
Genome Wide Analysis of U-Box E3 Ubiquitin Ligases in Wheat (Triticum aestivum L.)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Identification of U-Box E3 Genes in the Wheat Genome
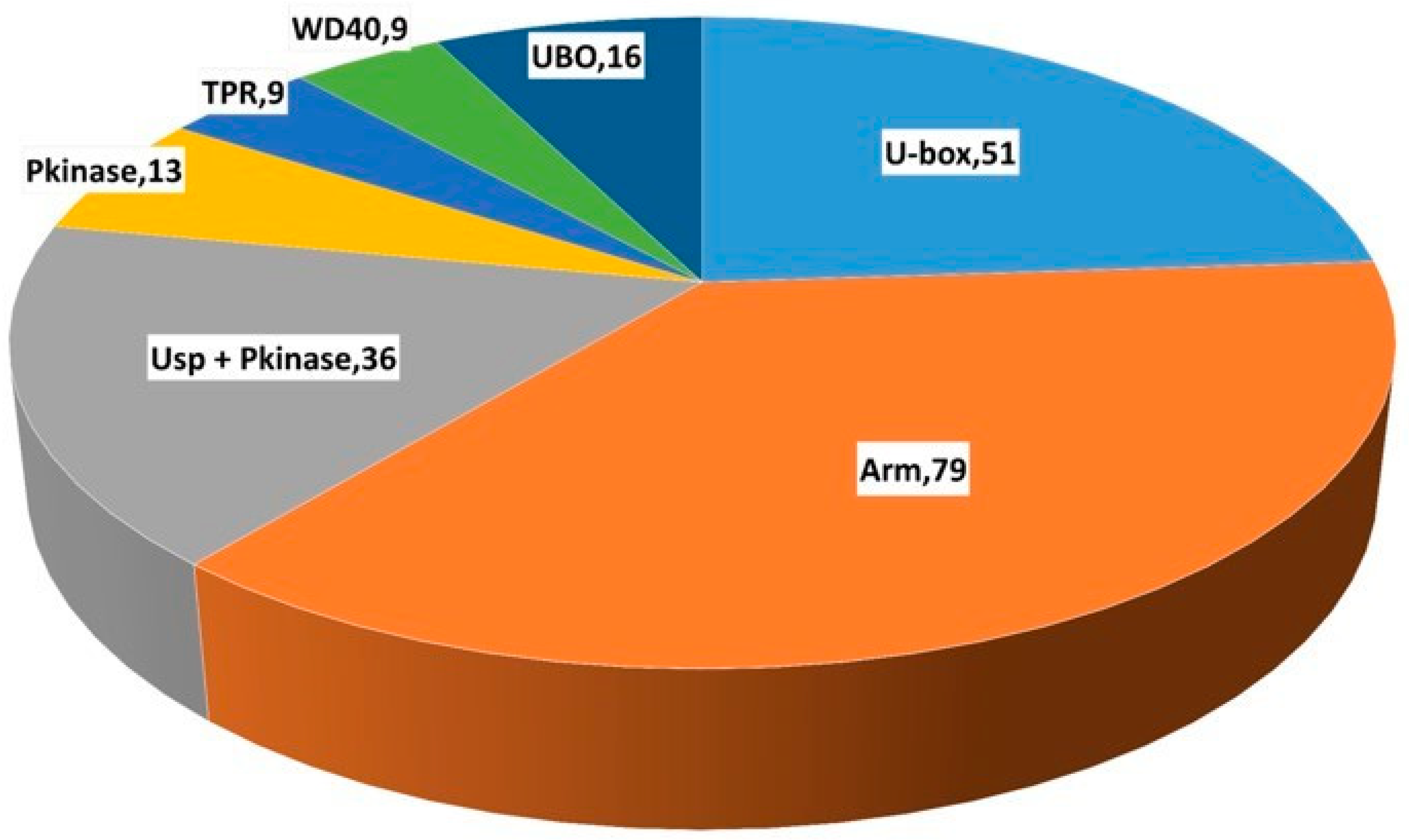
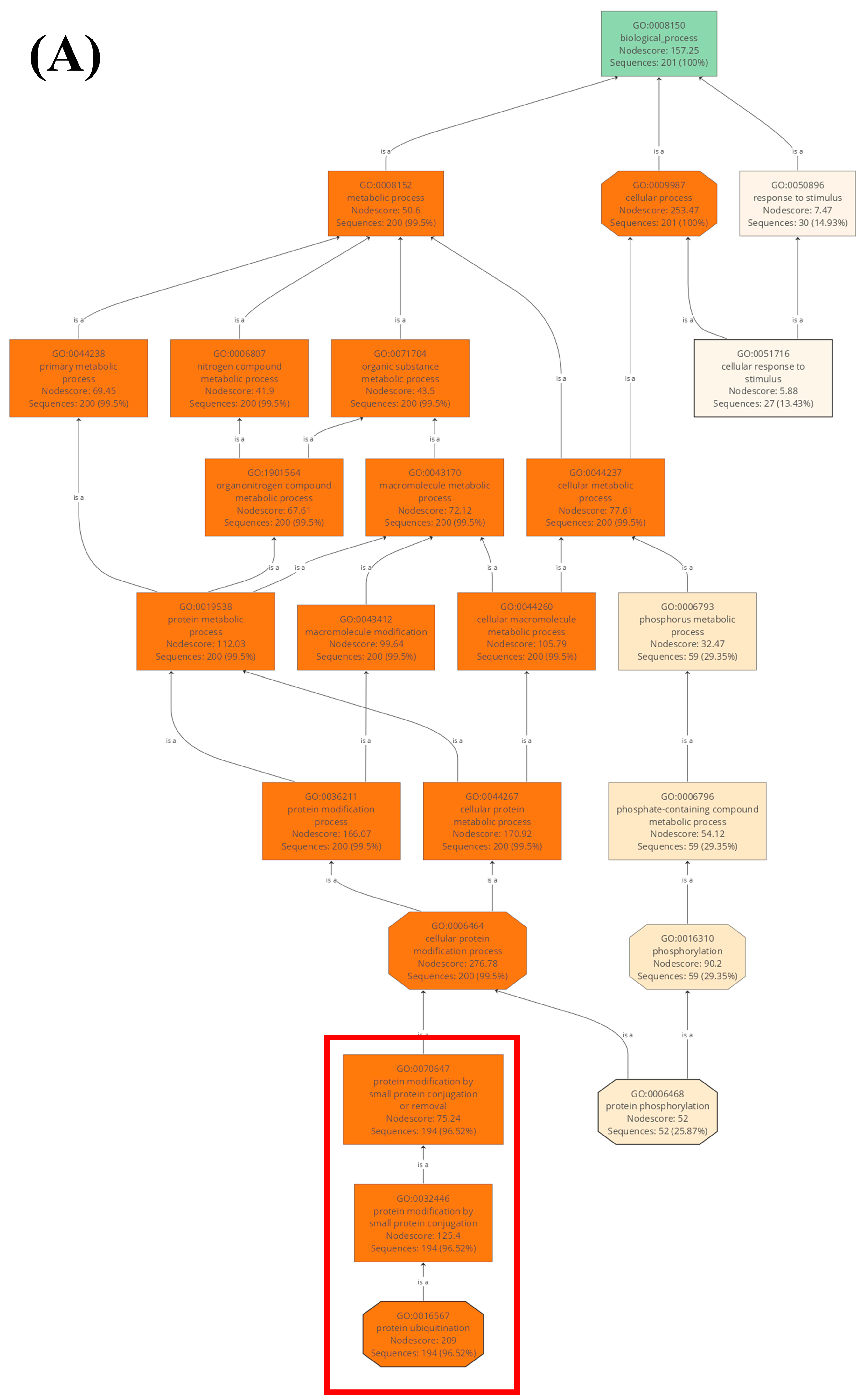
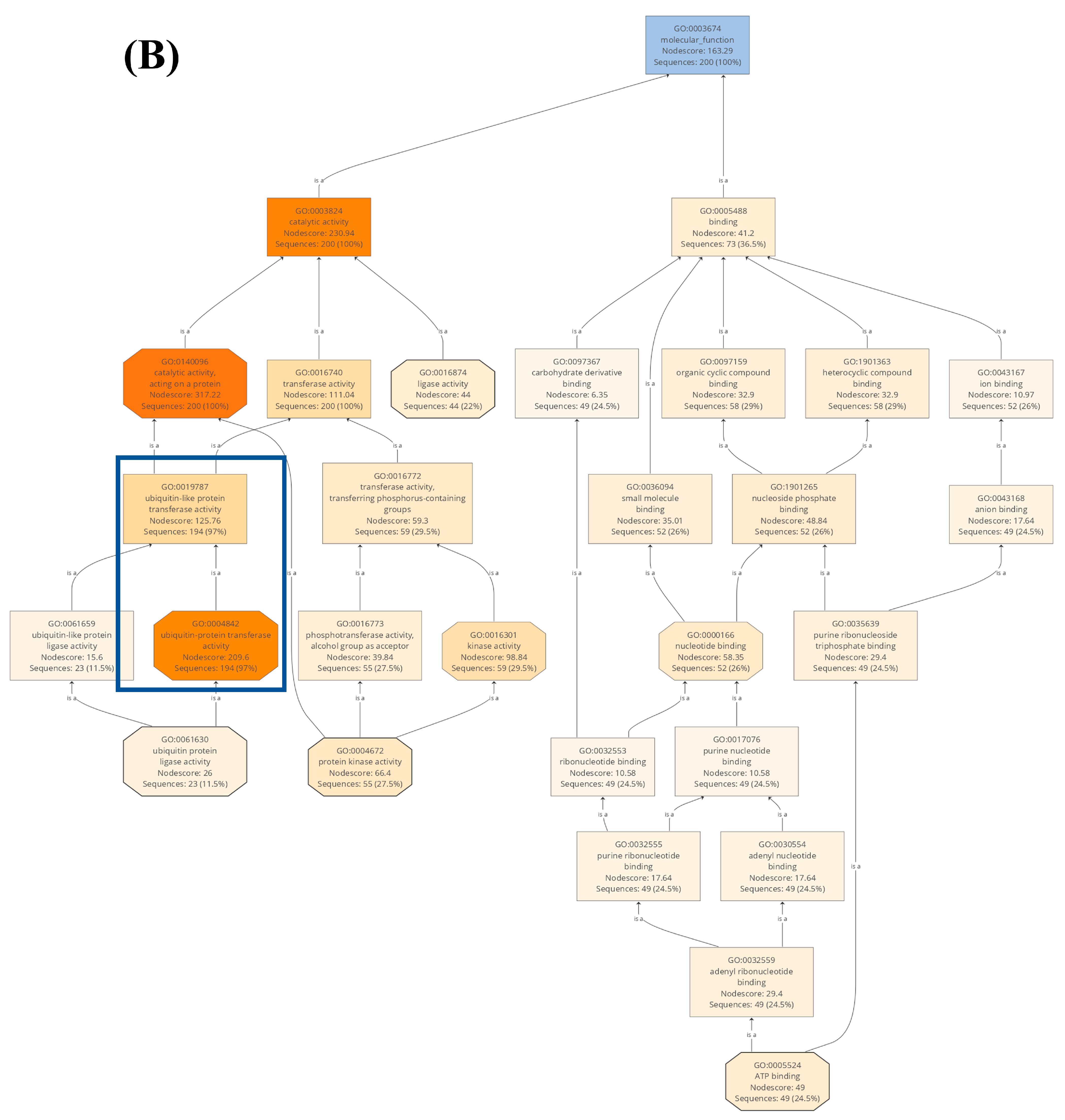
2.2. Gene Ontology Analysis of U-Box E3 Genes in the Wheat Genome
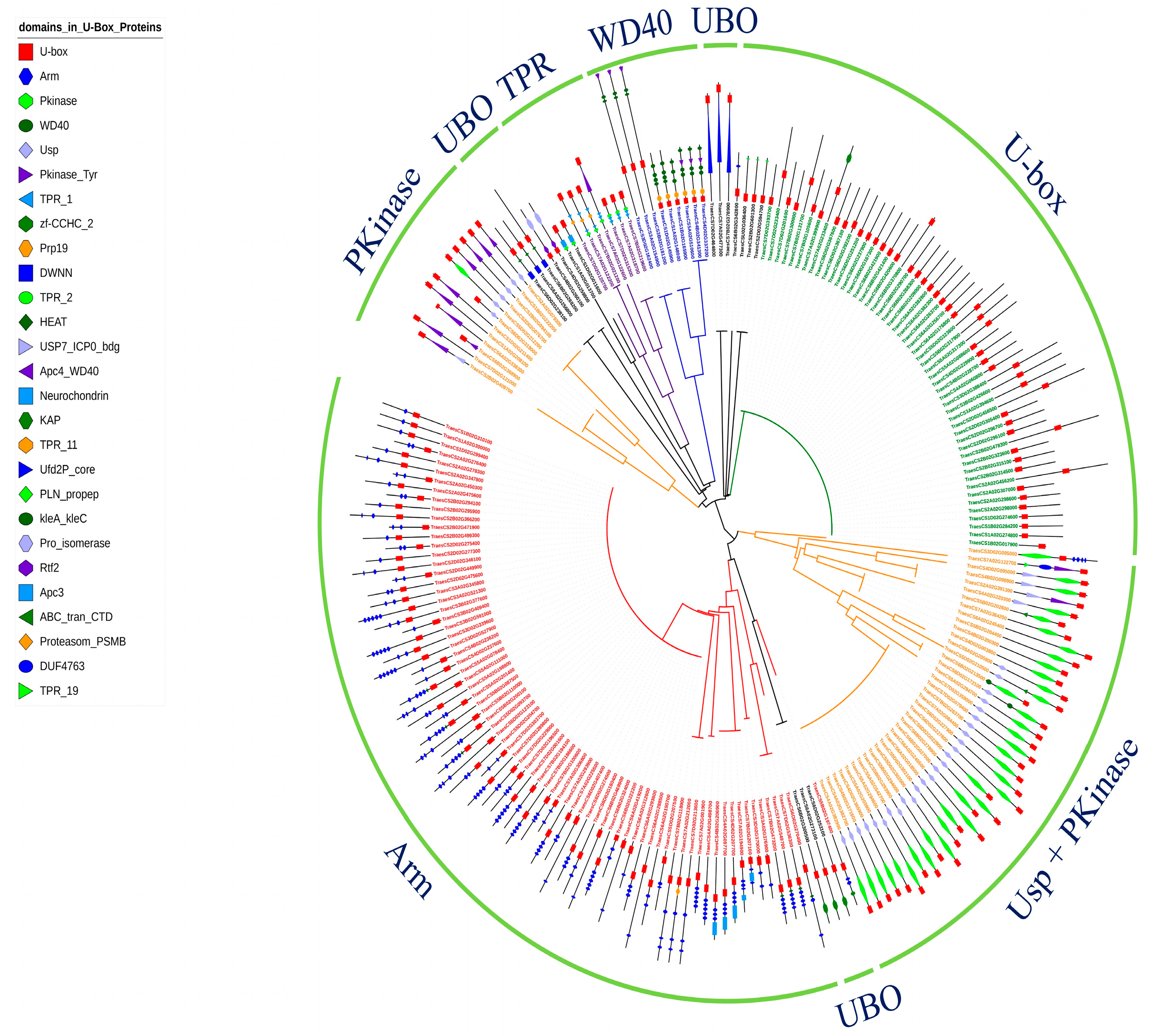
2.3. Phylogenetic Analysis and Chromosomal Location of U-Box E3 Genes
2.4. Detection of Homoeologous U-Box E3 Genes with High Sequence Homology between Triticum uratu, Aegilops tauschii, and Wheat
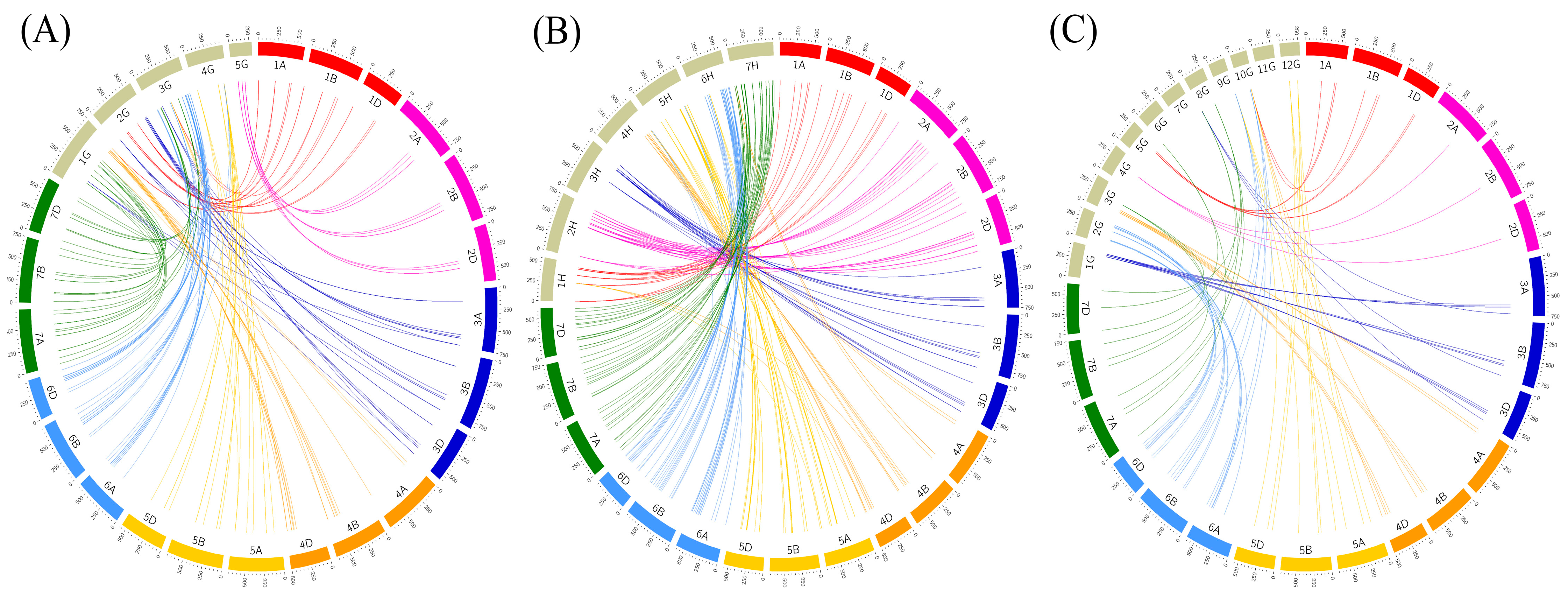
2.5. Synteny Analysis of U-Box E3 Genes in Wheat and Other Plants
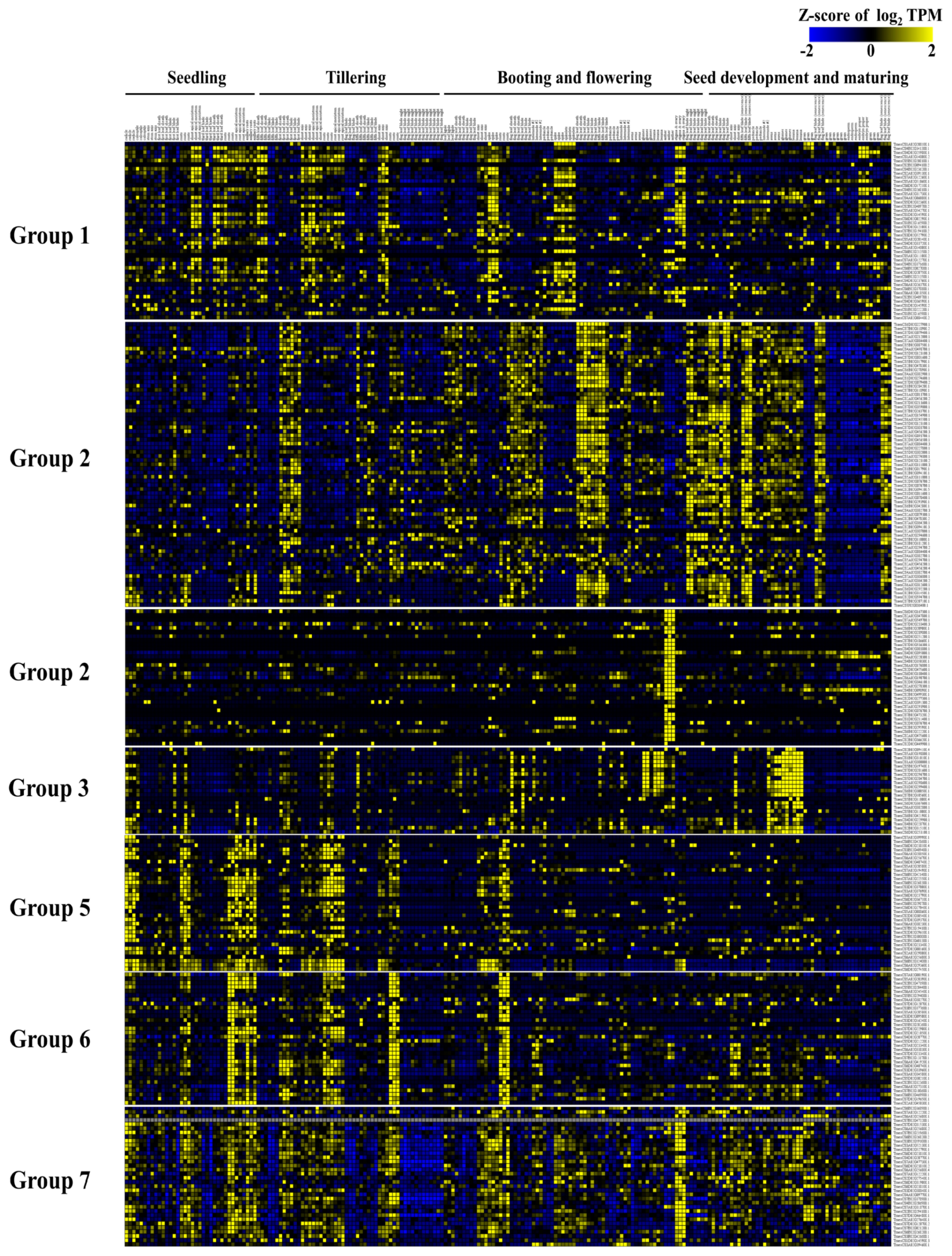
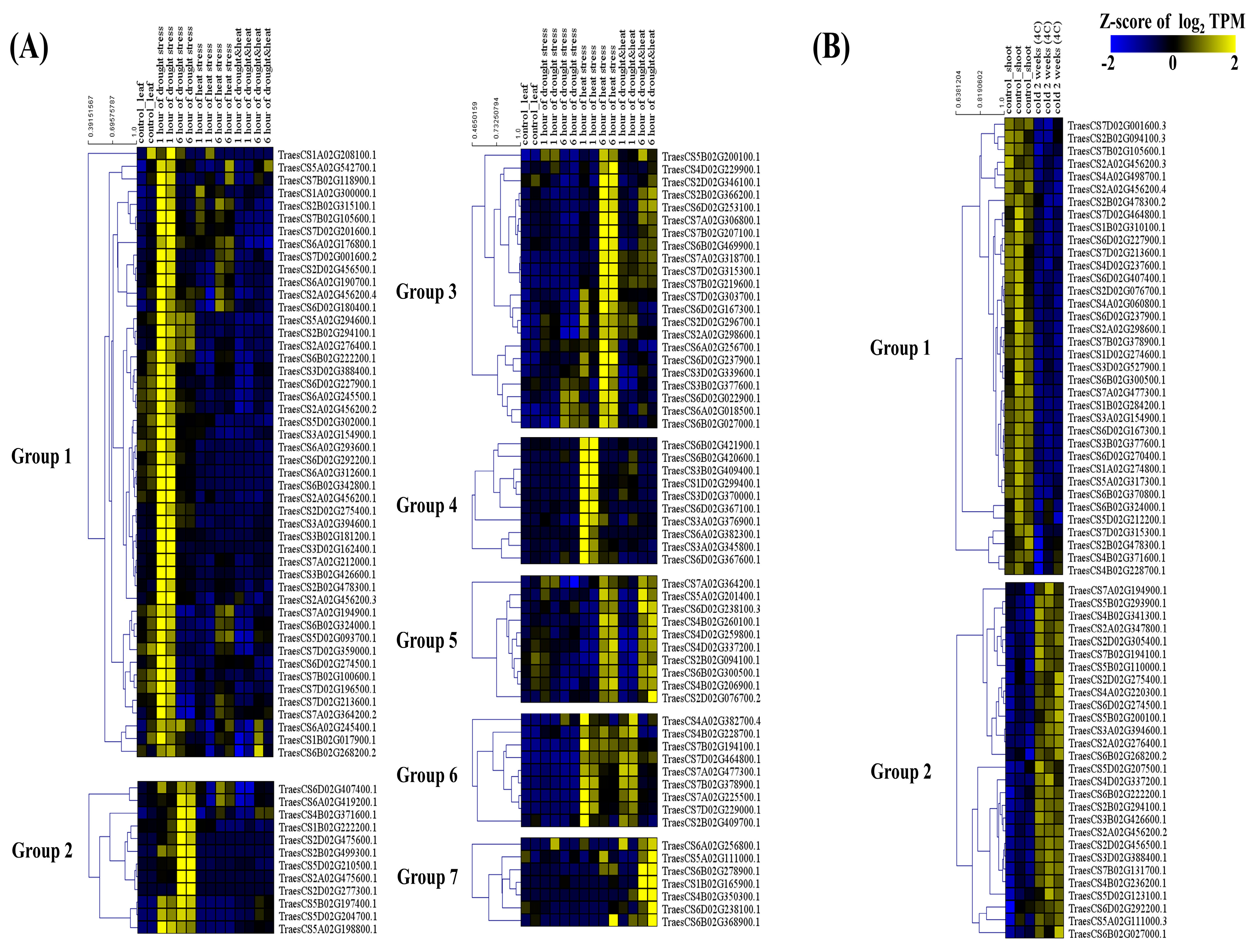
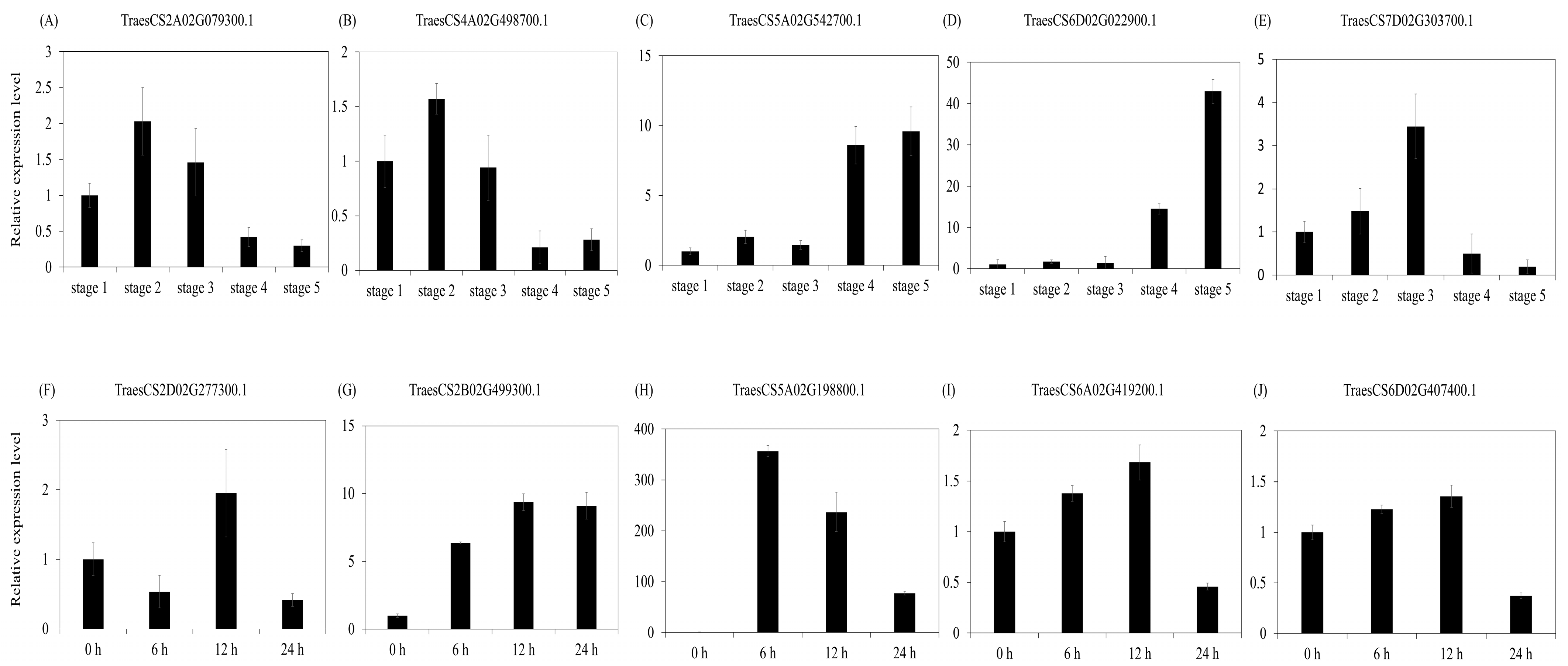
2.6. Expression Analysis of U-Box E3 Genes
3. Discussion
4. Materials and Methods
4.1. Identification and Sequence Analysis of U-Box E3 Genes in the Wheat Genome
4.2. Phylogenetic Analysis and Chromosomal Locations of U-Box E3 Genes
4.3. Genome Distribution with High Sequence Homology and Gene Duplication of U-Box E3 Genes
4.4. Synteny Analysis of U-Box E3 Genes in Wheat and Other Plants
4.5. Expression Analysis of U-Box E3 Genes Using RNA-Seq Data
4.6. Plant Materials and Real-Time Quantitative RT-PCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hershko, A.; Ciechanover, A. The Ubiquitin System. Annu. Rev. Biochem. 1998, 67, 425–479. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L.; Callis, J. Ubiquitin Ligases Mediate Growth and Development by Promoting Protein Death. Curr. Opin. Plant Biol. 2007, 10, 624–632. [Google Scholar] [CrossRef]
- Smalle, J.; Vierstra, R.D. The Ubiquitin 26s Proteasome Proteolytic Pathway. Annu. Rev. Plant Biol. 2004, 55, 555–590. [Google Scholar] [CrossRef] [PubMed]
- Serrano, I.; Campos, L.; Rivas, S. Roles of E3 Ubiquitin-Ligases in Nuclear Protein Homeostasis During Plant Stress Responses. Front. Plant Sci. 2018, 9, 139. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L.; Hauksdóttir, H.; Troy, A.; Herschleb, J.; Kraft, E.; Callis, J. Functional Analysis of the Ring-Type Ubiquitin Ligase Family of Arabidopsis. Plant Physiol. 2005, 137, 13–30. [Google Scholar] [CrossRef]
- Hua, Z.; Vierstra, R.D. The Cullin-Ring Ubiquitin-Protein Ligases. Annu. Rev. Plant Biol. 2011, 62, 299–334. [Google Scholar] [CrossRef]
- Johnson, E.S.; Ma, P.C.M.; Ota, I.M.; Varshavsky, A. A Proteolytic Pathway That Recognizes Ubiquitin as a Degradation Signal. J. Biol. Chem. 1995, 270, 17442–17456. [Google Scholar] [CrossRef] [PubMed]
- Aravind, L.; Koonin, E.V. The U Box Is a Modified Ring Finger—A Common Domain in Ubiquitination. Curr. Biol. 2000, 10, R132–R134. [Google Scholar] [CrossRef]
- Saurin, A.J.; Borden, K.L.B.; Boddy, M.N.; Freemont, P.S. Does This Have a Familiar Ring? Trends Biochem. Sci. 1996, 21, 208–214. [Google Scholar] [CrossRef]
- Wiborg, J.; O’Shea, C.; Skriver, K. Biochemical Function of Typical and Variant Arabidopsis thaliana U-Box E3 Ubiquitin-Protein Ligases. Biochem. J. 2008, 413, 447–457. [Google Scholar] [CrossRef]
- Zeng, L.-R.; Park, C.H.; Venu, R.C.; Gough, J.; Wang, G.L. Classification, Expression Pattern, and E3 Ligase Activity Assay of Rice U-Box-Containing Proteins. Mol. Plant 2008, 1, 800–815. [Google Scholar] [CrossRef]
- Wang, N.; Liu, Y.; Cong, Y.; Wang, T.; Zhong, X.; Yang, S.; Li, Y.; Gai, J. Genome-Wide Identification of Soybean U-Box E3 Ubiquitin Ligases and Roles of Gmpub8 in Negative Regulation of Drought Stress Response in Arabidopsis. Plant Cell Physiol. 2016, 57, 1189–1209. [Google Scholar] [CrossRef]
- Ryu, M.Y.; Cho, S.K.; Hong, Y.; Kim, J.; Kim, J.H.; Kim, G.M.; Chen, Y.J.; Knoch, E.; Møller, B.L.; Kim, W.T.; et al. Classification of Barley U-Box E3 Ligases and Their Expression Patterns in Response to Drought and Pathogen Stresses. BMC Genom. 2019, 20, 326. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-J.; Yi, J.; Yoon, J.; Cho, L.-H.; Ping, J.; Jeong, H.J.; Cho, S.K.; Kim, W.T.; An, G. Ospub15, an E3 Ubiquitin Ligase, Functions to Reduce Cellular Oxidative Stress During Seedling Establishment. Plant J. 2011, 65, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.; Gao, Z.; Xu, X.; Wang, Y.; Lin, Y.; Ye, P.; Huang, T. Plant U-Box E3 Ligases Pub 25 and Pub 26 Control Organ Growth in Arabidopsis. N. Phytol. 2021, 229, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Yoo, Y.H.; Jiang, X.; Jung, K.-H. An Abiotic Stress Responsive U-Box E3 Ubiquitin Ligase Is Involved in Osgi-Mediating Diurnal Rhythm Regulating Mechanism. Plants 2020, 9, 1071. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, M.; Guo, Y. Ring/U-Box Protein Atusr1 Functions in Promoting Leaf Senescence Through ja Signaling Pathway in Arabidopsis. Front. Plant Sci. 2020, 11, 608589. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.R.; Qu, S.; Bordeos, A.; Yang, C.; Baraoidan, M.; Yan, H.; Xie, Q.; Nahm, B.H.; Leung, H.; Wang, G.L. Spotted leaf11, A Negative Regulator of Plant Cell Death and Defense, Encodes a U-Box/Armadillo Repeat Protein Endowed with E3 Ubiquitin Ligase Activity. Plant Cell 2004, 16, 2795–2808. [Google Scholar] [CrossRef] [PubMed]
- Samuel, M.A.; Mudgil, Y.; Salt, J.N.; Delmas, F.; Ramachandran, S.; Chilelli, A.; Goring, D.R. Interactions Between the S-Domain Receptor Kinases and Atpub-Arm E3 Ubiquitin Ligases Suggest a Conserved Signaling Pathway in Arabidopsis. Plant Physiol. 2008, 147, 2084–2095. [Google Scholar] [CrossRef]
- Amador, V.; Monte, E.; García-Martínez, J.-L.; Prat, S. Gibberellins Signal Nuclear Import of Phor1, a Photoperiod-Responsive Protein with Homology to Drosophila armadillo. Cell 2001, 106, 343–354. [Google Scholar] [CrossRef]
- Cho, S.K.; Bae, H.; Ryu, M.Y.; Wook, Y.S.; Kim, W.T. Pub 22 and Pub23 U-Box E3 Ligases Directly Ubiquitinate Rpn6, a 26s Proteasome Lid Subunit, for Subsequent Degradation in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2015, 464, 994–999. [Google Scholar] [CrossRef]
- Cho, S.K.; Chung, H.S.; Ryu, M.Y.; Park, M.J.; Lee, M.M.; Bahk, Y.Y.; Kim, J.; Pai, H.S.; Kim, W.T. Heterologous Expression and Molecular and Cellular Characterization of Capub1 Encoding a Hot Pepper U-Box E3 Ubiquitin Ligase Homolog. Plant Physiol. 2006, 142, 1664–1682. [Google Scholar] [CrossRef]
- International Wheat Genome Sequencing Consortium (IWGSC). Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [PubMed]
- Buetow, L.; Huang, D.T. Structural Insights into the Catalysis and Regulation of E3 Ubiquitin Ligases. Nat. Rev. Mol. Cell Biol. 2016, 17, 626–642. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, A.; Ten Hove, C.A.; Tabata, R.; Yamada, M.; Shimizu, N.; Ishida, T.; Yamaguchi, K.; Shigenobu, S.; Takebayashi, Y.; Iuchi, S.; et al. A Plant U-Box Protein, Pub 4, Regulates Asymmetric Cell Division and Cell Proliferation in the Root Meristem. Development 2015, 142, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.; Zhao, P.; Seo, J.S.; Mitsuda, N.; Deng, S.; Chua, N.-H. Plant U-Box protein 10 Regulates Myc2 Stability in Arabidopsis. Plant Cell 2015, 27, 2016–2031. [Google Scholar] [CrossRef]
- Yu, F.; Wu, Y.; Xie, Q. Ubiquitin–Proteasome System in ABA Signaling: From Perception to Action. Mol. Plant 2016, 9, 21–33. [Google Scholar] [CrossRef]
- Stone, S.L. The Role of Ubiquitin and the 26s Proteasome in Plant Abiotic Stress Signaling. Front. Plant Sci. 2014, 5, 135. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, G.Q.; Kang, H.-H.; Zhou, S.-M.; Wang, W. Tapub1, a Putative E3 Ligase Gene from Wheat, Enhances Salt Stress Tolerance in Transgenic Nicotiana Benthamiana. Plant Cell Physiol. 2017, 58, 1673–1688. [Google Scholar] [CrossRef]
- Andersen, P.; Kragelund, B.B.; Olsen, A.N.; Larsen, F.H.; Chua, N.-H.; Poulsen, F.M.; Skriver, K. Structure and Biochemical Function of a Prototypical Arabidopsis U-Box Domain. J. Biol. Chem. 2004, 279, 40053–40061. [Google Scholar] [CrossRef]
- Trujillo, M. News from the Pub: Plant U-Box type E3 Ubiquitin Ligases. J. Exp. Bot. 2018, 69, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Furlan, G.; Nakagami, H.; Eschen-Lippold, L.; Jiang, X.; Majovsky, P.; Kowarschik, K.; Hoehenwarter, W.; Lee, J.; Trujillo, M. Changes in Pub 22 Ubiquitination Modes Triggered by Mitogen-Activated Protein Kinase3 Dampen the Immune Response. Plant Cell 2017, 29, 726–745. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, C.; Santos-Rosa, M.J.; Shirasu, K. The U-Box Protein Family in Plants. Trends Plant Sci. 2001, 6, 354–358. [Google Scholar] [CrossRef]
- Consortium, International Wheat Genome Sequencing. A Chromosome-Based Draft Sequence of the Hexaploid Bread Wheat (Triticum aestivum) Genome. Science 2014, 345, 1251788. [Google Scholar] [CrossRef]
- Lallemand, T.; Leduc, M.; Landès, C.; Rizzon, C.; Lerat, E. An Overview of Duplicated Gene Detection Methods: Why the Duplication Mechanism Has to Be Accounted for in Their Choice. Genes 2020, 11, 1046. [Google Scholar] [CrossRef] [PubMed]
- Bolot, S.; Abrouk, M.; Masood-Quraishi, U.; Stein, N.; Messing, J.; Feuillet, C.; Salse, J. The “Inner Circle” of the Cereal Genomes. Curr. Opin. Plant Biol. 2009, 12, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Chalupska, D.; Lee, H.Y.; Faris, J.D.; Evrard, A.; Chalhoub, B.; Haselkorn, R.; Gornicki, P. Acc Homoeoloci and the Evolution of Wheat Genomes. Proc. Natl. Acad. Sci. USA 2008, 105, 9691–9696. [Google Scholar] [CrossRef] [PubMed]
- Brkljacic, J.; Grotewold, E.; Scholl, R.; Mockler, T.; Garvin, D.F.; Vain, P.; Brutnell, T.; Sibout, R.; Bevan, M.; Budak, H.; et al. Brachypodium as a Model for the Grasses: Today and the Future. Plant Physiol. 2011, 157, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, N.A. Sequencing the Rice Genome. Plant Cell 2000, 12, 2011–2017. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, A.; Seo, M.; Kamiya, Y.; Sawa, S. Mystery in Genetics: Pub 4 Gives a Clue to the Complex Mechanism of Clv Signaling Pathway in the Shoot Apical Meristem. Plant Signal. Behav. 2015, 10, e1028707. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, Y.; Wu, Y.; Yu, B.; Yin, Z.; Xia, Y. Extra-Large G Proteins Interact with E3 Ligases Pub 4 and Pub 2 and Function in Cytokinin and Developmental Processes. Plant Physiol. 2017, 173, 1235–1246. [Google Scholar] [CrossRef]
- Deb, S.; Sankaranarayanan, S.; Wewala, G.; Widdup, E.; Samuel, M.A. The S-Domain Receptor Kinase Arabidopsis Receptor Kinase2 and the U Box/Armadillo Repeat-Containing E3 Ubiquitin Ligase9 Module Mediates Lateral Root Development Under Phosphate Starvation in Arabidopsis. Plant Physiol. 2014, 165, 1647–1656. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lu, D.; Xu, G.; Finlayson, S.A.; He, P.; Shan, L. The Dominant Negative Arm Domain Uncovers Multiple Functions of Pub 13 in Arabidopsis Immunity, Flowering, and Senescence. J. Exp. Bot. 2015, 66, 3353–3366. [Google Scholar] [CrossRef]
- Vega-Sánchez, M.E.; Zeng, L.; Chen, S.; Leung, H.; Wang, G.L. Spin1. a K Homology Domain Protein Negatively Regulated and Ubiquitinated by the E3 Ubiquitin Ligase Spl11, Is Involved in Flowering Time Control in Rice. Plant Cell 2008, 20, 1456–1469. [Google Scholar] [CrossRef]
- Woodson, J.D.; Joens, M.S.; Sinson, A.B.; Gilkerson, J.; Salomé, P.A.; Weigel, D.; Fitzpatrick, J.A.; Chory, J. Ubiquitin Facilitates a Quality-Control Pathway That Removes Damaged Chloroplasts. Science 2015, 350, 450–454. [Google Scholar] [CrossRef]
- Adler, G.; Konrad, Z.; Zamir, L.; Mishra, A.K.; Raveh, D.; Bar-Zvi, D. The Arabidopsis Paralogs, Pub 46 and Pub 48, Encoding U-Box E3 Ubiquitin Ligases, Are Essential for Plant Response to Drought Stress. BMC Plant Biol. 2017, 17, 8. [Google Scholar] [CrossRef] [PubMed]
- Bergler, J.; Hoth, S. Plant U-Box Armadillo Repeat Proteins Atpub18 and Atpub19 Are Involved in Salt Inhibition of Germination in Arabidopsis. Plant Biol. 2011, 13, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, W.; Wu, Y.; Li, Q.; Zhang, G.; Shi, R.; Yang, J.; Wang, Y.; Wang, W. The Involvement of Wheat U-Box E3 Ubiquitin Ligase Tapub1 in Salt Stress Tolerance. J. Integr. Plant Biol. 2020, 62, 631–651. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The Protein Families Database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent Updates to the Protein Domain Annotation Resource. Nucleic Acids Res. 2012, 40, D302–D305. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S. Blast2GO: A Comprehensive Suite for Functional Analysis in Plant Genomics. Int. J. Plant Genom. 2008, 2008, 619832. [Google Scholar] [CrossRef]
- Moore, A.D.; Held, A.; Terrapon, N.; Weiner, J., 3rd; Bornberg-Bauer, E. Domosaics: Software for Domain Arrangement Visualization and Domain-Centric Analysis of Proteins. Bioinformatics 2014, 30, 282–283. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (Itol): An Online Tool for Phylogenetic Tree Display and Annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef]
- Voorrips, R.E. Mapchart: Software for the Graphical Presentation of Linkage Maps and Qtls. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An Information Aesthetic for Comparative Genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-González, R.H.; Borrill, P.; Lang, D.; Harrington, S.A.; Brinton, J.; Venturini, L.; Davey, M.; Jacobs, J.; Van Ex, F.; Pasha, A.; et al. The Transcriptional Landscape of Hexaploid Wheat. Science 2018, 361, eaar6089. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Xin, M.; Qin, J.; Peng, H.; Ni, Z.; Yao, Y.; Sun, Q. Temporal Transcriptome Profiling Reveals Expression Partitioning of Homeologous Genes Contributing to Heat and Drought Acclimation in Wheat (Triticum aestivum L.). BMC Plant Biol. 2015, 15, 152. [Google Scholar] [CrossRef] [PubMed]
- Howe, E.A.; Sinha, R.; Schlauch, D.; Quackenbush, J. RNA-Seq Analysis in Mev. Bioinformatics 2011, 27, 3209–3210. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal Code for the Growth Stages of Cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.Y.; Lee, Y.J.; Hong, M.J.; Kim, J.H.; Seo, Y.W. Genome Wide Analysis of U-Box E3 Ubiquitin Ligases in Wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2021, 22, 2699. https://doi.org/10.3390/ijms22052699
Kim DY, Lee YJ, Hong MJ, Kim JH, Seo YW. Genome Wide Analysis of U-Box E3 Ubiquitin Ligases in Wheat (Triticum aestivum L.). International Journal of Molecular Sciences. 2021; 22(5):2699. https://doi.org/10.3390/ijms22052699
Chicago/Turabian StyleKim, Dae Yeon, Yong Jin Lee, Min Jeong Hong, Jae Ho Kim, and Yong Weon Seo. 2021. "Genome Wide Analysis of U-Box E3 Ubiquitin Ligases in Wheat (Triticum aestivum L.)" International Journal of Molecular Sciences 22, no. 5: 2699. https://doi.org/10.3390/ijms22052699
APA StyleKim, D. Y., Lee, Y. J., Hong, M. J., Kim, J. H., & Seo, Y. W. (2021). Genome Wide Analysis of U-Box E3 Ubiquitin Ligases in Wheat (Triticum aestivum L.). International Journal of Molecular Sciences, 22(5), 2699. https://doi.org/10.3390/ijms22052699