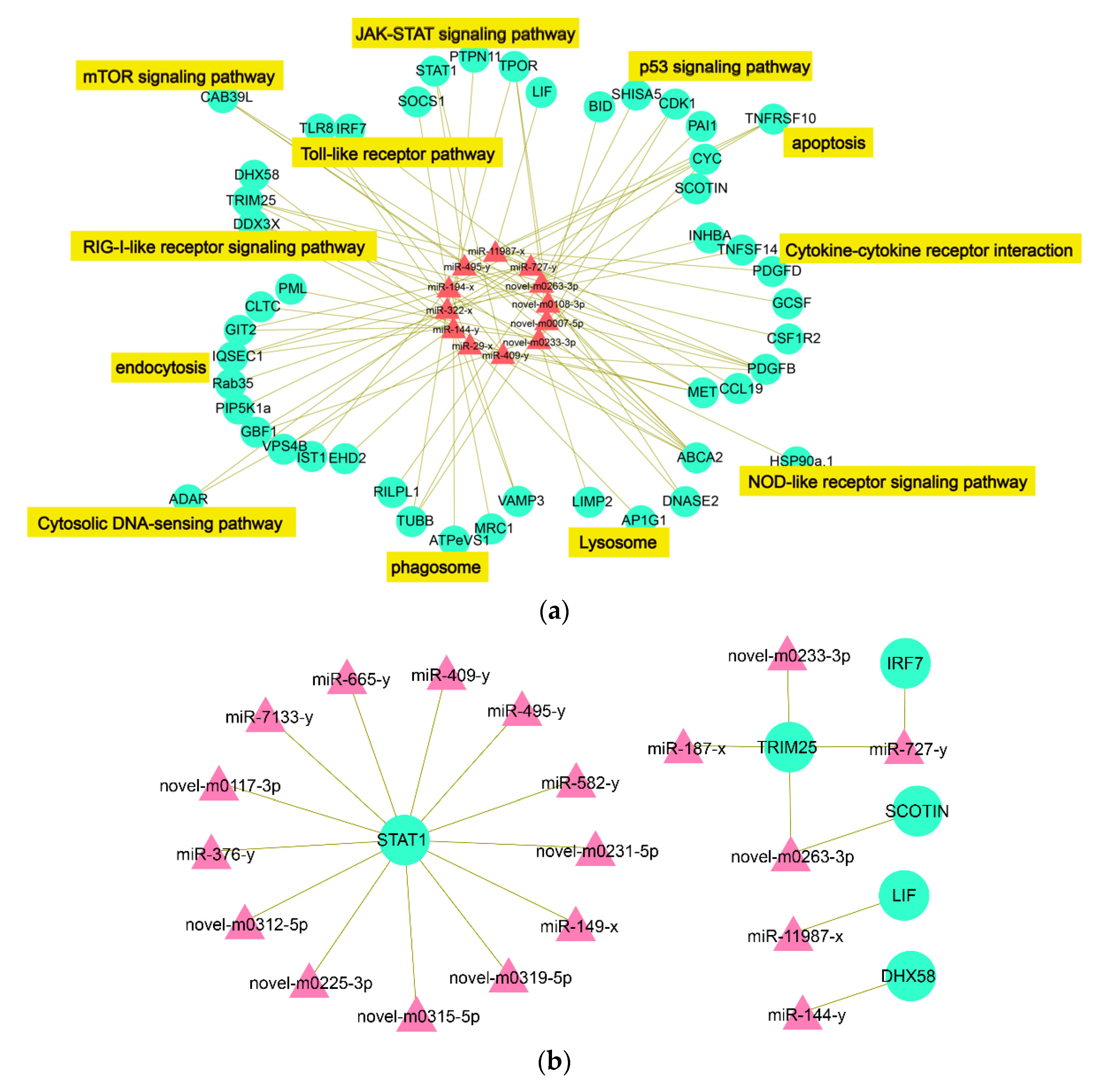
2.4. The Interactive Network of Immune-Related DEmiRs-DETmRs
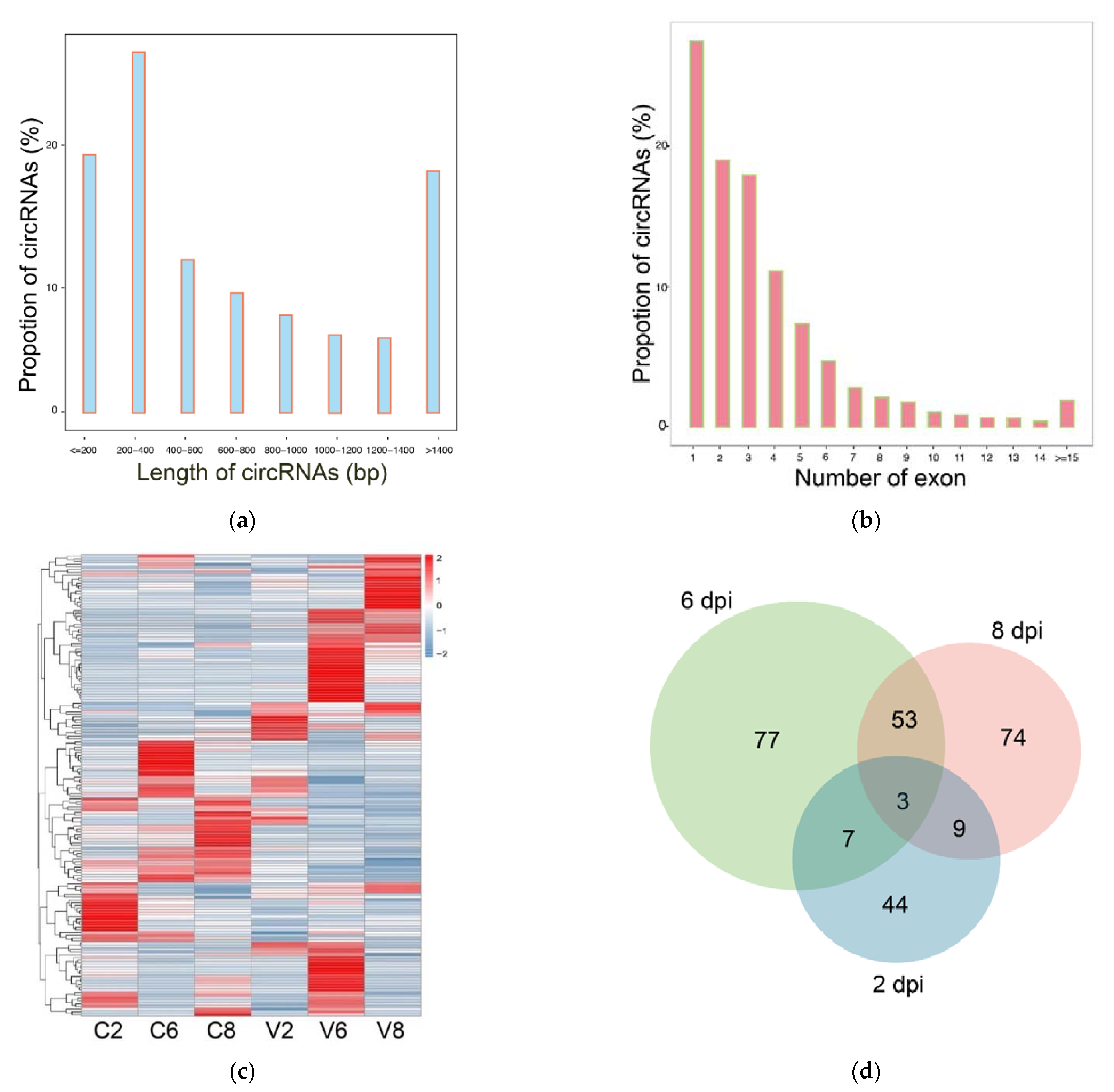
2.5. Overview of the circRNA Sequencing Data
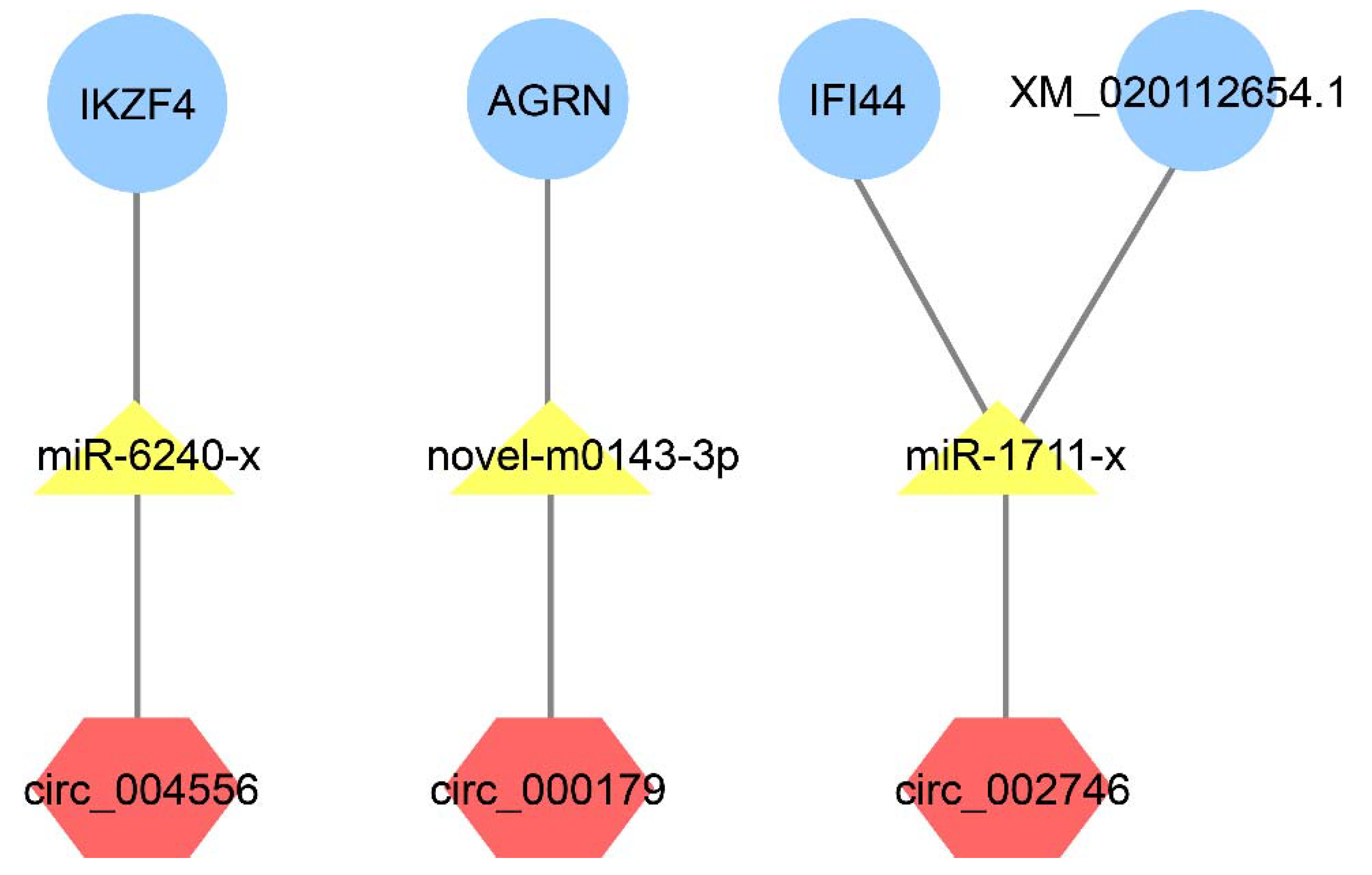
2.6. Competing Endogenous RNAs (ceRNAs)
3. Discussion
3.1. Endocytosis and Lysosome
3.2. Toll- and RIG-I-Like Receptor Signaling Pathways
3.3. Cytokine–Cytokine Receptor Interaction
3.4. JAK-STAT Signaling Pathway
4. Materials and Methods
4.1. Sample Collection
4.2. Small RNA Sequencing
4.3. Data Processing
4.4. MiRNA Identification and Differential Expression Analysis
4.5. CircRNA Identification and Differential Expression Analysis
4.6. Identification and Functional Enrichment Analysis of the DETmRs
4.7. Construction of Immune-Related DEmiR-DEmR Network
4.8. Identification of Interactive circRNA-miRNA and circRNA-miRNA-mRNA Regulatory Units
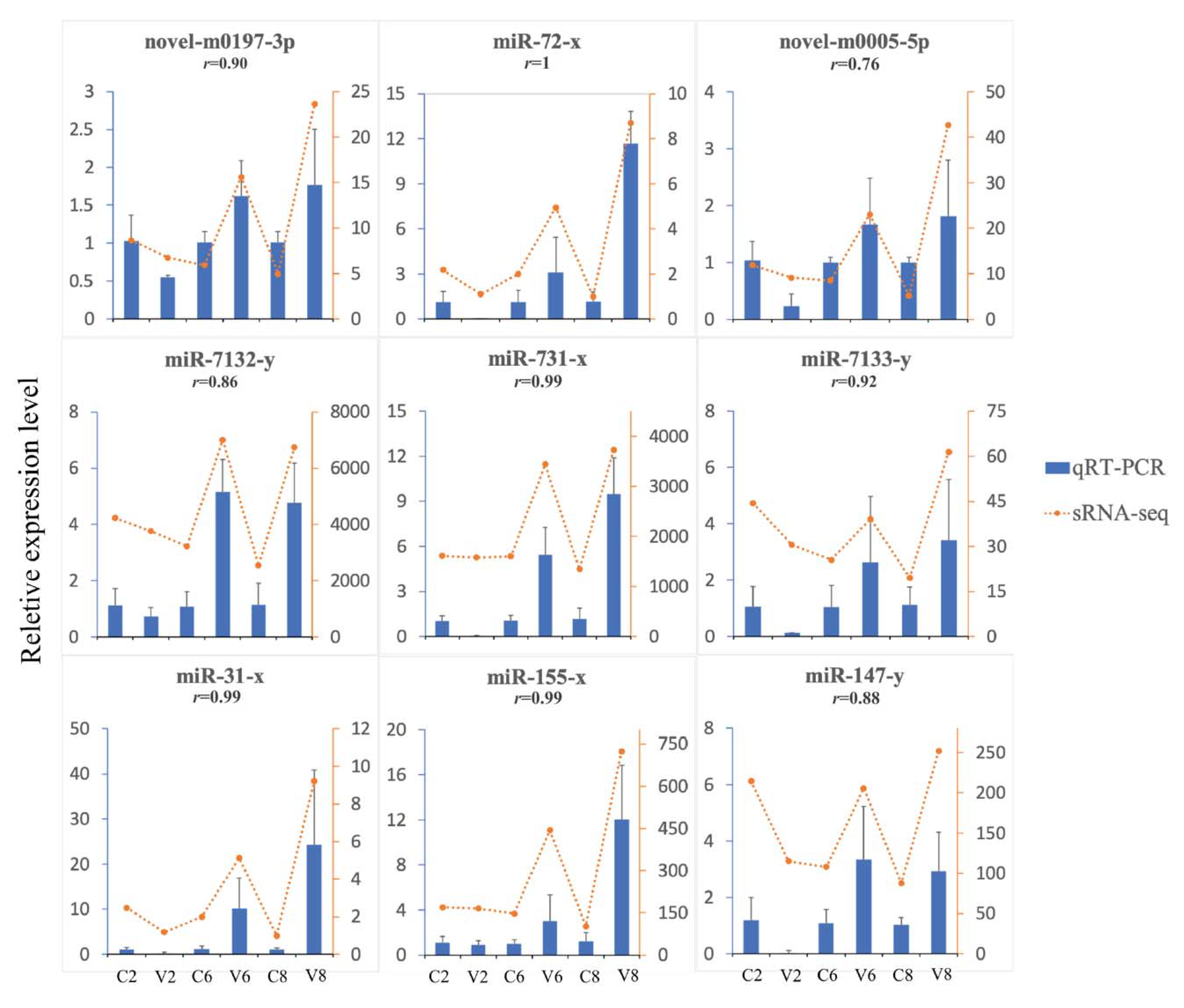
4.9. Experimental Validation of DEmiRs
4.10. Data Availability
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eaton, H.E.; Metcalf, J.; Penny, E.; Tcherepanov, V.; Upton, C.; Brunetti, C.R. Comparative genomic analysis of the family Iridoviridae: Re-annotating and defining the core set of iridovirus genes. Virol. J. 2007, 4, 11. [Google Scholar] [CrossRef]
- Kurita, J.N.K.; Hirono, I.; Aoki, T. Complete genome sequencing of red sea bream iridovirus (RSIV). Fish. Sci. 2002, 68, 1113–1115. [Google Scholar] [CrossRef]
- He, J.G.; Deng, M.; Weng, S.P.; Li, Z.; Zhou, S.Y.; Long, Q.X.; Wang, X.Z.; Chan, S.M. Complete genome analysis of the mandarin fish infectious spleen and kidney necrosis iridovirus. Virology 2001, 291, 126–139. [Google Scholar] [CrossRef]
- Do, J.W.; Moon, C.H.; Kim, H.J.; Ko, M.S.; Kim, S.B.; Son, J.H.; Kim, J.S.; An, E.J.; Kim, M.K.; Lee, S.K.; et al. Complete genomic DNA sequence of rock bream iridovirus. Virology 2004, 325, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Zhou, S.Y.; Chen, C.; Weng, S.P.; Chan, S.M.; He, J.G. Complete genome sequence analysis of an iridovirus isolated from the orange-spotted grouper, Epinephelus coioides. Virology 2005, 339, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.S.; Shih, H.H.; Ku, C.C.; Chen, S.N. Studies on epizootic iridovirus infection among red sea bream, Pagrus major (Temminck & Schlegel), cultured in Taiwan. J. Fish. Dis. 2003, 26, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Whittington, R.J.; Becker, J.A.; Dennis, M.M. Iridovirus infections in finfish—critical review with emphasis on ranaviruses. J. Fish. Dis. 2010, 33, 95–122. [Google Scholar] [CrossRef]
- Zhang, B.C.; Zhang, J.; Sun, L. In-depth profiling and analysis of host and viral microRNAs in Japanese flounder (Paralichthys olivaceus) infected with megalocytivirus reveal involvement of microRNAs in host-virus interaction in teleost fish. BMC Genom. 2014, 15, 878. [Google Scholar] [CrossRef] [PubMed]
- Kloosterman, W.P.; Plasterk, R.H. The diverse functions of microRNAs in animal development and disease. Dev. Cell 2006, 11, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Urbich, C.; Kuehbacher, A.; Dimmeler, S. Role of microRNAs in vascular diseases, inflammation, and angiogenesis. Cardiovasc. Res. 2008, 79, 581–588. [Google Scholar] [CrossRef]
- Slonchak, A.; Shannon, R.P.; Pali, G.; Khromykh, A.A. Human microRNA miR-532-5p exhibits antiviral activity against west nile virus via suppression of host genes SESTD1 and TAB3 required for virus replication. J. Virol. 2015, 90, 2388–2402. [Google Scholar] [CrossRef]
- Trobaugh, D.W.; Klimstra, W.B. MicroRNA regulation of RNA virus replication and pathogenesis. Trends Mol. Med. 2017, 23, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.C.; Zhou, Z.J.; Sun, L. pol-miR-731, a teleost miRNA upregulated by megalocytivirus, negatively regulates virus-induced type I interferon response, apoptosis, and cell cycle arrest. Sci. Rep. 2016, 6, 28354. [Google Scholar] [CrossRef]
- Hazra, B.; Kumawat, K.L.; Basu, A. The host microRNA miR-301a blocks the IRF1-mediated neuronal innate immune response to Japanese encephalitis virus infection. Sci. Signal. 2017, 10, eaaf5185. [Google Scholar] [CrossRef]
- Wang, B.; Fu, M.; Liu, Y.; Wang, Y.; Li, X.; Cao, H.; Zheng, S.J. gga-miR-155 enhances type I interferon expression and suppresses infectious burse disease virus replication via targeting SOCS1 and TANK. Front. Cell Infect. Microbiol. 2018, 8, 55. [Google Scholar] [CrossRef]
- Greene, J.; Baird, A.M.; Brady, L.; Lim, M.; Gray, S.G.; McDermott, R.; Finn, S.P. Circular RNAs: Biogenesis, function and role in human diseases. Front. Mol. Biosci. 2017, 4, 38. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Lin, W.; Guo, M.; Zou, Q. A comprehensive overview and evaluation of circular RNA detection tools. PLoS Comput. Biol. 2017, 13, e1005420. [Google Scholar] [CrossRef]
- Patop, I.L.; Wust, S.; Kadener, S. Past, present, and future of circRNAs. EMBO J. 2019, 38, e100836. [Google Scholar] [CrossRef] [PubMed]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Shao, T.; Pan, Y.-H.; Xiong, X.-D. Circular RNA: An important player with multiple facets to regulate its parental gene expression. Mol. Ther. Nucleic Acids 2021, 23, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, L.; Chen, L.L. The biogenesis, functions, and challenges of circular RNAs. Mol. Cell 2018, 71, 428–442. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.G.; Kim, M.V.; Chen, X.; Batista, P.J.; Aoyama, S.; Wilusz, J.E.; Iwasaki, A.; Chang, H.Y. Sensing self and foreign circular RNAs by intron identity. Mol. Cell 2017, 67, 228–238.e225. [Google Scholar] [CrossRef]
- Li, X.; Liu, C.X.; Xue, W.; Zhang, Y.; Jiang, S.; Yin, Q.F.; Wei, J.; Yao, R.W.; Yang, L.; Chen, L.L. Coordinated circRNA biogenesis and function with NF90/NF110 in viral infection. Mol. Cell 2017, 67, 214–227.e217. [Google Scholar] [CrossRef]
- Wang, M.; Yu, F.; Wu, W.; Zhang, Y.; Chang, W.; Ponnusamy, M.; Wang, K.; Li, P. Circular RNAs: A novel type of non-coding RNA and their potential implications in antiviral immunity. Int. J. Biol. Sci. 2017, 13, 1497–1506. [Google Scholar] [CrossRef]
- Chu, Q.; Zheng, W.; Su, H.; Zhang, L.; Chang, R.; Gao, W.; Xu, T. A Highly conserved circular RNA circRasGEF1B enhances antiviral immunity by regulating miR-21-3p/MITA pathway in lower vertebrates. J. Virol. 2021, JVI.02145-20. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Chu, Q.; Zheng, W.; Chang, R.; Gao, W.; Zhang, L.; Xu, T. Circular RNA circPIKfyve acts as a sponge of miR-21-3p to enhance antiviral immunity through regulating MAVS in teleost fish. J. Virol. 2021, JVI.02296-20. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Kang, H.; Liang, Q.J.; Hu, R.; Li, Z.H.; Liu, Y.; Wang, W.N. Integrative mRNA-miRNA interaction analysis associated with the immune response of Epinephelus coioddes to Vibrio alginolyticus infection. Fish. Shellfish Immunol. 2019, 90, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Sarropoulou, E.; Galindo-Villegas, J.; Garcia-Alcazar, A.; Kasapidis, P.; Mulero, V. Characterization of European sea bass transcripts by RNA SEQ after oral vaccine against V. anguillarum. Mar. Biotechnol. 2012, 14, 634–642. [Google Scholar] [CrossRef]
- Wu, Q.; Ning, X.; Jiang, S.; Sun, L. Transcriptome analysis reveals seven key immune pathways of Japanese flounder (Paralichthys olivaceus) involved in megalocytivirus infection. Fish. Shellfish Immunol. 2020, 103, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Gokhale, A.S.; Gangaplara, A.; Lopez-Occasio, M.; Thornton, A.M.; Shevach, E.M. Selective deletion of Eos (Ikzf4) in T-regulatory cells leads to loss of suppressive function and development of systemic autoimmunity. J. Autoimmun. 2019, 105, 102300. [Google Scholar] [CrossRef]
- Pan, F.; Yu, H.; Dang, E.V.; Barbi, J.; Pan, X.; Grosso, J.F.; Jinasena, D.; Sharma, S.M.; McCadden, E.M.; Getnet, D.; et al. Eos mediates Foxp3-dependent gene silencing in CD4+ regulatory T cells. Science 2009, 325, 1142–1146. [Google Scholar] [CrossRef]
- Rajkumar, T.; Sabitha, K.; Vijayalakshmi, N.; Shirley, S.; Bose, M.V.; Gopal, G.; Selvaluxmy, G. Identification and validation of genes involved in cervical tumourigenesis. BMC Cancer 2011, 11, 80. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Min, S.; Xie, F.; Yang, J.; Chen, J. Alleviating sepsis-induced neuromuscular dysfunction linked with acetylcholine receptors by agrin. J. Surg. Res. 2019, 241, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Power, D.; Santoso, N.; Dieringer, M.; Yu, J.; Huang, H.; Simpson, S.; Seth, I.; Miao, H.; Zhu, J. IFI44 suppresses HIV-1 LTR promoter activity and facilitates its latency. Virology 2015, 481, 142–150. [Google Scholar] [CrossRef]
- Busse, D.C.; Habgood-Coote, D.; Clare, S.; Brandt, C.; Bassano, I.; Kaforou, M.; Herberg, J.; Levin, M.; Eleouet, J.F.; Kellam, P.; et al. Interferon-induced protein 44 and interferon-induced protein 44-like restrict replication of respiratory syncytial virus. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Grove, J.; Marsh, M. The cell biology of receptor-mediated virus entry. J. Cell Biol. 2011, 195, 1071–1082. [Google Scholar] [CrossRef]
- Guo, C.J.; Wu, Y.Y.; Yang, L.S.; Yang, X.B.; He, J.; Mi, S.; Jia, K.T.; Weng, S.P.; Yu, X.Q.; He, J.G. Infectious spleen and kidney necrosis virus (a fish iridovirus) enters Mandarin fish fry cells via caveola-dependent endocytosis. J. Virol. 2012, 86, 2621–2631. [Google Scholar] [CrossRef]
- Wang, S.; Huang, X.; Huang, Y.; Hao, X.; Xu, H.; Cai, M.; Wang, H.; Qin, Q. Entry of a novel marine DNA virus, Singapore grouper iridovirus, into host cells occurs via clathrin-mediated endocytosis and macropinocytosis in a pH-dependent manner. J. Virol. 2014, 88, 13047–13063. [Google Scholar] [CrossRef]
- Mousavi, S.A.; Malerod, L.; Berg, T.; Kjeken, R. Clathrin-dependent endocytosis. Biochem. J. 2004, 377, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Zhu, L.; Chang, X.; Zhou, J.; Guo, R.; Zhao, Y.; Shi, D.; Niu, B.; Gu, J.; Yu, Z.; et al. Mortalin restricts porcine epidemic diarrhea virus entry by downregulating clathrin-mediated endocytosis. Vet. Microbiol. 2019, 239, 108455. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Xu, C.; Lei, C.; Hu, J.; Sun, X. Autographa californica multiple nucleopolyhedrovirus enters host cells via clathrin-mediated endocytosis and direct fusion with the plasma membrane. Viruses 2018, 10, 632. [Google Scholar] [CrossRef]
- Kouranti, I.; Sachse, M.; Arouche, N.; Goud, B.; Echard, A. Rab35 regulates an endocytic recycling pathway essential for the terminal steps of cytokinesis. Curr. Biol. 2006, 16, 1719–1725. [Google Scholar] [CrossRef] [PubMed]
- Minowa-Nozawa, A.; Nozawa, T.; Okamoto-Furuta, K.; Kohda, H.; Nakagawa, I. Rab35 GTPase recruits NDP52 to autophagy targets. EMBO J. 2017, 36, 3405. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, Y.; Tsuda, M.; Nanbo, A.; Hattori, T.; Sasaki, J.; Sasaki, T.; Miyazaki, T.; Ohba, Y. A Ca(2+)-dependent signalling circuit regulates influenza A virus internalization and infection. Nat. Commun. 2013, 4, 2763. [Google Scholar] [CrossRef]
- Vazquez-Calvo, A.; Sobrino, F.; Martin-Acebes, M.A. Plasma membrane phosphatidylinositol 4,5 bisphosphate is required for internalization of foot-and-mouth disease virus and vesicular stomatitis virus. PLoS ONE 2012, 7, e45172. [Google Scholar] [CrossRef][Green Version]
- Wang, K.; Yang, Z.; Liu, X.; Mao, K.; Nair, U.; Klionsky, D.J. Phosphatidylinositol 4-kinases are required for autophagic membrane trafficking. J. Biol. Chem. 2012, 287, 37964–37972. [Google Scholar] [CrossRef]
- Gee, K.; Zamora, D.; Horm, T.; George, L.; Upchurch, C.; Randall, J.; Weaver, C.; Sanford, C.; Miller, A.; Hernandez, S.; et al. Regulators of lysosome function and dynamics in caenorhabditis elegans. G3 (Bethesda) 2017, 7, 991–1000. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Settembre, C.; Ballabio, A. Lysosomal adaptation: How the lysosome responds to external cues. Cold Spring Harb. Perspect. Biol. 2014, 6. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Friedman, R.C.; Marquez, R.T.; Keck, K.; Kong, B.; Icardi, M.S.; Brown, K.E.; Burge, C.B.; Schmidt, W.N.; Wang, Y.; et al. Hepatitis C virus infection and hepatic stellate cell activation downregulate miR-29: miR-29 overexpression reduces hepatitis C viral abundance in culture. J. Infect. Dis. 2011, 203, 1753–1762. [Google Scholar] [CrossRef] [PubMed]
- Xing, T.J.; Jiang, D.F.; Huang, J.X.; Xu, Z.L. Expression and clinical significance of miR-122 and miR-29 in hepatitis B virus-related liver disease. Genet. Mol. Res. 2014, 13, 7912–7918. [Google Scholar] [CrossRef]
- Ortega, P.A.S.; Saulle, I.; Mercurio, V.; Ibba, S.V.; Lori, E.M.; Fenizia, C.; Masetti, M.; Trabattoni, D.; Caputo, S.L.; Vichi, F.; et al. Interleukin 21 (IL-21)/microRNA-29 (miR-29) axis is associated with natural resistance to HIV-1 infection. Aids 2018, 32, 2453–2461. [Google Scholar] [CrossRef]
- Tavares, L.A.; da Silva, E.M.; da Silva-Januario, M.E.; Januario, Y.C.; de Cavalho, J.V.; Czernisz, E.S.; Mardones, G.A.; da Silva, L.L. CD4 downregulation by the HIV-1 protein Nef reveals distinct roles for the gamma1 and gamma2 subunits of the AP-1 complex in protein trafficking. J. Cell Sci. 2017, 130, 429–443. [Google Scholar] [CrossRef]
- Kuronita, T.; Eskelinen, E.L.; Fujita, H.; Saftig, P.; Himeno, M.; Tanaka, Y. A role for the lysosomal membrane protein LGP85 in the biogenesis and maintenance of endosomal and lysosomal morphology. J. Cell Sci. 2002, 115, 4117–4131. [Google Scholar] [CrossRef] [PubMed]
- Tan, F.; Cao, M.; Ge, X.; Li, C.; Tian, M.; Zhang, L.; Fu, Q.; Song, L.; Yang, N. Identification and initial functional characterization of lysosomal integral membrane protein type 2 (LIMP-2) in turbot (Scophthalmus maximus L.). Dev. Comp. Immunol. 2019, 99, 103412. [Google Scholar] [CrossRef]
- Wang, K.; Lai, C.; Gu, H.; Zhao, L.; Xia, M.; Yang, P.; Wang, X. miR-194 inhibits innate antiviral immunity by targeting FGF2 in influenza H1N1 virus infection. Front. Microbiol. 2017, 8, 2187. [Google Scholar] [CrossRef]
- Khanizadeh, S.; Ravanshad, M.; Hosseini, S.Y.; Davoodian, P.; Almasian, M.; Khanlari, Z. The effect of the hepatitis C virus (HCV) NS3 protein on the expression of miR-150, miR-199a, miR-335, miR-194 and miR-27a. Microb. Pathog. 2017, 110, 688–693. [Google Scholar] [CrossRef]
- Mekky, R.Y.; El-Ekiaby, N.M.; Hamza, M.T.; Elemam, N.M.; El-Sayed, M.; Esmat, G.; Abdelaziz, A.I. Mir-194 is a hepatocyte gate keeper hindering HCV entry through targeting CD81 receptor. J. Infect. 2015, 70, 78–87. [Google Scholar] [CrossRef]
- Zhou, C.; Zhao, L.; Inagaki, N.; Guan, J.; Nakajo, S.; Hirabayashi, T.; Kikuyama, S.; Shioda, S. Atp-binding cassette transporter ABC2/ABCA2 in the rat brain: A novel mammalian lysosome-associated membrane protein and a specific marker for oligodendrocytes but not for myelin sheaths. J. Neurosci. 2001, 21, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Sakai, H.; Tanaka, Y.; Tanaka, M.; Ban, N.; Yamada, K.; Matsumura, Y.; Watanabe, D.; Sasaki, M.; Kita, T.; Inagaki, N. ABCA2 deficiency results in abnormal sphingolipid metabolism in mouse brain. J. Biol. Chem. 2007, 282, 19692–19699. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Kaisho, T.; Akira, S. Toll-like receptors. Annu. Rev. Immunol. 2003, 21, 335–376. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Villegas, J.; Garcia-Moreno, D.; de Oliveira, S.; Meseguer, J.; Mulero, V. Regulation of immunity and disease resistance by commensal microbes and chromatin modifications during zebrafish development. Proc. Natl. Acad. Sci. USA 2012, 109, E2605–E2614. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Villegas, J.; Montalban-Arques, A.; Liarte, S.; de Oliveira, S.; Pardo-Pastor, C.; Rubio-Moscardo, F.; Meseguer, J.; Valverde, M.A.; Mulero, V. TRPV4-mediated detection of hyposmotic stress by skin keratinocytes activates developmental immunity. J. Immunol. 2016, 196, 738–749. [Google Scholar] [CrossRef]
- Rosenberger, C.M.; Podyminogin, R.L.; Diercks, A.H.; Treuting, P.M.; Peschon, J.J.; Rodriguez, D.; Gundapuneni, M.; Weiss, M.J.; Aderem, A. miR-144 attenuates the host response to influenza virus by targeting the TRAF6-IRF7 signaling axis. PLoS Pathog. 2017, 13. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef]
- Da Silva, M.H.M.; Moises, R.N.C.; Alves, B.E.B.; Pereira, H.W.B.; de Paiva, A.A.P.; Morais, I.C.; Nascimento, Y.M.; Monteiro, J.D.; de Souto, J.T.; Nascimento, M.S.L.; et al. Innate immune response in patients with acute Zika virus infection. Med. Microbiol. Immunol. 2019, 208, 703–714. [Google Scholar] [CrossRef]
- Che, Q.; Wang, W.; Duan, P.; Fang, F.; Liu, C.; Zhou, T.; Li, H.; Xiong, C.; Zhao, K. Downregulation of miR-322 promotes apoptosis of GC-2 cell by targeting Ddx3x. Reprod. Biol. Endocrinol. 2019, 17, 63. [Google Scholar] [CrossRef]
- Oshiumi, H.; Sakai, K.; Matsumoto, M.; Seya, T. DEAD/H BOX 3 (DDX3) helicase binds the RIG-I adaptor IPS-1 to up-regulate IFN-beta-inducing potential. Eur. J. Immunol. 2010, 40, 940–948. [Google Scholar] [CrossRef]
- Charo, I.F.; Ransohoff, R.M. The many roles of chemokines and chemokine receptors in inflammation. N. Engl. J. Med. 2006, 354, 610–621. [Google Scholar] [CrossRef]
- O’Shea, J.J.; Holland, S.M.; Staudt, L.M. Mechanisms of disease JAKs and STATs in immunity, immunodeficiency, and cancer. N. Engl. J. Med. 2013, 368, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Secombes, C.J. The cytokine networks of adaptive immunity in fish. Fish. Shellfish Immunol. 2013, 35, 1703–1718. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Villegas, J.; Mulero, I.; Garcia-Alcazar, A.; Munoz, I.; Penalver-Mellado, M.; Streitenberger, S.; Scapigliati, G.; Meseguer, J.; Mulero, V. Recombinant TNFalpha as oral vaccine adjuvant protects European sea bass against vibriosis: Insights into the role of the CCL25/CCR9 axis. Fish. Shellfish Immunol. 2013, 35, 1260–1271. [Google Scholar] [CrossRef]
- De Oliveira, S.; Lopez-Munoz, A.; Martinez-Navarro, F.J.; Galindo-Villegas, J.; Mulero, V.; Calado, A. Cxcl8-l1 and Cxcl8-l2 are required in the zebrafish defense against Salmonella Typhimurium. Dev. Comp. Immunol. 2015, 49, 44–48. [Google Scholar] [CrossRef]
- Yan, Y.; Chen, R.; Wang, X.; Hu, K.; Huang, L.; Lu, M.; Hu, Q. CCL19 and CCR7 expression, signaling pathways, and adjuvant functions in viral infection and prevention. Front. Cell Dev. Biol. 2019, 7, 212. [Google Scholar] [CrossRef]
- Morris, D.; Ansar, M.; Speshock, J.; Ivanciuc, T.; Qu, Y.; Casola, A.; Garofalo, R. Antiviral and immunomodulatory activity of silver nanoparticles in experimental RSV infection. Viruses 2019, 11, 732. [Google Scholar] [CrossRef] [PubMed]
- Du, H.H.; Huang, H.Q.; Si, K.W.; Dai, H.F.; Hu, Y.H. Granulocyte colony stimulating factor (GCSF) of Japanese flounder (Paralichthys olivaceus): Immunoregulatory property and anti-infectious function. Fish. Shellfish Immunol. 2019, 89, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Mauri, D.N.; Ebner, R.; Montgomery, R.I.; Kochel, K.D.; Cheung, T.C.; Yu, G.L.; Ruben, S.; Murphy, M.; Eisenberg, R.J.; Cohen, G.H.; et al. LIGHT, a new member of the TNF superfamily, and lymphotoxin alpha are ligands for herpesvirus entry mediator. Immunity 1998, 8, 21–30. [Google Scholar] [CrossRef]
- Renauld, J.C. Class II cytokine receptors and their ligands: Key antiviral and inflammatory modulators. Nat. Rev. Immunol. 2003, 3, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.P.; Zhang, Y.J. Antagonizing cytokine-mediated JAK-STAT signaling by porcine reproductive and respiratory syndrome virus. Vet. Microbiol. 2017, 209, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.B.; Xu, W.; Chen, Y.N.; Xie, X.P.; Zhang, Y.C.; Ma, C.Q.; Yang, Q.Y.; Han, Y.; Zhu, C.L.; Xiong, Y.; et al. Matrix metalloproteinase 9 facilitates hepatitis B virus replication through binding with type I interferon (IFN) receptor 1 to repress IFN/JAK/STAT signaling. J. Virol. 2017, 91. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, F.; Yang, Y.; Wang, W.; Niu, L.; Zuo, D.; Li, X.; Hua, H.; Zhang, B.; Kou, Y.; et al. MiR-409-3p and MiR-1896 co-operatively participate in IL-17-induced inflammatory cytokine production in astrocytes and pathogenesis of EAE mice via targeting SOCS3/STAT3 signaling. Glia 2019, 67, 101–112. [Google Scholar] [CrossRef]
- Liau, N.P.D.; Laktyushin, A.; Lucet, I.S.; Murphy, J.M.; Yao, S.; Whitlock, E.; Callaghan, K.; Nicola, N.A.; Kershaw, N.J.; Babon, J.J. The molecular basis of JAK/STAT inhibition by SOCS1. Nat. Commun. 2018, 9, 1558. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, C.; Xue, M.; Fu, F.; Zhang, X.; Li, L.; Yin, L.; Xu, W.; Feng, L.; Liu, P. The coronavirus transmissible gastroenteritis virus evades the type I interferon response through IRE1alpha-mediated manipulation of the microRNA miR-30a-5p/SOCS1/3 Axis. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- Chitranshi, N.; Dheer, Y.; Gupta, V.; Abbasi, M.; Mirzaei, M.; You, Y.; Chung, R.; Graham, S.L.; Gupta, V. PTPN11 induces endoplasmic stress and apoptosis in SH-SY5Y cells. Neuroscience 2017, 364, 175–189. [Google Scholar] [CrossRef]
- Ruess, D.A.; Heynen, G.J.; Ciecielski, K.J.; Ai, J.; Berninger, A.; Kabacaoglu, D.; Gorgulu, K.; Dantes, Z.; Wormann, S.M.; Diakopoulos, K.N.; et al. Mutant KRAS-driven cancers depend on PTPN11/SHP2 phosphatase. Nat. Med. 2018, 24, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Qu, C.K. Protein tyrosine phosphatases in the JAK/STAT pathway. Front. Biosci. 2008, 13, 4925–4932. [Google Scholar] [CrossRef]
- Langmead, B. Aligning short sequencing reads with Bowtie. Curr. Protoc. Bioinform. 2010. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, Q.; Wang, J.; Guo, X.; Jiang, X.; Ren, Z.; Weng, C.; Sun, G.; Wang, X.; Liu, Y.; et al. Identification and characterization of novel amphioxus microRNAs by Solexa sequencing. Genome Biol. 2009, 10, R78. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.S.; Shi, Z.Y.; Wu, M.L.; Zhang, J.L.; Jia, L.; Chen, X.W. Identification and differential expression of microRNAs during metamorphosis of the Japanese flounder (Paralichthys olivaceus). PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Clean Tags (%) | Known miRNA | Novel miRNA |
|---|---|---|---|
| V2d-1 | 11,837,245 (96.83%) | 365 | 259 |
| V2d-2 | 11,280,092 (96.22%) | 374 | 239 |
| V2d-3 | 9,924,270 (96.10%) | 382 | 247 |
| V6d-1 | 17,710,805 (96.20%) | 518 | 296 |
| V6d-2 | 10,365,825 (96.37%) | 425 | 239 |
| V6d-3 | 11,893,288 (96.59%) | 451 | 272 |
| V8d-1 | 12,212,869 (96.67%) | 489 | 286 |
| V8d-2 | 13,140,335 (95.92%) | 365 | 288 |
| V8d-3 | 13,010,734 (95.98%) | 476 | 293 |
| C2d-1 | 12,573,839 (96.38%) | 432 | 248 |
| C2d-2 | 8,926,038 (94.98%) | 364 | 233 |
| C2d-3 | 8,576,422 (94.70%) | 368 | 225 |
| C6d-1 | 11,653,795 (96.21%) | 413 | 235 |
| C6d-2 | 14,097,705 (97.33%) | 367 | 260 |
| C6d-3 | 12,708,286 (95.14%) | 396 | 261 |
| C8d-1 | 10,633,416 (95.26%) | 351 | 260 |
| C8d-2 | 11,449,436 (95.92%) | 357 | 218 |
| C8d-3 | 11,412,190 (94.55%) | 394 | 228 |
| Time Point | Up | Down | Total |
|---|---|---|---|
| 2 dpi | 7 | 26 | 33 |
| 6 dpi | 69 | 8 | 77 |
| 8 dpi | 83 | 10 | 93 |
| Hub DEmiR | Fold Change (Log2) | Degree | DETmR | ||
|---|---|---|---|---|---|
| 2 dpi | 6 dpi | 8 dpi | |||
| novel-m0263-3p | −5.95 | −4.16 | 11 | CDK1, CSF1R2, DNASE2, INHBB, IQSEC1, RAB35, RILPL1, PAI1, SCOTIN, SHISA5, TRIM25 | |
| novel-m0233-3p | −6.21 | −6.09 | 10 | CDK1, CLTC, CYC, DNASE2, GBF1, LIMP2, PML, BID, TRIM25, HSP90α.1 | |
| miR-322-x | 5.66 | 7.14 | 9 | ADAR, DDX3X, TNFSF14, TPOR, PIP5K1α, TUBB, TLR8, TNFRSF10, VAMP3 | |
| miR-144-y | −2.20 | 8 | ATPeVS1, DHX58, GIT2, IQSEC1, MRC1, PTPN11, SOCS1, VPS4B | ||
| miR-409-y | 6.39 | 7 | ABCA2, DDX3X, MET, PDGFB, STAT1, TUBB, TLR8 | ||
| miR-11987-x | 2.50 | 2.64 | 7 | ABCA2, CCL19, CYC, PDGFB, PDGFD, GCSF, LIF | |
| miR-495-y | 5.78 | 6 | ABCA2, CAB39L, MET, PDGFB, STAT1, TNFRSF10 | ||
| novel-m0108-3p | 3.23 | 6 | ABCA2, CAB39L, DDX3X, MET, TPOR, PDGFB | ||
| miR-29-x | 2.15 | 2.12 | 5 | ABCA2, ADAR, AP1G1, EHD2, MET | |
| miR-194-x | 4.51 | 5 | ABCA2, DDX3X, IST1, TNFRSF10, VAMP3 | ||
| miR-727-y | −1.89 | 5 | GBF1, GIT2, IQSEC1, IRF7, TRIM25 | ||
| novel-m0007-5p | 5.43 | 5 | ABCA2, CAB39L, TPOR, PDGFB, TUBB | ||
| Time Point | Up | Down | Total |
|---|---|---|---|
| 2 dpi | 19 | 25 | 44 |
| 6 dpi | 46 | 31 | 77 |
| 8 dpi | 41 | 33 | 74 |
| MicroRNA | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|
| novel-m0197-3p | GACCACCCCCGAGCTTCTACGA | GTATCAACGCAGAGTACTTT |
| novel-m0005-5p | TACCACCCCCGAGCTTCTGCGA | GTATCAACGCAGAGTACTTT |
| miR-731-x | AATGACACGTTTTCTCCCGGATT | GTATCAACGCAGAGTACTTT |
| miR-72-x | AGGCAAGATGTTGGCATAGCT | GTATCAACGCAGAGTACTTT |
| miR-7133-y | TAGTTTGATACACAGCACAATG | GTATCAACGCAGAGTACTTT |
| miR-7132-x | GACTTGGTCAAAGCTCCTCAGC | GTATCAACGCAGAGTACTTT |
| miR-31-x | AGGCAAGATGTTGGCATAGCT | GTATCAACGCAGAGTACTTT |
| miR-155-x | TTAATGCTAATCGTGATAGGGGT | GTATCAACGCAGAGTACTTT |
| miR-147-y | GTGTGCGGAAAAGCTTCTGCTC | GTATCAACGCAGAGTACTTT |
| 5s | CCATACCACCCTGAACAC | CGGTCTCCCATCCAAGTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.; Ning, X.; Sun, L. Megalocytivirus Induces Complicated Fish Immune Response at Multiple RNA Levels Involving mRNA, miRNA, and circRNA. Int. J. Mol. Sci. 2021, 22, 3156. https://doi.org/10.3390/ijms22063156
Wu Q, Ning X, Sun L. Megalocytivirus Induces Complicated Fish Immune Response at Multiple RNA Levels Involving mRNA, miRNA, and circRNA. International Journal of Molecular Sciences. 2021; 22(6):3156. https://doi.org/10.3390/ijms22063156
Chicago/Turabian StyleWu, Qian, Xianhui Ning, and Li Sun. 2021. "Megalocytivirus Induces Complicated Fish Immune Response at Multiple RNA Levels Involving mRNA, miRNA, and circRNA" International Journal of Molecular Sciences 22, no. 6: 3156. https://doi.org/10.3390/ijms22063156
APA StyleWu, Q., Ning, X., & Sun, L. (2021). Megalocytivirus Induces Complicated Fish Immune Response at Multiple RNA Levels Involving mRNA, miRNA, and circRNA. International Journal of Molecular Sciences, 22(6), 3156. https://doi.org/10.3390/ijms22063156