
Palmitic Acid Impairs Myogenesis and Alters Temporal Expression of miR-133a and miR-206 in C2C12 Myoblasts

 , , and
, , and 
Abstract
1. Introduction
2. Results
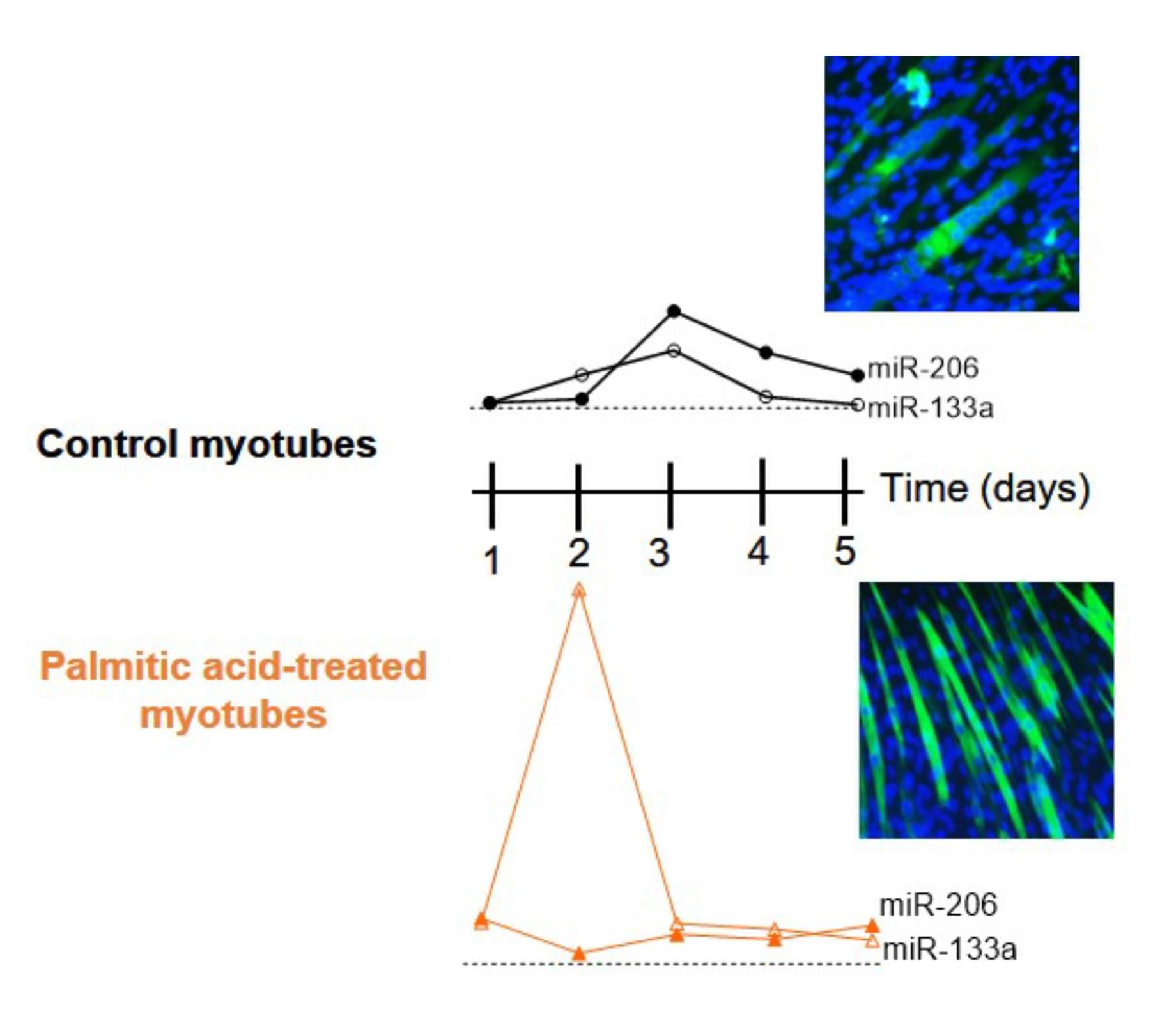
2.1. Effect of PA on Viability, DNA Fragmentation and Proliferation of C2C12 Cells
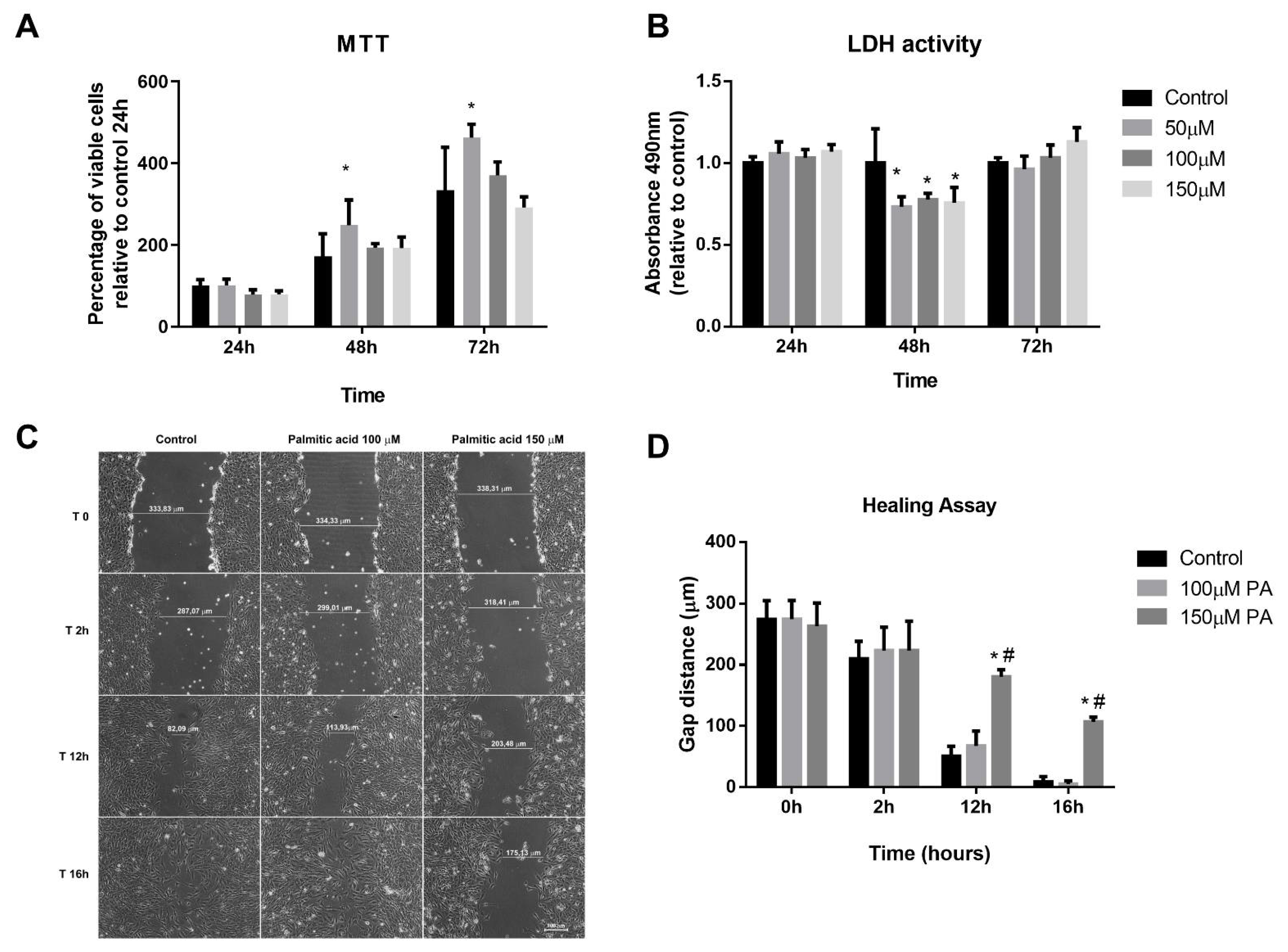
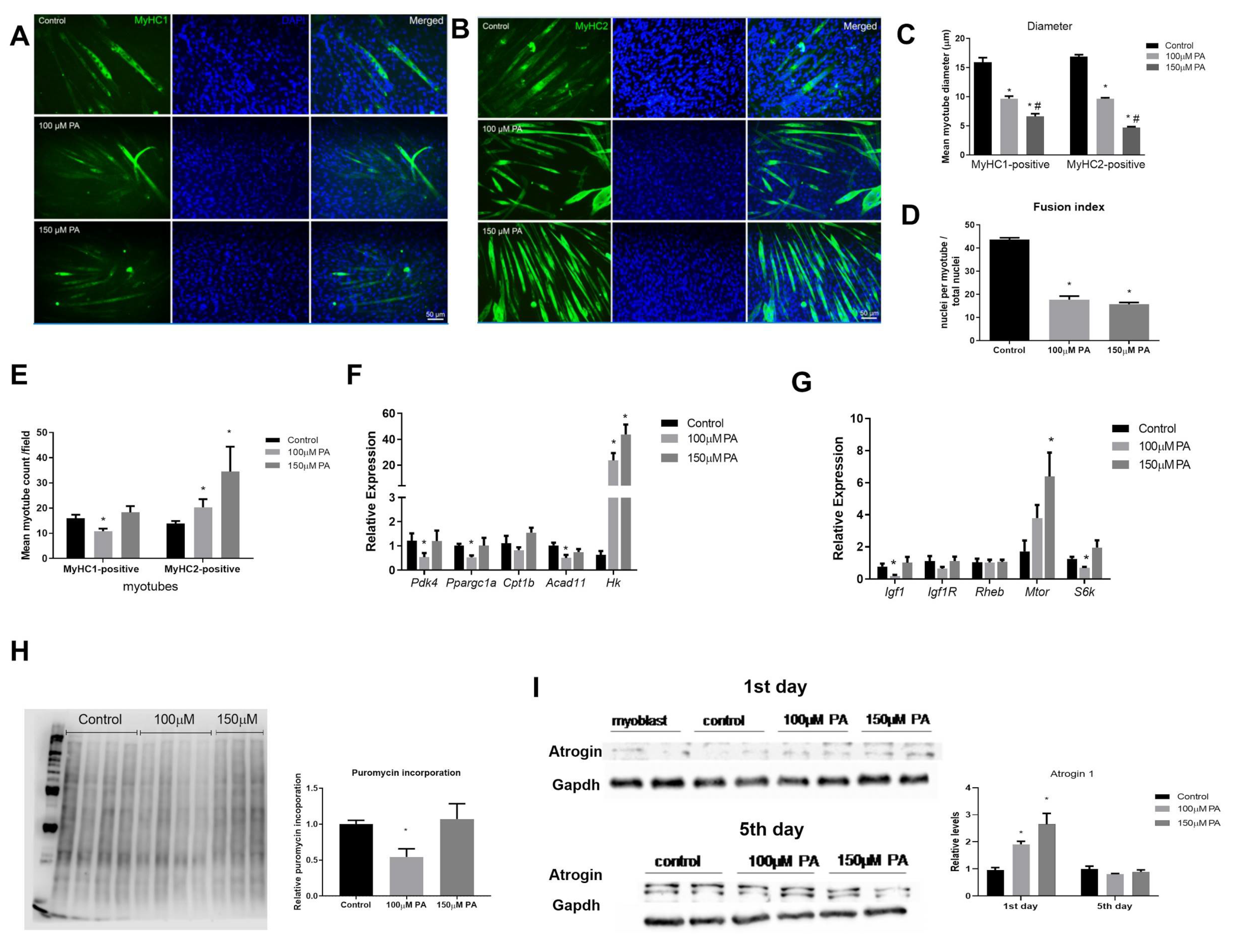
2.2. PA Induces Smaller C2C12 Myotubes and Metabolic Remodeling
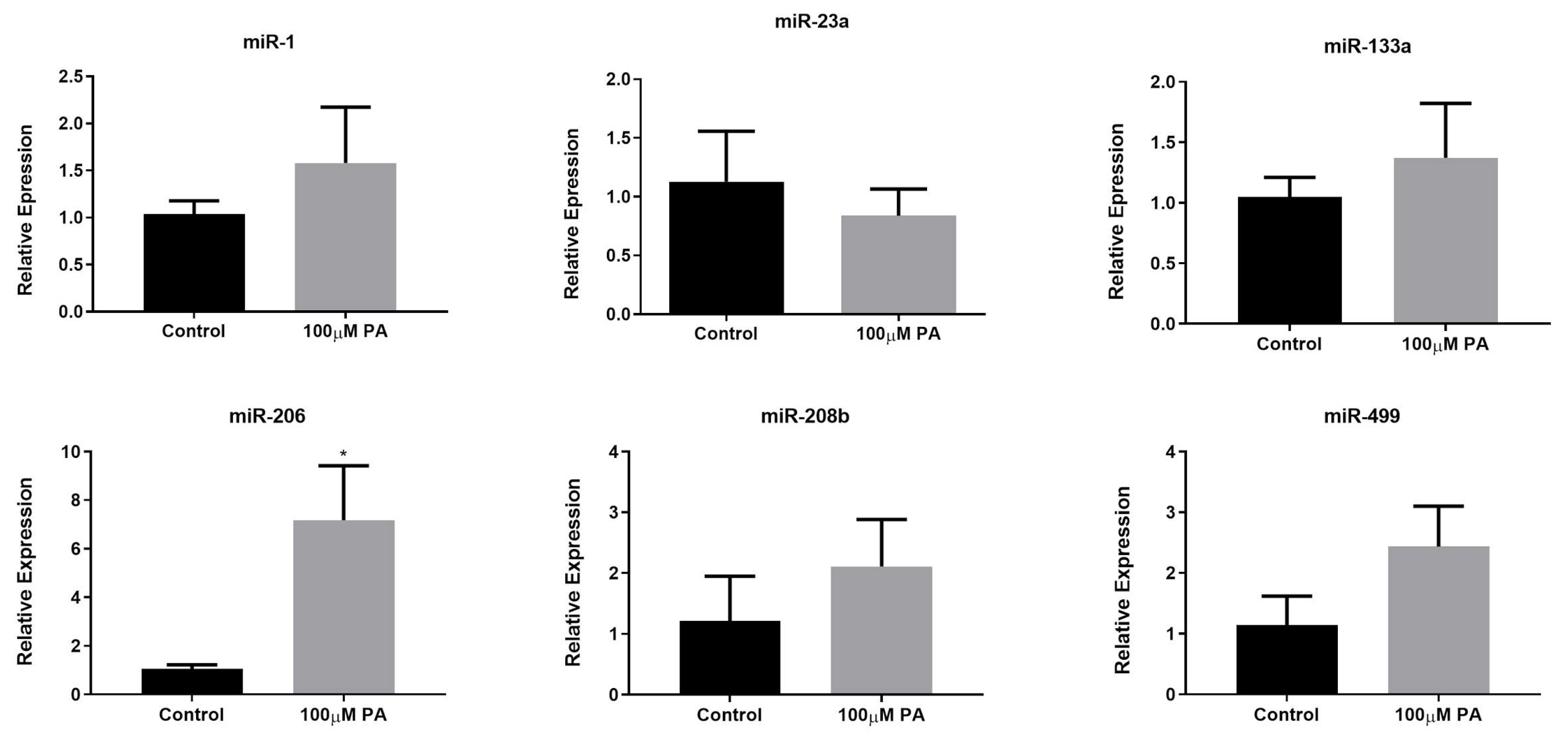
2.3. PA Increases miR-206 Expression in C2C12 Myotubes
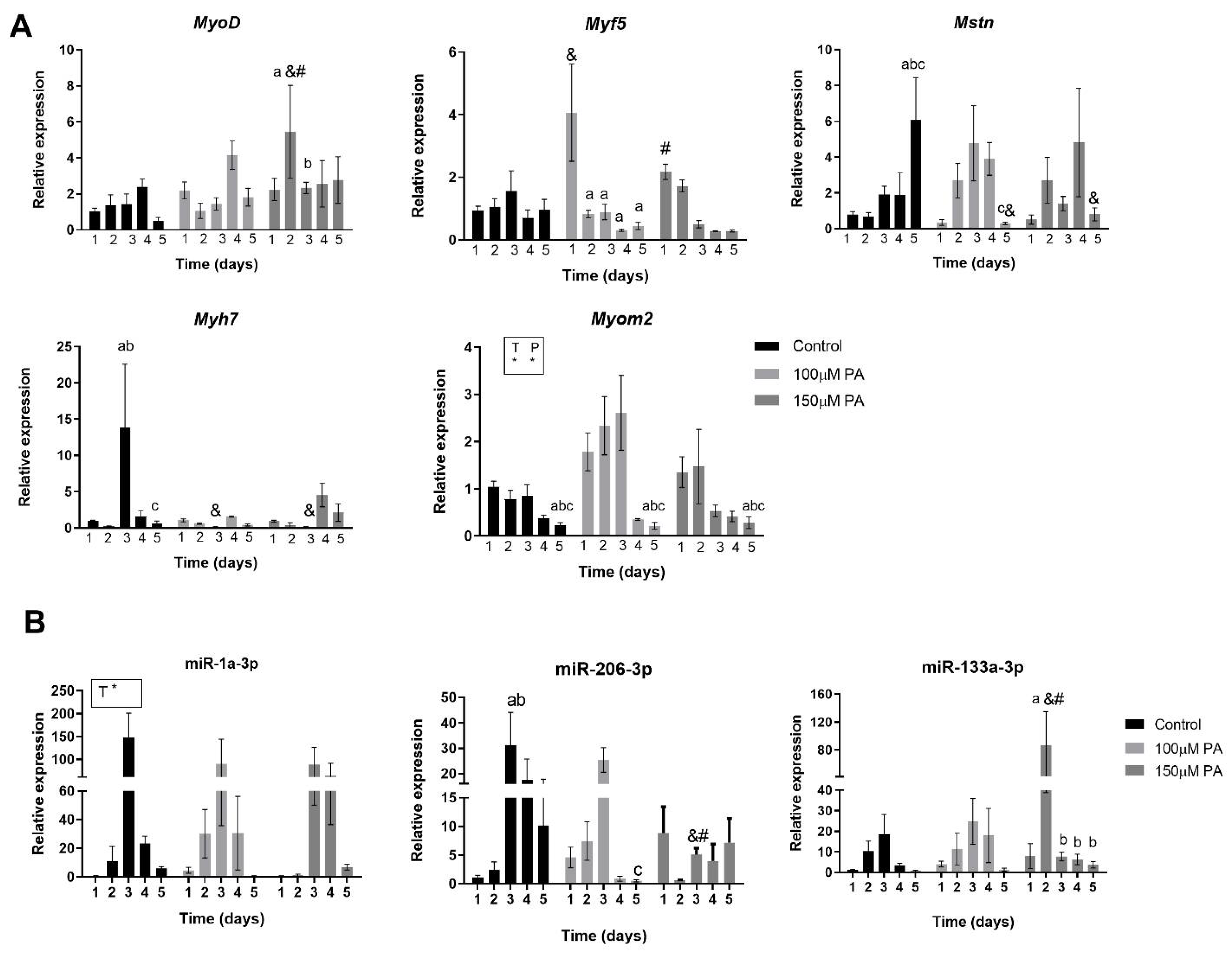
2.4. PA Alters Expression of Myogenic Markers during Myoblast Differentiation
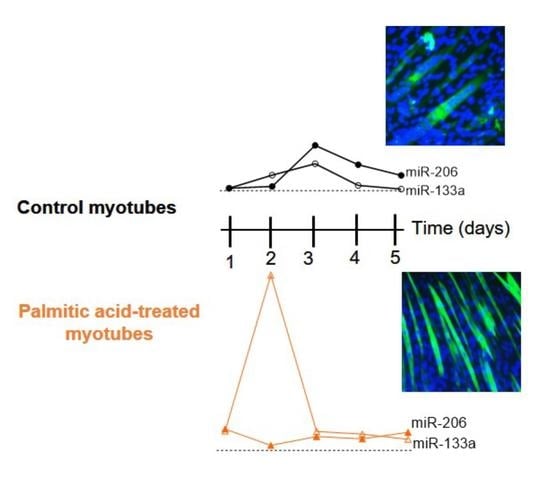
2.5. PA Affects Temporal Expression of miR-133a and miR-206 during Myoblast Differentiation
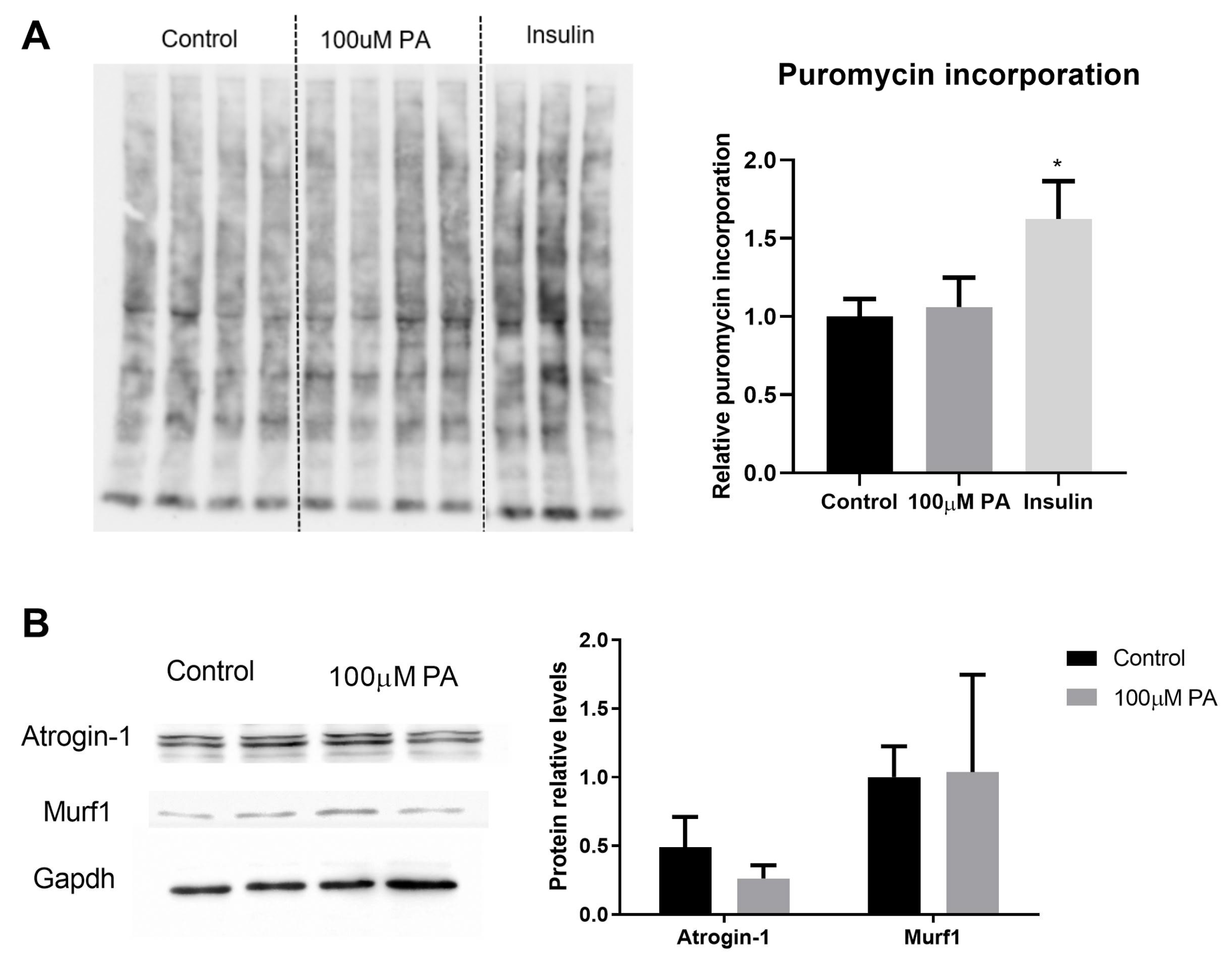
2.6. PA Impairs the Growth and Promotes Metabolic Remodeling of C2C12 Myotubes
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Cell Treatments
4.2. Cytotoxicity Assays
4.3. Healing Assay
4.4. Immunofluorescence
4.5. RNA Preparation and RT-qPCR
4.6. Western Blotting
4.7. Puromycin Incorporation Assay
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Astrup, A.; Ryan, L.; Grunwald, G.K.; Storgaard, M.; Saris, W.; Melanson, E.; Hill, J.O. The role of dietary fat in body fatness: Evidence from a preliminary meta-analysis of ad libitum low-fat dietary intervention studies. Br. J. Nutr. 2000, 83 (Suppl. 1), S25–S32. [Google Scholar] [CrossRef]
- Townley-Tilson, W.H.D.; Callis, T.E.; Wang, D. MicroRNAs 1, 133, and 206: Critical factors of skeletal and cardiac muscle development, function, and disease. Int. J. Biochem. Cell Biol. 2010, 42, 1252–1255. [Google Scholar] [CrossRef]
- Chavez, J.A.; Summers, S.A. Characterizing the effects of saturated fatty acids on insulin signaling and ceramide and diacylglycerol accumulation in 3T3-L1 adipocytes and C2C12 myotubes. Arch. Biochem. Biophys. 2003, 419, 101–109. [Google Scholar] [CrossRef]
- Cheon, H.G.; Cho, Y.S. Protection of palmitic acid-mediated lipotoxicity by arachidonic acid via channeling of palmitic acid into triglycerides in C2C12. J. Biomed. Sci. 2014, 21, 13. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.-T.; Wang, T.-Z.; Leng, J.; Chen, Y.; Liu, J.-B.; Liu, Y.; Wang, W.-J. Palmitate contributes to insulin resistance through downregulation of the Src-mediated phosphorylation of Akt in C2C12 myotubes. Biosci. Biotechnol. Biochem. 2012, 76, 1356–1361. [Google Scholar] [CrossRef]
- Wang, X.; Yu, W.; Nawaz, A.; Guan, F.; Sun, S.; Wang, C. Palmitate induced insulin resistance by PKCtheta-dependent activation of mTOR/S6K pathway in C2C12 myotubes. Exp. Clin. Endocrinol. Diabetes 2010, 118, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Bryner, R.W.; Woodworth-Hobbs, M.E.; Williamson, D.L.; Alway, S.E. Docosahexaenoic Acid protects muscle cells from palmitate-induced atrophy. ISRN Obes 2012, 2012, 647348. [Google Scholar] [CrossRef]
- Lee, H.; Lim, J.-Y.; Choi, S.-J. Oleate Prevents Palmitate-Induced Atrophy via Modulation of Mitochondrial ROS Production in Skeletal Myotubes. Oxid. Med. Cell. Longev. 2017, 2017, 2739721. [Google Scholar] [CrossRef] [PubMed]
- Woodworth-Hobbs, M.E.; Hudson, M.B.; Rahnert, J.A.; Zheng, B.; Franch, H.A.; Price, S.R. Docosahexaenoic acid prevents palmitate-induced activation of proteolytic systems in C2C12 myotubes. J. Nutr. Biochem. 2014, 25, 868–874. [Google Scholar] [CrossRef] [PubMed]
- de Frias, F.T.; de Mendonça, M.; Martins, A.R.; Gindro, A.F.; Cogliati, B.; Curi, R.; Rodrigues, A.C. MyomiRs as Markers of Insulin Resistance and Decreased Myogenesis in Skeletal Muscle of Diet-Induced Obese Mice. Front. Endocrinol. 2016, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.A.; Thomou, T.; Boucher, J.; Lee, K.Y.; Lallukka, S.; Kim, J.K.; Torriani, M.; Yki-Järvinen, H.; Grinspoon, S.K.; Cypess, A.M.; et al. Altered miRNA processing disrupts brown/white adipocyte determination and associates with lipodystrophy. J. Clin. Investig. 2014, 124, 3339–3351. [Google Scholar] [CrossRef]
- Wang, F.; Wang, J.; He, J.; Li, W.; Li, J.; Chen, S.; Zhang, P.; Liu, H.; Chen, X. Serum miRNAs miR-23a, 206, and 499 as Potential Biomarkers for Skeletal Muscle Atrophy. Biomed. Res. Int. 2017, 2017, 8361237. [Google Scholar] [CrossRef]
- Bjorkman, K.K.; Guess, M.G.; Harrison, B.C.; Polmear, M.M.; Peter, A.K.; Leinwand, L.A. miR-206 enforces a slow muscle phenotype. J. Cell Sci. 2020, 133. [Google Scholar] [CrossRef]
- McPherron, A.C.; Lee, S.-J. Suppression of body fat accumulation in myostatin-deficient mice. J. Clin. Investig. 2002, 109, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Elkina, Y.; von Haehling, S.; Anker, S.D.; Springer, J. The role of myostatin in muscle wasting: An overview. J. Cachexia Sarcopenia Muscle 2011, 2, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Tintignac, L.A.; Lagirand, J.; Batonnet, S.; Sirri, V.; Leibovitch, M.P.; Leibovitch, S.A. Degradation of MyoD mediated by the SCF (MAFbx) ubiquitin ligase. J. Biol. Chem. 2005, 280, 2847–2856. [Google Scholar] [CrossRef]
- Tachtsis, B.; Whitfield, J.; Hawley, J.A.; Hoffman, N.J. Omega-3 Polyunsaturated Fatty Acids Mitigate Palmitate-Induced Impairments in Skeletal Muscle Cell Viability and Differentiation. Front. Physiol. 2020, 11, 563. [Google Scholar] [CrossRef]
- Cnop, M.; Hannaert, J.C.; Hoorens, A.; Eizirik, D.L.; Pipeleers, D.G. Inverse relationship between cytotoxicity of free fatty acids in pancreatic islet cells and cellular triglyceride accumulation. Diabetes 2001, 50, 1771–1777. [Google Scholar] [CrossRef]
- Richieri, G.V.; Anel, A.; Kleinfeld, A.M. Interactions of long-chain fatty acids and albumin: Determination of free fatty acid levels using the fluorescent probe ADIFAB. Biochemistry 1993, 32, 7574–7580. [Google Scholar] [CrossRef]
- Masi, L.N.; Rodrigues, A.C.; Curi, R. Fatty acids regulation of inflammatory and metabolic genes. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 418–424. [Google Scholar] [CrossRef]
- Winbanks, C.E.; Beyer, C.; Hagg, A.; Qian, H.; Sepulveda, P.V.; Gregorevic, P. miR-206 Represses Hypertrophy of Myogenic Cells but Not Muscle Fibers via Inhibition of HDAC4. PLoS ONE 2013, 8, e73589. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.I.; Guimarães, D.; Sponton, C.H.; Bajgelman, M.C.; Palameta, S.; Toscaro, J.M.; Reis, O.; Silveira, L.R. Essential role of the PGC-1α/PPARβ axis in Ucp3 gene induction. J. Physiol. 2019, 597, 4277–4291. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, D.M.; Trajcevski, K.E.; Al-Sajee, D.; Wang, D.C.; Thomas, M.; Anderson, J.E.; Hawke, T.J. Diet-induced obesity impairs muscle satellite cell activation and muscle repair through alterations in hepatocyte growth factor signaling. Physiol. Rep. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Dedieu, S.; Mazères, G.; Cottin, P.; Brustis, J.-J. Involvement of myogenic regulator factors during fusion in the cell line C2C12. Int. J. Dev. Biol. 2002, 46, 235–241. [Google Scholar] [PubMed]
- Panda, A.C.; Abdelmohsen, K.; Martindale, J.L.; Di Germanio, C.; Yang, X.; Grammatikakis, I.; Noh, J.H.; Zhang, Y.; Lehrmann, E.; Dudekula, D.B.; et al. Novel RNA-binding activity of MYF5 enhances Ccnd1/Cyclin D1 mRNA translation during myogenesis. Nucleic Acids Res. 2016, 44, 2393–2408. [Google Scholar] [CrossRef]
- Chen, X.; Wang, K.; Chen, J.; Guo, J.; Yin, Y.; Cai, X.; Guo, X.; Wang, G.; Yang, R.; Zhu, L.; et al. In vitro evidence suggests that miR-133a-mediated regulation of uncoupling protein 2 (UCP2) is an indispensable step in myogenic differentiation. J. Biol. Chem. 2009, 284, 5362–5369. [Google Scholar] [CrossRef]
- Rao, P.K.; Kumar, R.M.; Farkhondeh, M.; Baskerville, S.; Lodish, H.F. Myogenic factors that regulate expression of muscle-specific microRNAs. Proc. Natl. Acad. Sci. USA 2006, 103, 8721–8726. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.H.; Ro, S.-H.; Cao, J.; Otto, N.M.; Kim, D.-H. mTOR regulation of autophagy. FEBS Lett. 2010, 584, 1287–1295. [Google Scholar] [CrossRef]
- Kalista, S.; Schakman, O.; Gilson, H.; Lause, P.; Demeulder, B.; Bertrand, L.; Pende, M.; Thissen, J.P. The type 1 insulin-like growth factor receptor (IGF-IR) pathway is mandatory for the follistatin-induced skeletal muscle hypertrophy. Endocrinology 2012, 153, 241–253. [Google Scholar] [CrossRef]
- Papaconstantinou, J.; Hsieh, C.-C. IGF-1 mediated phosphorylation of specific IRS-1 serines in Ames dwarf fibroblasts is associated with longevity. Oncotarget 2015, 6, 35315–35323. [Google Scholar] [CrossRef]
- Hirsch, T.; Spielmann, M.; Velander, P.; Zuhaili, B.; Bleiziffer, O.; Fossum, M.; Steinstraesser, L.; Yao, F.; Eriksson, E. Insulin-like growth factor-1 gene therapy and cell transplantation in diabetic wounds. J. Gene Med. 2008, 10, 1247–1252. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Wang, H.; Lee, I.H.; Modi, S.; Wang, X.; Du, J.; Mitch, W.E. PTEN inhibition improves muscle regeneration in mice fed a high-fat diet. Diabetes 2010, 59, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Stuart, C.A.; McCurry, M.P.; Marino, A.; South, M.A.; Howell, M.E.A.; Layne, A.S.; Ramsey, M.W.; Stone, M.H. Slow-twitch fiber proportion in skeletal muscle correlates with insulin responsiveness. J. Clin. Endocrinol. Metab. 2013, 98, 2027–2036. [Google Scholar] [CrossRef] [PubMed]
- Talbot, J.; Maves, L. Skeletal muscle fiber type: Using insights from muscle developmental biology to dissect targets for susceptibility and resistance to muscle disease. Wiley Interdiscip. Rev. Dev. Biol. 2016, 5, 518–534. [Google Scholar] [CrossRef]
- Webster, C.; Silberstein, L.; Hays, A.P.; Blau, H.M. Fast muscle fibers are preferentially affected in Duchenne muscular dystrophy. Cell 1988, 52, 503–513. [Google Scholar] [CrossRef]
- Quiat, D.; Voelker, K.A.; Pei, J.; Grishin, N.V.; Grange, R.W.; Bassel-Duby, R.; Olson, E.N. Concerted regulation of myofiber-specific gene expression and muscle performance by the transcriptional repressor Sox6. Proc. Natl. Acad. Sci. USA 2011, 108, 10196–10201. [Google Scholar] [CrossRef]
- Grove, B.K.; Holmbom, B.; Thornell, L.E. Myomesin and M protein: Differential expression in embryonic fibers during pectoral muscle development. Differentiation 1987, 34. [Google Scholar] [CrossRef]
- Steiner, F.; Weber, K.; Fürst, D.O. Structure and expression of the gene encoding murine M-protein, a sarcomere-specific member of the immunoglobulin superfamily. Genomics 1998, 49, 83–95. [Google Scholar] [CrossRef]
- McCarthy, J.J.; Esser, K.A.; Andrade, F.H. MicroRNA-206 is overexpressed in the diaphragm but not the hindlimb muscle of mdx mouse. Am. J. Physiol. Cell Physiol. 2007, 293, C451–C457. [Google Scholar] [CrossRef]
- Gallagher, I.J.; Scheele, C.; Keller, P.; Nielsen, A.R.; Remenyi, J.; Fischer, C.P.; Roder, K.; Babraj, J.; Wahlestedt, C.; Hutvagner, G.; et al. Integration of microRNA changes in vivo identifies novel molecular features of muscle insulin resistance in type 2 diabetes. Genome Med. 2010, 2, 9. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|
| Pdk4 | GTTCCTTCACACCTTCACCACA | CCTCCTCGGTCAGAAATCTTGA |
| Ppargc1a | CACCAAACCCACAGAAAACAG | GGGTCAGAGGAAGAGATAAAGTTG |
| Cpt1b | CCTCCGAAAAGCACCAAAAC | GCTCCAGGGTTCAGAAAGTAC |
| Igf1 | GTGAGCCAAAGACACACCCA | ACCTCTGATTTTCCGAGTTGC |
| Igf1R | CTCTGTTACCTCTCCACCAT | CTTCTCACACATGGGCTTCT |
| S6K | GGAGGGACAGAAGAGAATCA | AACCTAGAACCACACCAATG |
| Rheb | CGATCCAACCATAGAGAACAC | AATATTCATCCTGCCCCGCT |
| Mtor | TGCCGCTGAGAGATGACAATG | GTTGTTAATGCTGATGAGGG |
| Mstn | GCAAAATTGGCTCAAACAGCC | AGGGATTCAGCCCATCTTCTC |
| Myf5 | TGACGGCATGCCTGAATGT | GCTGGACAAGCAATCCAAGC |
| MyoD | GCCCGCGCTCCAACTGCTCTGAT | CCTACGGTGGTGCGCCCTCTGC |
| MyH7 | GCCAACTATGCTGGAGCTGATGCCC | GGTGCGTGGAGCGCAAGTTTGTCATAAG |
| Myom 2 | GATCAACAGGGCCAACTTTGA | TGGTAGACACTTGTTCATGGGAAT |
| Acad11 | AGATGCTTCAGTTATCGGAACG | ATGTAGCCATGCCAGGGTTTC |
| Hk | GCTCAGAAAAGGGGGATTTC | TCAGCGACGTGATCAAAAAG |
| Rpl0 | TAAAGACTGGAGACAAGGTG | GTGTACTCAGTCTCCACAGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Paixão, A.O.; Bolin, A.P.; Silvestre, J.G.; Rodrigues, A.C. Palmitic Acid Impairs Myogenesis and Alters Temporal Expression of miR-133a and miR-206 in C2C12 Myoblasts. Int. J. Mol. Sci. 2021, 22, 2748. https://doi.org/10.3390/ijms22052748
da Paixão AO, Bolin AP, Silvestre JG, Rodrigues AC. Palmitic Acid Impairs Myogenesis and Alters Temporal Expression of miR-133a and miR-206 in C2C12 Myoblasts. International Journal of Molecular Sciences. 2021; 22(5):2748. https://doi.org/10.3390/ijms22052748
Chicago/Turabian Styleda Paixão, Ailma O., Anaysa Paola Bolin, João G. Silvestre, and Alice Cristina Rodrigues. 2021. "Palmitic Acid Impairs Myogenesis and Alters Temporal Expression of miR-133a and miR-206 in C2C12 Myoblasts" International Journal of Molecular Sciences 22, no. 5: 2748. https://doi.org/10.3390/ijms22052748
APA Styleda Paixão, A. O., Bolin, A. P., Silvestre, J. G., & Rodrigues, A. C. (2021). Palmitic Acid Impairs Myogenesis and Alters Temporal Expression of miR-133a and miR-206 in C2C12 Myoblasts. International Journal of Molecular Sciences, 22(5), 2748. https://doi.org/10.3390/ijms22052748