
Omics Approaches in Adipose Tissue and Skeletal Muscle Addressing the Role of Extracellular Matrix in Obesity and Metabolic Dysfunction

 ,
, 
 ,
,  , and
, and 
Abstract
1. Introduction
2. Methodology
2.1. Systematic Review Strategy
2.2. Study Selection
3. Current Evidence of Adipose and SM ECM Remodeling in Obesity and Metabolic Dysfunction
4. Whole-Genome Genetic Approaches Reveal the Implication of ECM in Obesity and Metabolic Dysfunction
5. Whole-Genome DNA Methylation Approaches Reveal the Implication of ECM in Obesity and Metabolic Dysfunction
6. Whole-Genome Gene Expression Approaches Reveal the Implication of ECM in Obesity and Metabolic Dysfunction
7. Conclusions and Further Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schoettl, T.; Fischer, I.P.; Ussar, S. Heterogeneity of adipose tissue in development and metabolic function. J. Exp. Biol. 2018, 221 (Suppl. 1), jeb162958. [Google Scholar] [CrossRef]
- Sun, K.; Kusminski, C.M.; Scherer, P.E. Adipose tissue remodeling and obesity. J. Clin. Invest. 2011, 121, 2094–2101. [Google Scholar] [CrossRef]
- Sun, K.; Tordjman, J.; Clement, K.; Scherer, P.E. Fibrosis and adipose tissue dysfunction. Cell Metab. 2013, 18, 470–477. [Google Scholar] [CrossRef]
- Ruiz-Ojeda, F.J.; Méndez-Gutiérrez, A.; Aguilera, C.M.; Plaza-Díaz, J. Extracellular Matrix Remodeling of Adipose Tissue in Obesity and Metabolic Diseases. Int. J. Mol. Sci. 2019, 20, 4888. [Google Scholar] [CrossRef]
- Cox, T.R.; Erler, J.T. Remodeling and homeostasis of the extracellular matrix: Implications for fibrotic diseases and cancer. Dis. Model. Mech. 2011, 4, 165–178. [Google Scholar] [CrossRef]
- Lin, D.; Chun, T.H.; Kang, L. Adipose extracellular matrix remodelling in obesity and insulin resistance. Biochem. Pharm. 2016, 119, 8–16. [Google Scholar] [CrossRef]
- Williams, A.S.; Kang, L.; Wasserman, D.H. The extracellular matrix and insulin resistance. Trends Endocrinol. Metab. 2015, 26, 357–366. [Google Scholar] [CrossRef]
- Ruiz-Ojeda, F.J.; Wang, J.; Backer, T.; Krueger, M.; Zamani, S.; Rosowski, S.; Gruber, T.; Onogi, Y.; Feuchtinger, A.; Schulz, T.J.; et al. Active integrins regulate white adipose tissue insulin sensitivity and brown fat thermogenesis. Mol. Metab. 2021, 101147. [Google Scholar] [CrossRef]
- Bluher, M. Are there still healthy obese patients? Curr. Opin. Endocrinol. Diabetes Obes. 2012, 19, 341–346. [Google Scholar] [CrossRef]
- Mazzali, G.; Di Francesco, V.; Zoico, E.; Fantin, F.; Zamboni, G.; Benati, C.; Bambara, V.; Negri, M.; Bosello, O.; Zamboni, M. Interrelations between fat distribution, muscle lipid content, adipocytokines, and insulin resistance: Effect of moderate weight loss in older women. Am. J. Clin. Nutr. 2006, 84, 1193–1199. [Google Scholar] [CrossRef]
- Guillet, C.; Delcourt, I.; Rance, M.; Giraudet, C.; Walrand, S.; Bedu, M.; Duche, P.; Boirie, Y. Changes in basal and insulin and amino acid response of whole body and skeletal muscle proteins in obese men. J. Clin. Endocrinol. Metab. 2009, 94, 3044–3050. [Google Scholar] [CrossRef]
- Richardson, D.K.; Kashyap, S.; Bajaj, M.; Cusi, K.; Mandarino, S.J.; Finlayson, J.; DeFronzo, R.A.; Jenkinson, C.P.; Mandarino, L.J. Lipid Infusion Decreases the Expression of Nuclear Encoded Mitochondrial Genes and Increases the Expression of Extracellular Matrix Genes in Human Skeletal Muscle. J. Biol. Chem. 2005, 280, 10290–10297. [Google Scholar] [CrossRef]
- Berria, R.; Wang, L.; Richardson, D.K.; Finlayson, J.; Belfort, R.; Pratipanawatr, T.; De Filippis, E.A.; Kashyap, S.; Mandarino, L.J. Increased collagen content in insulin-resistant skeletal muscle. Am. J. Physiol Endocrinol. Metab 2006, 290, E560–E565. [Google Scholar] [CrossRef]
- Van Dijk, S.J.; Tellam, R.L.; Morrison, J.L.; Muhlhausler, B.S.; Molloy, P.L. Recent developments on the role of epigenetics in obesity and metabolic disease. Clin. Epigenet. 2015, 7, 66. [Google Scholar] [CrossRef]
- Davegardh, C.; Garcia-Calzon, S.; Bacos, K.; Ling, C. DNA methylation in the pathogenesis of type 2 diabetes in humans. Mol. Metab. 2018, 14, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Ling, C.; Ronn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 29, 1028–1044. [Google Scholar] [CrossRef] [PubMed]
- Gancheva, S.; Ouni, M.; Jelenik, T.; Koliaki, C.; Szendroedi, J.; Toledo, F.G.S.; Markgraf, D.F.; Pesta, D.H.; Mastrototaro, L.; De Filippo, E.; et al. Dynamic changes of muscle insulin sensitivity after metabolic surgery. Nat. Commun. 2019, 10, 4179. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Prisma Group. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Shamseer, L.; Moher, D.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015: Elaboration and explanation. BMJ Br. Med. J. 2015, 349, g7647. [Google Scholar] [CrossRef]
- Nakajima, I.; Yamaguchi, T.; Ozutsumi, K.; Aso, H. Adipose tissue extracellular matrix: Newly organized by adipocytes during differentiation. Differentiation 1998, 63, 193–200. [Google Scholar] [CrossRef]
- Mori, S.; Kiuchi, S.; Ouchi, A.; Hase, T.; Murase, T. Characteristic expression of extracellular matrix in subcutaneous adipose tissue development and adipogenesis; comparison with visceral adipose tissue. Int. J. Biol. Sci. 2014, 10, 825–833. [Google Scholar] [CrossRef]
- Poltavets, V.; Kochetkova, M.; Pitson, S.M.; Samuel, M.S. The Role of the Extracellular Matrix and Its Molecular and Cellular Regulators in Cancer Cell Plasticity. Front. Oncol. 2018, 8, 431. [Google Scholar] [CrossRef]
- Liu, X.; Xu, Q.; Liu, W.; Yao, G.; Zhao, Y.; Xu, F.; Hayashi, T.; Fujisaki, H.; Hattori, S.; Tashiro, S.-I.; et al. Enhanced migration of murine fibroblast-like 3T3-L1 preadipocytes on type I collagen-coated dish is reversed by silibinin treatment. Mol. Cell. Biochem. 2018, 441, 35–62. [Google Scholar] [CrossRef]
- Mariman, E.C.M.; Wang, P. Adipocyte extracellular matrix composition, dynamics and role in obesity. Cell. Mol. Life Sci. 2010, 67, 1277–1292. [Google Scholar] [CrossRef]
- Huber, J.; Löffler, M.; Bilban, M.; Reimers, M.; Kadl, A.; Todoric, J.; Zeyda, M.; Geyeregger, R.; Schreiner, M.; Weichhart, T.; et al. Prevention of high-fat diet-induced adipose tissue remodeling in obese diabetic mice by n-3 polyunsaturated fatty acids. Int. J. Obes. 2007, 31, 1004–1013. [Google Scholar] [CrossRef]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef]
- Bourboulia, D.; Stetler-Stevenson, W.G. Matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs): Positive and negative regulators in tumor cell adhesion. Semin. Cancer Biol. 2010, 20, 161–168. [Google Scholar] [CrossRef]
- Maquoi, E.; Munaut, C.; Colige, A.; Collen, D.; Lijnen, H.R. Modulation of Adipose Tissue Expression of Murine Matrix Metalloproteinases and Their Tissue Inhibitors with Obesity. Diabetes 2002, 51, 1093–1101. [Google Scholar] [CrossRef]
- Miksztowicz, V.; Morales, C.; Zago, V.; Friedman, S.; Schreier, L.; Berg, G. Effect of insulin-resistance on circulating and adipose tissue MMP-2 and MMP-9 activity in rats fed a sucrose-rich diet. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 294–300. [Google Scholar] [CrossRef]
- Murphy, G. Tissue inhibitors of metalloproteinases. Genome Biol. 2011, 12, 233. [Google Scholar] [CrossRef]
- Hopps, E.; Lo Presti, R.; Montana, M.; Noto, D.; Averna, M.R.; Caimi, G. Gelatinases and their tissue inhibitors in a group of subjects with metabolic syndrome. J. Investig. Med. 2013, 61, 978–983. [Google Scholar] [CrossRef]
- Jaworski, D.M.; Sideleva, O.; Stradecki, H.M.; Langlois, G.D.; Habibovic, A.; Satish, B.; Tharp, W.G.; Lausier, J.; LaRock, K.; Jetton, T.L.; et al. Sexually Dimorphic Diet-Induced Insulin Resistance in Obese Tissue Inhibitor of Metalloproteinase-2 (TIMP-2)-Deficient Mice. Endocrinology 2011, 152, 1300–1313. [Google Scholar] [CrossRef]
- Duarte, F.O.; Gomes-Gatto, C.D.V.; Oishi, J.C.; Lino, A.D.D.S.; Stotzer, U.S.; Rodrigues, M.F.C.; Gatti da Silva, G.H.; Selistre-de-Araújo, H.S. Physical training improves visceral adipose tissue health by remodelling extracellular matrix in rats with estrogen absence: A gene expression analysis. Int. J. Exp. Pathol. 2017, 98, 203–213. [Google Scholar] [CrossRef]
- Frontera, W.R.; Ochala, J. Skeletal muscle: A brief review of structure and function. Calcif. Tissue Int. 2015, 96, 183–195. [Google Scholar] [CrossRef]
- Gillies, A.R.; Lieber, R.L. Structure and function of the skeletal muscle extracellular matrix. Muscle Nerve 2011, 44, 318–331. [Google Scholar] [CrossRef]
- Martinez-Huenchullan, S.; McLennan, S.V.; Verhoeven, A.; Twigg, S.M.; Tam, C.S. The emerging role of skeletal muscle extracellular matrix remodelling in obesity and exercise. Obes. Rev. 2017, 18, 776–790. [Google Scholar] [CrossRef]
- Light, N.; Champion, A.E. Characterization of muscle epimysium, perimysium and endomysium collagens. Biochem. J. 1984, 219, 1017–1026. [Google Scholar] [CrossRef]
- Marvulli, D.; Volpin, D.; Bressan, G.M. Spatial and temporal changes of type VI collagen expression during mouse development. Dev. Dyn. 1996, 206, 447–454. [Google Scholar] [CrossRef]
- Weng, X.; Lin, D.; Huang, J.T.J.; Stimson, R.H.; Wasserman, D.H.; Kang, L. Collagen 24 α1 Is Increased in Insulin-Resistant Skeletal Muscle and Adipose Tissue. Int. J. Mol. Sci. 2020, 21, 5738. [Google Scholar] [CrossRef]
- Alameddine, H.S.; Morgan, J.E. Matrix Metalloproteinases and Tissue Inhibitor of Metalloproteinases in Inflammation and Fibrosis of Skeletal Muscles. J. Neuromuscul. Dis. 2016, 3, 455–473. [Google Scholar] [CrossRef]
- Kang, L.; Mayes, W.H.; James, F.D.; Bracy, D.P.; Wasserman, D.H. Matrix metalloproteinase 9 opposes diet-induced muscle insulin resistance in mice. Diabetologia 2014, 57, 603–613. [Google Scholar] [CrossRef]
- Knoblauch, A.; Will, C.; Goncharenko, G.; Ludwig, S.; Wixler, V. The binding of Mss4 to alpha-integrin subunits regulates matrix metalloproteinase activation and fibronectin remodeling. FASEB J. 2007, 21, 497–510. [Google Scholar] [CrossRef]
- Liu, J.; DeYoung, S.M.; Zhang, M.; Zhang, M.; Cheng, A.; Saltiel, A.R. Changes in integrin expression during adipocyte differentiation. Cell Metab. 2005, 2, 165–177. [Google Scholar] [CrossRef]
- Zong, H.; Bastie, C.C.; Xu, J.; Fassler, R.; Campbell, K.P.; Kurland, I.J.; Pessin, J.E. Insulin resistance in striated muscle-specific integrin receptor beta1-deficient mice. J. Biol. Chem. 2009, 284, 4679–4688. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Ayala, J.E.; Lee-Young, R.S.; Zhang, Z.; James, F.D.; Neufer, P.D.; Pozzi, A.; Zutter, M.M.; Wasserman, D.H. Diet-Induced Muscle Insulin Resistance Is Associated with Extracellular Matrix Remodeling and Interaction with Integrin α2β1 in Mice. Diabetes 2011, 60, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Pessentheiner, A.R.; Quach, A.; Al-Azzam, N.; Liu, S.; Downes, M.; Evans, R.M.; Gordts, P.L.M.S. Adipose tissue heparan sulfate proteoglycans—Critical regulators of adipocyte metabolism and glucose homeostasis. FASEB J. 2020, 34 (Suppl. 1), 1. [Google Scholar] [CrossRef]
- Brandan, E.; Gutierrez, J. Role of skeletal muscle proteoglycans during myogenesis. Matrix Biol. 2013, 32, 289–297. [Google Scholar] [CrossRef]
- Kamiya, S.; Kato, R.; Wakabayashi, M.; Tohyama, T.; Enami, I.; Ueki, M.; Yajima, H.; Ishii, T.; Nakamura, H.; Katayama, T.; et al. Fibronectin peptides derived from two distinct regions stimulate adipocyte differentiation by preventing fibronectin matrix assembly. Biochemistry 2002, 41, 3270–3277. [Google Scholar] [CrossRef]
- Kühl, U.; Öcalan, M.; Timpl, R.; von der Mark, K. Role of laminin and fibronectin in selecting myogenic versus fibrogenic cells from skeletal muscle cells in vitro. Dev. Biol. 1986, 117, 628–635. [Google Scholar] [CrossRef]
- Leshem, Y.; Spicer, D.B.; Gal-Levi, R.; Halevy, O. Hepatocyte growth factor (HGF) inhibits skeletal muscle cell differentiation: A role for the bHLH protein twist and the cdk inhibitor p27. J. Cell Physiol. 2000, 184, 101–109. [Google Scholar] [CrossRef]
- Saiki, A.; Watanabe, F.; Murano, T.; Miyashita, Y.; Shirai, K. Hepatocyte growth factor secreted by cultured adipocytes promotes tube formation of vascular endothelial cells in vitro. Int. J. Obes. 2006, 30, 1676–1684. [Google Scholar] [CrossRef][Green Version]
- Adachi, H.; Kurachi, H.; Homma, H.; Adachi, K.; Imai, T.; Morishige, K.; Matsuzawa, Y.; Miyake, A. Epidermal growth factor promotes adipogenesis of 3T3-L1 cell in vitro. Endocrinology 1994, 135, 1824–1830. [Google Scholar] [CrossRef]
- Leroy, M.C.; Perroud, J.; Darbellay, B.; Bernheim, L.; Konig, S. Epidermal growth factor receptor down-regulation triggers human myoblast differentiation. PLoS ONE 2013, 8, e71770. [Google Scholar] [CrossRef]
- Miyata, S.; Yada, T.; Ishikawa, N.; Taheruzzaman, K.; Hara, R.; Matsuzaki, T.; Nishikawa, A. Insulin-like growth factor 1 regulation of proliferation and differentiation of Xenopus laevis myogenic cells in vitro. In Vitro Cell. Dev. Biol. Anim. 2017, 53, 231–247. [Google Scholar] [CrossRef] [PubMed]
- Garten, A.; Schuster, S.; Kiess, W. The insulin-like growth factors in adipogenesis and obesity. Endocrinol. Metab. Clin. N. Am. 2012, 41, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Prelle, K.; Wobus, A.M.; Krebs, O.; Blum, W.F.; Wolf, E. Overexpression of Insulin-like Growth Factor-II in Mouse Embryonic Stem Cells Promotes Myogenic Differentiation. Biochem. Biophys. Res. Commun. 2000, 277, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Wang, F.; Huang, J.; Lou, Y.; Xie, J.; Li, H.; Cao, D.; Huang, X. Insulin-like growth factor 2 promotes the adipogenesis of hemangioma-derived stem cells. Exp. Ther. Med. 2019, 17, 1663–1669. [Google Scholar] [CrossRef] [PubMed]
- Lilla, J.; Stickens, D.; Werb, Z. Metalloproteases and adipogenesis: A weighty subject. Am. J. Pathol. 2002, 160, 1551–1554. [Google Scholar] [CrossRef]
- Chen, X.; Li, Y. Role of matrix metalloproteinases in skeletal muscle: Migration, differentiation, regeneration and fibrosis. Cell Adh. Migr. 2009, 3, 337–341. [Google Scholar] [CrossRef]
- Ahmad, K.; Choi, I.; Lee, Y.H. Implications of Skeletal Muscle Extracellular Matrix Remodeling in Metabolic Disorders: Diabetes Perspective. Int. J. Mol. Sci. 2020, 21, 3845. [Google Scholar] [CrossRef]
- Ghanemi, A.; Melouane, A.; Yoshioka, M.; St-Amand, J. Secreted protein acidic and rich in cysteine and bioenergetics: Extracellular matrix, adipocytes remodeling and skeletal muscle metabolism. Int. J. Biochem. Cell Biol. 2019, 117, 105627. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.S.; Heard-Costa, N.; Cupples, L.A.; Dupuis, J.; Vasan, R.S.; Atwood, L.D. Genome-wide association to body mass index and waist circumference: The Framingham Heart Study 100K project. BMC Med. Genet. 2007, 8 (Suppl. 1), S18. [Google Scholar] [CrossRef] [PubMed]
- León-Mimila, P.; Villamil-Ramírez, H.; Villalobos-Comparán, M.; Villarreal-Molina, T.; Romero-Hidalgo, S.; López-Contreras, B.; Gutiérrez-Vidal, R.; Vega-Badillo, J.; Jacobo-Albavera, L.; Posadas-Romeros, C.; et al. Contribution of common genetic variants to obesity and obesity-related traits in mexican children and adults. PLoS ONE 2013, 8, e70640. [Google Scholar]
- Hinney, A.; Albayrak, Ö.; Antel, J.; Volckmar, A.-L.; Sims, R.; Chapman, J.; Harold, D.; Gerrish, A.; Heid, I.M.; Winkler, T.W.; et al. Genetic variation at the CELF1 (CUGBP, elav-like family member 1 gene) locus is genome-wide associated with Alzheimer’s disease and obesity. Am. J. Med. Genet. Part. B Neuropsychiatr. Genet. 2014, 165, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ban, M.R.; Zou, G.Y.; Cao, H.; Lin, T.; Kennedy, B.A.; Anand, S.; Yusuf, S.; Huff, M.W.; Pollex, R.L.; et al. Polygenic determinants of severe hypertriglyceridemia. Hum. Mol. Genet. 2008, 17, 2894–2899. [Google Scholar] [CrossRef]
- Chiang, K.-M.; Chang, H.-C.; Yang, H.-C.; Chen, C.-H.; Chen, H.-H.; Lee, W.-J.; Pan, W.-H. Genome-wide association study of morbid obesity in Han Chinese. BMC Genet. 2019, 20, 97. [Google Scholar] [CrossRef]
- Comuzzie, A.G.; Cole, S.A.; Laston, S.L.; Voruganti, V.S.; Haack, K.; Gibbs, R.A.; Butte, N.F. Novel Genetic Loci Identified for the Pathophysiology of Childhood Obesity in the Hispanic Population. PLoS ONE 2012, 7, e51954. [Google Scholar] [CrossRef]
- Namjou, B.; Keddache, M.; Marsolo, K.; Wagner, M.; Lingren, T.; Cobb, B.; Perry, C.; Kennebeck, S.; Holm, I.; Li, R.; et al. EMR-linked GWAS study: Investigation of variation landscape of loci for body mass index in children. Front. Genet. 2013, 4, 268. [Google Scholar] [CrossRef]
- Pei, Y.-F.; Zhang, L.; Liu, Y.; Li, J.; Shen, H.; Liu, Y.-Z.; Tian, Q.; He, H.; Wu, S.; Ran, S.; et al. Meta-analysis of genome-wide association data identifies novel susceptibility loci for obesity. Hum. Mol. Genet. 2013, 23, 820–830. [Google Scholar] [CrossRef]
- Wu, Y.; Duan, H.; Tian, X.; Xu, C.; Wang, W.; Jiang, W.; Pang, Z.; Zhang, D.; Tan, Q. Genetics of Obesity Traits: A Bivariate Genome-Wide Association Analysis. Front. Genet. 2018, 9, 179. [Google Scholar] [CrossRef]
- Naukkarinen, J.; Surakka, I.; Pietiläinen, K.H.; Rissanen, A.; Salomaa, V.; Ripatti, S.; Yki-Järvinen, H.; van Duijn, C.M.; Wichmann, H.E.; Kaprio, J.; et al. Use of Genome-Wide Expression Data to Mine the “Gray Zone” of GWA Studies Leads to Novel Candidate Obesity Genes. PLoS Genet. 2010, 6, e1000976. [Google Scholar] [CrossRef]
- Li, W.-D.; Jiao, H.; Wang, K.; Zhang, C.K.; Glessner, J.T.; Grant, S.F.A.; Zhao, H.; Hakonarson, H.; Arlen Price, R. A genome wide association study of plasma uric acid levels in obese cases and never-overweight controls. Obesity 2013, 21, E490–E494. [Google Scholar] [CrossRef]
- Liu, H.Y.; Alyass, A.; Abadi, A.; Peralta-Romero, J.; Suarez, F.; Gomez-Zamudio, J.; Audirac, A.; Parra, E.J.; Cruz, M.; Meyre, D. Fine-mapping of 98 obesity loci in Mexican children. Int. J. Obes. 2019, 43, 23–32. [Google Scholar] [CrossRef]
- Zhao, W.; Wineinger, N.E.; Tiwari, H.K.; Mosley, T.H.; Broeckel, U.; Arnett, D.K.; Kardia, S.L.R.; Kabagambe, E.K.; Sun, Y.V. Copy Number Variations Associated with Obesity-Related Traits in African Americans: A Joint Analysis between GENOA and HyperGEN. Obesity 2012, 20, 2431–2437. [Google Scholar] [CrossRef]
- Liu, C.-T.; Monda, K.L.; Taylor, K.C.; Lange, L.; Demerath, E.W.; Palmas, W.; Wojczynski, M.K.; Ellis, J.C.; Vitolins, M.Z.; Liu, S.; et al. Genome-Wide Association of Body Fat Distribution in African Ancestry Populations Suggests New Loci. PLoS Genet. 2013, 9, e1003681. [Google Scholar] [CrossRef]
- Felix, J.F.; Bradfield, J.P.; Monnereau, C.; van der Valk, R.J.P.; Stergiakouli, E.; Chesi, A.; Gaillard, R.; Feenstra, B.; Thiering, E.; Kreiner-Møller, E.; et al. Genome-wide association analysis identifies three new susceptibility loci for childhood body mass index. Hum. Mol. Genet. 2015, 25, 389–403. [Google Scholar] [CrossRef]
- Kong, X.; Zhang, X.; Xing, X.; Zhang, B.; Hong, J.; Yang, W. The Association of Type 2 Diabetes Loci Identified in Genome-Wide Association Studies with Metabolic Syndrome and Its Components in a Chinese Population with Type 2 Diabetes. PLoS ONE 2015, 10, e0143607. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.; Wang, K.; Yang, F.; Grant, S.F.A.; Hakonarson, H.; Price, R.A.; Li, W.-D. Pathway-Based Genome-Wide Association Studies for Plasma Triglycerides in Obese Females and Normal-Weight Controls. PLoS ONE 2015, 10, e0134923. [Google Scholar] [CrossRef]
- Strawbridge, R.J.; Dupuis, J.; Prokopenko, I.; Barker, A.; Ahlqvist, E.; Rybin, D.; Petrie, J.R.; Travers, M.E.; Bouatia-Naji, N.; Dimas, A.S.; et al. Genome-wide association identifies nine common variants associated with fasting proinsulin levels and provides new insights into the pathophysiology of type 2 diabetes. Diabetes 2011, 60, 2624–2634. [Google Scholar] [CrossRef]
- Choquet, H.; Labrune, Y.; De Graeve, F.; Hinney, A.; Hebebrand, J.; Scherag, A.; Lecoeur, C.; Tauber, M.; Balkau, B.; Elliot, P.; et al. Lack of association of CD36 SNPs with early onset obesity: A meta-analysis in 9,973 European subjects. Obesity (Silver Spring) 2011, 19, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.C.; Hester, J.M.; Wing, M.R.; Li, J.; Xu, J.; Hicks, P.J.; Roh, B.H.; Lu, L.; Divers, J.; Langefeld, C.D.; et al. Genome-wide association of BMI in African Americans. Obesity 2012, 20, 622–627. [Google Scholar] [CrossRef]
- Wilson, C.L.; Liu, W.; Yang, J.J.; Kang, G.; Ojha, R.P.; Neale, G.A.; Srivastava, D.K.; Gurney, J.G.; Hudson, M.M.; Robison, L.L.; et al. Genetic and clinical factors associated with obesity among adult survivors of childhood cancer: A report from the St. Jude Lifetime Cohort. Cancer 2015, 121, 2262–2270. [Google Scholar] [CrossRef]
- Kocarnik, J.M.; Pendergrass, S.A.; Carty, C.L.; Pankow, J.S.; Schumacher, F.R.; Cheng, I.; Durda, P.; Ambite, J.L.; Deelman, E.; Cook, N.R.; et al. Multiancestral analysis of inflammation-related genetic variants and C-reactive protein in the population architecture using genomics and epidemiology study. Circ. Cardiovasc. Genet. 2014, 7, 178–188. [Google Scholar] [CrossRef]
- Ortega-Azorín, C.; Coltell, O.; Asensio, E.M.; Sorlí, J.V.; González, J.I.; Portolés, O.; Saiz, C.; Estruch, R.; Ramírez-Sabio, J.B.; Pérez-Fidalgo, A.; et al. Candidate Gene and Genome-Wide Association Studies for Circulating Leptin Levels Reveal Population and Sex-Specific Associations in High Cardiovascular Risk Mediterranean Subjects. Nutrients 2019, 11, 2751. [Google Scholar] [CrossRef]
- Hotta, K.; Kitamoto, A.; Kitamoto, T.; Mizusawa, S.; Teranishi, H.; Matsuo, T.; Nakata, Y.; Hyogo, H.; Ochi, H.; Nakamura, T.; et al. Genetic variations in the CYP17A1 and NT5C2 genes are associated with a reduction in visceral and subcutaneous fat areas in Japanese women. J. Hum. Genet. 2012, 57, 46–51. [Google Scholar] [CrossRef]
- Demerath, E.W.; Liu, C.-T.; Franceschini, N.; Chen, G.; Palmer, J.R.; Smith, E.N.; Chen, C.T.L.; Ambrosone, C.B.; Arnold, A.M.; Bandera, E.V.; et al. Genome-wide association study of age at menarche in African-American women. Hum. Mol. Genet. 2013, 22, 3329–3346. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Wang, N.; Guo, X.; Ziegler, J.T.; Taylor, K.D.; Xiang, A.H.; Hai, Y.; Kridel, S.J.; Nadler, J.L.; Kandeel, F.; et al. A Comprehensive Analysis of Common and Rare Variants to Identify Adiposity Loci in Hispanic Americans: The IRAS Family Study (IRASFS). PLoS ONE 2015, 10, e0134649. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.-W.; Chung, M.; Cho, S.B. Meta-analysis of genome-wide association study of homeostasis model assessment β cell function and insulin resistance in an East Asian population and the European results. Mol. Genet. Genom. 2014, 289, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Rinella, E.S.; Still, C.; Shao, Y.; Wood, G.C.; Chu, X.; Salerno, B.; Gerhard, G.S.; Ostrer, H. Genome-wide Association of Single-Nucleotide Polymorphisms with Weight Loss Outcomes After Roux-en-Y Gastric Bypass Surgery. J. Clin. Endocrinol. Metab. 2013, 98, E1131–E1136. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.R.; Brower, M.A.; Xu, N.; Cui, J.; Mengesha, E.; Chen, Y.-D.I.; Taylor, K.D.; Azziz, R.; Goodarzi, M.O. Systems Genetics Reveals the Functional Context of PCOS Loci and Identifies Genetic and Molecular Mechanisms of Disease Heterogeneity. PLoS Genet. 2015, 11, e1005455. [Google Scholar] [CrossRef]
- Sarzynski, M.A.; Jacobson, P.; Rankinen, T.; Carlsson, B.; Sjöström, L.; Bouchard, C.; Carlsson, L.M. Associations of markers in 11 obesity candidate genes with maximal weight loss and weight regain in the SOS bariatric surgery cases. Int. J. Obes. 2011, 35, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, W.-D.; Zhang, C.K.; Wang, Z.; Glessner, J.T.; Grant, S.F.A.; Zhao, H.; Hakonarson, H.; Price, R.A. A Genome-Wide Association Study on Obesity and Obesity-Related Traits. PLoS ONE 2011, 6, e18939. [Google Scholar] [CrossRef] [PubMed]
- Warrington, N.M.; Howe, L.D.; Paternoster, L.; Kaakinen, M.; Herrala, S.; Huikari, V.; Wu, Y.Y.; Kemp, J.P.; Timpson, N.J.; Pourcain, B.S.; et al. A genome-wide association study of body mass index across early life and childhood. Int. J. Epidemiol. 2015, 44, 700–712. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-J.; Liu, X.-G.; Wang, L.; Dina, C.; Yan, H.; Liu, J.-F.; Levy, S.; Papasian, C.J.; Drees, B.M.; Hamilton, J.J.; et al. Genome-wide association scans identified CTNNBL1 as a novel gene for obesity. Hum. Mol. Genet. 2008, 17, 1803–1813. [Google Scholar] [CrossRef] [PubMed]
- Scuteri, A.; Sanna, S.; Chen, W.-M.; Uda, M.; Albai, G.; Strait, J.; Najjar, S.; Nagaraja, R.; Orrú, M.; Usala, G.; et al. Genome-Wide Association Scan Shows Genetic Variants in the FTO Gene Are Associated with Obesity-Related Traits. PLoS Genet. 2007, 3, e115. [Google Scholar] [CrossRef]
- Salinas, Y.D.; Wang, L.; DeWan, A.T. Multiethnic genome-wide association study identifies ethnic-specific associations with body mass index in Hispanics and African Americans. BMC Genet. 2016, 17, 78. [Google Scholar] [CrossRef]
- Bian, L.; Traurig, M.; Hanson, R.L.; Marinelarena, A.; Kobes, S.; Muller, Y.L.; Malhotra, A.; Huang, K.; Perez, J.; Gale, A.; et al. MAP2K3 is associated with body mass index in American Indians and Caucasians and may mediate hypothalamic inflammation. Hum. Mol. Genet. 2013, 22, 4438–4449. [Google Scholar] [CrossRef]
- Wei, F.J.; Cai, C.Y.; Yu, P.; Lv, J.; Ling, C.; Shi, W.T.; Jiao, H.X.; Chang, B.C.; Yang, F.H.; Tian, Y.; et al. Quantitative candidate gene association studies of metabolic traits in Han Chinese type 2 diabetes patients. Genet. Mol. Res. 2015, 14, 15471–15481. [Google Scholar] [CrossRef]
- Dorajoo, R.; Blakemore, A.I.; Sim, X.; Ong, R.T.; Ng, D.P.; Seielstad, M.; Wong, T.Y.; Saw, S.M.; Froguel, P.; Liu, J.; et al. Replication of 13 obesity loci among Singaporean Chinese, Malay and Asian-Indian populations. Int. J. Obes. 2012, 36, 159–163. [Google Scholar] [CrossRef]
- Hong, K.W.; Oh, B. Recapitulation of genome-wide association studies on body mass index in the Korean population. Int. J. Obes. 2012, 36, 1127–1130. [Google Scholar] [CrossRef]
- Lv, D.; Zhang, D.D.; Wang, H.; Zhang, Y.; Liang, L.; Fu, J.F.; Xiong, F.; Liu, G.L.; Gong, C.X.; Luo, F.H.; et al. Genetic variations in SEC16B, MC4R, MAP2K5 and KCTD15 were associated with childhood obesity and interacted with dietary behaviors in Chinese school-age population. Gene 2015, 560, 149–155. [Google Scholar] [CrossRef]
- Mei, H.; Chen, W.; Jiang, F.; He, J.; Srinivasan, S.; Smith, E.N.; Schork, N.; Murray, S.; Berenson, G.S. Longitudinal replication studies of GWAS risk SNPs influencing body mass index over the course of childhood and adulthood. PLoS ONE 2012, 7, e31470. [Google Scholar] [CrossRef]
- Wang, H.J.; Hinney, A.; Song, J.Y.; Scherag, A.; Meng, X.R.; Grallert, H.; Illig, T.; Hebebrand, J.; Wang, Y.; Ma, J. Association of common variants identified by recent genome-wide association studies with obesity in Chinese children: A case-control study. BMC Med. Genet. 2016, 17, 7. [Google Scholar] [CrossRef]
- Wen, W.; Cho, Y.S.; Zheng, W.; Dorajoo, R.; Kato, N.; Qi, L.; Chen, C.H.; Delahanty, R.J.; Okada, Y.; Tabara, Y.; et al. Meta-analysis identifies common variants associated with body mass index in east Asians. Nat. Genet. 2012, 44, 307–311. [Google Scholar] [CrossRef]
- Hebebrand, J.; Peters, T.; Schijven, D.; Hebebrand, M.; Grasemann, C.; Winkler, T.W.; Heid, I.M.; Antel, J.; Föcker, M.; Tegeler, L.; et al. The role of genetic variation of human metabolism for BMI, mental traits and mental disorders. Mol. Metab. 2018, 12, 1–11. [Google Scholar] [CrossRef]
- Chen, G.; Bentley, A.; Adeyemo, A.; Shriner, D.; Zhou, J.; Doumatey, A.; Huang, H.; Ramos, E.; Erdos, M.; Gerry, N.; et al. Genome-wide association study identifies novel loci association with fasting insulin and insulin resistance in African Americans. Hum. Mol. Genet. 2012, 21, 4530–4536. [Google Scholar] [CrossRef] [PubMed]
- Corfitsen, H.T.; Krantz, B.; Larsen, A.; Drago, A. Molecular pathway analysis associates alterations in obesity-related genes and antipsychotic-induced weight gain. Acta Neuropsychiatr. 2020, 32, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Zhang, L.; Chen, T.; Shi, A.; Xie, K.; Li, Z.; Xu, J.; Chen, Z.; Ji, C.; Wen, J. Genetic Susceptibility to Gestational Diabetes Mellitus in a Chinese Population. Front. Endocrinol. 2020, 11, 247. [Google Scholar] [CrossRef]
- Kim, H.J.; Yoo, Y.J.; Ju, Y.S.; Lee, S.; Cho, S.I.; Sung, J.; Kim, J.I.; Seo, J.S. Combined linkage and association analyses identify a novel locus for obesity near PROX1 in Asians. Obesity 2013, 21, 2405–2412. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Guo, T.; Traurig, M.; Mason, C.C.; Kobes, S.; Perez, J.; Knowler, W.C.; Bogardus, C.; Hanson, R.L.; Baier, L.J. SIRT1 is associated with a decrease in acute insulin secretion and a sex specific increase in risk for type 2 diabetes in Pima Indians. Mol. Genet. Metab. 2011, 104, 661–665. [Google Scholar] [CrossRef]
- Jung, S.Y.; Scott, P.A.; Papp, J.C.; Sobel, E.M.; Pellegrini, M.; Yu, H.; Han, S.; Zhang, Z.F. Genome-wide Association Analysis of Proinflammatory Cytokines and Gene-lifestyle Interaction for Invasive Breast Cancer Risk: The WHI dbGaP Study. Cancer Prev. Res. 2020, 14, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Speakman, J.R. Functional Analysis of Seven Genes Linked to Body Mass Index and Adiposity by Genome-Wide Association Studies: A Review. Hum. Hered. 2013, 75, 57–79. [Google Scholar] [CrossRef]
- Vogel, C.I.G.; Greene, B.; Scherag, A.; Müller, T.D.; Friedel, S.; Grallert, H.; Heid, I.M.; Illig, T.; Wichmann, H.E.; Schäfer, H.; et al. Non-replication of an association of CTNNBL1polymorphisms and obesity in a population of Central European ancestry. BMC Med. Genet. 2009, 10, 14. [Google Scholar] [CrossRef] [PubMed]
- Hoggart, C.J.; Venturini, G.; Mangino, M.; Gomez, F.; Ascari, G.; Zhao, J.H.; Teumer, A.; Winkler, T.W.; Tšernikova, N.; Luan, J.A.; et al. Novel Approach Identifies SNPs in SLC2A10 and KCNK9 with Evidence for Parent-of-Origin Effect on Body Mass Index. PLoS Genet. 2014, 10, e1004508. [Google Scholar] [CrossRef]
- Perfilyev, A.; Dahlman, I.; Gillberg, L.; Rosqvist, F.; Iggman, D.; Volkov, P.; Nilsson, E.; Riserus, U.; Ling, C. Impact of polyunsaturated and saturated fat overfeeding on the DNA-methylation pattern in human adipose tissue: A randomized controlled trial. Am. J. Clin. Nutr. 2017, 105, 991–1000. [Google Scholar] [CrossRef]
- Guenard, F.; Tchernof, A.; Deshaies, Y.; Biron, S.; Lescelleur, O.; Biertho, L.; Marceau, S.; Perusse, L.; Vohl, M.C. Genetic regulation of differentially methylated genes in visceral adipose tissue of severely obese men discordant for the metabolic syndrome. Transl. Res. 2017, 184, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Crujeiras, A.B.; Diaz-Lagares, A.; Moreno-Navarrete, J.M.; Sandoval, J.; Hervas, D.; Gomez, A.; Ricart, W.; Casanueva, F.F.; Esteller, M.; Fernandez-Real, J.M. Genome-wide DNA methylation pattern in visceral adipose tissue differentiates insulin-resistant from insulin-sensitive obese subjects. Transl. Res. 2016, 178, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Crujeiras, A.B.; Pissios, P.; Moreno-Navarrete, J.M.; Diaz-Lagares, A.; Sandoval, J.; Gomez, A.; Ricart, W.; Esteller, M.; Casanueva, F.F.; Fernandez-Real, J.M. An Epigenetic Signature in Adipose Tissue Is Linked to Nicotinamide N-Methyltransferase Gene Expression. Mol. Nutr. Food Res. 2018, 62, e1700933. [Google Scholar] [CrossRef]
- Demerath, E.W.; Guan, W.; Grove, M.L.; Aslibekyan, S.; Mendelson, M.; Zhou, Y.H.; Hedman, A.K.; Sandling, J.K.; Li, L.A.; Irvin, M.R.; et al. Epigenome-wide association study (EWAS) of BMI, BMI change and waist circumference in African American adults identifies multiple replicated loci. Hum. Mol. Genet. 2015, 24, 4464–4479. [Google Scholar] [CrossRef]
- Ronn, T.; Volkov, P.; Gillberg, L.; Kokosar, M.; Perfilyev, A.; Jacobsen, A.L.; Jorgensen, S.W.; Brons, C.; Jansson, P.A.; Eriksson, K.F.; et al. Impact of age, BMI and HbA1c levels on the genome-wide DNA methylation and mRNA expression patterns in human adipose tissue and identification of epigenetic biomarkers in blood. Hum. Mol. Genet. 2015, 24, 3792–3813. [Google Scholar] [CrossRef]
- Anguita-Ruiz, A.; Mendez-Gutierrez, A.; Ruperez, A.I.; Leis, R.; Bueno, G.; Gil-Campos, M.; Tofe, I.; Gomez-Llorente, C.; Moreno, L.A.; Gil, A.; et al. The protein S100A4 as a novel marker of insulin resistance in prepubertal and pubertal children with obesity. Metabolism 2020, 105, 154187. [Google Scholar] [CrossRef]
- Nilsson, E.; Jansson, P.A.; Perfilyev, A.; Volkov, P.; Pedersen, M.; Svensson, M.K.; Poulsen, P.; Ribel-Madsen, R.; Pedersen, N.L.; Almgren, P.; et al. Altered DNA methylation and differential expression of genes influencing metabolism and inflammation in adipose tissue from subjects with type 2 diabetes. Diabetes 2014, 63, 2962–2976. [Google Scholar] [CrossRef]
- Ronn, T.; Volkov, P.; Davegardh, C.; Dayeh, T.; Hall, E.; Olsson, A.H.; Nilsson, E.; Tornberg, A.; Dekker Nitert, M.; Eriksson, K.F.; et al. A six months exercise intervention influences the genome-wide DNA methylation pattern in human adipose tissue. PLoS Genet. 2013, 9, e1003572. [Google Scholar] [CrossRef]
- Multhaup, M.L.; Seldin, M.M.; Jaffe, A.E.; Lei, X.; Kirchner, H.; Mondal, P.; Li, Y.; Rodriguez, V.; Drong, A.; Hussain, M.; et al. Mouse-human experimental epigenetic analysis unmasks dietary targets and genetic liability for diabetic phenotypes. Cell Metab. 2015, 21, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Bollepalli, S.; Kaye, S.; Heinonen, S.; Kaprio, J.; Rissanen, A.; Virtanen, K.A.; Pietilainen, K.H.; Ollikainen, M. Subcutaneous adipose tissue gene expression and DNA methylation respond to both short- and long-term weight loss. Int. J. Obes. 2018, 42, 412–423. [Google Scholar] [CrossRef]
- Arner, P.; Sahlqvist, A.S.; Sinha, I.; Xu, H.; Yao, X.; Waterworth, D.; Rajpal, D.; Loomis, A.K.; Freudenberg, J.M.; Johnson, T.; et al. The epigenetic signature of systemic insulin resistance in obese women. Diabetologia 2016, 59, 2393–2405. [Google Scholar] [CrossRef]
- Keller, M.; Hopp, L.; Liu, X.; Wohland, T.; Rohde, K.; Cancello, R.; Klos, M.; Bacos, K.; Kern, M.; Eichelmann, F.; et al. Genome-wide DNA promoter methylation and transcriptome analysis in human adipose tissue unravels novel candidate genes for obesity. Mol. Metab. 2017, 6, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Andersen, E.; Ingerslev, L.R.; Fabre, O.; Donkin, I.; Altintas, A.; Versteyhe, S.; Bisgaard, T.; Kristiansen, V.B.; Simar, D.; Barres, R. Preadipocytes from obese humans with type 2 diabetes are epigenetically reprogrammed at genes controlling adipose tissue function. Int. J. Obes. 2019, 43, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Kamei, Y.; Suganami, T.; Ehara, T.; Kanai, S.; Hayashi, K.; Yamamoto, Y.; Miura, S.; Ezaki, O.; Okano, M.; Ogawa, Y. Increased expression of DNA methyltransferase 3a in obese adipose tissue: Studies with transgenic mice. Obesity 2010, 18, 314–321. [Google Scholar] [CrossRef]
- Gillberg, L.; Perfilyev, A.; Brøns, C.; Thomasen, M.; Grunnet, L.G.; Volkov, P.; Rosqvist, F.; Iggman, D.; Dahlman, I.; Risérus, U.; et al. Adipose tissue transcriptomics and epigenomics in low birthweight men and controls: Role of high-fat overfeeding. Diabetologia 2016, 59, 799–812. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gu, H.F.; Frystyk, J.; Efendic, S.; Brismar, K.; Thorell, A. Analyses of IGFBP2 DNA methylation and mRNA expression in visceral and subcutaneous adipose tissues of obese subjects. Growth Horm. IGF Res. 2019, 45, 31–36. [Google Scholar] [CrossRef]
- Petrus, P.; Bialesova, L.; Checa, A.; Kerr, A.; Naz, S.; Backdahl, J.; Gracia, A.; Toft, S.; Dahlman-Wright, K.; Heden, P.; et al. Adipocyte Expression of SLC19A1 Links DNA Hypermethylation to Adipose Tissue Inflammation and Insulin Resistance. J. Clin. Endocrinol. Metab. 2018, 103, 710–721. [Google Scholar] [CrossRef]
- Ryden, M.; Petrus, P.; Andersson, D.P.; Medina-Gomez, G.; Escasany, E.; Corrales Cordon, P.; Dahlman, I.; Kulyte, A.; Arner, P. Insulin action is severely impaired in adipocytes of apparently healthy overweight and obese subjects. J. Intern. Med. 2019, 285, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Dayeh, T.; Tuomi, T.; Almgren, P.; Perfilyev, A.; Jansson, P.-A.; de Mello, V.D.; Pihlajamäki, J.; Vaag, A.; Groop, L.; Nilsson, E.; et al. DNA methylation of loci within ABCG1 and PHOSPHO1 in blood DNA is associated with future type 2 diabetes risk. Epigenetics 2016, 11, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.-C.; Glastonbury, C.A.; Eliot, M.N.; Bollepalli, S.; Yet, I.; Castillo-Fernandez, J.E.; Carnero-Montoro, E.; Hardiman, T.; Martin, T.C.; Vickers, A.; et al. Smoking induces coordinated DNA methylation and gene expression changes in adipose tissue with consequences for metabolic health. Clin. Epigenet. 2018, 10, 126. [Google Scholar] [CrossRef]
- Chu, S.H.; Kelsey, K.T.; Koestler, D.C.; Loucks, E.B.; Huang, Y.T. Leveraging cell-specific differentially methylated regions to identify leukocyte infiltration in adipose tissue. Genet. Epidemiol. 2019, 43, 1018–1029. [Google Scholar] [CrossRef]
- De Toro-Martín, J.; Guénard, F.; Tchernof, A.; Deshaies, Y.; Pérusse, L.; Biron, S.; Lescelleur, O.; Biertho, L.; Marceau, S.; Vohl, M.C. A CpG-SNP Located within the ARPC3 Gene Promoter Is Associated with Hypertriglyceridemia in Severely Obese Patients. Ann. Nutr. Metab. 2016, 68, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Bluher, M. Adipose tissue inflammation: A cause or consequence of obesity-related insulin resistance? Clin. Sci. 2016, 130, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Shimobayashi, M.; Albert, V.; Woelnerhanssen, B.; Frei, I.C.; Weissenberger, D.; Meyer-Gerspach, A.C.; Clement, N.; Moes, S.; Colombi, M.; Meier, J.A.; et al. Insulin resistance causes inflammation in adipose tissue. J. Clin. Investig. 2018, 128, 1538–1550. [Google Scholar] [CrossRef]
- Lagathu, C.; Christodoulides, C.; Tan, C.Y.; Virtue, S.; Laudes, M.; Campbell, M.; Ishikawa, K.; Ortega, F.; Tinahones, F.J.; Fernández-Real, J.M.; et al. Secreted frizzled-related protein 1 regulates adipose tissue expansion and is dysregulated in severe obesity. Int. J. Obes. 2010, 34, 1695–1705. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ha, X.; Li, W.; Xu, P.; Zhang, Z.; Wang, T.; Li, J.; Wang, Y.; Li, S.; Xie, J.; et al. Comparative gene expression profile and DNA methylation status in diabetic patients of Kazak and Han people. Medicine 2018, 97, e11982. [Google Scholar] [CrossRef]
- Schadt, E.E.; Monks, S.A.; Drake, T.A.; Lusis, A.J.; Che, N.; Colinayo, V.; Ruff, T.G.; Milligan, S.B.; Lamb, J.R.; Cavet, G.; et al. Genetics of gene expression surveyed in maize, mouse and man. Nature 2003, 422, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Bolton, J.; Montastier, E.; Carayol, J.; Bonnel, S.; Mir, L.; Marques, M.A.; Astrup, A.; Saris, W.; Iacovoni, J.; Villa-Vialaneix, N.; et al. Molecular Biomarkers for Weight Control in Obese Individuals Subjected to a Multiphase Dietary Intervention. J. Clin. Endocrinol. Metab. 2017, 102, 2751–2761. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Hao, K.; Ji, H.; Peng, S.; Sherwood, B.; Di Narzo, A.; Tsai, H.-J.; Liu, X.; Burd, I.; Wang, G.; et al. Genome-wide approach identifies a novel gene-maternal pre-pregnancy BMI interaction on preterm birth. Nat. Commun. 2017, 8, 15608. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Aron-Wisnewsky, J.; Marcelin, G.; Genser, L.; Le Naour, G.; Torcivia, A.; Bauvois, B.; Bouchet, S.; Pelloux, V.; Sasso, M.; et al. Accumulation and Changes in Composition of Collagens in Subcutaneous Adipose Tissue After Bariatric Surgery. J. Clin. Endocrinol. Metab. 2016, 101, 293–304. [Google Scholar] [CrossRef]
- Pettersson, M.; Viljakainen, H.; Loid, P.; Mustila, T.; Pekkinen, M.; Armenio, M.; Andersson-Assarsson, J.C.; Mäkitie, O.; Lindstrand, A. Copy Number Variants Are Enriched in Individuals with Early-Onset Obesity and Highlight Novel Pathogenic Pathways. J. Clin. Endocrinol. Metab. 2017, 102, 3029–3039. [Google Scholar] [CrossRef]
- Pellegrinelli, V.; Heuvingh, J.; du Roure, O.; Rouault, C.; Devulder, A.; Klein, C.; Lacasa, M.; Clément, E.; Lacasa, D.; Clément, K. Human adipocyte function is impacted by mechanical cues. J. Pathol. 2014, 233, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, M.; Inoue, M.; Jiang, Y.; Barnes, R.H., II; Buchner, D.A.; Chun, T.-H. Fat depot-specific gene signature and ECM remodeling of Sca1(high) adipose-derived stem cells. Matrix Biol. 2014, 36, 28–38. [Google Scholar] [CrossRef]
- Li, X.; Thomason, P.A.; Withers, D.J.; Scott, J. Bio-informatics analysis of a gene co-expression module in adipose tissue containing the diet-responsive gene Nnat. BMC Syst. Biol. 2010, 4, 175. [Google Scholar] [CrossRef]
- Aguilera, C.M.; Gomez-Llorente, C.; Tofe, I.; Gil-Campos, M.; Cañete, R.; Gil, Á. Genome-wide expression in visceral adipose tissue from obese prepubertal children. Int. J. Mol. Sci. 2015, 16, 7723–7737. [Google Scholar] [CrossRef]
- Kępczyńska, M.A.; Zaibi, M.S.; Alomar, S.Y.; Trayhurn, P. PCR arrays indicate that the expression of extracellular matrix and cell adhesion genes in human adipocytes is regulated by IL-1β (interleukin-1β). Arch. Physiol. Biochem. 2017, 123, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Kogelman, L.J.; Fu, J.; Franke, L.; Greve, J.W.; Hofker, M.; Rensen, S.S.; Kadarmideen, H.N. Inter-Tissue Gene Co-Expression Networks between Metabolically Healthy and Unhealthy Obese Individuals. PLoS ONE 2016, 11, e0167519. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, M.; De Sibio, M.T.; Mathias, L.S.; Rodrigues, B.M.; Sakalem, M.E.; Nogueira, C.R. Irisin modulates genes associated with severe coronavirus disease (COVID-19) outcome in human subcutaneous adipocytes cell culture. Mol. Cell. Endocrinol. 2020, 515, 110917. [Google Scholar] [CrossRef]
- Samuvel, D.J.; Sundararaj, K.P.; Li, Y.; Lopes-Virella, M.F.; Huang, Y. Adipocyte-mononuclear cell interaction, Toll-like receptor 4 activation, and high glucose synergistically up-regulate osteopontin expression via an interleukin 6-mediated mechanism. J. Biol. Chem. 2010, 285, 3916–3927. [Google Scholar] [CrossRef] [PubMed]
- Wu, C. Focal adhesion: A focal point in current cell biology and molecular medicine. Cell Adh. Migr. 2007, 1, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Luk, C.T.; Shi, S.Y.; Cai, E.P.; Sivasubramaniyam, T.; Krishnamurthy, M.; Brunt, J.J.; Schroer, S.A.; Winer, D.A.; Woo, M. FAK signalling controls insulin sensitivity through regulation of adipocyte survival. Nat. Commun. 2017, 8, 14360. [Google Scholar] [CrossRef]
- Liu, Y.; Ji, Y.; Li, M.; Wang, M.; Yi, X.; Yin, C.; Wang, S.; Zhang, M.; Zhao, Z.; Xiao, Y. Integrated analysis of long noncoding RNA and mRNA expression profile in children with obesity by microarray analysis. Sci. Rep. 2018, 8, 8750. [Google Scholar] [CrossRef] [PubMed]
- Tam, C.S.; Covington, J.D.; Bajpeyi, S.; Tchoukalova, Y.; Burk, D.; Johannsen, D.L.; Zingaretti, C.M.; Cinti, S.; Ravussin, E. Weight Gain Reveals Dramatic Increases in Skeletal Muscle Extracellular Matrix Remodeling. J. Clin. Endocrinol. Metab. 2014, 99, 1749–1757. [Google Scholar] [CrossRef]
- Tam, C.S.; Chaudhuri, R.; Hutchison, A.T.; Samocha-Bonet, D.; Heilbronn, L.K. Skeletal muscle extracellular matrix remodeling after short-term overfeeding in healthy humans. Metabolism 2017, 67, 26–30. [Google Scholar] [CrossRef]
- Walton, R.G.; Kosmac, K.; Mula, J.; Fry, C.S.; Peck, B.D.; Groshong, J.S.; Finlin, B.S.; Zhu, B.; Kern, P.A.; Peterson, C.A. Human skeletal muscle macrophages increase following cycle training and are associated with adaptations that may facilitate growth. Sci. Rep. 2019, 9, 969. [Google Scholar] [CrossRef]
- Aleksandrova, K.; Egea Rodrigues, C.; Floegel, A.; Ahrens, W. Omics Biomarkers in Obesity: Novel Etiological Insights and Targets for Precision Prevention. Curr. Obes. Rep. 2020, 9, 219–230. [Google Scholar] [CrossRef] [PubMed]


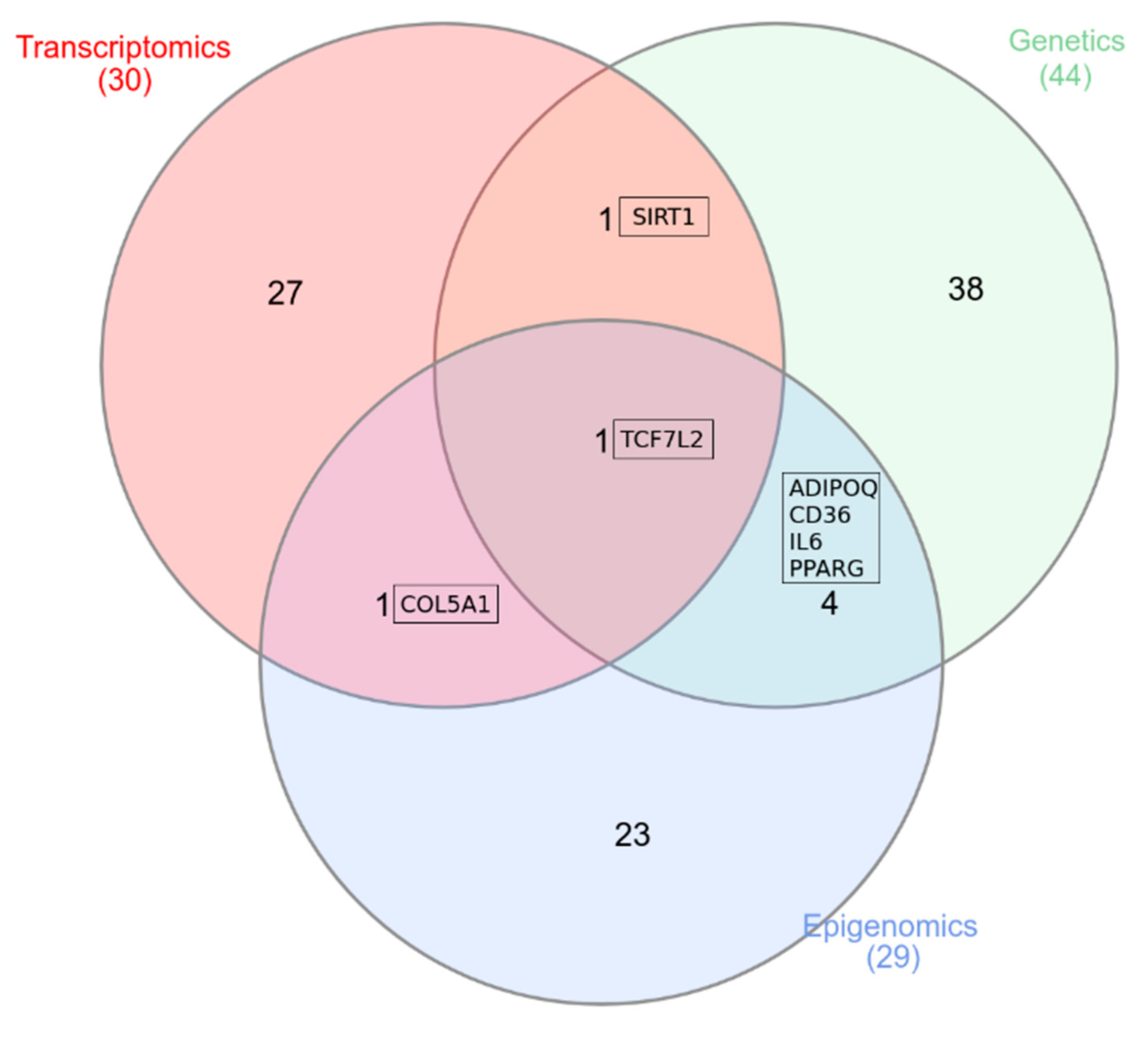{kind=link}
| Cell Signaling/ Term | Gene Symbol and Name | Population | Study/Technology | Author |
|---|---|---|---|---|
| Collagens | ||||
| Collagen-containing extracellular matrix | ADIPOQ (Adiponectin) | 1341 Framingham Heart Study participants in 310 families | GWAS on adiposity | Fox et al. 2007 [62] |
| Collagen-containing extracellular matrix | ADIPOQ (Adiponectin) | 1156 Mexican mestizos with obesity and 473 normal weight controls | Candidate gene approach on obesity | León-Mimila et al. 2013 [63] |
| Collagen-containing extracellular matrix | APOE (Apolipoprotein E) | GIANT cohort (BMI as measure of obesity, n = 123,865) | Meta-analysis of GWAS on obesity | Hinney et al. 2014 [64] |
| Collagen-containing extracellular matrix | APOE (Apolipoprotein E) | 132 European patients with severe HTG and 351 controls | GWAS on lipid traits | Wang et al. 2008 [65] |
| Collagen-containing extracellular matrix | CBLN4 (Cerebellin-4) | 1110 and morbid obesity bariatric patients from Taiwan | GWAS on obesity | Chiang et al. 2019 [66] |
| Collagen-containing extracellular matrix | COL4A1 (Collagen alpha-1(IV) chain) | 815 Hispanic children | Candidate gene approach on obesity | Comuzzie et al. 2012 [67] |
| Collagen-containing extracellular matrix | COL6A5 (Collagen alpha-5(VI) chain) | 5049 Europeans participants | GWAS on adiposity | Namjou et al. 2013 [68] |
| Collagen-containing extracellular matrix | CTSS | 21,969 individuals from diverse ethnic populations | Meta-analysis of GWAS on obesity | Pei et al. 2014 [69] |
| (Cathepsin S) | ||||
| Collagen-containing extracellular matrix | F12 | 242 monozygotic and 140 dizygotic twin pairs from a Northern Han Chinese population | GWAS on adiposity | Wu et al. 2018 [70] |
| (Coagulation factor XII) | ||||
| Collagen-containing extracellular matrix | F13A1 (Coagulation factor XIII A chain) | 13 pairs of monozygotic Chinese twin pairs discordant for BMI and 77 controls | GWAS on adiposity | Naukkarinen et al. 2010 [71] |
| Collagen-containing extracellular matrix | FRAS1 (Extracellular matrix protein FRAS1) | 1060 obese cases and never-overweight controls | GWAS on uric acid levels | Li et al. 2013 [72] |
| Collagen-containing extracellular matrix | ITIH4 | 390 Mexican children with obesity and 405 normal weight controls | Candidate gene approach on obesity | Liu et al. 2019 [73] |
| (Inter-alpha-trypsin inhibitor heavy chain H4) | ||||
| Dystroglycan complex | SGCZ | 1335 African American subjects from GENOA cohort and 1224 from HyperGEN cohort | CNVs-GWAS on adiposity | Zhao et al. 2012 [74] |
| (Zeta-sarcoglycan) | ||||
| Dystrophin associated glycoprotein complex | SSPN | 27,350 African ancestry individuals | GWAS on adiposity | Liu et al. 2013 [75] |
| (Sarcospan) | ||||
| Cell adhesion | ||||
| Cell adhesion molecule binding | ADAM23 (Disintegrin and metalloproteinase domain-containing protein 23) | 35,668 children from 20 studies in the discovery phase and 11,873 children from 13 studies in the replication phase | Meta-analysis of GWAS on adiposity | Felix et al. 2015 [76] |
| Cell matrix adhesion | ADAMTS9 (A disintegrin and metalloproteinase with thrombospondin motifs 9) | 5169 T2D cases and 4560 normal glycemic controls | GWAS on metabolic syndrome | Kong et al. 2015 [77] |
| Cell matrix adhesion | ADAMTS9 (A disintegrin and metalloproteinase with thrombospondin motifs 9) | 33,591 and 27,350 African ancestry individuals with waist circumference or waist-hip ratio | GWAS on adiposity | Liu et al. 2013 [75] |
| Cell matrix adhesion | BCL6 (B-cell lymphoma 6 protein) | 1028 unrelated European-American extremely obese females and normal weight controls | GWAS on lipid traits | Jiao et al. 2015 [78] |
| Regulation of cell adhesion | C2CD4A (C2 calcium-dependent domain-containing protein 4A) | 10,701 non-diabetic adults of European ancestry under follow-up | GWAS on T2D | Strawbridge et al. 2011 [79] |
| Cell matrix adhesion | CD36 (Platelet glycoprotein 4) | 9973 European subjects | Meta-analysis of GWAS on obesity | Choquet et al. 2011 [80] |
| Cell adhesion molecule binding | CDH12 (Cadherin-12) | 1715 African American subjects | GWAS on adiposity | Ng et al. 2012 [81] |
| Cell adhesion molecule binding | CDH18 (Cadherin-18) | 1996 adult survivors of childhood cancer | GWAS on obesity | Wilson et al. 2015 [82] |
| Regulation of cell adhesion | CELSR2 (Cadherin EGF LAG seven-pass G-type receptor 2) | 40,473 African American, American Indian, Asian/Pacific Islander, European American, and Hispanic participants from 7 studies | Candidate gene approach on CRP levels | Kocarnik et al. 2014 [83] |
| Cell adhesion molecule binding | COBLL1 (Cordon-bleu protein-like 1 | 1011 mediterranean subjects | GWAS on leptin levels | Ortega-Azorin et al. 2019 [84] |
| Regulation of cell adhesion | CSK (Tyrosine-protein kinase CSK) | 1279 Japanese subjects (556 men and 723 women) | Candidate gene approach on adiposity | Hotta et al. 2012 [85] |
| Cell adhesion involved in heart morphogenesis | FLRT2 (Leucine-rich repeat transmembrane protein FLRT2) | 8089 African American women | GWAS and age of menarche | Demerath et al. 2013 [86] |
| Cell adhesion molecule binding | IDH1 (Isocitrate dehydrogenase [NADP] cytoplasmic) | 1263 Hispanic Americans | GWAS on adiposity | Gao et al. 2015 [87] |
| Cell adhesion molecule binding | IGF1 (Insulin-like growth factor I) | 5974 non-diabetic subjects | Meta-analysis of GWAS on IR | Hong et al. 2014 [88] |
| Focal adhesion | ||||
| Focal adhesion | IGF1R (Insulin like growth factor 1 receptor) | Obese patients after bariatric surgery | GWAS on adiposity | Rinella et al. 2013 [89] |
| Adherents junction | INSR (Insulin receptor) | 23 obese PCOS women and 13 control women | GWAS on PCOS and obesity | Jones et al. 2015 [90] |
| Regulation of cell adhesion | LEP (Leptin) | 1011 mediterranean subjects | GWAS on leptin levels | Ortega-Azorín et al. 2019 [84] |
| Regulation of cell adhesion | LEP (Leptin) | Swedish obese subjects after bariatric surgery | Candidate gene approach on adiposity | Sarzynski et al. 2011 [91] |
| Cell adhesion molecule binding | NRXN3 (Neurexin-3) | 520 cases (BMI > 35 kg/m(2)) and 540 control subjects (BMI < 25 kg/m(2)) of European ancestry | GWAS on obesity and related traits | Wang et al. 2011 [92] |
| Cell adhesion molecule binding | OLFM4 (Olfactomedin-4) | 35,668 children from 20 studies in the discovery phase and 11,873 children from 13 studies in the replication phase. | Meta-analysis of GWAS on adiposity | Felix et al. 2015 [76] |
| Cell adhesion molecule binding | OLFM4 (Olfactomedin-4) | 9377 children from the from the ALSPAC and the Raine Study | Meta-analysis of GWAS on adiposity | Warrington et al. 2015 [93] |
| Cell adhesion molecule binding | PFKP (ATP-dependent 6-phosphofructokinase, platelet type) | 1000 US Caucasians | GWAS on obesity | Liu et al. 2008 [94] |
| Cell adhesion molecule binding | PFKP (ATP-dependent 6-phosphofructokinase, platelet type) | European Americans (n = 1496) and Hispanic Americans (n = 839) | GWAS on obesity | Scuteri et al. 2007 [95] |
| Regulation of cell adhesion | RREB1 (Ras-responsive element-binding protein 1) | 1060 obese cases and never-overweight controls | GWAS on uric acid levels | Li et al. 2013 [72] |
| Adherents junction | TCF7L2 (transcription factor 7 like 2) | 1235 Hispanic, 706 Asian, 1549 African American, and 2395 European American subjects from the Multi-ethnic Study of Atherosclerosis | GWAS on BMI | Salinas et al. 2016 [96] |
| Adherents junction | TCF7L2 (transcription factor 7 like 2) | 10,701 non-diabetic adults of European ancestry under follow-up | GWAS on T2D | Strawbridge et al. 2011 [79] |
| Positive regulation of endothelial cell migration | MAP2K3 (Dual specificity mitogen-activated protein kinase kinase 3) | 3562 American Indians | Candidate gene approach on adiposity | Bian et al. 2013 [97] |
| Positive regulation of endothelial cell migration | MAP2K3 (Dual specificity mitogen-activated protein kinase kinase 3) | 1975 Han Chinese type 2 diabetes patients | Candidate gene approach on Metabolic traits | Wei et al. 2015 [98] |
| Endothelial Cell migration | ||||
| Negative regulation of endothelial cell migration | MAP2K5 (Dual specificity mitogen-activated protein kinase kinase 5) | Singaporean Chinese, Malay, and Asian-Indian populations (n = 10,482) | Meta-analysis of GWAS on adiposity | Dorajoo et al. 2012 [99] |
| Negative regulation of endothelial cell migration | MAP2K5 (Dual specificity mitogen-activated protein kinase kinase 5) | 8842 individuals from the Korean Association Resource data | Candidate gene approach on adiposity | Hong et al. 2012 [100] |
| Negative regulation of endothelial cell migration | MAP2K5 (Dual specificity mitogen-activated protein kinase kinase 5) | Chinese children (n = 2977, 853 obese and 2124 controls | Candidate gene approach on adiposity | Lv et al. 2015 [101] |
| Negative regulation of endothelial cell migration | MAP2K5 (Dual specificity mitogen-activated protein kinase kinase 5) | Childhood (3–17 years) and adulthood (18–45 years) follow-up for 658 subjects | Candidate gene approach on adiposity | Mei et al. 2012 [102] |
| Negative regulation of endothelial cell migration | MAP2K5 (Dual specificity mitogen-activated protein kinase kinase 5) | 2 030 unrelated Chinese children, including 607 normal weight, 718 overweight, and 705 obese | Candidate gene approach on adiposity | Wang et al. 2016 [103] |
| Negative regulation of endothelial cell migration | MAP2K5 (Dual specificity mitogen-activated protein kinase kinase 5) | 27,715; 37,691 and 17,642 individuals from three east Asian populations | Meta-analysis of GWAS on adiposity | Wen et al. 2012 [104] |
| Positive regulation of endothelial cell migration | MET (Hepatocyte growth factor receptor) | Search in GWAS catalog | GWAS on BMI and mental traits disorders | Hebebrand et al. 2018 [105] |
| Negative regulation of endothelial cell migration | PPARG (Peroxisome proliferator-activated receptor gamma) | 927 non-diabetic African Americans | GWAS on IR | Chen et al. 2012 [106] |
| Negative regulation of endothelial cell migration | PPARG (Peroxisome proliferator-activated receptor gamma) | 765 (556 males) from the NIMH CATIE sample | GWAS on drug-induced weight-regain | Corfitsen et al. 2020 [107] |
| Negative regulation of endothelial cell migration | PPARG (Peroxisome proliferator-activated receptor gamma) | 1156 Mexican mestizos with obesity and 473 normal weight controls | Candidate gene approach on obesity | León Mimila et al. 2013 [63] |
| Negative regulation of endothelial cell migration | PPARG (Peroxisome proliferator-activated receptor gamma) | Review Swedish population | Candidate gene approach on obesity | Sarzynski et al. 2011 [91] |
| Positive regulation of endothelial cell migration | PROX1 (Prospero homeobox protein 1) | 964 chinese pregnant women with GMD and 1021 chinese pregnant women with normal glucose tolerance | GWAS on GMD | Cao et al. 2020 [108] |
| Positive regulation of endothelial cell migration | PROX1 (Prospero homeobox protein 1) | 756 individuals from a Mongolian sample | GWAS on obesity | Kim et al. 2013 [109] |
| Positive regulation of endothelial cell migration | SIRT1 (Sirtuin 1) | 3501 Pima Indians | Candidate gene approach on IR and T2D | Dong et al. 2011 [110] |
| ECM assembly and disassembly | ||||
| Extracellular matrix structural constituent | GP2 (Pancreatic secretory granule membrane major glycoprotein GP2) | 27,715; 37,691 and 17,642 individuals from three east Asian populations | Meta-analysis of GWAS on adiposity | Wen et al. 2012 [104] |
| Extracellular matrix disassembly | IL6 (Interleukin-6) | 16,088 postmenopausal women stratified by obesity status | GWAS on Proinflammatory Cytokines | Jung et al. 2020 [111] |
| Extracellular matrix | LINGO2 (Leucine-rich repeat and immunoglobulin-like domain-containing nogo receptor-interacting protein 2) | 100,418 adults from the single large multi-ethnic Genetic Epidemiology Research on Adult Health and Aging (GERA) cohort | Review of GWAS on adiposity | Speakman et al. 2013 [112] |
| Extracellular matrix disassembly | SH3PXD2B (SH3 and PX domain-containing protein 2B) | German case control population of 487 extremely obese children and adolescents and 442 healthy lean individuals; and an adult population of 1644 individuals from the German population-based study (KORA) | Candidate gene approach on obesity | Vogel et al. 2009 [113] |
| Extracellular matrix organization | SLC2A10 (Solute carrier family 2, facilitated glucose transporter member 10) | 56,000 unrelated individuals of several ethnics and cohorts | GWAS on adiposity | Hoggart et al. 2014 [114] |
| Cell Signaling/Term | Gene Symbol and Name | Study/Tissue/Technology | Author, In Vitro/In Vivo |
|---|---|---|---|
| Collagens | |||
| Collagen-containing extracellular matrix | ADIPOQ (Adiponectin) | Epigenome of human AT is affected by dietary fat composition and overfeeding in RCT/SAT/Array EWAS Illumina | Perfilyev et al. 2017, [115] In vivo |
| Collagen-containing extracellular matrix | COL11A2 (Collagen alpha-2(XI) chain) | Severely obese men discordant for Metabolic syndrome/VAT/Infinium Human Methylation 450K Beadchip (Illumina) | Guénard et al. 2017, [116] In vivo |
| Collagen-containing extracellular matrix | COL11A2 (Collagen alpha-2(XI) chain) | DNA methylation pattern in VAT differentiates insulin-resistant from insulin-sensitive obese subjects/Array EWAS Infinium Human Methylation 450K BeadChips (Illumina) | Crujeiras et al. 2016, [117] In vivo |
| Collagen-containing extracellular matrix | COL23A1 (Collagen alpha-1(XXIII) chain) | An epigenetic signature in association with nicotinamide N-methyltransferase gene expression VAT/Infinium Human Methylation 450K BeadChip array (Illumina) | Crujeiras et al. 2018, [118] In vivo |
| Collagen-containing extracellular matrix | LGALS3BP (Galectin-3-binding protein) | Methylation signatures associated with obesity traits using leukocyte DNA samples from 2097 African American adults/SAT/EWAS Infinium Human Methylation 450K Beadchip (Illumina) | Demerath et al. 2015, [119] In vivo |
| Collagen-containing extracellular matrix | PLG (Plasminogen) | Age, BMI, and HbA1c levels on DNA methylation and mRNA expression patterns in human AT biomarkers in blood/AT/Array EWAS Infinium Human Methylation 450K BeadChips (Illumina) | Rönn et al. 2015, [120] In vivo |
| Collagen-containing extracellular matrix | S100A4 (Protein S100-A4) | Evaluate the association between S100A4 and different obesity and insulin resistance parameters/vAT/Array AffymetrixHG-V133Plus 2.0 and RT-qPCR | Anguita-Ruiz et al. 2020, [121] In vivo |
| Cell adhesion | |||
| Adherents junction | TCF7L2 (transcription factor 7 like 2) | DNA methylation data in AT from monozygotic twin pairs discordant for T2D and independent case control cohorts/AT/Array EWAS Infinium Human Methylation 450K BeadChips (Illumina). | Nilsson et al. 2014, [122] In vivo |
| Adherents junction | TCF7L2 (transcription factor 7 like 2) | Six months exercise intervention/AT/Array EWAS Infinium Human Methylation 450K BeadChips (Illumina). | Rönn et al. 2013, [123] In vivo |
| Adherents junction | TCF7L2 (transcription factor 7 like 2) | Differentially DNA-methylated genomic regions in mouse and then replicated in human/AT/Array Comprehensive High-throughput Array-based Relative Methylation (CHARM) 7.5 million CpG sites | Multhaup et al. 2015, [124] In vitro e In vivo |
| Cell adhesion molecule binding | SPP1 (Osteopontin) | DNA methylation data in AT from monozygotic twin pairs discordant for T2D and independent case control cohorts/AT/Array EWAS Infinium Human Methylation 450K BeadChips (Illumina) | Nilsson et al. 2014, [122] In vivo |
| Cell adhesion molecule binding | SPP1 (Osteopontin) | Age, BMI, and HbA1c levels on DNA methylation and mRNA expression patterns in human AT biomarkers in blood/AT/Array EWAS Infinium Human Methylation 450K BeadChips (Illumina) | Rönn et al. 2015, [120] In vivo |
| Cell adhesion molecule binding | BAG3 (BAG family molecular chaperone regulator 3) | Gene expression and DNA methylation respond to both short- and long-term weight loss/SAT/Infinium HumanMethylation 450K BeadChip | Bollepalli et al. 2018, [125] In vivo |
| Cell adhesion molecule binding | COL5A1 (Collagen alpha-1(V) chain) | The epigenetic signature of systemic insulin resistance in obese women/SAT and VAT/Array EWAS Infinium Human Methylation 450K BeadChips (Illumina). | Arner et al. 2016, [126] In vivo |
| Cell adhesion molecule binding | ADAM2 (Disintegrin and metalloproteinase domain-containing protein 2) | Genome-wide DNA methylation pattern differentiates insulin-resistant from insulin-sensitive obese subjects/VAT/Array EWAS Infinium Human Methylation 450K BeadChips (Illumina). | Crujeiras et al. 2016, [117] |
| Cell matrix adhesion | CD36 (Platelet glycoprotein 4) | Case control study in 2 independent cohorts of obese/non-obese individuals/SAT and VAT/EZ DNA Methylation kit (Zymo) | Keller et al. 2017, [127] In vivo/In silico |
| Cell matrix adhesion | CD36 (Platelet glycoprotein 4) | Case control study in obese subjects (Obese+Obese T2D) to evaluate if extracellular factors in obesity epigenetically reprogram adipogenesis potential and metabolic function of preadipocytes/VAT/Array EWAS RRBS | Andersen et al. 2019, [128] In vivo |
| Cell matrix adhesion | SFRP1 (Secreted frizzled-related protein 1) | Role of a DNA methyltransferase (Dnmt3a) in obese AT from transgenic mice overexpressing Dnmt3a/AT/Array GWAS Affimetrics Mouse Genome 430 2.0 | Kamei et al. 2010, [129] In vivo/In vitro |
| Cell matrix adhesion | CDK5 (Cyclin-dependent-like kinase 5) | Case control study/SAT/Affymetrix Human Gene 1.0 ST arrays and DNA methylation using Illumina 450K BeadChip arrays. | Gillberg et al. 2016, [130] In vivo |
| Cell matrix adhesion | CD44 (CD44 antigen) | Genome-wide DNA methylation pattern in insulin-resistant from insulin-sensitive obese subjects/VAT/Array EWAS Infinium Human Methylation 450K BeadChips (Illumina). | Crujeiras et al. 2016, [117] In vivo |
| Regulation of cell adhesion | HLA-DPB1 (HLA class II histocompatibility antigen, DP beta 1 chain) | Age, BMI and HbA1c levels on DNA methylation and mRNA expression patterns in human AT biomarkers in blood/AT/Array EWAS Infinium Human Methylation 450K BeadChips (Illumina) | Rönn et al. 2015, [120] In vivo |
| Regulation of cell adhesion | MAPK7 (Mitogen-activated protein kinase 7) | A RCT to study if the epigenome of human AT is affected differently by dietary fat composition/SAT/Array (Infinium Human Methylation 450K BeadChip, Illumina) | Perfilyev et al. 2017, [115] In vivo |
| Regulation of cell adhesion | IGFBP2 (Insulin-like growth factor-binding protein 2) | Epigenetic changes of the IGFBP2 gene associated with obesity by DNA methylation and mRNA expression in adipocytes from different depots/SAT and VAT/Array (Bisulfite pyrosequencing, PyroMark Q96) | Zhang et al. 2019, [131] In vivo |
| Regulation of cell adhesion | CCL2 (C-C motif chemokine 2) | Adipocyte-expressed 1CC genes linked to WAT inflammation and IR from obese individuals/SAT, isolated adipocytes and in vitro adipocytes/Array (GeneChip® Human Transcriptome Array 2.0 (Affymetrix) and EZ DNA Methylation-Gold Kit | Petrus et al. 2018, [132] In vivo/In vitro |
| Tight junction | CLDN1 (claudin 1) | Genome-wide DNA promoter methylation along with mRNA profiles in non-obese vs. obese individuals/SAT and VAT/Array (Infinium HumanMethylation450K BeadChips (Illumina) | Keller et al. 2017, [127] In vivo |
| Focal adhesion | |||
| Focal adhesion | AKT2 (AKT serine/threonine kinase 2) | Case control study in subjects subdivided according BMI aimed to identify AT dysfunction involved in decreasing insulin action in adipocytes/SAT/Array EWAS (Illumina HumanMethylation27 BeadChip) + TWAS (Affymetrix Human 1.0 or 1.1 ST arrays) + pyrosequencing + RT-PCR | Rydén et al. 2019, [133] In vivo |
| ECM assembly and disassembly | |||
| Extracellular matrix | PHOSPH+B33O1 (Phosphoethanolamine/phosphocholine phosphatase) | DNA methylation loci in blood DNA (ABCG1, PHOSPHO1, SOCS3, SREBF1, and TXNIP) as predictors of future T2D/AT, blood, human pancreatic islets, liver, and skeletal muscle/Array EWAS Illumina Infinium Human Methylation 450K Beadchip | Dayeh et al. 2016, [134] In vivo |
| Extracellular matrix assembly | NOTCH1 (Neurogenic locus notch homolog protein 1) | Smoking-associated DNA methylation and gene expression variation in AT biopsies from 542 healthy female twins/SAT, blood/Array EWAS Illumina Infinium Human Methylation 450K Beadchip | Tsai et al. 2018, [135] In vivo |
| Extracellular matrix structural constituent | CD4 (T-cell surface glycoprotein CD4) | DNA methylation data to identify leukocyte cell types in obesity/SAT, blood/Array EWAS Illumina Infinium HumanMethylation450K Beadchip | Chu et al. 2019, [136] In vivo |
| Extracellular matrix structural constituent | CD4 (T-cell surface glycoprotein CD4) | Methylation signatures associated with obesity traits using leukocyte DNA samples from 2097 African American adults/SAT/EWAS Infinium Human Methylation 450K Beadchip (Illumina) | Demerath et al. 2015, [119] In vivo |
| Extracellular matrix disassembly | IL6 (Interleukin-6) | Epigenome of human AT affected differently by dietary fat composition and general overfeeding in a randomized trial/SAT/Array EWAS Illumina Infinium Human Methylation 450K Beadchip | Perfilyev et al. 2017, [115] In vivo |
| Extracellular matrix organization | TNF (Tumor necrosis factor) | Role of a DNA methyltransferase (Dnmt3a) in obese AT from transgenic mice overexpressing Dnmt3a/AT/Array GWAS Affimetrics Mouse Genome 430 2.0 | Kamei et al. 2010, [129] In vivo/In vitro |
| Extracellular matrix organization | TNF (Tumor necrosis factor) | A RCT to study if the epigenome of human AT is affected differently by dietary fat composition/SAT/Array (Infinium Human Methylation 450K BeadChip, Illumina) | Perfilyev et al. 2017 [115] In vivo |
| Extracellular matrix organization | TNF (Tumor necrosis factor) | Case control study in obese subjects (Obese+Obese T2D) to evaluate if extracellular factors in obesity epigenetically reprogram adipogenesis potential and metabolic function of preadipocytes/VAT/Array EWAS RRBS | Andersen et al. 2019 [128] In vivo |
| Negative regulation of endothelial cell migration | PPARG (Peroxisome proliferator-activated receptor gamma) | A RCT to study if the epigenome of human AT is affected differently by dietary fat composition/SAT/Array (Infinium Human Methylation 450K BeadChip, Illumina) | Perfilyev et al. 2017, [115] In vivo |
| Negative regulation of endothelial cell migration | PPARG (Peroxisome proliferator-activated receptor gamma) | DNA methylation data in AT from monozygotic twin pairs discordant for T2D and independent case control cohorts/AT/Array EWAS Infinium Human Methylation 450K BeadChips (Illumina) | Nilsson et al. 2014, [122] In vivo |
| Negative regulation of endothelial cell migration | PPARG (Peroxisome proliferator-activated receptor gamma) | Genome-wide DNA promoter methylation along with mRNA profiles in non-obese vs. obese individuals/SAT and VAT/Array (Infinium HumanMethylation450K BeadChips (Illumina) | Keller et al. 2017, [127] In vivo/In silico |
| Regulation of cytoskeleton | |||
| Regulation of actin cytoskeleton | ARHGEF1 (Rho guanine nucleotide exchange factor 1) | Gene expression and methylation data from diabetic and healthy individuals/VAT/Array EZ-DNA methylation kit and Illumina Human Methylation 450K BeadChip | Wang et al. 2018, [65] In vivo |
| Regulation of actin cytoskeleton | ARPC3 (actin-related protein 2/3 complex subunit 3) | Association of CpG-SNPs located within ARPC3, which is linked to adipogenesis/VAT/Array High-throughput array technology QuantStudio 12K Flex System | Toro-Martín et al. 2016, [137] In vivo |
| Cell Signaling/Term | Gene Symbol and Name | Study/Tissue/Technology | Author, In Vitro/In Vivo |
|---|---|---|---|
| Collagens | |||
| Collagen-containing extracellular matrix | ASPN (Asporin) | Maintained weight loss or regained weight/AT/Microarray | Bolton et al. 2017, [143] In vivo |
| Collagen-containing extracellular matrix | COL24A1 (Collagen alpha-1(XXIV) chain) | Gene × Environment analyses/AT/Microarray | Hong et al. 2017, [144] In vivo |
| Collagen-containing extracellular matrix | COL6A1 (Collagen alpha-1(VI) chain) | ECM remodeling during the first year of bariatric surgery/SAT/Microarray and RT-PCR | Liu et al. 2016, [145] In vivo |
| Collagen-containing extracellular matrix | COL6A2 (Collagen alpha-2(VI) chain) | ECM remodeling during the first year of bariatric surgery/SAT/Microarray and RT-PCR | Liu et al. 2016, [145] In vivo |
| Collagen-containing extracellular matrix | EFEMP1 (EGF-containing fibulin-like extracellular matrix protein 1) | Copy number variants to early-onset obesity/SAT/Microarray | Petterson et al. 2017, [146] In vivo |
| Collagen-containing extracellular matrix | ELN (Elastin) | ECM remodeling during the first year of bariatric surgery/SAT/Microarray and RT-PCR | Liu et al. 2016, [145] In vivo |
| Collagen-containing extracellular matrix | LOXL2 (Lysyl oxidase homolog 2) | In vitro 3D culture system/AT/Microarray and RT-PCR | Pellegrinelli et al. 2014, [147] In vitro |
| Collagen-containing extracellular matrix | MMP8 (Neutrophil collagenase) | Gene signature and ECM remodeling/AT/RNA-seq and RT-PCR | Tokunaga et al. 2014, [148] In vitro |
| Collagen-containing extracellular matrix | SOD3 (Superoxide dismutase) | Bio-informatics analysis/WAT of C57BL/6J mice/Microarray of four datasets | Li et al. 2010, [149] In vivo |
| Collagen-containing extracellular matrix | SPARC (Secreted Protein Acidic And Cysteine Rich) | Maintained weight loss or regained weight/AT/Microarray | Bolton et al. 2017, [143] In vivo |
| Collagen-containing extracellular matrix | TGFB1 (Transforming growth factor beta-1 proprotein) | Maintained weight loss or regained weight/AT/Microarray | Bolton et al. 2017, [143] In vivo |
| Cell adhesion | |||
| Adherents junction | TCF7L2 (transcription factor 7 like 2) | Bio-informatics analysis/WAT of C57BL/6J mice/Microarray of four datasets | Li et al. 2010, [149] In vivo |
| Cell adhesion molecule binding | ANXA1 (Annexin A1) | VAT from obese prepubertal children/Microarray | Aguilera et al. 2015, [150] In vivo |
| Cell adhesion molecule binding | COL5A1 (Collagen alpha-1(V) chain) | SAT and VAT microarray | Arner et al. 2016, [126] In vivo |
| Cell adhesion molecule binding | EGFL6 (Epidermal growth factor-like protein 6) | Maintained weight loss or regained weight/Adipose/Microarray | Bolton et al. 2017, [143] In vivo |
| Regulation of cell adhesion | FSTL3 (Follistatin-related protein 3) | Dietary intervention/AT/Microarray and RT-PCR | Bolton et al. 2017, [143] In vivo |
| Cell adhesion molecule binding | ICAM1 (Intercellular adhesion molecule 1) | Culture of human Adipocytes exposed to IL1B/AT/Microarray | Kępczyńska et al. 2017, [151] In vitro |
| Cell adhesion molecule binding | IL1B (Interleukin-1 beta) | Culture of human Adipocytes exposed to IL1B/AT/Microarray | Kępczyńska et al. 2017, [151] In vitro |
| Cell adhesion molecule binding | IL1B (Interleukin-1 beta) | Inter-Tissue Gene Co-Expression Networks liver/muscle, SAT, and VAT/HumanOmni-BeadChips (Illumina) | Kogelman et al. 2016, [152] In vivo |
| Regulation of cell adhesion | IRF1 (Interferon regulatory factor 1) | Bio-informatics analysis/WAT of C57BL/6J mice/Microarray of four datasets | Li et al. 2010, [149] In vivo |
| Cell adhesion molecule binding | LGALS12 (Galectin-12) | Bio-informatics analysis/WAT of C57BL/6J mice/Microarray of four datasets | Li et al. 2010, [149] In vivo |
| Cell adhesion molecule binding | PTPRJ (Receptor-type tyrosine-protein phosphatase eta) | SAT and VAT microarray | Arner et al. 2016, [126] In vivo |
| Cell adhesion molecule binding | VCAM1 (Vascular cell adhesion protein 1) | Culture of human Adipocytes exposed to IL1B/AT/Microarray | Kępczyńska et al. 2017, [151] In vitro |
| Focal adhesion | |||
| Focal adhesion | JUN (Jun proto-oncogene, AP-1 transcription factor subunit) | Bio-informatics analysis/WAT of C57BL/6J mice/Microarray of four datasets | Li et al. 2010, [149] In vivo |
| ECM assembly and disassembly | |||
| Extracellular matrix disassembly | FURIN | Culture human subcutaneous adipocytes/AT/RNA-seq | de Oliveira et al. 2020, [153] In vitro |
| Extracellular matrix cell signaling | FZD4 (Frizzled-4) | Bio-informatics analysis/WAT of C57BL/6J mice/Microarray of four datasets | Li et al. 2010, [149] In vivo |
| Extracellular matrix disassembly | MMP13 (Matrix metalloproteinase-13) | ASCs isolated from subcutaneous and visceral adipose tissues/AT/RNA-seq and qRT-PCR | Tokunaga et al. 2014, [148] In vivo |
| Extracellular matrix | TLR3 (Toll-like receptor 3) | Culture human subcutaneous adipocytes/AT/RNA-seq | de Oliveira et al. 2020, [153] In vivo |
| Matrix metallopeptidase secretion | TLR4 (Toll-like receptor 4) | Culture of mononuclear cells with adipocytes/AT/RT-PCR and PCR array | Samuvel et al. 2010, [154] In vivo |
| Endothelial Cell migration | |||
| Positive regulation of endothelial cell migration | GATA3 (Trans-acting T-cell-specific transcription factor GATA-3) | Bio-informatics analysis/WAT of C57BL/6J mice/Microarray of four datasets | Li et al. 2010, [149] In vivo |
| Positive regulation of endothelial cell migration | SIRT1 (NAD-dependent protein deacetylase sirtuin-1) | Culture human subcutaneous adipocytes/AT/RNA-seq | de Oliveira et al. 2020, [153] In vivo |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anguita-Ruiz, A.; Bustos-Aibar, M.; Plaza-Díaz, J.; Mendez-Gutierrez, A.; Alcalá-Fdez, J.; Aguilera, C.M.; Ruiz-Ojeda, F.J. Omics Approaches in Adipose Tissue and Skeletal Muscle Addressing the Role of Extracellular Matrix in Obesity and Metabolic Dysfunction. Int. J. Mol. Sci. 2021, 22, 2756. https://doi.org/10.3390/ijms22052756
Anguita-Ruiz A, Bustos-Aibar M, Plaza-Díaz J, Mendez-Gutierrez A, Alcalá-Fdez J, Aguilera CM, Ruiz-Ojeda FJ. Omics Approaches in Adipose Tissue and Skeletal Muscle Addressing the Role of Extracellular Matrix in Obesity and Metabolic Dysfunction. International Journal of Molecular Sciences. 2021; 22(5):2756. https://doi.org/10.3390/ijms22052756
Chicago/Turabian StyleAnguita-Ruiz, Augusto, Mireia Bustos-Aibar, Julio Plaza-Díaz, Andrea Mendez-Gutierrez, Jesús Alcalá-Fdez, Concepción María Aguilera, and Francisco Javier Ruiz-Ojeda. 2021. "Omics Approaches in Adipose Tissue and Skeletal Muscle Addressing the Role of Extracellular Matrix in Obesity and Metabolic Dysfunction" International Journal of Molecular Sciences 22, no. 5: 2756. https://doi.org/10.3390/ijms22052756
APA StyleAnguita-Ruiz, A., Bustos-Aibar, M., Plaza-Díaz, J., Mendez-Gutierrez, A., Alcalá-Fdez, J., Aguilera, C. M., & Ruiz-Ojeda, F. J. (2021). Omics Approaches in Adipose Tissue and Skeletal Muscle Addressing the Role of Extracellular Matrix in Obesity and Metabolic Dysfunction. International Journal of Molecular Sciences, 22(5), 2756. https://doi.org/10.3390/ijms22052756